canale de sociu si calciu
DESCRIPTION
biologie celulara, membrane celulare, canale membranareTRANSCRIPT

6. Modificatori ai gating-ului şi blocanţi
6.1 pH
Efectele variaţiilor de pH asupra canalelor ionice reprezintă o temă îndelung studiată în electrofiziologie. Astfel, o serie de experimentatori au pus în evidenţă o scădere a permeabilităţii maxime pentru Na+ la acidifierea mediului extracelular, curbele de titrare indicând un pKa
aparent între 4.6 şi 5.4 la 0 mV în diverse preparate: noduri Ranvier la amfibieni, muşchi scheletici, axoni giganţi de calmar sau Myxicola (Woodhull 1973, Drouin & Neumcke 1974, Schauf & Davis 1976, Campbell & Hille 1976, Mozhayeva et al. 1981-1982, Campbell 1982, Begenisich & Danko 1983, etc.). Ţinând cont de faptul că reacţiile de protonare-deprotonare în acest interval de valori pH sunt rapide, au fost formulate cel puţin trei ipoteze:- scăderea pH poate afecta cinetica de gating a canalelor de Na+
- scăderea pH poate afecta conductanţa canalelor de Na+ prin titrarea unui număr mare de sarcini negative distribuite difuz la gura porului (teoria potenţialului de suprafaţă)
- scăderea pH poate afecta conductanţa canalelor de Na+ prin titrarea unui reziduu aminoacil esenţial din peretele porului (teoria grupării acide).
Deşi pentru canalele de Na+ voltage-gated prima ipoteză a fost infirmată (în 1980, Sigworth a arătat prin analiza fluctuaţiilor că expunerea nodurilor Ranvier la pH 5.0 scade conductanţa unicanal la 40% din control), recent s-au descoperit diferite canale ionice care funcţionează ca senzori de pH, fiind activate la acidifierea mediului extracelular. Din această categorie fac parte canalele numite ASIC (acid sensing ion channels), care aparţin familiei Deg/ENaC, larg

răspândite în sistemul nervos central, periferic, şi în alte ţesuturi. Un studiu recent de mutageneză (Baron et al. 2001) a identificat în secvenţa ASIC2a un reziduu extracelular (His72) a cărui transformare aboleşte sensibilitatea la pH a canalului. Un alt exemplu este receptorul vaniloid VR1, care integrează diferiţi stimuli nocivi: căldură, aciditate şi compuşi vaniloizi. Jordt et al. (2000) au identificat un reziduu din a treia regiune extracelulară a subunităţii VR1 (E648), pentru care mutaţiile punctiforme cu neutralizarea sarcinii (E648Q sau E648A) reduc sensibilitatea la variaţii de pH, fără a afecta activarea prin căldură sau vaniloizi. Pentru un alt reziduu (E600), mutaţiile cu neutralizarea (E600N) sau inversarea sarcinii (E600K) duc la o activare permanentă a canalului şi reduc efectul de potenţare de către protoni a stimulării prin căldură sau compuşi vaniloizi, în timp ce mutaţiile cu conservarea sarcinii (E600D) nu influenţează semnificativ acest sinergism.

Pentru canalul de K+ din Streptomyces lividans a cărui structură 3D a fost rezolvată (Kcsa), studii prin spectroscopie de rezonanţă electronică paramagnetică ale mutanţilor în care reziduurile TM1 şi TM2 au fost înlocuite pe rând cu cisteină pentru a lega MTS cu marker de spin, au relevat că activarea canalului prin reducerea pH implică o mişcare de ansamblu de rotaţie a celor două -helixuri, cu deschiderea orificiului intern al porului (Perozo, Cortes & Cuello 1999). Prin această rotaţie, atât cuplajul spin-spin între subunităţi (dat de amplitudinea peakului principal de rezonanţă), ce reflectă distanţa între markeri, cât şi lărgimea rezonanţei, ce reflectă mobilitatea markerilor, prezintă variaţii distincte pentru reziduurile de pe TM1 şi TM2.Revenind la studiile mai vechi pe canale de Na+ voltage-gated din axonul gigant de calmar, putem sintetiza constatările lor astfel: există trei efecte distincte ale reducerii pH extracelular asupra dependenţei de voltaj a conductanţei de vârf pentru Na+ (care la pH normal urmează o distribuţie Boltzmann):- o deplasare spre dreapta (right voltage shift) a activării cu 20-30
mV- o reducere a permeabilităţii maxime, chiar la depolarizări mari
Fig. 6.1 Activarea Kcsa la reducerea pH implică o rotaţie de ansamblu a -helixurilor transmembranare. Prin cuplarea cu markeri de spin a reziduurilor mutate în cisteină, se pot evidenţia variaţii de distanţă (exprimată prin cuplajul spin-spin) şi de mobilitate (exprimată prin lărgimea peakului de rezonanţă H0) (din Perozo et al. 1999).

- chiar la potenţiale pozitive mari curba nu mai este plată, ci păstrează o anumită pantă.
AM Woodhull a reuşit să modeleze aceste efecte considerând un situs de protonare aflat la distanţa relativă faţă de exterior în câmpul electric transmembranar. Atunci probabilitatea stării de conductanţă a canalelor, p, este complementară ocupanţei de către protoni a situsului respectiv. Neglijând competiţia cu alţi ioni, şi notând ratele de acces şi de părăsire a situsului din/spre interior şi exterior cu k1 şi k-1, respectiv k2 şi k-2, obţinem la echilibru:
În continuare, exprimând ratele în funcţie de înălţimea barierei de energie liberă (la care se adaugă lucrul mecanic de deplasare a protonilor în câmpul electric transmembranar până în punctul de maxim, considerat la jumătatea distanţei de la exterior la situs - /2), şi presupunând o barieră energetică infinită la accesul dinspre interior,
obţinem:
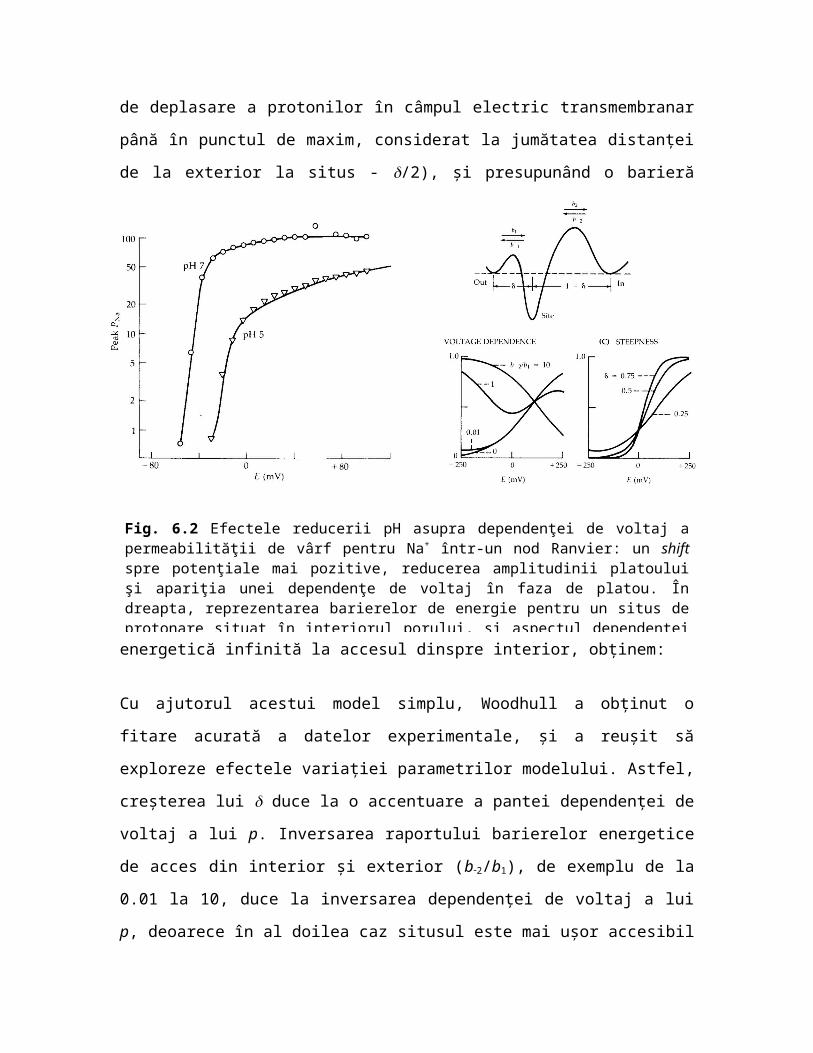
Cu ajutorul acestui model simplu, Woodhull a obţinut o fitare acurată a datelor experimentale, şi a reuşit să exploreze efectele variaţiei parametrilor modelului. Astfel, creşterea lui duce la o accentuare a pantei dependenţei de voltaj a lui p. Inversarea raportului barierelor energetice de acces din interior şi exterior (b2/b1), de exemplu de la 0.01 la 10, duce la inversarea dependenţei de voltaj a lui p, deoarece în al doilea caz situsul este mai uşor accesibil dinspre interior, deci protonarea lui este favorizată de depolarizare.
6.2 Ca2+
Efectele calciului, şi, în general, ale cationilor divalenţi, asupra sistemelor excitabile reprezintă un domeniu de studiu cu o îndelungată istorie. În 1880 Sydney Ringer demonstra că prezenţa calciului este absolut necesară pentru menţinerea bătăilor cordului izolat de broască. Ulterior s-a dovedit că nivelul calciului liber intracitoplasmatic este
Fig. 6.2 Efectele reducerii pH asupra dependenţei de voltaj a permeabilităţii de vârf pentru Na+ într-un nod Ranvier: un shift spre potenţiale mai pozitive, reducerea amplitudinii platoului şi apariţia unei dependenţe de voltaj în faza de platou. În dreapta, reprezentarea barierelor de energie pentru un situs de protonare situat în interiorul porului, şi aspectul dependenţei de voltaj a permeabilităţii pentru diferite rapoarte ale înălţimii barierelor externă şi internă (din Woodhull 1973).

extrem de redus, aproximativ 10-7 M (100 nM), iar creşterea sa prin activarea canalelor de Ca2+ din membrana celulară sau a diferitelor depozite intracelulare reprezintă o cale de semnalizare. Rolul calciului ca mesager secund intracelular nu poate fi înlocuit de alţi ioni similari, precum Mg2+ sau Ba2+. Ce se poate spune însă despre calciul extracelular? Experimentele au arătat că efectul său este stabilizator, de reducere a excitabilităţii neuromusculare, iar aceste acţiuni sunt comune cationilor divalenţi. După punerea la punct a metodei voltage clamp pe axonul gigant de calmar, Frankenhaeuser & Hodgkin (1957) au studiat efectele reducerii concentraţiei Ca2+ extracelular asupra dependenţei de voltaj a cineticii de activare şi inactivare a conductanţelor de Na+ şi K+. Ei au considerat două explicaţii posibile ale rezultatelor lor:
- modificarea potenţialului de suprafaţă al membranei prin adsorbţia Ca2+ şi neutralizarea reziduurilor negative, echivalentă cu o
depolarizare (ipoteză avansată de A.F. Huxley)- influenţarea sarcinii echivalente de gating prin interacţiunea cu
Ca2+. Această teorie poate fi respinsă în forma ei extremă, şi anume cea care pretinde că ionii de Ca2+ reprezintă ei înşişi particulele de gating. Deşi contraargumentul iniţial se baza pe considerente energetice (distanţa electrică echivalentă a blocului), ulterior au fost aduse noi infirmări (discutate de Hille): identificarea sarcinilor de gating ca aparţinând unor reziduuri de Arg şi Lys din S4, persistenţa efectelor cinetice la canale permeabile pentru Ca2+, cinetica rapidă a blocării de către Ca2+
a canalelor de Na2+ (microsecunde) comparativ cu cinetica lor de
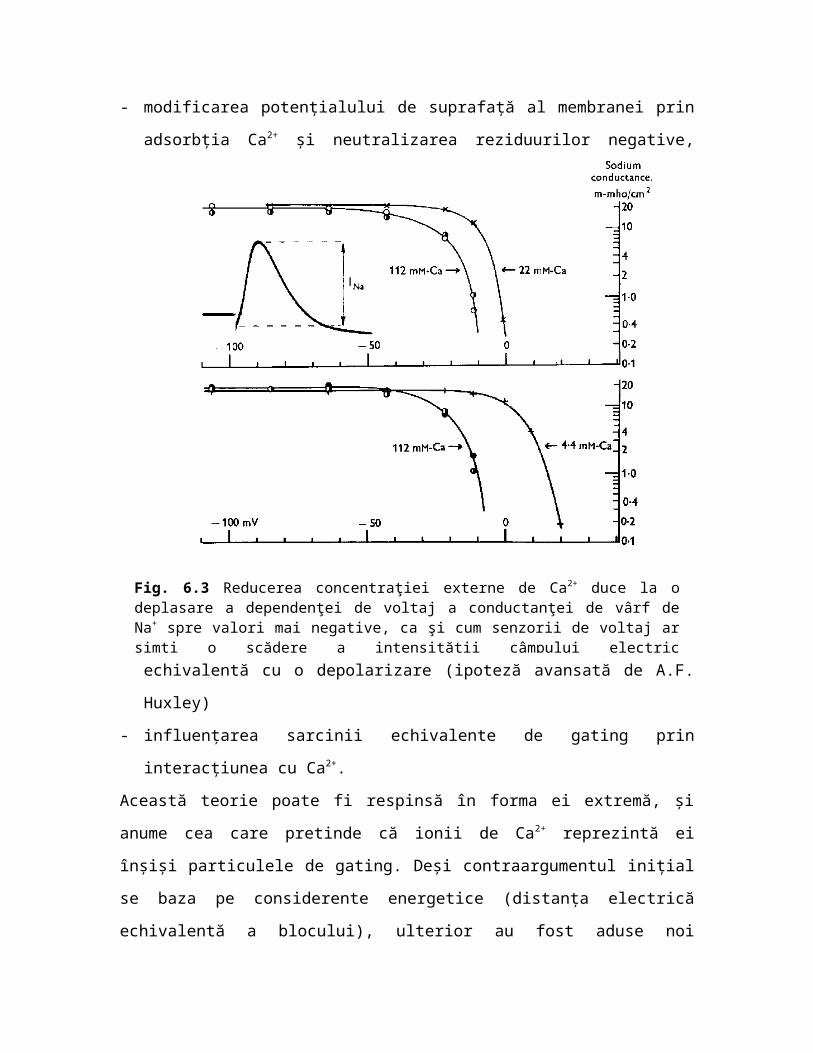
Fig. 6.3 Reducerea concentraţiei externe de Ca2+ duce la o deplasare a dependenţei de voltaj a conductanţei de vârf de Na+ spre valori mai negative, ca şi cum senzorii de voltaj ar simţi o scădere a intensităţii câmpului electric transmembranar (din Frankenhaeuser & Hodgkin 1957).

gating (milisecunde), şi în fine, mutaţiile care cresc sau scad afinitatea pentru Ca2+ a filtrului de selectivitate cu mai multe ordine de mărime (Heinemann et al 1992) nu influenţează dependenţa de voltaj a cineticii canalelor. Rămâne însă valabilă cealaltă formulare, anume posibilitatea existenţei unei interacţiuni între Ca2+ şi sarcinile senzorului de voltaj.Înainte de a examina această direcţie, să recapitulăm pe scurt prima teorie, cea a potenţialului de suprafaţă. În forma sa cea mai simplă, ea pretinde existenţa unei distribuţii uniforme de sarcină pe suprafaţa membranei, şi defineşte o densitate de sarcină de suprafaţă, . Aceasta poate fi calculată pornind de la concentraţiile relative ale speciilor ionice cok la suprafaţa membranei, deduse la rândul lor dintr-o distribuţie Boltzmann, în funcţie de valoarea potenţialului de suprafaţă 0 şi de concentraţiile în soluţie (bulk), ck:
În formularea iniţială, aparţinând fizico-chimiştilor americani Gouy (1911) şi Chapman (1913), relaţia între densitatea de sarcină de suprafaţă şi concentraţia ionică c într-un electrolit binar simetric este:
Grahame (1947) generalizează această ecuaţie:
Deşi o astfel de reprezentare funcţionează corect în descrierea efectelor de screening (ecranarea interacţiunilor electrostatice între grupările polare ale fosfolipidelor de membrană), care nu depind de natura ionului, ci doar de sarcina sa, efectele de legare (binding) trebuie să joace un rol important, deoarece influenţa calciului asupra

potenţialului de suprafaţă este mult mai pronunţată decât în cazul ionilor monovalenţi. Legarea poate fi cuantificată printr-o dezvoltare a ecuaţiei Grahame în modelul Gouy-Chapman-Stern, ce presupune o distribuţie neuniformă a situsurilor de legare pe suprafaţa membranei, cu o densitate S. Putem exprima independent densitatea de sarcină de suprafaţă în funcţie de concentraţia ionică printr-o izotermă de adsorbţie Langmuir:Pentru bistraturi fosfolipidice, S are o valoare maximă de aproximativ 1/80 Å2, corespunzător ariei transversale medii per moleculă de fosfolipid. Astfel, considerând interacţiunea cu un singur tip de ion, potenţialul de suprafaţă devine:
În cazul unei competiţii între mai multe specii de ioni, constantele de disociere aparente se modifică:
unde KDI reprezintă constanta de disociere a cationului competitor.

Aşa cum au recunoscut Frankenhaeuser şi Hodgkin, teoria potenţialului de suprafaţă explică doar parţial rezultatele experimentale. Astfel, dependenţa de voltaj a conductanţei maxime de Na+ şi de K+ sunt ambele decalate cu aproximativ 15 mV prin scăderea de 5 ori a activităţii Ca2+ extracelular (de la 112 mM la 22 mM şi respectiv la 4.4 mM), ceea ce era de aşteptat, deoarece variaţia intensităţii câmpului electric transmembranar produsă prin modificarea potenţialului de suprafaţă ar trebui să afecteze în aceeaşi măsură senzorii de voltaj ai canalelor de Na+ şi de K+. Acelaşi decalaj (voltage shift) se înregistrează şi pentru potenţialul prag de activare, un parametru mai uşor de măsurat experimental. Hagiwara & Takahashi (1967) au folosit acest parametru pentru a studia efectul diferiţilor cationi divalenţi şi trivalenţi asupra membranei fibrei musculare de barnaclu (mic crustaceu marin din subclasa Cirripede), găsind următoarea secvenţă a
afinităţilor de legare: La3+, UO22+>Zn2+, Co2+, Fe2+>Mn2+>Ni2+>Ca2+>Mg2+, Sr2+. În schimb, efectele asupra inactivării sunt mai complexe, reducerea Ca2+ producând, în afară de decalaj, şi o scădere aparentă a sarcinii echivalente de gating a inactivării, şi o mărire considerabilă a constantelor de timp de inactivare.

Inadecvarea teoriei potenţialului de suprafaţă în a explica rezultatele experimentelor cu cationi di- şi trivalenţi a fost relevată în special de Clay M. Armstrong, într-o serie de studii. Astfel, în 1982 Gilly & Armstrong arată că Zn2+, Cd2+ şi Hg2+ încetinesc constanta de timp de activare a curenţilor ionici de Na+ şi K+, precum şi constanta de timp a curenţilor de gating de activare, în schimb au o influenţă redusă asupra constantei de timp de închidere a curenţilor ionici şi a celor de gating. Ulterior, Armstrong & Cota (1990, 1999) arată că La3+
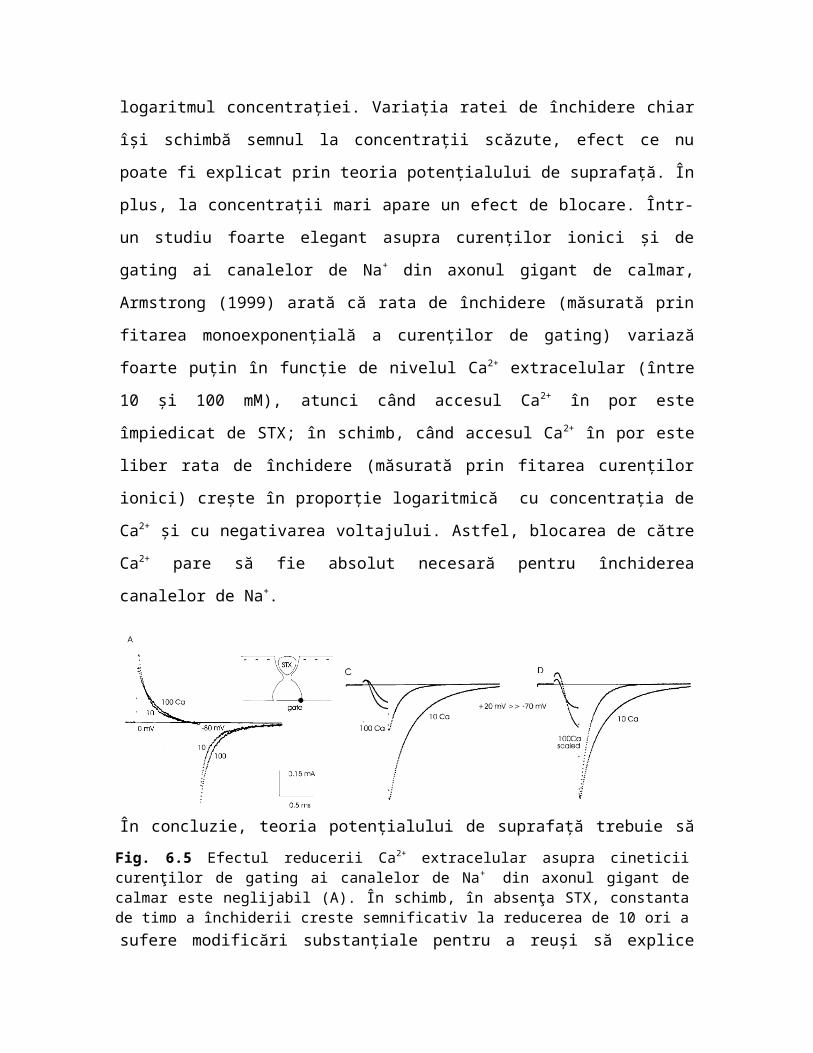
decalează dependenţa de voltaj a ratelor de deschidere şi închidere ale canalelor de Na+ din celule pituitare de mamifere (linia GH3) proporţional cu logaritmul concentraţiei. Variaţia ratei de închidere chiar îşi schimbă semnul la concentraţii scăzute, efect ce nu poate fi explicat prin teoria potenţialului de suprafaţă. În plus, la concentraţii mari apare un efect de blocare. Într-un studiu foarte elegant asupra curenţilor ionici şi de gating ai canalelor de Na+ din axonul gigant de calmar, Armstrong (1999) arată că rata de închidere (măsurată prin fitarea monoexponenţială a curenţilor de gating) variază foarte puţin în
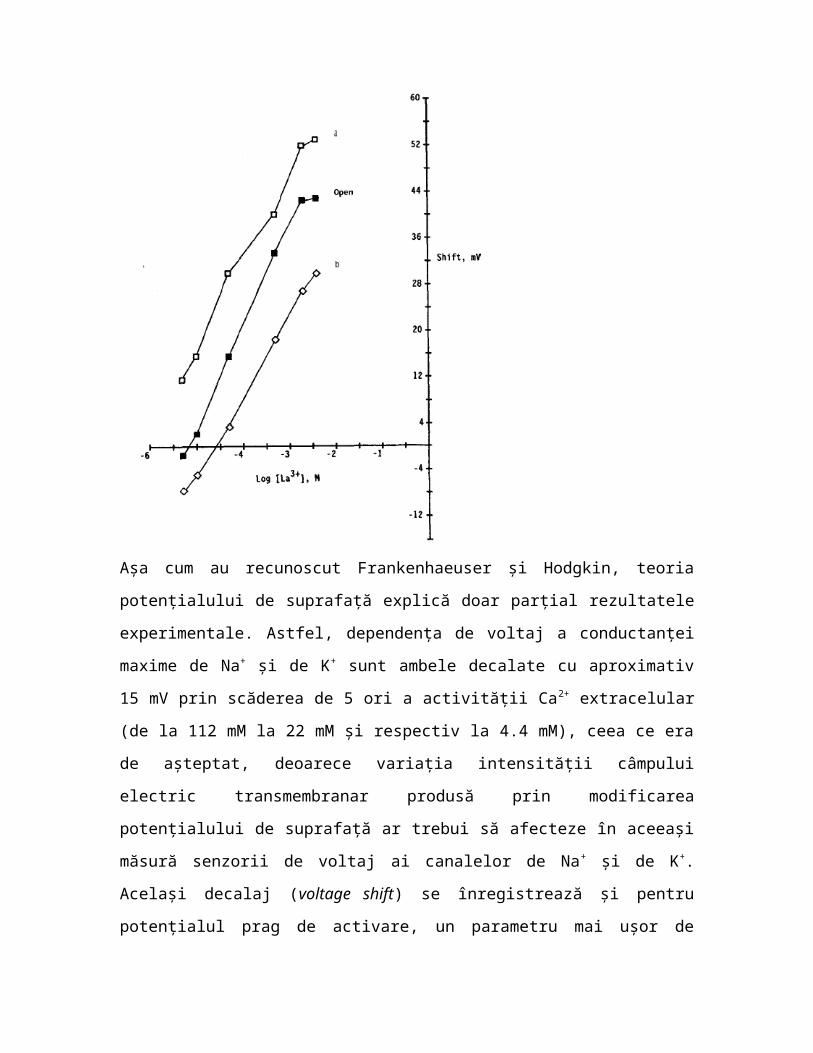
Fig. 6.4 Efectele La3+
asupra dependenţei de voltaj a ratei de deschidere (a), ratei de închidere (b) şi fracţiunii de canale de Na+
deschise, pe o linie de celule pituitare de şobolan (GH3). Efectele nu pot fi explicate integral prin teoria potenţialului de suprafaţă (din Armstrong & Cota 1990).
funcţie de nivelul Ca2+ extracelular (între 10 şi 100 mM), atunci când accesul Ca2+ în por este împiedicat de STX; în schimb, când accesul Ca2+ în por este liber rata de închidere (măsurată prin fitarea curenţilor ionici) creşte în proporţie logaritmică cu concentraţia de Ca2+ şi cu negativarea voltajului. Astfel, blocarea de către Ca2+ pare să fie absolut necesară pentru închiderea canalelor de Na+.În concluzie, teoria potenţialului de suprafaţă trebuie să sufere modificări substanţiale pentru a reuşi să explice particularităţile relevate experimental. Evident, viitorul aparţine metodelor de calculare a potenţialului local mai degrabă decât a potenţialului de suprafaţă, de exemplu cu ajutorul ecuaţiei Poisson-Boltzmann linearizate, în momentul în care structura canalelor ionice dependente de voltaj va fi cunoscută la nivel de rezoluţie atomică.
6.3 Anestezicele locale – state-dependent binding
La fel ca în cazul majorităţii compuşilor farmacologici activi, anestezicele locale au fost utilizate în practica medicală cu mult înainte de a li se cunoaşte mecanismul de acţiune. Sigmund Freud, discipol al
lui Ernst von Brücke la Universitatea din Viena, şi ulterior al psihiatrului
Theodore Meynert şi al profesorului de medicină internă Hermann
Fig. 6.5 Efectul reducerii Ca2+ extracelular asupra cineticii curenţilor de gating ai canalelor de Na+ din axonul gigant de calmar este neglijabil (A). În schimb, în absenţa STX, constanta de timp a închiderii creşte semnificativ la reducerea de 10 ori a concentraţiei de Ca2+ (C, D) (din Armstrong 1999).

Nothnagel, studiază în anii 1880 acţiunea farmacologică a cocainei. Cu toate succesele obţinute de prietenul său Carl Koller în chirurgia oftalmologică, administrarea generală s-a dovedit dezastruoasă datorită dependenţei faţă de drog, ducând la moartea unui alt prieten apropiat, Ernst Fleischl von Marxow. Aceste insuccese au zdruncinat încrederea lui Freud în medicina “somatică” şi au afectat reputaţia sa ca practician, ceea ce l-a determinat să aleagă calea psihanalizei. Ulterior, problema dependenţei avea să fie rezolvată prin sinteza unor compuşi înrudiţi, precum lidocaina (xilina), procaina, benzocaina, tetracaina, etc. Anestezicele locale sunt folosite fie pentru blocarea trunchiurilor sau terminaţiilor nervoase din piele sau mucoase, fie ca antiaritmice ventriculare. Datorită liposolubilităţii, ele traversează direct membranele celulare pentru a ajunge la locul de acţiune. La pH acid (caracteristic ţesuturilor inflamate) ele trec în forma protonată, pentru care membrana este impermeabilă. La începutul anilor 1970, sub impulsul descoperirilor lui Armstrong despre acţiunea aminelor cuaternare asupra canalelor de K+ şi a modelului de open channel block, s-a dovedit că analogii sintetici cu sarcină permanentă ai anestezicelor locale, de exemplu QX-314, acţionează într-un mod similar asupra canalelor de Na+ axonale atunci când sunt aplicaţi intracelular (Frazier et al. 1970, Strichartz 1973). În prezenţa anestezicului, stimulii de depolarizare aplicaţi succesiv elicită curenţi

din ce în ce mai mici, fenomen denumit use-dependent block sau phasic block. Strichartz a explicat acest fenomen printr-un model de state-dependent binding, şi anume open channel block, în care moleculele blocantului nu pot ajunge la situsul de legare decât dinspre interior şi în starea deschisă a canalului. Întrucât există şi o dependenţă de voltaj a blocului (distanţa electrică echivalentă dinspre interior fiind = 0.6), înseamnă că porţile canalului se află la interior, şi dincolo de ele există un vestibul cu pereţi hidrofobi suficient de larg pentru a acomoda anestezicul. Ulterior s-a constatat că use-dependent block produs de anestezicele liposolubile este tranzitoriu, deci ele pot părăsi situsul direct prin pereţii laterali ai porului (aşa-numita cale hidrofobă). Interesant, situsul de legare este accesibil pentru protonii din exterior, nu şi pentru cei din interior, deoarece scăderea pH-ului extracelular duce la protonarea moleculei de anestezic, ceea ce face ca ea să nu mai poată părăsi situsul (Cahalan & Almers 1979, Shapiro 1977). În plus, ionii de Na+ din exterior pot îndepărta blocul, fracţiunea de canale blocate crescând la scăderea [Na+] extracelular sau la aplicarea TTX.Interferenţele anestezicului local cu mecanismele de gating ale canalului sunt multiple: afinitatea situsului de legare este dependentă de stare, iar legarea anestezicului modifică ratele cinetice ale canalului. Explicaţia ar fi accea că anestezicul se leagă mai puternic atunci când poarta de inactivare este închisă, deci hiperpolarizarea cu deschiderea porţii poate duce la îndepărtarea sa. Existenţa mai multor stări cu afinităţi şi rate de legare diferite poartă numele de modelul receptorului modulat (Hille 1977), similar cu variaţia dependentă de
Fig. 6.6 Situsul de legare al anestezicelor locale, situat în vestibulul intern al canalelor de Na+ voltaj-dependente. Compuşii cu sarcină permanentă (de exemplu QX-314) pătrund numai dinspre interior, în timp ce compuşii hidrofobi trec şi prin pereţii laterali. Situsul este accesibil pentru protoni din exterior.

conformaţie a afinităţilor de legare ale enzimelor allosterice (Monod et al 1965).Anestezicele locale acţionează şi asupra receptorilor nicotinici de ACh, tot după un model de state-dependent binding, blocându-i în stare deschisă şi prevenind închiderea lor. Acesta a fost primul efect blocant evidenţiat direct prin metoda patch clamp (Neher & Steinbach 1978). În acest caz, anestezicul local acţionează numai dinspre exterior, pătrunzând la o distanţă electrică = 0.5 – 0.8. Acesta pare a fi modul de acţiune al leptocurarelor, precum D-tubocurarina. De aceea, putem presupune că poarta receptorilor nicotinici se află la exterior, iar filtrul de selectivitate la interior. În segmentul transmembranar M2 au fost identificate un număr de reziduuri care participă la situsul de legare al anestezicelor locale.
6.4 Blocanţi direcţi ai porului – dependenţa de voltaj a blocului
O problemă la fel de importantă în plan teoretic este efectul blocant al unor ioni. De exemplu, canalele de Ca2+ sunt blocate de Mn2+, Co2+ sau Ni2+ aplicaţi extracelular, iar canalele de K+ de TEA+, Cs+, Na+ şi Ba2+
intracelular.O înregistrare patch clamp de mare acurateţe a blocării canalelor de K+
de tip inward rectifier, care prezintă deschideri lungi – peste 1 s – la potenţiale negative, de către Cs+ extern, arată că frecvenţa blocărilor creşte cu gradul de hiperpolarizare, dând un aspect de flickering opening (Fukushima 1982). Diferiţi ioni produc durate medii diferite ale blocului, fapt verificat în canalele de K+ de tip delayed rectifier cu Na+
sau Cs+ intracelular (Hille 1975, French & Wells 1977). Blocul prezintă şi în acest caz o dependenţă de voltaj, fiind intensificat la depolarizare, însă la depolarizări mari scade din nou, ca şi cum ionii blocanţi ar acumula suficientă energie pentru a penetra regiunea îngustă a porului.

Pentru unele canale, încercarea de a aplica modelul Woodhull pentru a exprima dependenţa de voltaj rezultă în distanţe electrice echivalente mai mari decât 1, iar curba doză-răspuns are o pantă mai mare decât cea prezisă în cazul acţiunii unui singur ion. Aceste date sugerează existenţa unor situsuri de blocare multiple, corespunzând situsurilor multiple de legare a ionilor permeanţi de-a lungul porului în canalele multi-ion, prezise de Hodgkin & Keynes (1955).Experimentele lui Neyton & Miller (1988) au relevat un caz interesant în blocarea canalelor de K+ de mare conductanţă activate de Ca2+ – BK K(Ca) – de către Ba2+ intern. Lucrând la diferite combinaţii ale concentraţiilor K+ extra- şi intracelular, ei au relevat existenţa a 4 situsuri de legare a K+ care facilitează blocarea sau împiedică/facilitează deblocarea, la distanţe electrice de 0.7, 0.6 (situsul blocant), 0.5 şi 0.15 faţă de exterior. După descifrarea structurii 3D a Kcsa, aceste situsuri au fost identificate: două situsuri în
Fig. 6.7 Ionii de Cs+ extracelulari produc un bloc dependent de voltaj al canalelor de K+ de tip inward rectifier, dând un aspect de flickering curenţilor single-channel (din Fukushima 1982).

interiorul filtrului de selectivitate, situsul blocant pentru Ba+ la intrarea internă a filtrului, şi încă un situs pentru K+ în vestibulul intern.
Referinţe
Armstrong CM. 1999. Distinguishing surface effects of calcium ion from pore-occupancy effects in Na+ channels. Proc Natl Acad Sci U S A 96(7):4158-63.
Armstrong CM, Cota G. 1990. Modification of sodium channel gating by lanthanum. Some effects that cannot be explained by surface charge theory. J Gen Physiol 96(6):1129-40.
Baron A, Schaefer L, Lingueglia E, Champigny G, Lazdunski M. 2001. Zn2+ and H+ are coactivators of acid-sensing ion channels. J Biol Chem 276(38):35361-7.
Ermakov YA, Averbakh AZ, Yusipovich AI, Sukharev S. 2001. Dipole potentials indicate restructuring of the membrane interface induced by gadolinium and beryllium ions. Biophysical Journal 80(4):1851-62.
Frankenhaeuser B, Hodgkin AL. 1957. The action of calcium on the electrical properties of squid axons. The Journal of Physiology 137:218-244.
Fig. 6.8 Experimente asupra blocării de către Ba2+ a canalelor de K+ activate de Ca2+ - BK(Ca) încorporate în bistraturi artificiale. Diferenţa de potenţial de +40 mV favorizează părăsirea de către Ba2+ a situsului blocant spre exterior, însă creşterea concentraţiei K+ la exterior duce la ocuparea altor situsuri de-a lungul căii de permeaţie, ducând la imobilizarea prelungită a Ba2+
şi eliminarea sa spre interior. În aceste condiţii, creşterea concentraţiei de K+ la interior prelungeşte blocul, relevând un nou situs interior (din Neyton & Miller 1988).

Fukushima Y. 1982. Blocking kinetics of the anomalous potassium rectifier of tunicate egg studied by single channel recording. J Physiol 331:311-31.
Grahame DC. 1947. The electrical double layer and the theory of electrocapillarity. Chem Rev 41:441-501.
Hagiwara S, Takahashi K. 1967. Surface density of calcium ions and calcium spikes in the barnacle muscle fiber membrane. Journal of General Physiology 50(3):583-601.
Heinemann SH, Terlau H, Stuhmer W, Imoto K, Numa S. 1992. Calcium channel characteristics conferred on the sodium channel by single mutations. Nature 356(6368):441-3.
Hille B. 1977. Local anesthetics: hydrophilic and hydrophobic pathways for the drug-receptor reaction. J Gen Physiol 69(4):497-515.
Hille B. 2001. Ionic channels of excitable membranes. Sunderland, MA: Sinauer Associates. 814 p. ill p.
Hodgkin AL, Keynes RD. 1955. The potassium permeability of a giant nerve fiber. J Physiol 128:61-88.
Jordt SE, Tominaga M, Julius D. 2000. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc Natl Acad Sci U S A 97(14):8134-9.
Monod J, Wyman J, Changeux JP. 1965. On the nature of allosteric transitions: a plausible model. J Mol Biol 12:88-118.
Neher E, Steinbach JH. 1978. Local anaesthetics transiently block currents through single acetylcholine-receptor channels. J Physiol 277:153-76.
Neyton J, Miller C. 1988. Discrete Ba2+ block as a probe of ion occupancy and pore structure in the high-conductance Ca2+ -activated K+ channel. J Gen Physiol 92(5):569-86.
Perozo E, Cortes DM, Cuello LG. 1999. Structural rearrangements underlying K+-channel activation gating. Science 285(5424):73-8.
Ringer S. 1883. A further contribution regarding the influence of the different constituents of the blood on the contraction of the heart. J Physiol 4:29-42.
Sigworth FJ. 1980. The conductance of sodium channels under conditions of reduced current at the node of Ranvier. J Physiol 307:131-42.
Woodhull AM. 1973. Ionic blockage of sodium channels in nerve. J Gen Physiol 61(6):687-708.