microbiologie 2011
TRANSCRIPT

Prof.dr. Eugen ULEA Asist.dr. Florin Daniel LIPŞA
Universitatea de Ştiinţe Agricole şi Medicină Veterinară “Ion Ionescu de la Brad” Iaşi
MICROBIOLOGIE
Editura "Ion Ionescu de la Brad"
IAŞI - 2011

Referenţi ştiinţifici:
Prof.univ.dr. Mihai CARP-CĂRARE Universitatea de Ştiinţe Agricloe şi Medicină Veterinară „Ion Ionescu de la Brad” Iaşi Facultatea de Medicină Veterinară
Prof.univ.dr. Cătălin TĂNASE Universitatea „Al.I.Cuza” Iaşi Facultatea de Biologie
ISBN 978-973-147-091-7
Descrierea CIP a Bibliotecii Naţionale a României ULEA, EUGEN
Microbiologie / Eugen Ulea, Florin Daniel Lipşa. – Iaşi : Editura Ion Ionescu de la Brad, 2011 203 pg., 84 il., Bibliogr. ISBN 978-973-147-091-7
I. Lipşa, Florin Daniel 579

În memoria mentorului nostru, Prof.dr.doc. Mircea HATMAN


CUPRINS
CAPITOLUL 1 - OBIECTUL DE STUDIU AL MICROBIOLOGIEI......... 9 1.1. Date selective privind istoricul Microbiologiei ........................... 11 1.2. Şcoala românească de Microbiologie .......................................... 19
CAPITOLUL 2 - CARACTERELE GENERALE ŞI POZIŢIA MICROORGANISMELOR ÎN LUMEA VIE............................................. 21
2.1. Poziţia microorganismelor în lumea vie...................................... 23 2.2. Prionii .......................................................................................... 25 2.3. Virusurile ..................................................................................... 27
2.3.1. Morfologia virusurilor ......................................................... 27 2.3.2. Structura virusurilor............................................................. 28 2.3.3. Virusurile plantelor .............................................................. 29 2.3.4. Bacteriofagii......................................................................... 31 2.3.5. Fagii filamentoşi .................................................................. 37 2.3.6. Cianofagii............................................................................. 38 2.3.7. Micovirusurile...................................................................... 38 2.3.8. Sistematica, nomenclatura şi identificarea virusurilor......... 38
2.4. Bacteriile...................................................................................... 40 2.4.1. Morfologia bacteriilor.......................................................... 41 2.4.2. Structura celulei bacteriene.................................................. 47 2.4.3. Compoziţia chimică a bacteriilor ......................................... 57 2.4.4. Creşterea şi multiplicarea bacteriilor ................................... 59 2.4.5. Nutriţia bacteriilor................................................................ 62 2.4.6. Respiraţia microorganismelor.............................................. 62 2.4.7. Grupe particulare de bacterii................................................ 63 2.4.8. Sistematica, nomenclatura şi identificarea bacteriilor ......... 67 2.4.9. Rolul bacteriilor în natură .................................................... 69
2.5. Cyanobacteriile (algele albastre-verzi)........................................ 69 2.5.1. Morfologia cyanobacteriilor ................................................ 70 2.5.2. Structura cyanobacteriilor.................................................... 70 2.5.3. Sistematica cyanobacteriilor ................................................ 71 2.5.4. Rolul cyanobacteriilor în natură .......................................... 72
2.6. Protozoarele ................................................................................. 72 2.6.1. Morfologia şi fiziologia protozoarelor................................. 72

2.6.2. Structura protozoarelor ........................................................ 73 2.6.3. Reproducerea protozoarelor................................................. 74 2.6.4. Nutriţia protozoarelor .......................................................... 74 2.6.5. Sistematica protozoarelor .................................................... 75 2.6.6. Rolul protozoarelor în natură ............................................... 76
2.7. Diatomeele................................................................................... 76 2.7.1. Morfologia diatomeelor ....................................................... 76 2.7.2. Structura diatomeelor........................................................... 77 2.7.3. Reproducerea diatomeelor ................................................... 78 2.7.4. Sistematica diatomeelor ....................................................... 81 2.7.5. Rolul diatomeelor în natură ................................................. 83
2.8. Ciupercile .................................................................................... 83 2.8.1. Morfologia ciupercilor......................................................... 83 2.8.2. Structura celulară a ciupercilor ............................................ 85 2.8.3. Organele de fixare şi de absorbţie ale ciupercilor................ 87 2.8.4. Formele de rezistenţă ale ciupercilor ................................... 91 2.8.5. Înmulţirea ciupercilor .......................................................... 92 2.8.6. Alternanţa de faze la ciuperci .............................................. 99 2.8.7. Nutriţia ciupercilor............................................................. 100 2.8.8. Originea şi evoluţia parazitismului la ciuperci .................. 102 2.8.9. Cerinţe faţă de mediu......................................................... 103 2.8.10. Sistematica ciupercilor..................................................... 103
CAPITOLUL 3 - INFLUENŢA FACTORILOR ECOLOGICI ASUPRA MICROORGANISMELOR....................................................................... 104
3.1. Influenţa pH-ului ....................................................................... 104 3.2. Influenţa temperaturii ................................................................ 105 3.3. Influenţa apei ............................................................................. 106 3.4. Influenţa energiei radiante ......................................................... 108 3.5. Activitatea microbiană a diferitelor soluri................................. 109 3.6. Profilul microbian al solului ...................................................... 110
CAPITOLUL 4 - INTERRELAŢIILE ECOLOGICE ÎNTRE ORGANISME ............................................................................................................... 111
4.1. Interrelaţiile dintre populaţiile de microorganisme ................... 111 4.2. Interrelaţiile dintre plantele superioare şi microorganismele

din sol ........................................................................................ 115 4.2.1. Influenţa sistemului radicular al plantelor asupra
microflorei solului .............................................................. 115 4.2.2. Natura secreţiilor radicelare şi influenţa lor
asupra structuriilor taxonomice a microflorei solului ........ 117 4.2.3. Vârsta plantelor şi efectul de rizosferă .............................. 118 4.2.4. Micorizele .......................................................................... 118 4.2.5. Interrelaţiile între ciuperci şi plante în cazul micorizelor .. 120
CAPITOLUL 5 - SOLUL CA MEDIU DE EXISTENŢĂ PENTRU MICROORGANISME............................................................................... 122
5.1. Alcătuirea generală a solului ..................................................... 122 5.2. Reacţia solului ........................................................................... 125 5.3. Formarea solului ........................................................................ 125
5.3.1. Compoziţia chimică a solului ............................................ 126 5.3.2. Rolul microorganismelor în formarea şi evoluţia
materiei organice ................................................................ 127 5.3.3. Teoria microbiologică a formării humusului ..................... 128
5.4. Populaţia solului ........................................................................ 130 CAPITOLUL 6 - ROLUL MICROORGANISMELOR ÎN CIRCUITUL MATERIEI ÎN NATURĂ.......................................................................... 132
6.1. Circuitul azotului ....................................................................... 132 6.1.1. Fixarea azotului molecular................................................. 135 6.1.2. Amonificarea ..................................................................... 145 6.1.3. Nitrificarea ......................................................................... 147 6.1.4. Denitrificarea ..................................................................... 149
6.2. Circuitul carbonului................................................................... 150 6.2.1. Metabolismul carbonului mineral ...................................... 152 6.2.2. Metabolismul carbonului organic ...................................... 152
6.3. Circuitul sulfului........................................................................ 162 6.4. Circuitul fierului ........................................................................ 165 6.5. Circuitul fosforului .................................................................... 167 6.6. Circuitul potasiului .................................................................... 169 6.7. Transformările microbiene ale calciului.................................... 169 6.8. Transformările microbiene ale magneziului.............................. 169

6.9. Transformările microbiene ale microelementelor ..................... 170 6.9.1. Manganul ........................................................................... 170 6.9.2. Seleniul .............................................................................. 170 6.9.3. Zincul ................................................................................. 171 6.9.4. Cuprul ................................................................................ 171
CAPITOLUL 7 - MICROBIOLOGIA FERMENTAŢIILOR................... 172 7.1. Fermentaţia alcoolică................................................................. 173
7.1.1. Utilizarea microorganismelor în vinificaţie ....................... 175 7.1.2. Utilizarea microorganismelor în industria berii................. 190
7.2. Fermentaţia lactică..................................................................... 190 7.2.1. Rolul microrganismelor în obţinerea produselor
lactate.................................................................................. 192 7.2.2. Rolul microorganismelor lactice în conservarea
alimentelor murate............................................................. 193 7.2.3. Rolul microorganismelor în producerea furajelor
însilozate............................................................................. 194 7.3. Fermentaţia acetică .................................................................... 195
BIBLIOGRAFIE........................................................................................ 199

9
Capitolul 1
OBIECTUL DE STUDIU AL MICROBIOLOGIEI
Microbiologia este ştiinţa care se ocupă cu studiul microorganismelor.
Microorganismele constituie un grup foarte mare de organisme, diferite ca natură, formă şi activitate biologică, cu o organizare unicelulară şi o structură internă relativ simplă, având însă caractere comune dimensiunile microscopice, care le fac invizibile cu ochiul liber.
Din grupul microorganismelor fac parte bacteriile, ciupercile microscopice (mucegaiurile şi levurile), algele unicelulare şi protozoarele. Lumii microorganismelor i-au fost alăturate şi virusurile, care sunt entităţi infecţioase, alcătuite din componentele chimice esenţiale ale organismelor vii, proteine şi acizi nucleici cu o organizare acelulară, care le fac inapte de a se multiplica în afara unei celule vii, fapt ce le obligă la un parazitism intracelular absolut.
Deşi Microbiologia este o ştiinţă relativ tânără, datorită domeniului de studiu foarte vast, în cadrul ei s-au diferenţiat o serie de discipline microbiologice independente. Acestea pot fi grupate după mai multe criterii:
A. După grupul taxonomic studiat:
• Virologia (virusologia) este ştiinţa ce se ocupă cu studiul virusurilor şi acţiunea lor asupra organismelor vii.
• Bacteriologia se ocupă cu studierea, identificarea şi cultivarea bacteriilor, precum şi cu aplicaţiile lor în medicină, agricultură, industrie şi biotehnologie. Micologia (gr. mykes - ciupercă) studiază morfologia, genetica, taxonomia, proprietăţile biochimice şi ecologia ciupercilor.
• Algologia (ficologia) este ştiinţa ce se ocupă cu studiul algelor. • Protozoologia este ramura microbiologiei care studiază morfologia,
genetica şi taxonomia protozoarelor.

10
B. După activitatea microorganismelor în raport cu mediul în care trăiesc:
• Microbiologia acvatică se ocupă cu studiul microorganismelor care trăiesc în mediu acvatic şi care participă la transformarea substanţelor organice şi anorganice din mări şi oceane.
• Microbiologia solului studiază activitatea microbiotei solului şi rolul ei în nutriţia minerală a plantelor, fertilitatea solului, geneza şi formarea structurii solului.
C. După natura problemelor studiate: • Fiziologia microorganismelor studiază procesele vitale ale
microorganismelor (respiraţie, nutriţie etc). • Ecologia microorganismelor studiază legităţile generale de evoluţie
şi interacţiunea microorganismelor în natură, interacţiunile şi interrelaţiile dintre micro şi macroorganisme.
• Genetica microorganismelor se ocupă cu studiul eredităţii şi variabilităţii microorganismelor precum şi cu mecanismele de transfer al materialului genetic.
D. După aplicaţiile practice: • Microbiologia medicală umană şi veterinară studiază
microorganismele patogene şi relaţiile ecologice care se stabilesc cu organismele infectate.
• Imunologia studiază starea de rezistenţă sau de imunitate faţă de agenţii infecţioşi precum şi diferite funcţii şi proprietăţi ale celulelor şi ţesuturilor organismelor animale şi vegetale care determină prezenţa şi apariţia sau modificarea acestei stări.
• Microbiologia industrială (microbiologia tehnică sau a fermentaţiilor) studiază rolul microorganismelor în obţinerea pe cale fermentativă a unor substanţe chimice utile ca: alcoolul etilic, alcoolul butiric, acetona, acidul acetic, glicerina, antibioticele. În industria alimentară, microorganismele au rol în panificaţie, la conservarea alimentelor şi furajelor, precum şi în procese industriale ca topitul inului şi cânepei pe cale biologică, tăbăcirea pieilor etc.

11
• Microbiologia solului (agricolă) se ocupă de microflora naturală din sol şi de rolul acesteia în circuitul elementelor în natură, în asigurarea fertilităţii solului, în geneza şi formarea structurii solului, precum şi de relaţiile microorganismelor cu plantele. Cunoaşterea microflorei solului şi a activităţii ei biologice permite dirijarea unor procese naturale din sol şi folosirea lor în scopul sporirii producţiei agricole.
• Microbiologia insectelor studiază microflora normală a insectelor şi, în general, a artropodelor, relaţiile sale cu organismele gazdă, mecanismele de transmitere a unor agenţi patogeni la om, animale şi plante, precum şi microorganismele ce sunt patogene insectelor dăunătoare agriculturii şi omului.
• Microbiologia geologică se ocupă cu studiul rolului unor organisme în formarea unor depozite geologice (fier, mangan, sulf) în geneza rocilor calcaroase, a hidrocarburilor, a zăcămintelor de petrol, cărbuni, gaze naturale şi coroziunea rocilor; ea studiază, de asemenea, folosirea bacteriilor în detectarea depozitelor de petrol etc.
• Microbiologia marină (pelagică) studiază rolul microorganismelor în transformarea substanţelor organice şi anorganice din mări, influenţa lor asupra productivităţii biologice a mărilor şi în formarea diferitelor roci biogene şi a minereurilor organogene în mări.
• Microbiologia cosmică este ramura cea mai nouă a microbiologiei aplicate ce studiază probleme legate de influenţa spaţiului cosmic asupra viabilităţii şi variabilităţii microorganismelor, precum şi de posibilitatea exportului şi importului de microorganisme.
• Patologia vegetală (Fitopatologia) se ocupă cu studiul microorganismelor ce produc boli la plante, stabilind simptomatologia, căile de transmitere, relaţiile imunologice ale gazdelor, precum şi profilaxia şi tratamentul infecţiilor produse plantelor cultivate şi celor din flora spontană.
1.1. Date selective privind istoricul Microbiologiei Microbiologia este o ştiinţa relativ tânără, datorită faptului că, din
lipsa mijloacelor de investigaţie, oamenii de ştiinţă s-au ocupat, în primul rând, de vieţuitoarele vizibile. Existenţa microorganismelor ca agenţi etiologici ai unor boli era însă bănuită încă din antichitate.

12
Hipocrat (460-370 î.H.), întemeietorul medicinei, considera doi factori responsabili de aceste boli: unul intrinsec, reprezentat de contribuţia bolnavului, şi altul extrinsec, constând dintr-o alterare necunoscută a aerului, care devine nociv.
Această primă ipoteză asupra etiologiei bolilor infecţioase a persistat multă vreme sub forma teoriei miasmelor (gr. miasma = emanaţie putredă, murdărie).
Varon (100 î.H.) considera că alterarea aerului (gr. malaria = aer rău) este determinată de animale foarte mici, invizibile, care pătrund în organism prin gură şi prin nări.
Fracastor (1488-1553) intuieşte foarte corect una dintre caracteristicile esenţiale ale bolilor produse de microorganisme şi anume contagiozitatea lor. El arată că "infecţia este aceeaşi pentru cel care a primit-o şi pentru cel care a dat-o" şi că ea este produsă de "particule mici şi imperceptibile", care trec de la un organism bolnav la altul sănătos ("De Contagione", 1546).
Antony van Leeuwenhoek (1632-1723) reuşeşte descoperirea microorganismelor, folosind ca microscop un sistem de lentile biconvexe de construcţie proprie, cu o putere de mărire de 40-275x. El observă protozoare, alge, nematozi, levuri şi bacterii, pe care le descrie şi le desenează în lucrarea sa "Arcana naturae ope microscopiorum detecta" (Tainele naturii descoperite cu ajutorul microscoapelor, 1675). Deşi, descoperirile lui Leeuwenhoek au stârnit admiraţia şi interesul oamenilor de ştiinţă, studiul microorganismelor a rămas în continuare empiric, constituind doar o preocupare a amatorilor de curiozităţi ale naturii. În felul acesta se acumulează numai date disparate cu privire la forma, structura, mărimea şi distribuţia în natură a acestor microorganisme.
Fig. 1 – Hipocrat
Fig. 2 – Fracastoro

13
Karl Linnaeus, celebru botanist, în opera sa "Systema naturae"
(1735) încearcă o sistematizare a vieţuitoarelor cunoscute, reunind toate microorganismele într-un grup, denumit semnificativ "Chaos".
Terehovski, folosind pentru prima dată metode experimentale,
publică, în anul 1775, lucrarea "De Chao infusorii Linnaei", în care demonstrează că fiinţele foarte mici, observate în diferite infuzii bogate în substanţe organice, cresc, se divid şi se mişcă datorită unor forţe interne proprii, ajungând la concluzia că aceste organisme sunt adevărate animale în miniatură.
Louis Pasteur (1822-1895), savant francez, prin lucrările sale stabileşte principiile care stau şi azi la baza acestei ştiinţe.
Fig. 3 – Antony van Leeuwenhoek. (a) Microscop cu lentilă (1), dispozitiv de fixare (2) şi şuruburi de reglaj (3, 4). (b) Tipuri morfologice de bacterii desenate de Leeuwenhoeck.
Fig. 4 – Karl Linnaeus (1707 - 1778) şi lucrarea "Systema naturae".

14
Primele cercetări ale lui Pasteur au fost legate de fenomenul asimetriei moleculare. La acidul tartric racemic, compus inactiv pentru lumina polarizată, demonstrează că este un amestec echimolecular de acid dextrogir şi acid levogir, ale cărui cristale sunt de forme asimetrice şi enantiomorfe. De asemenea, a observat că, dacă în amestecul racemic, inactiv din punct de vedere optic, se dezvoltă un mucegai (Penicillium glaucum), soluţia devine optic activă, deviind, de astă dată, spre stânga planul luminii polarizate. Mucegaiul degradează izomerul dextrogir, lăsându-l intact pe cel levogir, care, astfel, poate fi izolat în stare pură. Această descoperire, pe lângă faptul că furniza un procedeu simplu de separare a izomerilor optici, a demonstrat şi intervenţia unei caracteristici fizice, ca disimetria moleculară în fenomenele chimice ale vieţii, dar, mai ales a atras atenţia lui Pasteur asupra unui fenomen spre care se va îndrepta în continuare curiozitatea sa de cercetător: microorganismele produc transformări ale substanţelor organice printr-o activitate selectivă foarte specifică.
Cercetările lui Pasteur asupra fermentaţiei vinului şi berii l-au condus la următoarele concluzii fundamentale:
• fermentaţiile sunt procese biologice, determinate de acţiunea unor microorganisme anaerobe, deci fermentaţia este viaţă fără aer;
• fiecare fermentaţie este produsă de un anumit tip de microorganism, care este specific, în sensul că determină o anumită transformare a mediului pe care creşte;
• dezvoltarea unui microorganism străin în mediul unde acţionează un microorganism cu acţiune fermentativă specifică deviază cursul normal al fermentaţiei în dauna calităţii (prin apariţia compuşilor nedoriţi) şi a randamentului ei în produs util, determinând o aşa-numită "boală" a fermentaţiei;
Fig. 5 – Louis Pasteur

15
• microorganismele străine care produc bolile vinului şi ale berii sunt consecinţa contaminărilor din aer, de pe vasele sau din ingredientele folosite în producţie, iar multiplicarea lor poate fi prevenită prin încălzire, procedeu utilizat sub denumirea de pasteurizare.
Până la Pasteur, ideea participării microorganismelor la determinarea unor boli era respinsă, ca nefiind probată pe cale experimentală. Pasteur extinde noţiunea de specificitate din domeniul fermentaţiilor în acela al patologiei omului şi animalelor, în sensul că orice boală infecţioasă este rezultatul activităţii vitale a unui anumit microorganism specific, care se dezvoltă ca parazit în organismul animal respectiv. Ulterior, cercetările acestuia au demonstrat principiul vaccinării şi stabilirea bazelor ştiinţifice ale preparării vaccinului, contribuind, în acelaşi timp, la descoperirea fenomenului de imunitate.
Studiind boala "holera găinilor", Pasteur observă că agentul ei patogen îşi pierde complet, prin învechirea culturii, capacitatea sa specifică de a produce această boală, adică devine avirulent. Inoculată la păsări sănătoase, cultura avirulentă le conferă însă rezistenţa faţă de holeră, în sensul că ele nu se mai îmbolnăvesc nici dacă sunt infectate cu o cultură proaspătă, virulentă, păsările "vaccinate" devenind "imune".
Pe acelaşi principiu, Pasteur a mai preparat şi utilizat cu succes vaccinul anticărbunos (1881), apoi vaccinul antirabic (1885).
L. Pasteur rezolvă şi problema generaţiei spontane (Fig. 5). Ignorarea unor principii esenţiale care, mai târziu, au stat la baza microbiologiei experimentale, în ceea ce priveşte sterilizarea şi asepsia, a făcut posibilă concluzia falsă că microorganismele pot lua naştere prin organizarea spontană a substanţelor organice din produsele supuse fermentaţiei sau putrefacţiei. Folosind baloane speciale cu "gât de lebădă", Pasteur demonstrează că un mediu de cultură sterilizat, în speţă bulionul de carne, poate rămâne steril timp îndelungat, dacă se evită contaminarea sa cu germeni din aer. Dacă prin înclinarea unui asemenea balon, mediul "spală" porţiunile incurbate ale gâtului rămas descoperit, porţiunile în care s-au depus bacteriile din aer, bulionul se infectează. Faptul că în baloanele cu mediu steril, închise ermetic, nu apar niciodată microorganisme, în timp ce acelaşi mediu se infectează, dacă este lăsat în contact cu aerul, dovedeşte că microorganismele nu iau naştere spontan din materia organică inanimată, ci

16
numai se înmulţesc în ea, pornind de la organisme similare, provenite din mediul înconjurător.
L. Pasteur a adoptat pentru denumirea organismelor microscopice
termenul de "microb" (trad. viaţă scurtă), creat de medicul francez Sedillot, iar noua ştiinţă, care studiază microbii, a rămas cu numele de microbiologie.
Robert Koch (1843-1910) a adus în dezvoltarea microbiologiei ca ştiinţă, contribuţii de ordin tehnic şi teoretic deosebit de valoroase, între care conceptul de "cultură pură", respectiv de organism cultivat într-un mediu lipsit de alte organisme. Realizarea culturilor pure este facilitată prin introducerea în tehnica microbiologiei de către Koch, a mediilor de cultură solidificate (cu gelatină sau agar).
Pe lângă faptul că a descoperit mai multe specii de bacterii patogene, între care bacilul tuberculozei (Mycobacterium tuberculosis) şi vibrionul holerei, Koch demonstrează, fără echivoc, că bacteria carbonoasă (Bacillus anthracis) este agentul patogen al bolii denumită antrax sau cărbune,
Fig. 5 – Experimentul lui Pasteur prin care combate teoria generaţiei spontane. A: mediul nutritiv este introdus într-un balon; B. curbarea gâtului balonului (gât de lebădă"); C. sterilizarea mediului nutritiv; D. mediul nutritiv rămâne steril fiind protejat de pătrunderea microorganismelor din atmosferă prin forma gâtului.
Fig. 6 – Robert Koch (1843-1910)

17
pe care îl cultivă "in vitro" în ser sanguin, studiind formarea de spori termorezistenţi, alternanţa de sporulare şi germinare, precum şi faptul că infecţia naturală a animalelor provine din sol prin iarba contaminată.
Pe baza acestor cercetări, R. Koch stabileşte criteriile indispensabile pentru ca un microorganism izolat dintr-un organism afectat de o anumită boală să poată fi considerat în mod justificat ca agentul cauzal al bolii respective. Aceste criterii, cunoscute sub numele de postulatele lui Koch, sunt următoarele: • microorganismul incriminat să poată fi pus întotdeauna în evidenţă în toate cazurile bolii respective, iar distribuţia lui să corespundă leziunilor caracteristice bolii; • să poată fi utilizat în cultură pură, pe medii artificiale; • după ce este cultivat "in vitro", mai multe generaţii să reproducă boala şi leziunile specifice la animalele sensibile, de la care să poată fi reizolat.
Postulatele lui Koch au reprezentat şi reprezintă încă un principiu eficace de lucru, a cărui respectare previne interpretările eronate, în materie de diagnostic bacteriologic sau de identificare a unor agenţi infecţioşi.
Ilia Metchnikoff (1845-1916), biolog rus, studiind digestia intracelulară la echinoderme, demonstrează capacitatea anumitor celule de a capta şi îngloba diferite particule cu care vin în contact şi emite ipoteza că celulele cu funcţii asemănătoare, denumite de el "fagocite", ar putea exista şi în organismul altor animale.
Ulterior (1884), studiind infecţia crustaceului Daphnia magna cu ciuperca Monospora (Metschnikowia) bicuspidata, observă că sporii aciculari ai levurii pătrund odată cu hrana în tubul digestiv al dafniei, de unde, străpungând peretele acestuia, trec în cavitatea generală, unde sunt atacaţi de celule mobile. Când infecţia este moderată, amoebocitele înglobează şi digeră toţi sporii ciupercii şi, astfel, crustaceul supravieţuieşte. Când însă
Fig. 7 – Ilia Metchnikoff

18
infecţia este masivă, sporii rămaşi nefagocitaţi germinează şi dau naştere formei vegetative a ciupercii, care, prin multiplicare, invadează tot organismul dafniei, determinându-i moartea.
Metchnikoff extinde rezultatele acestor observaţii la animalele superioare şi la om, demonstrând prezenţa şi importanţa celulelor cu proprietăţi fagocitare în reacţiile de apărare ale organismului faţă de bacteriile patogene, stabilind, astfel, bazele imunităţii celulare.
Dimitri Ivanovski (1864-1920) demonstrează că mozaicul tutunului este produs de un agent patogen invizibil la microscop, care traversează filtrele bacteriene şi poate fi transmis de la o plantă bolnavă la una sănătoasă prin intermediul filtratului acelular al trituratului de frunze, cu leziuni.
Martinus Beijerinck (1851-1931) confirmă în 1898 filtrabilitatea agentului patogen şi intuieşte natura deosebită a agentului patogen, pe care îl considera ca agent contagios viu
(contagium vivum fluidum). Această intuiţie a făcut din Beijerinck adevăratul intemeietor al virologiei ca ştiinţă.
Frederick Twort (1877-1950) şi Felix d'Herelle (1873-1949) descoperă, în perioada 1915-1917, bacteriofagii (trad. mâncător de bacterii) şi fenomenul bacteriofagic.
Sergei Winogradsky (1856-1953), lucrând la Institutul Pasteur din Paris, a descris procesul de asimilare la organismele chimiosintetizante şi fenomenul de fixare a azotului atmosferic de către microorganisme. În acelaşi timp el a elaborat metode speciale pentru cercetarea activităţii microorganismelor din sol, fiind considerat intemeietorul Microbiologiei solului.
Fig. 8 – Dimitri Ivanovski
Fig. 9 – Martinus Beijerinck

19
Alexander Fleming (1881-1955), observă în anul 1929 că unele culturi de Penicillium, elaborează o substanţă cu proprietăţi microbiene
specifice – penicilina. Acest prim antibiotic a fost mai târziu purificat de Florey şi Chain (1940). Prin lucrările sale, Fleming deschide era antibioticelor, de o importanţă excepţională în medicină şi biologie.
Stanley Prusiner (1942- ), neurolog şi biochimist american, laureat al Premiului Nobel în 1997. A descoperit că encefalopatia spongiformă la bovine şi boala Creutzfeldt-Jakob la om sunt produse de particule infecţioase de natură proteică, pe care le-a numit prioni.
1.2. Şcoala românească de Microbiologie Victor Babeş (1854-1926), fondatorul
şcolii româneşti de microbiologie, a desfăşurat o prodigioasă activitate de cercetător, studiind numeroase boli ale omului, printre care lepra, tuberculoza, holera, febra tifoidă şi, mai ales, turbarea.
În colaborare cu Cornil, este autorul primei lucrări de sinteză în bacteriologie şi
Fig. 10 – Alexander Fleming
Fig. 11 – Stanley Prusiner
Fig. 12 – Victor Babeş

20
anatomie patologică din lume, apărută în 1885: “Bacteriile şi rolul lor în etiologia, anatomia şi histologia patologică a bolilor infecţioase”.
A studiat, de asemenea, asociaţiile microbiene, fiind primul cercetător care, după Pasteur, şi-a dat seama de importanţa terapeutică a antagonismului microbian.
Ioan Cantacuzino (1863-1934) este creatorul Institutului de Microbiologie, care îi poartă numele, şi al şcolii contemporane de microbiologie din ţara noastră. Cele mai importante lucrări sunt referitoare la aparatele şi funcţiile fagocitare în regnul animal şi problema imunităţii la nevertebrate. Cantacuzino a studiat, de asemenea, diferite boli ca holera, febra tifoidă, scarlatina, tuberculoza, creând şi organizând condiţiile producţiei de seruri şi vaccinuri în ţara noastră.
Constantin Ionescu-Mihăieşti (1883-1962) a adus contribuţii ştiinţifice valoroase în domeniul virologiei, imunologiei şi microbiologiei generale.
Dumitru Combiescu (1887-1961) a desfăşurat o amplă activitate în domeniul rickettsiozelor şi zoonozelor.
Mihai Ciucă (1883-1969) este autorul unor lucrări deosebit de importante în domeniul bacteriofagilor, paludismului, salmonelozelor, difteriei.
Ştefan Nicolau (1896-1967) este creatorul şcolii româneşti de virologie şi fondatorul institutului respectiv. A publicat numeroase lucrări originale în domeniul herpesului, turbării, febrei aftoase, febrei galbene, hepatitelor virale.
Traian Săvulescu (1889-1963) este creatorul şi îndrumătorul şcolii româneşti de fitopatologie. Este autorul unor lucrări de sinteză ca Monografia uredinalelor, Monografia ustilaginalelor şi al unor lucrări originale în fitopatologie şi în domeniul imunităţii plantelor faţă de bolile bacteriene.
Fig. 13 – Ioan Cantacuzino
21
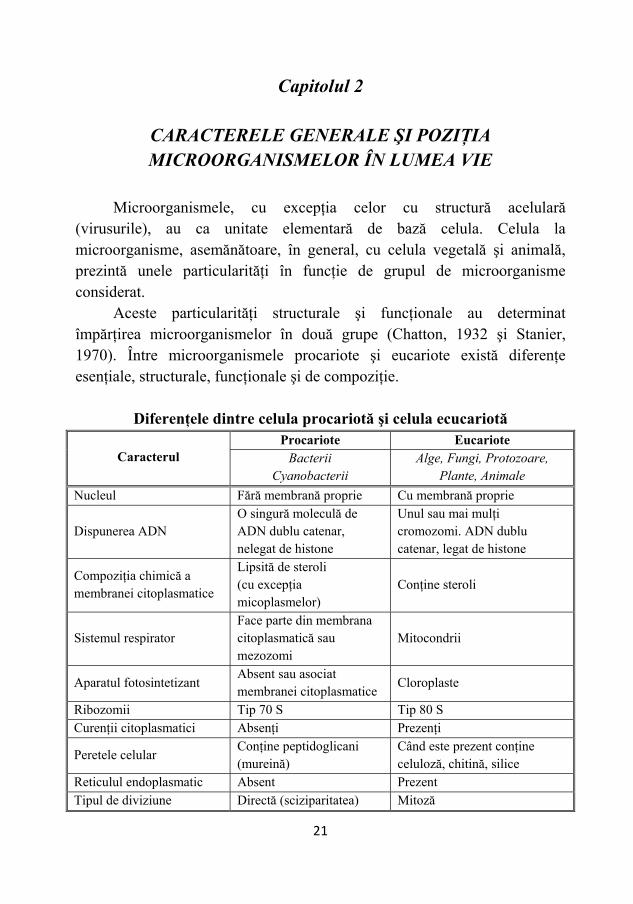
Capitolul 2
CARACTERELE GENERALE ŞI POZIŢIA MICROORGANISMELOR ÎN LUMEA VIE
Microorganismele, cu excepţia celor cu structură acelulară
(virusurile), au ca unitate elementară de bază celula. Celula la microorganisme, asemănătoare, în general, cu celula vegetală şi animală, prezintă unele particularităţi în funcţie de grupul de microorganisme considerat.
Aceste particularităţi structurale şi funcţionale au determinat împărţirea microorganismelor în două grupe (Chatton, 1932 şi Stanier, 1970). Între microorganismele procariote şi eucariote există diferenţe esenţiale, structurale, funcţionale şi de compoziţie.
Diferenţele dintre celula procariotă şi celula ecucariotă Procariote Eucariote
Caracterul Bacterii Cyanobacterii
Alge, Fungi, Protozoare, Plante, Animale
Nucleul Fără membrană proprie Cu membrană proprie
Dispunerea ADN O singură moleculă de ADN dublu catenar, nelegat de histone
Unul sau mai mulţi cromozomi. ADN dublu catenar, legat de histone
Compoziţia chimică a membranei citoplasmatice
Lipsită de steroli (cu excepţia micoplasmelor)
Conţine steroli
Sistemul respirator Face parte din membrana citoplasmatică sau mezozomi
Mitocondrii
Aparatul fotosintetizant Absent sau asociat membranei citoplasmatice
Cloroplaste
Ribozomii Tip 70 S Tip 80 S Curenţii citoplasmatici Absenţi Prezenţi
Peretele celular Conţine peptidoglicani (mureină)
Când este prezent conţine celuloză, chitină, silice
Reticulul endoplasmatic Absent Prezent Tipul de diviziune Directă (sciziparitatea) Mitoză
22
Ribozom
CapsulăPerete celular
Membrană citoplasmatică
Incluziune
Citoplasmă Material nuclear (ADN)
Fimbrii
Plasmid
Flageli
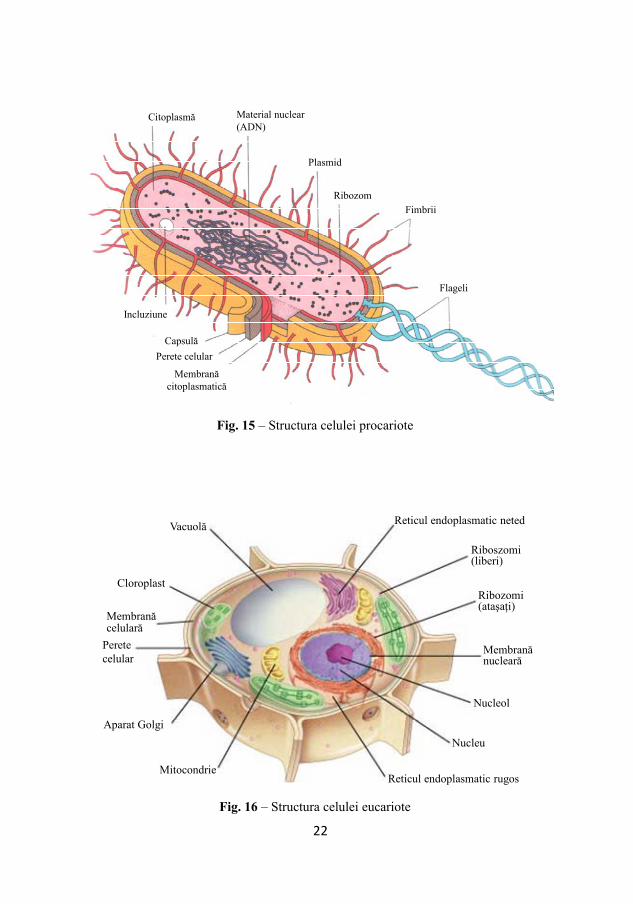
Fig. 15 – Structura celulei procariote
Membrană nucleară
Ribozomi(ataşaţi)
Riboszomi(liberi)
Reticul endoplasmatic neted
Nucleu
Reticul endoplasmatic rugos
Nucleol
Aparat Golgi
Mitocondrie
Perete celular
Membranăcelulară
Cloroplast
Vacuolă
Fig. 16 – Structura celulei eucariote

23
2.1. Poziţia microorganismelor în lumea vie
După acumularea a numeroase cunoştinţe asupra microorganismelor a fost necesară introducerea acestora într-un grup taxonomic. Iniţial microorganismele au fost incluse în regnul Plantae, iar ulterior au fost incluse în diferite sisteme de clasificare.
A. Sistemul tradiţional:
1. Regnul Plantae; 2. Regnul Animalia.
B. Sistemul lui Hogg şi Haeckel (1866): 1. Regnul Monera; 2. Regnul Plantae; 3. Regnul Animalia.
C. Sistemul lui Copeland (1938): 1. Regnul Procaryotae (Monera); 2. Regnul Protista; 3. Regnul Plantae; 4. Regnul Animalia.
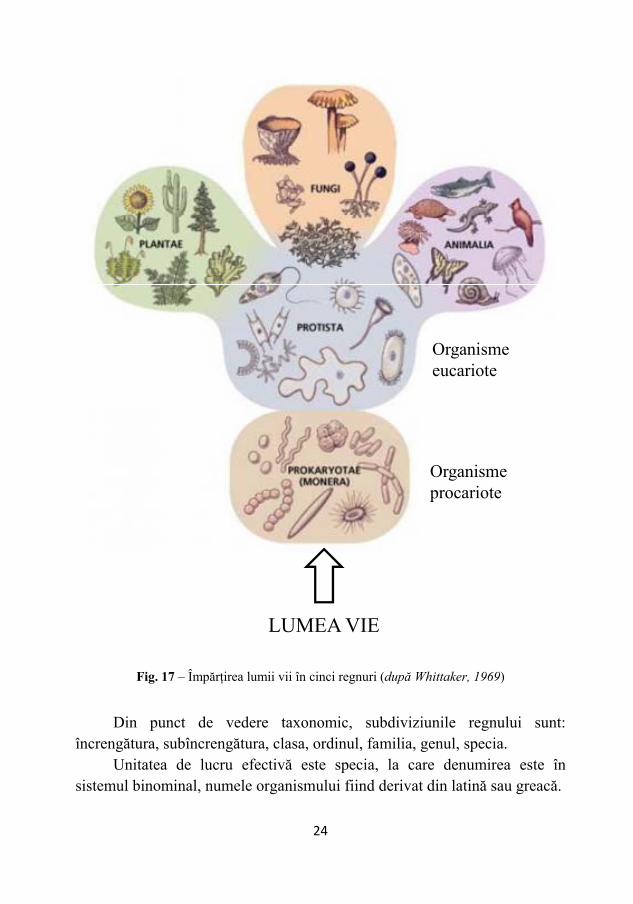
D. Sistemul lui Whittaker (1969): 1. Regnul Procaryotae (Monera) (bacterii, cianobacterii = alge albastre-verzi); 2. Regnul Protista (protozoare, diatomee); 3. Regnul Fungi (ciuperci); 4. Regnul Plantae (alge verzi, brune, roşii, brofite, traheofite); 5. Regnul Animalia (animale multicelulare).
Astăzi, majoritatea specialiştilor din domeniul microbiologiei au
acceptat sistemul de clasificare elaborat de Whittaeker (Fig. 17) în anul 1969, care corespunde necesităţilor actuale.
24
LUMEA VIE
Organisme procariote
Organisme eucariote
Fig. 17 – Împărţirea lumii vii în cinci regnuri (după Whittaker, 1969)
Din punct de vedere taxonomic, subdiviziunile regnului sunt:
încrengătura, subîncrengătura, clasa, ordinul, familia, genul, specia. Unitatea de lucru efectivă este specia, la care denumirea este în
sistemul binominal, numele organismului fiind derivat din latină sau greacă.

25
În sistemele de clasificare prezentate anterior sunt cuprinse numai organismele care au ca unitate de bază, celula, eucariotă sau procariotă, nefiind introduse entităţile infecţioase, cum sunt virusurile sau prionii, şi care au fost "alăturate" acestor sisteme de clasificare.
2.2. Prionii
Prionii sunt agenţi infecţioşi neconvenţionali de natură proteică, lipsiţi de orice tip de acid nucleic, care produc un grup de boli neurodegenerative, transmisibile la animale şi om, numite boli prionice. Procesul care declanşează boala este reprezentat de conversia unei proteine normale, sintetizată în mod natural în creierul tuturor mamiferelor (PrPc), într-una mutantă, patogenă (PrPSc).
Prima semnalare a unei boli de natură prionică a fost făcută în Anglia, în anul 1732, la ovine.
Astăzi, sunt cunoscute ca boli certe, produse de prioni, următoarele: - la animale: 1. encefalopatia spongiformă la ovine, denumită popular scrapie
(engl.); tramblanta (fr.). În România mai este utilizat, impropriu, termenul de "căpiala" oilor, dar aceasta este o boală bine cunoscută şi este produsă de un parazit animal. Medicii veterinari avizaţi folosesc termenul francez de tramblanta oilor.
2. encefalopatia spongiformă bovină (BSE sau "boala vacii nebune"); 3. encefalopatia spongiformă la feline; 4. encefalopatia spongiformă la nurci; 5. encefalopatia spongiformă la hamsteri. - la om: 1. Boala "Kuru". Aceasta este prima semnalată şi cel mai bine
studiată. A fost depistată la un trib local în Papua - Noua Guinee. După interzicerea consumului de carne în stare proaspătă, această boală a fost eradicată.
2. Boala Creutzfeldt-Jacob (CJD); 3. Sindromul Alpers (la copii); 4. Sindromul Gerstmann-Straussler-Scheinker (GSS); 5. Insomnia fatală familială (FFI).

26
Agentul infecţios În prezent, se consideră că agentul infecţios este o moleculă proteică,
cu greutatea moleculară de 28 kilodaltoni, alcătuită dintr-un lanţ de 208-220 de aminoacizi, în funcţie de specie, şi care prezintă o configuraţie spaţială, asemănătoare unui metru de tâmplărie, parţial destins. Prezintă un capăt NH2-terminal, o regiune centrală şi un capăt COOH-terminal.
Această proteină a fost izolată din creierul tuturor cazurilor bolnave studiate. Surpriza a sosit în momentul studierii în paralel (la oi) a creierului la animale sănătoase şi care prezintă o proteină celulară similară. Lanţul de aminoacizi este identic cu excepţia unui singur aminoacid (schimbat).
Proteina celulară normală (PrPc) prezintă o configuraţie spaţială
asemănătoare unui resort. Specialiştii din biochimie consideră că, prin schimbarea unui singur aminoacid din lanţ, nu se poate modifica configuraţia spaţială. În acelaşi timp, proteina infecţioasă (PrPSc) rezistă la atacul proteazelor celulare.
Se consideră că procesul infecţios se desfăşoară astfel: celula prepară proteina celulară normală care se acumulează în lizozomi, aceştia o transportă la exteriorul celulei şi în momentul în care această proteină pătrunde din nou în celulă este degradată de proteaze.
Fig. 18 – Proteina prionică (huPrP) responsabilă pentru o serie de boli
prionice la om (www.itqb.unl.pt).

27
În cazul proteinei infecţioase, aceasta se acumulează în lizozomi în mod continuu, până când aceştia "crapă". Enzimele din interiorul lor degradează componentele celulare şi, în final, întreaga celulă nervoasă. În locul acesteia rămâne un orificiu. Din acest motiv, după un timp, creierul atacat arată ca un burete (cu numeroase orificii). În stadiul final al bolii, creierul degenerează complet, având dimensiunea unei nuci.
Studiile cele mai ample sunt efectuate la animale, la om ele fiind extrem de puţine, deoarece boala poate fi studiată numai după moartea individului.
Prionii pot rezista la acţiunea multor factori fizico-chimici, dintre care amintim: formol 10% (timp de 28 de luni), caldură (rezistă la fierbere timp de 3 ore), factori inhibitori ai acizilor nucleici, precum şi la acţiunea radiaţiilor UV.
2.3. Virusurile
Virusurile (lat. virus – otravă, infecţie, lichid otrăvitor) reprezintă o
categorie specifică de agenţi infecţioşi, structural şi fiziologic fundamental diferiţi de oricare dintre microorganismele cunoscute (Zarnea, 1983).
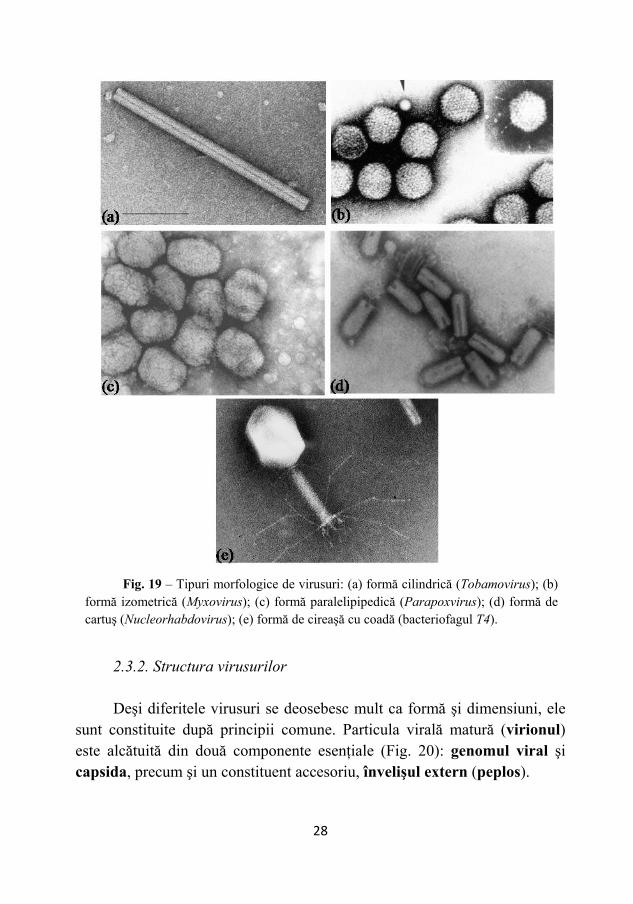
2.3.1. Morfologia virusurilor Din punct de vedere morfologic (Fig. 19), virusurile pot aparţine
următoarelor tipuri principale: formă cilindrică-alungită sau de bastonaş (virusul mozaicului tutunului - Tobamovirus); formă sferică (izometrică), sferoidală (virusul gripal - Myxovirus), formă paralelipipedică (virusul variolei - Parapoxvirus), formă de cartuş sau obuz (virusul piticirii galbene la cartof - Nucleorhabdovirus), formă de mormoloc, spermatozoid sau cireaşă cu coadă (unii bacteriofagi).
Dimensiunile virusurilor: 17 - 2500 nanometri (1 nm = 10-9 m).
28
2.3.2. Structura virusurilor Deşi diferitele virusuri se deosebesc mult ca formă şi dimensiuni, ele
sunt constituite după principii comune. Particula virală matură (virionul) este alcătuită din două componente esenţiale (Fig. 20): genomul viral şi capsida, precum şi un constituent accesoriu, învelişul extern (peplos).
Fig. 19 – Tipuri morfologice de virusuri: (a) formă cilindrică (Tobamovirus); (b)
formă izometrică (Myxovirus); (c) formă paralelipipedică (Parapoxvirus); (d) formă de cartuş (Nucleorhabdovirus); (e) formă de cireaşă cu coadă (bacteriofagul T4).

29
Genomul viral este reprezentat, în mod obişnuit, printr-o moleculă de acid nucleic (ADN sau ARN, niciodată ambele). Virusurile cu genom ADN sunt numite dezoxiribovirusuri, iar cele cu genom ARN – ribovirusuri. Genomul viral poartă informaţia genetică necesară replicării în sensul sintezei constituenţilor virali şi a precursorilor acestora.
Capsida virală (gr. kapsa – cutie) acoperă genomul, fiind alcătuită din subunităţi proteice, denumite capsomere. Capsomerele sunt constituite din molecule proteice, aşezate în mod regulat, formând, în ansamblu, structura specifică a virusului. Capsida protejează materialul genetic.
Capsida şi genomul viral formează nucleocapsida. La unele virusuri, nucleocapsida este acoperită de o structură
trilamelară, numită înveliş extern sau peplos (gr. peplos – manta), ce poate prezenta la exterior nişte proeminenţe de suprafaţă, denumite spicule.
2.3.3. Virusurile plantelor Virusurile plantelor au o mare importanţă ca agenţi patogeni, datorită
marii lor răspândiri şi faptului că acelaşi virus poate infecta plante, care aparţin de diferite familii botanice.
Bolile produse de virusuri la plante sunt denumite viroze. Până în prezent sunt cunoscute câteva sute de boli virale, produse de virusuri, care aparţin la 25 genuri distincte. Majoritatea virusurilor plantelor au genom ARN monocatenar.
Capsidă
Înveliş extern(peplos)
Spicule
Acid nucleic(ADN sau ARN)
Fig. 20 – Structura unor virusuri şi denumirea constituenţilor virali

30
Din punct de vedere morfologic, virusurile plantelor aparţin la două grupuri: grupul virusurilor izometrice (izodiametrice) sau sferice şi grupul virusurilor alungite.
În general, virusurile plantelor au o structură chimică mai simplă decât virusurile animale şi bacteriofagii.
Mecanismul de transmitere Virusurile se pot transmite de la planta bolnavă la planta sănătoasă pe
mai multe căi: A - transmitere mecanică: prin contact între frunze; anastomoze
radiculare; unelte de lucru; altoire. B - transmiterea prin vectori se realizează prin intermediul insectelor
(cca..400 specii): afide şi cicade. Relaţia dintre virus - insectă vectoare este variabilă şi corespunde următoarelor trei situaţii: • Virusuri nepersistente pot fi transmise până la circa 4 ore de la achiziţie. • Virusuri semipersistente au capacitatea de infectare variabilă de la 10-100 ore de la achiziţie. • Virusuri persistente cu o capacitate de infectare de peste 100 ore (uneori chiar toată durata de viaţă a vectorului).
În funcţie de răspândirea şi comportarea virusurilor în corpul vectorului, acestea se împart în trei categorii: • Virusuri localizate pe stilet (virusuri nepersistente). • Virusuri circulante (virusuri persistente) pot fi transmise timp îndelungat ajungând în hemolimfa insectei. • Virusuri propagative (virusuri persistente) se multiplică în corpul vectorului fiind transmise toată viaţa insectei.
C - transmiterea prin intermediul ciupercilor fitopatogene din sol, (ex. Polymyxa betae transmite virusul rizomaniei la sfecla pentru zahăr).
D - transmiterea prin intermediul nematozilor. E - transmiterea prin seminţe (cca.. 1/3 din virusuri). Circulaţia virusurilor în interiorul plantelor se realizează prin
parenchim (plasmodesme), prin floem şi prin xilem.

31
2.3.4. Bacteriofagii Bacteriofagii (gr. bakterion – bastonaş; gr. phago ein – a mânca) sunt
virusuri adaptate la viaţa parazitară în celulele bacteriene şi, care, prin multiplicare, produc liza acestora.
În anul 1915, Twort descoperă fenomenul de liză transmisibilă, dar nu poate explica cu exactitate cauza acesteia. D'Herelle (1917) demonstrează natura particulară a fagului şi îl consideră ca virus, determinându-l bacteriofag (mâncător de bacterii). În anul 1940, ia fiinţă un grup de cercetători, denumit "grupul fag", condus de Delbrück, sistemul bacterie-bacteriofag fiind folosit ca model experimental pentru cele mai importante studii de genetică moleculară.
Structura fagilor diferă de la un grup la altul, în majoritate aceştia încadrându-se în două tipuri de bază şi anume: • tipul icosaedric (poliedric); • tipul filamentos.
Datele cele mai numeroase, cunoscute în prezent, sunt cele referitoare la fagii din seria T, îndeosebi la fagi T-par (T2, T4, T6).
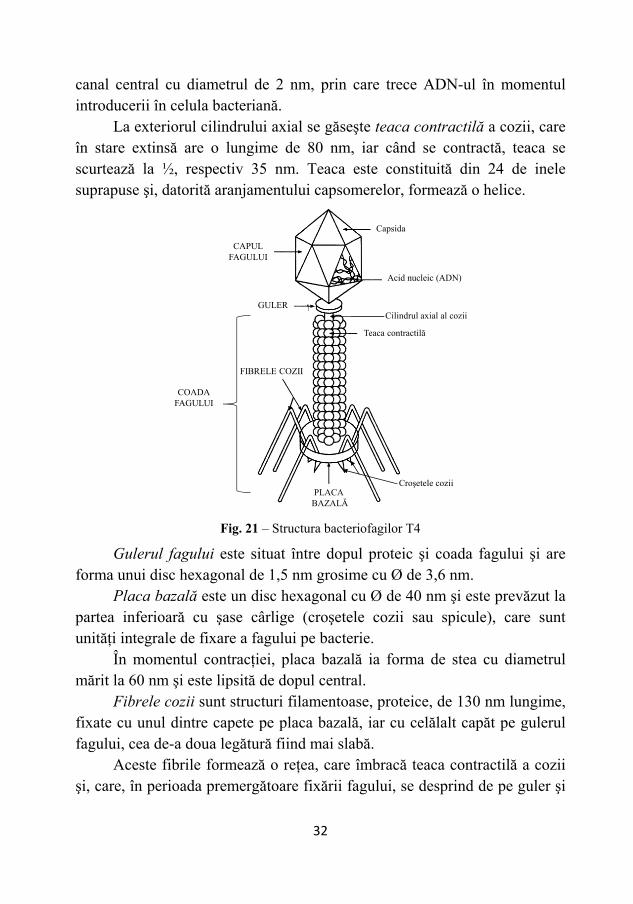
Anatomia fagilor T-par Particula virală matură a fagilor T-par cu g.m. 2,2x1010 este alcătuită
din ADN şi proteină şi, din punct de vedere anatomic, prezintă următoarele componente: cap, guler, coadă, placă bazală şi fibrele cozii (Fig. 21).
Capul fagului, la microscop, prezintă o formă poliedrică, iar în secţiune are o formă hexagonală, cu o lungime de 100 nm şi o lăţime de 65 nm. Acesta este constituit din capsomere cu diametrul de 4 nm, dar nu este cunoscut, încă, numărul şi modul de aşezare al acestora.
În interiorul capului, se găseşte genomul, alcătuit dintr-o moleculă de ADN d.c., liniară, cu lungimea de 50 μm, ce cuprinde 200 gene, fiind împachetată foarte strâns.
În partea bazală a capului, la locul de prindere al cozii, se află un dop proteic, cu rol de articulare mecanică.
Între acest dop şi placa bazală se găseşte un tub lung de 120 nm, reprezentat de cilindrul axial al cozii. El are un diametru de 7,5 nm şi un
32
canal central cu diametrul de 2 nm, prin care trece ADN-ul în momentul introducerii în celula bacteriană.
La exteriorul cilindrului axial se găseşte teaca contractilă a cozii, care în stare extinsă are o lungime de 80 nm, iar când se contractă, teaca se scurtează la ½, respectiv 35 nm. Teaca este constituită din 24 de inele suprapuse şi, datorită aranjamentului capsomerelor, formează o helice.
Gulerul fagului este situat între dopul proteic şi coada fagului şi are
forma unui disc hexagonal de 1,5 nm grosime cu Ø de 3,6 nm. Placa bazală este un disc hexagonal cu Ø de 40 nm şi este prevăzut la
partea inferioară cu şase cârlige (croşetele cozii sau spicule), care sunt unităţi integrale de fixare a fagului pe bacterie.
În momentul contracţiei, placa bazală ia forma de stea cu diametrul mărit la 60 nm şi este lipsită de dopul central.
Fibrele cozii sunt structuri filamentoase, proteice, de 130 nm lungime, fixate cu unul dintre capete pe placa bazală, iar cu celălalt capăt pe gulerul fagului, cea de-a doua legătură fiind mai slabă.
Aceste fibrile formează o reţea, care îmbracă teaca contractilă a cozii şi, care, în perioada premergătoare fixării fagului, se desprind de pe guler şi
CAPULFAGULUI
Capsida
Acid nucleic (ADN)
GULERCilindrul axial al cozii
Teaca contractilă
PLACABAZALĂ
FIBRELE COZII
Croşetele cozii
COADAFAGULUI
Fig. 21 – Structura bacteriofagilor T4

33
rămân legate numai pe placa bazală, înotând libere în mediu, având aspectul unor picioare de păianjen.
Structura genomului Bacteriofagii prezintă, în majoritate, un genom format dintr-o singură
moleculă de acid nucleic (ADN m.c., ADN d.c. , ARN m.c.), cu o singură excepţie, fagul Ø 6 de la Pseudomonas phaseolicola, care are un genom ARN segmentat, format din trei segmente.
Genomul bacteriofagilor se deosebeşte de genomul virusurilor animale şi vegetale prin prezenţa unor baze nucleice neobişnuite, cum ar fi la bacteriofagii T-par, în loc de citozină prezintă 5-hidroximetilcitozină, iar la fagii de la Bacillus subtilis, 5-hidroximetilluracil şi 5-4,5 hidroximetiluracil.
Legat de acest aspect s-a emis ipoteza că aceste baze nucleice ar avea rol de protecţie a acidului nucleic fagic împotriva acţiunii unor enzime virale (nucleaze) care atacă numai acidul nucleic străin şi, în felul acesta, poate degrada selectiv cromozomul bacterian în cursul sintezei de virus.
Infecţia celulei bacteriene. Replicarea Între fagi şi bacteriile infectate se pot stabili două tipuri de relaţii,
funcţie de ciclul de viaţă al bacteriofagilor. Dacă bacteriofagii sunt virulenţi, infectarea bacteriei duce la formarea
de noi virioni, care, în urma fenomenului de liză bacteriană, sunt eliberaţi în mediu. Acest tip de relaţie poartă numele de ciclu litic şi facilitează formarea şi eliberarea a 100-200 de virioni, în aproximativ 20 de minute.
În cazul în care bacteriofagii sunt temperaţi, genomul acestora se integrează în cromozomul bacterian, se replică concomitent cu acesta şi se distribuie la celulele fiice, fără a-şi manifesta funcţiile virale. Această manifestare poartă numele de ciclu lizogenic, iar genomul fagic integrat în genomul celulei bacteriene a fost numit profag (provirus). După mai multe generaţii, profagul se desprinde de cromozomul bacterian şi devine fag litic care lizează celula bacteriană şi eliberează în mediul înconjurător virioni lizogeni.
Ciclul de replicare vegetativă (ciclul litic) a fagului are următoarele etape:

34
I. Adsorbţia reprezintă procesul de fixare a particulelor virale pe suprafaţa bacteriilor. După o serie de ciocniri întâmplătoare, fagul se fixează pe peretele celular prin intermediul fibrelor cozii, realizând faza de fixare iniţială, care poate fi reversibilă. Urmează faza de fixare ireversibilă, realizată prin intermediul croşetelor, care este condiţionată de prezenţa receptorilor de fag (Fig. 22). Aceştia sunt reprezentaţi de orice structură existentă la suprafaţa celulei bacteriene sau pe pilii şi flagelii acesteia, iar sinteza lor este controlată de gene bacteriene.
II. Injectarea genomului viral în celula bacteriană. După fixarea
ireversibilă pe peretele celular are loc o contracţie a cozii fagului, axul central al acesteia pătrunzând în adâncimea peretelui celular 12 nm, iar genomul este injectat prin intermediul cilindrului axial tubular (Fig. 23). Nu se cunosc încă forţele care proiectează genomul viral în interiorul celulei, injectarea făcându-se rapid (15 secunde la T4).
Fig. 23 – Injectarea genomului viral în celula bacteriană
Material nuclear bacterian
Fig. 22 – Adsorbţia particulelor virale pe suprafaţa bacteriilor
35
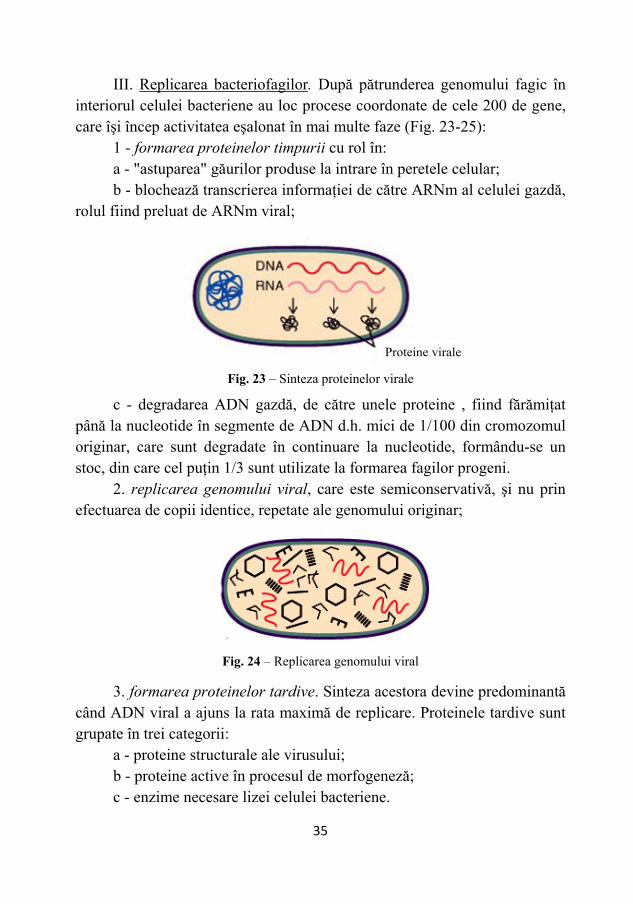
III. Replicarea bacteriofagilor. După pătrunderea genomului fagic în interiorul celulei bacteriene au loc procese coordonate de cele 200 de gene, care îşi încep activitatea eşalonat în mai multe faze (Fig. 23-25):
1 - formarea proteinelor timpurii cu rol în: a - "astuparea" găurilor produse la intrare în peretele celular; b - blochează transcrierea informaţiei de către ARNm al celulei gazdă,
rolul fiind preluat de ARNm viral;
c - degradarea ADN gazdă, de către unele proteine , fiind fărămiţat
până la nucleotide în segmente de ADN d.h. mici de 1/100 din cromozomul originar, care sunt degradate în continuare la nucleotide, formându-se un stoc, din care cel puţin 1/3 sunt utilizate la formarea fagilor progeni.
2. replicarea genomului viral, care este semiconservativă, şi nu prin efectuarea de copii identice, repetate ale genomului originar;
3. formarea proteinelor tardive. Sinteza acestora devine predominantă
când ADN viral a ajuns la rata maximă de replicare. Proteinele tardive sunt grupate în trei categorii:
a - proteine structurale ale virusului; b - proteine active în procesul de morfogeneză; c - enzime necesare lizei celulei bacteriene.
Fig. 24 – Replicarea genomului viral
Proteine virale Fig. 23 – Sinteza proteinelor virale
36
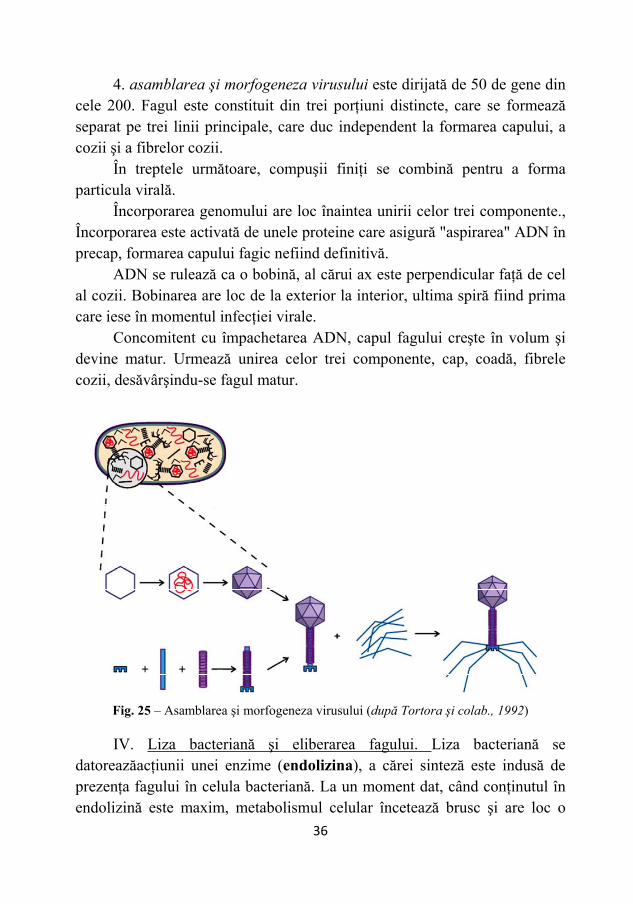
4. asamblarea şi morfogeneza virusului este dirijată de 50 de gene din cele 200. Fagul este constituit din trei porţiuni distincte, care se formează separat pe trei linii principale, care duc independent la formarea capului, a cozii şi a fibrelor cozii.
În treptele următoare, compuşii finiţi se combină pentru a forma particula virală.
Încorporarea genomului are loc înaintea unirii celor trei componente., Încorporarea este activată de unele proteine care asigură "aspirarea" ADN în precap, formarea capului fagic nefiind definitivă.
ADN se rulează ca o bobină, al cărui ax este perpendicular faţă de cel al cozii. Bobinarea are loc de la exterior la interior, ultima spiră fiind prima care iese în momentul infecţiei virale.
Concomitent cu împachetarea ADN, capul fagului creşte în volum şi devine matur. Urmează unirea celor trei componente, cap, coadă, fibrele cozii, desăvârşindu-se fagul matur.
IV. Liza bacteriană şi eliberarea fagului. Liza bacteriană se
datoreazăacţiunii unei enzime (endolizina), a cărei sinteză este indusă de prezenţa fagului în celula bacteriană. La un moment dat, când conţinutul în endolizină este maxim, metabolismul celular încetează brusc şi are loc o
Fig. 25 – Asamblarea şi morfogeneza virusului (după Tortora şi colab., 1992)

37
distrugere masivă a peretelui celular, cu un caracter exploziv, fagii fiind expulzaţi în exterior (Fig. 26).
2.3.5. Fagii filamentoşi Fagii filamentoşi (Fig. 27), care infectează E. coli, au forma unor
bastonaşe flexibile cu diametrul de 6 nm şi o lungime de 1000-2000 nm. Structura este aceea a unui
cilindru proteic gol, deschis la capete (capsidă), în interior aflându-se genomul fagic, format din ADN m.c., o moleculă circulară.
Fagii filamentoşi infectează numai bacteriile cu caracter mascul, ca şi fagii cu ARN, adsorbţia făcându-se pe receptori speciali situaţi la extremitatea liberă a pililor de sex.
Fagii maturi sunt eliminaţi, fără ca bacteria să fie lizată, prin intermediul unor pori, care se formează în membrana citoplasmatică. Bacteria continuă să se dividă pentru o perioadă de timp, iar ulterior conţinutul celular al acesteia trece la exterior prin porii deschişi de virioni.
Fig. 26 – Liza bacteriană
Fig. 27 – Structura bacteriofagului M13

38
2.3.6. Cianofagii Cianofagii (sin. ficovirusuri, algofagi) sunt agenţi virali, capabili să
producă infecţii la numeroase specii de cianobacterii (alge albastre-verzi), influenţând, astfel, dinamica acestor populaţii, în sensul menţinerii unui echilibru în bazinele acvatice naturale.
Sunt similari ca structură şi evoluţie cu bacteriofagii T-impar, genomul fiind reprezentat de ADN dublu catenar de 13,2 μm.
2.3.7. Micovirusurile Sunt virusuri care atacă fungii, fiind cunoscute şi sub denumirea de
micofagi. Din cele peste 50 de genuri atacate menţionăm: Penicillium, Aspergillus, Mucor, Fusarium, Saccharomyces, Candida etc.
Structura micovirusurilor. Cel mai bine studiat este virusul care infectează specia Penicillium chrysogenum, care se prezintă sub forma unor mici particule poliedrice, cu Ø de 33-41 nm. Genomul este reprezentat de o moleculă de ARN d.c.
La fungi, virusurile sunt, în general, latente, celulele se dezvoltă mai lent, dar nu lizează constant. Micovirusurile au fost evidenţiate şi în sporii fungilor.
Transmiterea micovirusurilor, în general, se face în urma plasmogamiei (heterocarioza).
2.3.8. Sistematica, nomenclatura şi identificarea virusurilor Prima clasificare a virusurilor poartă numele de sistemul LHT şi a fost
realizată în anul 1962 de către cercetătorii Lowoff, Horn şi Tournier. Sistemul oferă o primă clasificare ştiinţifică a virusurilor, bazată pe următoarele criterii:
1) natura materialului genetic; 2) tipul de simetrie a capsidei; 3) prezenţa sau absenţa învelişului extern; 4) dimensiunea virionului şi a capsidei.

39
În prezent, există două sisteme principale utilizate pentru clasificarea virusurilor:
1. Sistemul de clasificare ICTV . Centrul Internaţional de Taxonomie a Virusurilor (ICTV = International Comitee on Taxonomy of Viruses) a elaborat, la începutul anilor 1990, un sistem de clasificare şi nomenclatură a virusurilor. ICTV are sarcina de a actualiza şi de a menţine un sistem taxonomic universal.
Conform acestui sistem, la clasificarea virusurilor se ţine cont de următoarele aspecte : • proprietăţi morfologice (mărimea şi forma virionului, structura şi simetria
capsidei, etc.); • proprietăţi fizico-chimice şi fizice (greutatea moleculară, coeficientul de
sedimentare); • genomul (tipul acidului nucleic, monocatenar sau bicatenar); • organizarea şi replicarea genomului; • proprietăţile antigenice; • proprietăţi biologice (spectrul de gazde, modul de transmitere în natură,
relaţiile cu vectorii). Unităţile taxonomice pentru virusuri încep la nivel de ordin şi fiecare
taxon prezintă sufixul, scris cursiv între paranteze. • Ordin (-virales) • Familie (-viridae) • Subfamilie (-virinae) • Gen (-virus) • Specie
Conform datelor publicate în 2010 de Centrul Internaţional de Taxonomie a Virusurilor, în schema taxonomică sunt cuprinse: Sase ordine, 87 familii, 19 subfamilii, 348 genuri şi 2285 specii de virusuri. Cele şase ordine stabilite de ICTV sunt: Caudovirales, Herpesvirales, Mononegavirales, Nidovirales, Picornavirales şi Tymovirales.
2. Sistemul de clasificare Baltimore. Clasificarea Baltimore, iniţiată de
biologul american David Baltimore (1971), separă virusurile în şapte grupe. Are la bază împărţirea în funcţie de tipul genomului viral, sens, precum şi

40
metoda de replicare. Aceasta clasificare este deseori preferată datorită uşurinţei de identificare a diferitelor familii de virusuri: • Tipul I: Virusuri cu genom ADN, dublucatenar: Fam. Herpesviridae, Fam. Poxviridae, Fam. Adenoviridae şi Fam. Papovaviridae • Tipul II: Virusuri cu genom ADN monocatenar. Familiile: Circoviridae şi Parvoviridae • Tipul III: Virusuri cu genom ARN dublucatenar. Familiile: Reoviridae şi Birnaviridae • Tipul IV: Virusuri cu genom ARN(+), monocatenar. Familiile: Astroviridae, Caliciiridae, Coronaviridae, Flaviviridae, Picornaviridae, Arteriviridae şi Togaviridae • Tipul V: Virusuri cu genom ARN monocatenar (-). Familiile: Orthomyxoviridae, Arenaviridae, Paramyxoviridae, Bunyaviridae şi Rhabdoviridae. • Tipul VI: Virusuri diploide cu genom ARN monocatenar: Fam. Retroviridae. • Tipul VII: Virusuri cu genom ADN dublucatenar cu molecula de ARN monocatenar ca intermediar: Fam. Hepadnaviridae.
Pentru identificarea virusurilor se utilizează metode moderne, bazate pe studiul componentelor genomului viral, pe clonări şi hibridări nucleare şi enzimatice de tip ELISA sau PCR.
2.4. Bacteriile
Sunt organisme unicelulare, cu o structură complexă, care au un metobolism propriu, datorită unui aparat enzimatic complex, ce le asigură desfăşurarea proceselor vitale. În linii generale, bacteriile au o serie de caractere generale, ce pot fi sistematizate astfel: • Nucleul celulei bacteriene este de tip procariot, adică lipsit de membrană,
iar afinităţile tinctoriale nu sunt deosebite faţă de citoplasmă. • Bacteriile au un polimorfism accentuat, putând îmbrăca mai multe aspecte
morfologice, ca forme sferice, cilindrice, spiralate sau helicoidale, filamentoase şi pătrate.

41
• Mişcarea la speciile mobile este asigurată de organite speciale, numite cili sau flageli.
• Bacteriile sunt lipsite de clorofilă, dar unele grupe au pigmenţi sintetizanţi şi, ca atare, pot folosi energia luminoasă.
• Altele îşi procură energia necesară sintezelor celulare şi altor manifestări vitale dezintegrând diverse substanţe chimice prin intermediul proceselor fermentative, fiind, din acest punct de vedere, organisme chimiosintetizante (Chemotrofe).
• Se înmulţesc prin sciziparitate, dar se cunosc la unele grupe şi forme de conjugare de tip parasexuat, cu formare de celule sexuate de sens diferit; forma de sciziparitate rămâne însă forma esenţială de reproducere.
• Bacteriile pot forma spori, ce constituie un mijloc de conservare a speciei, la unele grupe.
2.4.1. Morfologia bacteriilor
Forma exterioară a bacteriilor (Fig. 28) este un caracter controlat
genetic, iar din acest punct de vedere se disting cinci tipuri de bază, după cum urmează: • bacterii sferice, denumite coci, cu genul principal Coccus; • bacterii cilindrice, denumite bacili, sub formă de bastonaş drept sau uşor curbat, din care face parte genul Bacillus; • bacterii spiralate sau elicoidale, în care se cuprind trei categorii de germeni şi anume: vibrion, spiril şi spirocheta; • bacterii filamentoase, cu celule alungite, reprezentate de actinomicete; • bacterii pătrate, întâlnite în apele hipersaline.

42
După forma celulelor şi modul de grupare în urma procesului de
diviziune, bacteriile se împart în următoarele tipuri şi subtipuri morfologice: 1. Bacteriile sferice sau cocii (gr. kokkos - bob) au formă sferică,
ovoidală, elipsoidală sau reniformă, cu cele două diametre aproximativ egale. În funcţie de poziţia celulelor fiice după diviziune, cocii prezintă următoarele moduri de grupare (Fig. 29-34): • Cocul simplu, izolat, la care celulele rămân independente după diviziune (ex. Micrococcus ureae).
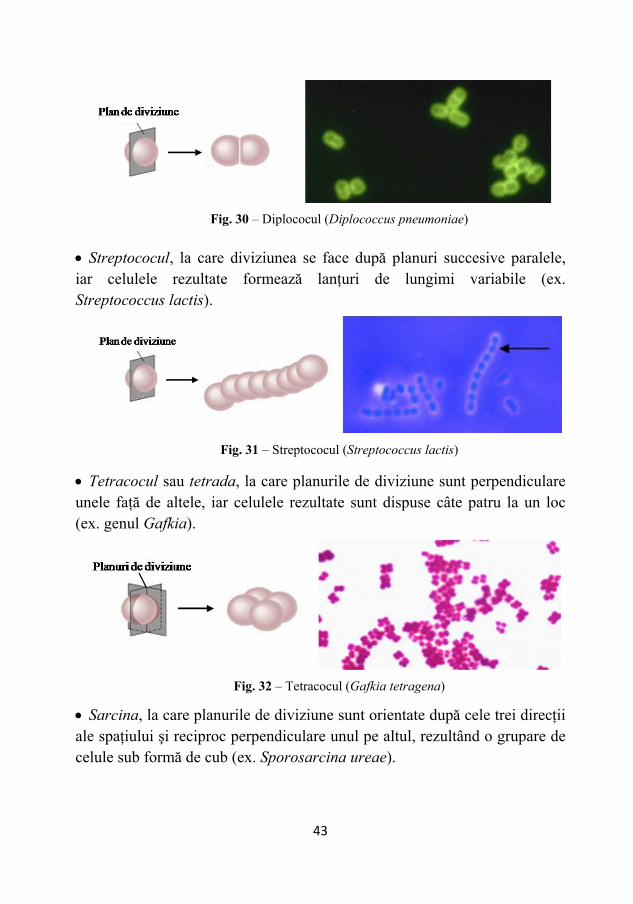
• Diplococul, la care diviziunea se face după planuri succesive paralele, celulele rezultate rămânând grupate câte două (ex. Diplococcus pneumoniae).
Fig. 29 – Cocul simplu (Micrococcus ureae)
Fig. 28 – Tipuri morfologice de bacterii: (a) sferice (coci); (b) cilindrice
(bacili); (c) spiralate (elicoidale); (d) filamentoase; (e) pătrate.
43
• Streptococul, la care diviziunea se face după planuri succesive paralele, iar celulele rezultate formează lanţuri de lungimi variabile (ex. Streptococcus lactis).
• Tetracocul sau tetrada, la care planurile de diviziune sunt perpendiculare unele faţă de altele, iar celulele rezultate sunt dispuse câte patru la un loc (ex. genul Gafkia).
• Sarcina, la care planurile de diviziune sunt orientate după cele trei direcţii ale spaţiului şi reciproc perpendiculare unul pe altul, rezultând o grupare de celule sub formă de cub (ex. Sporosarcina ureae).
Fig. 32 – Tetracocul (Gafkia tetragena)
Fig. 31 – Streptococul (Streptococcus lactis)
Fig. 30 – Diplococul (Diplococcus pneumoniae)

44
• Stafilococul, la care planurile succesive de diviziune sunt dispuse în mai multe direcţii, iar celulele rezultate se grupează ca un ciorchine (ex. Staphyllococcus aureus).
2. Bacteriile cilindrice, cunoscute sub denumirea de bacili (lat.
bacillus - bastonaş), au formă de bastonaşe, cu raportul între cele două axe foarte diferit, de la forme cu aspect filamentos, până la unele cu aspect aproape sferic (cocobacili, ex. Pasturella pestis). Bacilii sunt drepţi sau uşor curbaţi la mijloc sau la una din extremităţi, iar capetele lor pot fi tăiate drept (Bacillus anthracis) sau rotunjite (Escherichia coli). Marginile laterale ale celulei sunt, de obicei, paralele, dar pot fi şi apropiate la extremităţi, în formă de suveică (Fusiformis fusiformis), sau îndepărtate şi rotunjite la una sau ambele extremităţi, în formă de măciucă sau de pişcot (Corynebacterium sp.). Unele dintre bacteriile cilindrice au proprietatea de a forma spori.
Bacilii pot fi (Fig. 35): • izolaţi • grupaţi câte doi (diplobacili), în formă de V, X, Y (Mycobacterium sp.); • în lanţuri, cu lungimi variate (streptobacili);
Fig. 34 – Stafilococul (Staphyllococcus aureus)
Fig. 33 – Sarcina (Sarcina lutea)

45
• în palisdă, ca scândurile unui gard, celulele ramânând apropiate şi paralele în sensul axului lung, aşezarea fiind rezultatul unei mişcări de basculare a celulei-fiice, având ca punct de sprijin peretele transvers recent separat; • grupaţi în formă de rozetă sau de stea (Agrobacterium stellulatum, Ag. radiobacter, Phyllobacterium stappi).
3. Bacteriile spiralate (elicoidale) cuprind trei subtipuri morfologice
(Fig. 36): • Vibrionul, în formă de virgulă, cu un singur tur de spiră (ex. Vibrio cholerae); • Spirilul, în formă de spirală cu mai multe ture de spiră, rigide ca un baston (ex. Spirillum voluntas); • Spirocheta, în formă de spirală cu mai multe ture flexibile, care se poate strânge şi relaxa ( ex. Borrelia sp., Treponema sp. şi Leptospira sp.).
Fig. 35 – Bacterii cilindrice: (a) cocobacil; (b) bacil izolat; (c) diplobacil;
(d) streptobacil; (e) bacili grupaţi în palisadă.

46
4. Bacteriile filamentoase pot fi neramificate (Ord. Caryophanales),
cu ramificaţii false (Ord. Chlamydobacteriales) sau cu ramificaţii adevărate (Ord. Actinomycetales).
Sunt microorganisme cu asemănări morfologice cu fungii, ce au particularitatea de a forma hife, cu tendinţă de ramificare, de unde şi aspectul lor de miceliu.
În unele cazuri, spre exemplu la Sphaerotilus nutans (Fig. 37), aspectul filamentos este determinat de aşezarea celulelor individuale în lanţuri de celule, reunite printr-o teacă delicată cu perete neted.
5. Bacteriile pătrate, evidenţiate în apele hipersaline din peninsula
Sinai, au forma unor pătrate cu latura de 1,5-11 μm şi o grosime inegală (0,1 µm sau chiar mai mică în regiunea centrală şi 0,2-0,5 µm la periferie).
Multiplicarea se face prin diviziune directă (ex. Quadra sp.). În unele cazuri, fiecare bacterie pătrată creşte până ia formă unui dreptunghi, care se divide în două pătrate egale. Alteori, diviziunea se face în două planuri, ce alterneză în unghi drept, iar celulele rezultate din diviziune formează placarde de 8-16 celule (Fig. 38).
Fig. 36 – Bacterii spiralate: (a) vibrion; (b) spiril; (c) spirochetă.
Fig. 37 – Sphaerotilus nutans
Fig. 38 – Bacterii pătrate

47
În afara acestor cinci tipuri morfologice de bază există bacterii cu forme particulare: • Bacterii stelate - bacterii sub formă de stea; se înmulţesc prin sciziparitate (Stella sp.); • Bacterii care formează trichoame - grupare de celule care în urma diviziunii se prezintă sub forma unui filament multicelular, în care celulele adiacente au o suprafaţă relativ mare de contact şi sunt menţinute într-un înveliş parietal comun (Beggiatoa sp., Caryophanon sp., Sphaerotilus sp.); • Bacteriile prostecate (gr. prosteka – adaos) prezintă o complicaţie morfologică sub forma unui apendice semirigid, situat în continuarea celulei procariote (Caulobacter sp). Diametrul acestui apendice este mai mic decât al celulei bacteriene mature; • Bacteriile cu apendice acelulare prezintă un aspect filamentos, datorită apendicelor acelulare (substanţe secretate sau excretate), nedelimitate de peretele celular (Gallionella sp.).
Dimensiunile bacteriilor se exprimă în micrometri (1 μm = 10-6 m) şi, în medie, au 0,5-1 x 3-6 μm. Cele mai mici bacterii aparţin genului Mycoplasma (diametrul = 125-250 nm), iar cele mai mari pot ajunge la 10-150 μm lungime (Beggiatoa sp.). În cazul formelor filamentoase, dimensiunile bacteriilor ajung în mod excepţional la 500 μm lungime (Saprospira grandis).
Sub raportul dimensiunilor, cele mai mici bacterii se suprapun virusurilor mari (Poxvirus), vizibile la microscopul fotonic, iar cele mai mari depăşesc mãrimea celor mai mici protiste eucariote.
2.4.2. Structura celulei bacteriene
Celula bacteriană este o entitate morfologică şi funcţională echivalentă
cu celulele organismelor superioare, cu o structură complexă, formată din următoarele componente, de la exterior către interior, luând ca reper peretele celular: • Structura extraparietală, formată din: capsulă; cilii sau flagelii; aparatul fimbrial şi pilii de sex; • Peretele celular;

48
• Structura intraparietală, compusă din: membrana citoplasmatică, mezozomii, citoplasmă, aparatul nuclear, incluziunile, ribozomii, vacuolele, sporul bacterian. I. Structura extraparietală:
1. Capsula (Fig. 39) este o secreţie, care apare la exteriorul unor celule bacteriene, şi constituie un înveliş în jurul acestora. În funcţie de raportul faţă de celula bacteriană, capsula este de mai multe feluri: • microcapsula este un strat mucoid foarte fin, cu grosimea de până la 0,2 μm şi care poate fi pus în evidenţă prin metode imunologice; • capsula propriu-zisă are grosimea cuprinsă între 0,2-2 μm; • stratul mucos este caracterizat prin prezenţa unei mase amorfe, neorganizate în jurul celulei; • zooglea se prezintă ca un strat mucos, neorganizat, care înglobează mai multe celule microbiene.
Compoziţia chimică a capsulei diferă în funcţie de specia bacteriană la care apare şi de condiţiile de mediu, fiind formată din 98 % apă şi alţi constituenţi ca polizaharidele şi polipeptide.
Funcţiile biologice ale capsulei sunt următoarele: - funcţia de protecţie împotriva fagocitelor la bacteriile patogene; - funcţia de virulenţă; - funcţia de protecţie împotriva desicaţiei.
2. Cilii sau flagelii sunt formaţiuni filamentoase, cilindrice, lungi, subţiri, situate la suprafaţa celulei bacteriene şi reprezintă organite de mişcare ale acesteia. Prezenţa cililor este caracteristică numai speciilor de bacterii mobile, iar numărul acestora este variabil funcţie de specie, de la unu până la o sută.
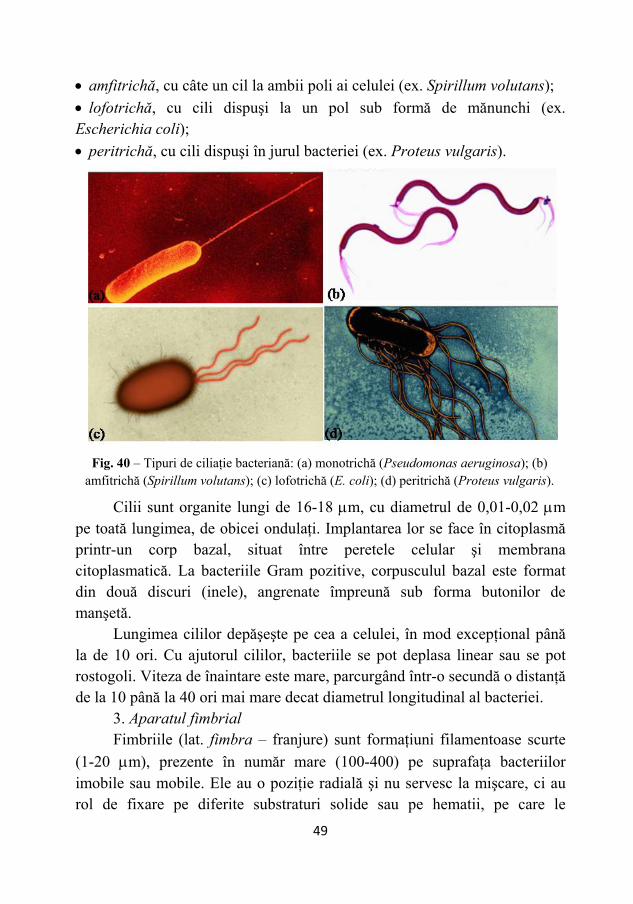
După modalitatea de inserţie pe corpul bacteriei, cilii pot avea următoarele poziţii (Fig. 40): • monotrihă, un cil la un singur pol (ex. Pseudomonas aeruginosa);
Fig. 39 – Capsula bacteriană
49
• amfitrichă, cu câte un cil la ambii poli ai celulei (ex. Spirillum volutans); • lofotrichă, cu cili dispuşi la un pol sub formă de mănunchi (ex. Escherichia coli); • peritrichă, cu cili dispuşi în jurul bacteriei (ex. Proteus vulgaris).
Cilii sunt organite lungi de 16-18 μm, cu diametrul de 0,01-0,02 μm
pe toată lungimea, de obicei ondulaţi. Implantarea lor se face în citoplasmă printr-un corp bazal, situat între peretele celular şi membrana citoplasmatică. La bacteriile Gram pozitive, corpusculul bazal este format din două discuri (inele), angrenate împreună sub forma butonilor de manşetă.
Lungimea cililor depăşeşte pe cea a celulei, în mod excepţional până la de 10 ori. Cu ajutorul cililor, bacteriile se pot deplasa linear sau se pot rostogoli. Viteza de înaintare este mare, parcurgând într-o secundă o distanţă de la 10 până la 40 ori mai mare decat diametrul longitudinal al bacteriei.
3. Aparatul fimbrial Fimbriile (lat. fimbra – franjure) sunt formaţiuni filamentoase scurte
(1-20 μm), prezente în număr mare (100-400) pe suprafaţa bacteriilor imobile sau mobile. Ele au o poziţie radială şi nu servesc la mişcare, ci au rol de fixare pe diferite substraturi solide sau pe hematii, pe care le
Fig. 40 – Tipuri de ciliaţie bacteriană: (a) monotrichă (Pseudomonas aeruginosa); (b)
amfitrichă (Spirillum volutans); (c) lofotrichă (E. coli); (d) peritrichă (Proteus vulgaris).

50
aglutinează. Fimbriile (Fig. 41) sunt de natură intracelulară, deoarece, după îndepărtarea peretelui celular, rămân fixate pe protoplaşti.
4. Pilii (Fig. 42) sunt apendici filamentoşi, tubulari, flexibili, care au
un canal prin care se face transferul cromozomului bacterian de la celula masculă (F+) la celula femelă (F-). Mai sunt denumiţi şi "pili de sex", iar procesul de transfer se numeşte conjugare bacteriană.
5. Spinii sunt formaţiuni extraparietale tubulare şi rigide, prezente la
suprafaţa unor bacterii Gram negative. Numărul lor variază între 1 şi 15 şi au o distribuţie întâmplătoare, perpendicular pe suprafaţa celulară. Prezintă rol taxonomic.
II. Peretele celular este invizibil sau foarte greu vizibil la microscopul
fotonic şi reprezintă aproximativ 20% din greutatea uscată a celulei şi 25% din volumul ei.
Peretele celular are rol: • de susţinere mecanică; • asigură individualitatea morfologică;
Fig. 42 – Pili de sex la E. coli
Fig. 41 – Aparat fimbrial

51
• oferă protecţie faţă de şocul osmotic; • participă la procesul de diviziune celulară; • conţine receptori de virus; • mediază schimbul de substanţe cu mediu.
Peretele celular este absent la genul Mycoplasma. Peretele celular (Fig. 43) reprezintă o structură macromoleculară,
formată dintr-un mucopeptid numit peptidoglican (mureină) şi substanţe care formează un matrix, în care este înglobată structura parietală bazală şi prezintă o compoziţie chimică particulară, în raport cu grupa de bacterii analizată.
Coloraţia Gram are o importanţă deosebită în taxonomia bacteriană şi a fost decoperită în 1844 de Hans Christian Gram. Această metodă porneşte de la observaţia că unele bacterii, colorate cu derivaţi bazici din grupul trifenil metanului (violetul de genţiana, violetul de metil, cristal violet) şi mordansate cu iod, rezistă la decolorarea cu anumiţi solvenţi organici (alcool sau acetonă), în timp ce altele nu rezistă. De aici, necesitatea recolorării frotiului cu o culoare de contrast (fucsină, safranină), care să recoloreze bacteriile, care nu au reţinut primul colorant.
Coloraţia Gram permite clasificarea bacteriilor în funcţie de structura şi compoziţia chimică a peretelelui celular în două mari categorii: Gram pozitive (G+, violet) şi Gram negative (G-, roşu). Între cele două categorii există diferenţe mari de compoziţie chimică şi anume: • Peptidoglicanul (mureina) este prezent la majoritatea bacteriilor, fiind de mai multe tipuri moleculare. Reprezintă 60-90% din greutatea uscată a peretelui celular la bacteriile Gram pozitive, în timp ce la bacteriile Gram negative reprezintă 10-20% şi se află în zona mediană a peretelui. • Acizii teichoici (gr. teichos - perete) sunt foarte abundenţi la bacteriile Gram pozitive şi lipsesc la bacterille Gram negative. Aceşti acizi sunt legaţi de straturile de peptidoglican sau de membrana citoplasmatică. • Acizii lipoteichoici sunt întâlniţi la bacteriile Gram pozitive, asociaţi la membrana citoplasmatică, dar se găsesc şi în peretele celular. • Acizii teichuronici sunt caracteristici bacteriilor Gram pozitive. • Lipopolizaharidele caracterizează bacteriile Gram negative şi sunt responsabile de primul control al permeabilităţii celulare. Bacteriile Gram

52
negative sunt, în general, mai rezistente la inhibitori (peniciline, coloranţi) decât cele Gram pozitive. • Lipoproteinele sunt întâlnite la bacteriile Gram negative sub forma unui strat lax, format din grupări lipidice şi proteice. Sunt situate pe suprafaţa superioară a peptidoglicanului. • Fosfolipidele sunt tipice pentru bacteriile Gram negative. • Acizii graşi se găsesc sub forma unor lanţuri lungi la nivelul peretelui celular la actinomicete şi la bacteriile coryneforme.
III. Structura intraparietală: 1. Membrana citoplasmatică, acoperă citoplasma bacteriilor şi o
separă de faţa internă a peretelui celular (Fig. 44). Are o grosime de 8-10 nm, este constituită din două straturi fosfolipidice şi reprezintă 10-20% din greutatea uscată a celulei.
Analiza chimică a membranei evidenţiază trei tipuri de molecule: lipide (fosfolipide), proteine şi glucide (polizaharide legate cu proteine – glicoproteine sau lipide - glicolipide). Membrana citoplasmatică este o barieră osmotică de permeabilitate, care reglează pătrunderea în celulă şi eliminarea selectivă a diferitelor substanţe.
Ea menţine în celulă o concentraţie ridicată de macromolecule, molecule mici şi chiar ioni, împedicându-le difuzarea în mediu, deşi concentraţia extracelulară este mai mică. Conţine permeazele care asigură
Membrană plasmatică
PeptidoglicanAcid teichoic
Acid lipoteichoic
Bacterie Gram pozitivă
Proteină
Fosfolipide
Proteină
Lipoproteine
Proteină
Porine
Membrană externă
Lipopolizaharide
Bacterie Gram negativă
Peptidoglican
Spaţiu periplasmicMembrană plasmatică
Proteină
(a) (b)
Fig. 43 – Structura peretelui celular la bacterii: (a) Gram pozitive; (b) Gram negative.

53
transportul activ în interiorul celulei a unor substanţe organice polare din mediu.
Membrana citoplasmatică are rol în creşterea şi diviziunea celulară; la nivelul său ia naştere semnalul care declanşează iniţierea replicării cromozomului bacterian.
În sfârşit, membrana citoplasmatică este suportul enzimelor care participă la sinteza ATP (la eucariote aceste enzime se află în mitocondrii).
2. Mezozomii sunt formaţiuni care derivă din membrana citoplasmatică sub forma unor invaginări, legate de ADN celular. Aceste organite celulare joacă un rol important în diviziunea nucleului şi în formarea septului, care separă cele două celule fiice.
La bacteriile purpurii, mezozomii conţin pigmenţi clorofilieni. La bacteriile fixatoare de azot, nitrogenaza, care este inhibată de O2, este protejată de mezozomi. Bacteriile nitrificatoare prezintă numeroase invaginări ale membranei citoplasmatice care măresc suprafaţa de schimb enzimatic.
3. Citoplasma este un sistem coloidal, constituit din săruri minerale, compuşi solubili de natură lipoproteică, nucleoproteine, lipide şi apă. Are un pH cuprins între 7 şi 7,2.
Citoplasma poate fi caracterizată ca un complex de stări fizice într-o continuă transformare, în funcţie de starea fiziologică şi vârsta celulei.
4. Ribozomii sunt formaţiuni citoplasmatice, sferice. O celulă bacteriană conţine, în medie, 18.000 ribozomi de tip 70 S, cu un diametru de 10-30 nm. Fiecare ribozom se disociază în două subunităţi, 30 S şi 50 S (Fig. 45). Fiecare subunitate 50 S este legată de
Strat fosfolipidic
Canalicul
Glicolipde
Proteină
Glicoproteine
Fig. 44 – Secţiune prin membrană
+50S 30S 70S
Fig. 45 – Structura ribozomilor bacterieni

54
o subunitate 30 S prin intermediul legăturilor ARN - proteină şi proteină-proteină. Ribozomii 70 S joacă un rol precis în cursul traducerii lanţurilor de ARN în proteine.
Conţin 50% apă, iar substanţa lor uscată se compune din circa 60% ARN (ARN ribozomal) şi circa 30% proteine. O parte din ribozomi sunt liberi în citoplasmă, iar ceilalţi sunt fixaţi de suprafaţa internă a membranei citoplasmatice.
5. Materialul nuclear ("nucleul") este constituit dintr-un cromozom format dintr-o buclă de ADN, aflat suspendat în citoplasmă. Lungimea acestuia este mare, ajungând la Escherichia coli la 1 mm, ceea ce implică o răsucire foarte accentuată. Din punct de vedere chimic, materialul nuclear conţine 60% ADN, 30% ARN şi 10% proteine.
6. Plasmidele sunt considerate material genetic extracromozomial, fiind independente de nucleu.
Plasmidele nu sunt indispensabile pentru viaţa celulară, dar pot aduce un avantaj selectiv. Ele poartă informaţia pentru degradarea unor substraturi şi pentru rezistenţa la antibiotice.
Replicarea plasmidelor se produce în două momente diferite: atunci când celula se divide şi atunci când se produce procesul de conjugare.
Bacteriile pot conţine mai multe plasmide. Escherichia coli, de exemplu, posedă pe cromozom unul sau două plasmide conjugate şi 10-15 plasmide neconjugate.
7. Vacuolele sunt formaţiuni sferice, înconjurate la periferie de o membrană lipoproteică, numită tonoplast, şi care apar în citoplasmă în faza de creştere activă a celulelor bacteriene. Numărul lor variază între 6 şi 20, iar în interiorul lor se găseşte apă sau gaz (cianobacterii, bacterii fotosintetizante roşii şi verzi etc.). Cele cu apă au rol în menţinerea presiunii osmotice în raport cu mediul extern şi în depozitarea substanţelor de rezervă, în timp ce vacuolele cu gaz au rol de flotaţie sau rol protector (faţă de intensitatea luminoasă).
8. Incluziunile sunt formaţiuni inerte, care apar în citoplasmă la sfârşitul periodei exponenţiale de creştere a celulei. Ele pot fi formate din glicogen, amidon, carbonat de calciu şi fosfat anorganic.
9. Sporul bacterian este o formaţiune care derivă din celula vegetativă a bacteriilor, în anumite condiţii de viaţă. Sporul este o formă de conservare

55
a speciei la condiţiile nefavorabile de mediu şi concentrează, într-un volum redus, toate componentele esenţiale ale celulei din care provine. Bacteriile sporogene, cu excepţia genului Desulfotomaculum (Gram variabil), sunt Gram pozitive, iar forma sporului poate fi sferică sau ovală.
Sporul apare ca o structură constantă la bacteriile anaerobe, aparţinând genului Clostridium, în mod facultativ la bacteriile aerobe din genul Bacillus şi foarte rar la coci (Sporosarcina ureae).
Dispunerea sporului în celula vegetativă poate fi centrală (Bacillus anthracis), subterminală (Bacillus cereus), terminală (Clostridium tetani) sau laterală (Fig. 46).
Dimensiunile sporilor variază între 0,2 şi 2 µm, iar diametrul acestora
poate fi mai mic (spor nedeformant sau bacteridie) sau mai mare (spor deformant sau clostridie) decât diametrul transversal al celulei în care se formează.
După modul de formare, structură şi rezistenţă la factorii de mediu, sporul bacterian poate fi de mai multe tipuri: • Endosporul (sporul propriu-zis) apare în interiorul unei celule vegetative, numită "sporangiu", şi prezintă o mare rezistenţă la condiţiile nefavorabile de mediu, în special la variaţiile temperaturii. O celulă vegetativă poate forma un singur endospor. Bacteriile susceptibile de a produce endospori aparţin genurilor: Bacillus, Clostridium, Desulfatomaculum, Sporosarcina, Sporolactobacillus, Thermoactinomyces. • Artrosporii se formează prin fragmentarea unor celule vegetative. Au o formă neregulată şi o rezistenţă intermediară între cea a endosporului şi celula vegetativă. Artrosporii sunt caracteristici bacteriilor actinomicete. • Chiştii sunt stadii de repaus sau supravieţuire, formate ca răspuns la modificările de mediu şi provin din transformarea unei celule vegetative, prin îngroşarea pereţilor şi acumularea de material de rezervă. Au o
(a) (b) (c) (d)
Fig. 46 – Localizarea sporului bacterian: (a) central; (b) subterminal; (c) terminal; (d) lateral.

56
rezistenţă mai mică decât endosporul şi nu sunt termorezistenţi. (ex. Azotobacter sp.) • Gonidia apare endocelular prin condensarea şi fragmentarea protoplasmei unei celule vegetative, numită gonidangiu. În interiorul unei celule apar, de regulă, mai multe gonidii, care sunt eliberate în mediu prin ruperea peretelui gonidangiului. Principala lor funcţie priveşte capacitatea de reproducere, neavând rezistenţa caracteristică sporului (ex. Leptothrix ochracea).
Compoziţia chimică a sporului. Între forma vegetativă şi forma sporulată a unei specii bacteriene există deosebiri de ordin calitativ şi cantitativ. Astfel, din punct de vedere cantitativ, sporii sunt bogaţi în Ca2+, Cu2+ şi Mg2+ şi săraci în P şi în substanţe proteice. De asemenea, sporii au un conţinut mai scăzut în H2O şi ARN, comparativ cu formele vegetative. În plus, sporii mai conţin, în cortexul sporului, acidul dipicolinic sub formă de dipicolinat de Ca.
Compoziţia chimică a sporului duce, implicit, la unele deosebiri în legătură cu rezistenţa sa faţă de agenţii fizici şi chimici.
Semnificaţia biologică a sporului Sporul bacterian, prin rezistenţa sporită faţă de factorii de mediu, are
rolul de conservare a speciei. El concentrează într-un volum redus toate componentele esenţiale ale celulei, sporogeneza făcând parte dintr-un ciclu vital obligatoriu.
Însuşirile de rezistenţă ale sporului pot fi sintetizate astfel: rezistenţa la temperatură, rezistenţa la uscăciune, rezistenţa faţă de factorii chimici. Metabolismul sporului este mult mai redus ca al celulei vegetative şi nu permite multiplicarea.
Citologia formării sporului În interiorul celulei bacteriene, formarea sporului presupune crearea
unei noi celule diferite de cea vegetativă, care se produce în mai multe faze (Fig. 47): • Stadiul preparator – viitorul nucleu al sporului ia naştere prin detaşarea unui segment din cromozomul bacterian, iar în jurul acestuia, ulterior, are loc condensarea citoplasmei; • Stadiul depresor – în care nucleul sporului, format şi înconjurat cu un strat de citoplasmă condensată, se separă de restul citoplasmei prin septul sporal; • Stadiul apariţiei învelişurilor sporale, tunica şi cortexul sporului;
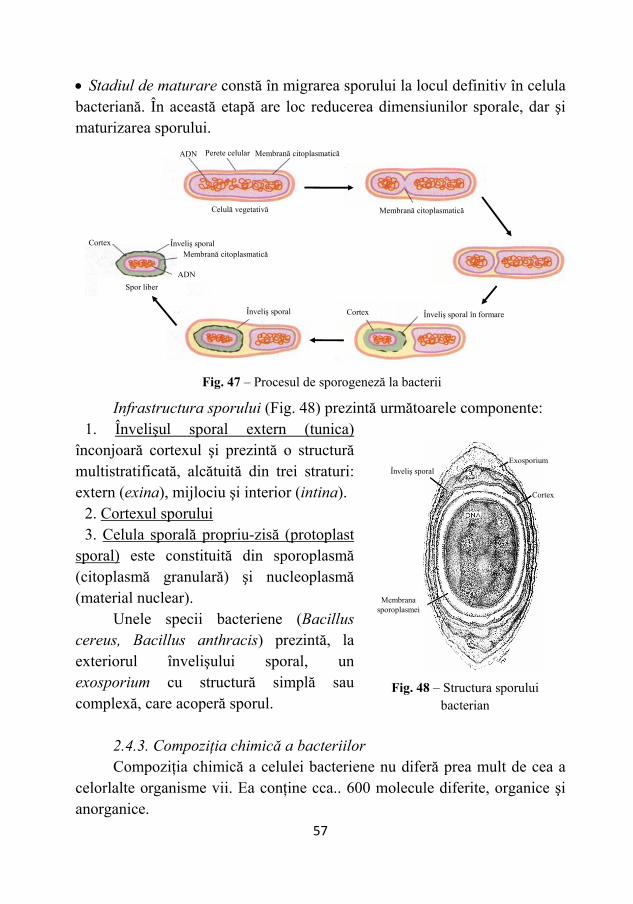
57
• Stadiul de maturare constă în migrarea sporului la locul definitiv în celula bacteriană. În această etapă are loc reducerea dimensiunilor sporale, dar şi maturizarea sporului.
Infrastructura sporului (Fig. 48) prezintă următoarele componente:
1. Învelişul sporal extern (tunica) înconjoară cortexul şi prezintă o structură multistratificată, alcătuită din trei straturi: extern (exina), mijlociu şi interior (intina).
2. Cortexul sporului 3. Celula sporală propriu-zisă (protoplast
sporal) este constituită din sporoplasmă (citoplasmă granulară) şi nucleoplasmă (material nuclear).
Unele specii bacteriene (Bacillus cereus, Bacillus anthracis) prezintă, la exteriorul învelişului sporal, un exosporium cu structură simplă sau complexă, care acoperă sporul.
2.4.3. Compoziţia chimică a bacteriilor Compoziţia chimică a celulei bacteriene nu diferă prea mult de cea a
celorlalte organisme vii. Ea conţine cca.. 600 molecule diferite, organice şi anorganice.
ADN Perete celular Membrană citoplasmatică
Celulă vegetativă Membrană citoplasmatică
Membrană citoplasmatică
ADNSpor liber
Cortex Înveliş sporal
Înveliş sporal Înveliş sporal în formareCortex
Fig. 47 – Procesul de sporogeneză la bacterii
Membrana sporoplasmei
Exosporium
Cortex
Înveliş sporal
Fig. 48 – Structura sporului
bacterian

58
Apa este mediul biogen esenţial şi reprezintă 78 % din greutatea medie a celulei bacteriene. Se găseşte sub formă de apă liberă sau legată de diferite componente celulare. Apa îndeplineşte numeroase funcţii: • este solvent al compuşilor hidrosolubili; • este mediu de dispersie pentru constituenţii chimici insolubili în apă; • condiţionează activitatea enzimelor; • asigură transportul elementelor de metabolism.
Sărurile minerale reprezintă 2-20 % din greutatea uscată a bacteriilor şi sunt reprezentate de principalele elemente. Cel mai bine este reprezentat fosforul (∼ 45 %). Substanţele minerale au rol în: • reglarea presiunii osmotice; • activează unele sisteme enzimatice; • reglează pH-ul; • influenţează permeabilitatea peretelui celular; • intră în compoziţia unor constituenţi celulari.
Acizii nucleici. ADN se găseşte sub forma unei singure molecule circulare, dublu helicale. Reprezintă 1/5 din conţinutul celular. ARN reprezintă cca. 17% din greutatea uscată a celulei bacteriene şi se găseşte localizat în citoplasmă sub trei forme: ARN mitocondrial (ARNm), ARN transfer (ARNt) şi ARN ribozomal (ARNr).
Proteinele reprezintă cca.. 60% din greutatea uscată a bacteriilor şi se găsesc sub formă de holoproteine şi heteroproteine. Proteinele au rol structural, iar unele îndeplinesc funcţia de enzime.
Glucidele reprezintă 4-25% din greutatea uscată a celulei bacteriene şi se găsesc sub formă de glucide simple (mono şi dizaharide) şi polizaharide.
Glucidele simple participă la metabolismul bacterian, iar polizaharidele au rol structural, energetic sau de material de rezervă.
Lipidele se găsesc în proporţie de 1-20% din greutatea uscată a bacteriilor, iar această variaţie este specifică, depinzând de vârsta celulei şi compoziţia mediului de cultură. Lipidele se găsesc în membrana citoplasmatică şi sub formă de granule intracitoplasmatice.
Pigmenţii sunt substanţe colorate care se găsesc în citoplasma bacteriilor cromogene. După localizarea pigmenţilor faţă de celulă, bacteriile se împart în două categorii:

59
• bacterii cromopare, la care pigmentul este eliminat în mediul pe care îl colorează (ex. Pseudomonas fluorescens); • bacterii cromofore, la care pigmentul rămâne localizat în celula bacteriană (ex. Staphylococcus aureus).
Pigmenţii pot fi de mai multe tipuri, după compoziţia chimică: bacterioclorofite, pigmenţi carotenoizi (roşii, galbeni, portocalii), antocianici, pigmenţi melanici (brun sau negru).
Pigmenţii pot avea rol în fotosinteză, rol antibiotic, de protecţie împotriva radiaţiilor luminoase şi ultraviolete, rol de vitamine.
Enzimele bacteriene constituie o grupă de proteine cu importanţă biologică deosebită, de catalizatori ai celulei vii, fiind solubile în apă, termolabile, criolabile şi sensibile la acţiunea radiaţiei ultraviolete.
După substratul asupra căruia acţionează, enzimele se clasifică în: • exoenzime, care sunt eliberate în mediul înconjurător, acţionând asupra unui substrat din afara celulei; acestea sunt angrenate în procesele de catabolism; • Endoenzime, care sunt secretate intracelular şi participă la reacţiile de sinteză (anabolism).
După viteza de acţiune, enzimele pot fi de două categorii: • enzime constitutive, care sunt preexistente şi care acţionează rapid asupra unui substrat. Sunt sintetizate în tot cursul vieţii bacteriei; • enzime adaptive, care acţionează lent, deoarece celula trebuie să se adapteze la substrat, apariţia enzimelor fiind legată de mediu.
2.4.4. Creşterea şi multiplicarea bacteriilor
Ciclul de dezvoltare al bacteriilor se compune din două faze: creşterea
şi multiplicarea. Prin creştere se înţelege procesul de neoformare, în condiţii
favorabile, în protoplasma bacteriei, a substanţei de constituţie. Această creştere se continuă până la atingerea unei limite, când survine diviziunea celulară.
Prin multiplicare se înţelege sporirea numărului de indivizi, care se face prin diviziune celulară.
Diviziunea celulară poate fi:
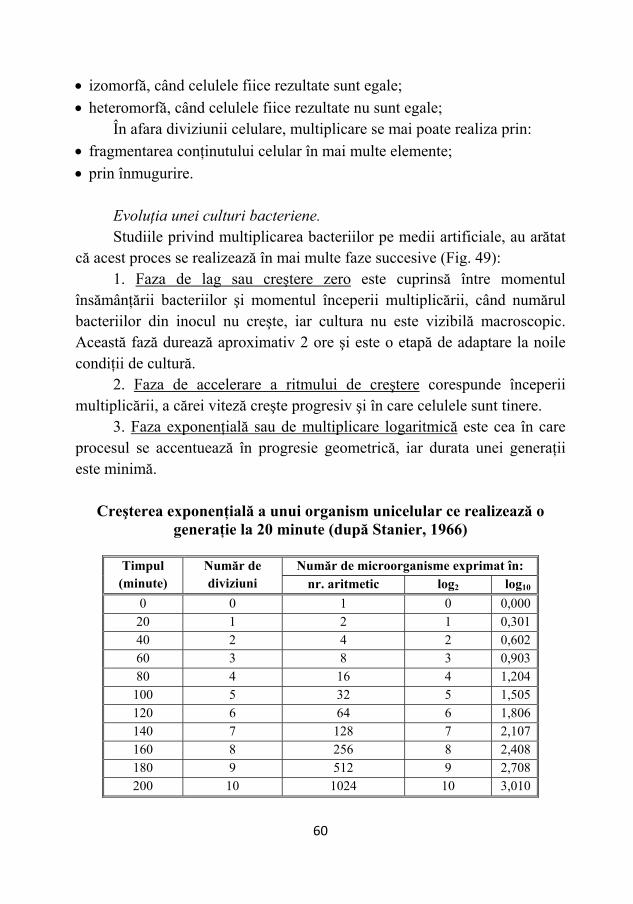
60
• izomorfă, când celulele fiice rezultate sunt egale; • heteromorfă, când celulele fiice rezultate nu sunt egale;
În afara diviziunii celulare, multiplicare se mai poate realiza prin: • fragmentarea conţinutului celular în mai multe elemente; • prin înmugurire.
Evoluţia unei culturi bacteriene. Studiile privind multiplicarea bacteriilor pe medii artificiale, au arătat
că acest proces se realizează în mai multe faze succesive (Fig. 49): 1. Faza de lag sau creştere zero este cuprinsă între momentul
însămânţării bacteriilor şi momentul începerii multiplicării, când numărul bacteriilor din inocul nu creşte, iar cultura nu este vizibilă macroscopic. Această fază durează aproximativ 2 ore şi este o etapă de adaptare la noile condiţii de cultură.
2. Faza de accelerare a ritmului de creştere corespunde începerii multiplicării, a cărei viteză creşte progresiv şi în care celulele sunt tinere.
3. Faza exponenţială sau de multiplicare logaritmică este cea în care procesul se accentuează în progresie geometrică, iar durata unei generaţii este minimă.
Creşterea exponenţială a unui organism unicelular ce realizează o
generaţie la 20 minute (după Stanier, 1966)
Număr de microorganisme exprimat în: Timpul (minute)
Număr de diviziuni nr. aritmetic log2 log10
0 0 1 0 0,000 20 1 2 1 0,301 40 2 4 2 0,602 60 3 8 3 0,903 80 4 16 4 1,204
100 5 32 5 1,505 120 6 64 6 1,806 140 7 128 7 2,107 160 8 256 8 2,408 180 9 512 9 2,708 200 10 1024 10 3,010
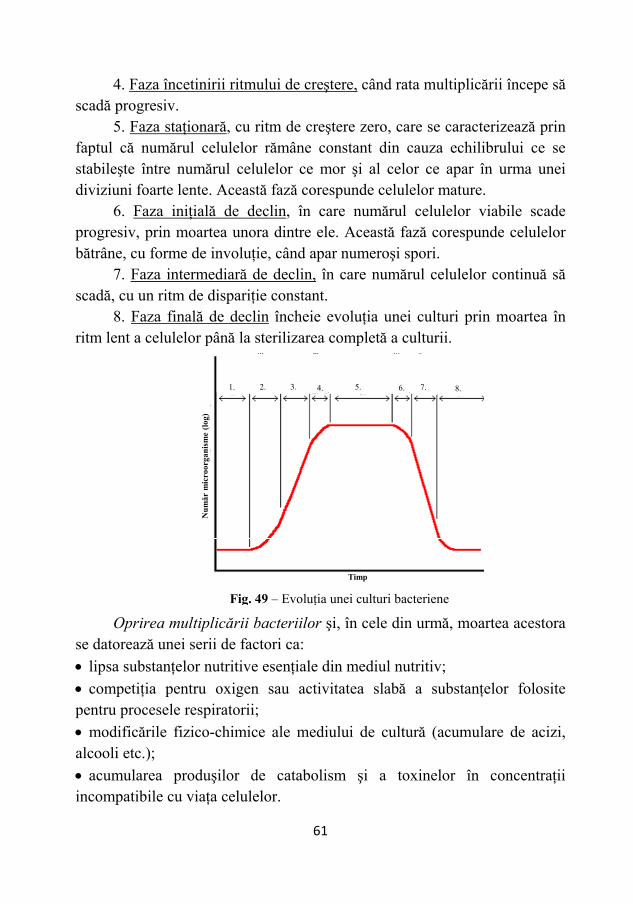
61
4. Faza încetinirii ritmului de creştere, când rata multiplicării începe să scadă progresiv.
5. Faza staţionară, cu ritm de creştere zero, care se caracterizează prin faptul că numărul celulelor rămâne constant din cauza echilibrului ce se stabileşte între numărul celulelor ce mor şi al celor ce apar în urma unei diviziuni foarte lente. Această fază corespunde celulelor mature.
6. Faza iniţială de declin, în care numărul celulelor viabile scade progresiv, prin moartea unora dintre ele. Această fază corespunde celulelor bătrâne, cu forme de involuţie, când apar numeroşi spori.
7. Faza intermediară de declin, în care numărul celulelor continuă să scadă, cu un ritm de dispariţie constant.
8. Faza finală de declin încheie evoluţia unei culturi prin moartea în ritm lent a celulelor până la sterilizarea completă a culturii.
Oprirea multiplicării bacteriilor şi, în cele din urmă, moartea acestora
se datorează unei serii de factori ca: • lipsa substanţelor nutritive esenţiale din mediul nutritiv; • competiţia pentru oxigen sau activitatea slabă a substanţelor folosite pentru procesele respiratorii; • modificările fizico-chimice ale mediului de cultură (acumulare de acizi, alcooli etc.); • acumularea produşilor de catabolism şi a toxinelor în concentraţii incompatibile cu viaţa celulelor.
1. 2. 3. 5.4. 7. 8.6.
Timp
Num
ăr m
icro
orga
nism
e (lo
g)
Fig. 49 – Evoluţia unei culturi bacteriene

62
2.4.5. Nutriţia bacteriilor Bacteriile pot folosi pentru manifestările lor vitale fie energia radiantă
luminoasă, în cazul celor capabile de fotosinteză, fie energia eliberată prin reacţii chimice oxidative, la întuneric, în cazul celor chimiotrofe.
După sursa de energie, bacteriile pot fi clasificate astfel: • fototrofe (fotosintetizante), a căror sursă primară de energie este lumina; • chemotrofe (chimiosintetizante), a căror sursă de energie este obţinută la întuneric, în urma reacţiei de oxido-reducere a unor substanţe organice sau anorganice. • paratrofe, la care creşterea este dependentă de energia furnizată de celula parazitată.
În funcţie de capacitatea de sinteză a metaboliţilor esenţiali, bacteriile prezintă următoarele tipuri de nutriţie: • Autotrofia corespunde capacităţii de sinteză a tuturor constituenţilor celulari prin mijloace proprii, pornind de la surse simple anorganice de carbon şi azot ca: CO2, NH3, NO2
-, NO3-.
• Heterotrofia implică incapacitatea de sinteză a unor metaboliţi esenţiali şi, de aceea, este nevoie de prezenţa substanţelor organice ca sursă de carbon sau azot. Bacteriile care folosesc o sursă organică de carbon sunt denumite bacterii de fermentaţie, iar cele care utilizează o sursă organică de azot mai sunt denumite bacterii de putrefacţie.
În concluzie, autotrofele, prin sintezele lor generează substanţa organică, plecând de la compuşi minerali simpli, pe când heterotrofele nu-şi pot realiza sintezele proprii, decât dacă dispun de substanţe organice, pe care le descompun până la produşi mai simpli şi pe care îi utilizează în metabolismul lor.
2.4.6. Respiraţia microorganismelor
După exigenţele faţă de oxigenul molecular, microorganismele se
grupează în patru tipuri respiratorii: 1. Microorganisme strict (obligatoriu) aerobe, care folosesc oxigenul
ca acceptor final de hidrogen, având nevoie de prezenţa continuă a oxigenului atmosferic.

63
2. Microorganisme strict (obligatoriu) anaerobe, care nu se pot dezvolta în prezenţa oxigenului molecular, putând fi cultivate numai pe medii sărăcite în oxigen. Ele nu dispun de enzimele respiratorii din categoria citocromilor.
Anaerobioza lor strictă constă în faptul că în prezenţa oxigenului, care se comportă ca acceptor final de hidrogen, se formează peroxidul de hidrogen (compus toxic) şi nu apă.
3. Microorganisme facultativ anaerobe, care sunt capabile să-şi orienteze metabolismul spre respiraţie sau spre fermentaţie, în funcţie de disponibilităţile de oxigen şi care au, în mod obişnuit, un metabolism anaerob, fără a fi însă sensibile la prezenţa oxigenului.
Unele microorganisme cu acest tip respirator (levurile) pot trece de la un metabolism aerob la unul anaerob, în funcţie de condiţiile de mediu.
4. Microorganismele microaerofile au nevoie de o concentraţie de oxigen mai mică decât cea din aerul atmosferic. Această particularitate se datorează sensibilităţii unora dintre enzimele lor la condiţiile de oxidare puternică.
2.4.7. Grupe particulare de bacterii 1. Bacterii glasogene Apa pură din punct de vedere chimic şi biologic, poate rămâne în
stare lichidă până la -40oC. Supusă la temperaturi inferioare pragului de 0oC, nu îngheaţă decât în prezenţa nucleelor glasogene. Aceste nuclee pot fi reprezentate de particulele minerale, care provoacă congelarea, începând de la -10oC. La temperaturi cuprinse între -10 şi 0oC, gheaţa nu se formează la plante decât în prezenţa bacteriilor specifice, ce posedă în membrana lor nuclee glasogene.
Suşele bacteriene în cauză aparţin speciilor Pseudomonas syringae (Fig. 50) şi Erwinia herbicola; ele pot fi saprofite (epifite) sau parazite pe organele plantelor în cauză (frunze, flori, fructe). Proporţia de nuclee glasogene este ridicat la Ps. syringae
Fig. 50 – Pseudomonas syringae

64
(60 %) şi mai mic la E. herbicola (1-5 %). Puterea glasogenă a bacteriilor se exprimă "in vitro" sau în "in vivo" pe organele plantelor. Pe de altă parte, suşele inductoare de gheaţă se multiplică abundent pe organele necrozate prin îngheţ, ceea ce le oferă un avantaj selectiv considerabil.
Când o suspensie de bacterii glasogene este pulverizată pe frunze apar simptomele îngheţului cu ofilirea ireversibilă, dacă sunt plasate (ţinute) o oră la -5oC, pe când plantele martor netratate cu bacterii rămân imune la toate alterările vizibile.
La viţa de vie s-a observat o corelaţie ridicată între numărul de bacterii glasogene prezente pe muguri şi pierderile cauzate de frig, la temperaturi apropiate de 0oC. Bacteriile glasogene, care provoacă îngheţarea, au fost observate la plante foarte diverse (pin, cireş, piersic, prun, cais, forsiţia, fasole, porumb, tomate, căpşun).
Suşele bacteriene active, datorită capacităţii de a induce formarea gheţii, prezintă o genă specifică, care codează o proteină foarte particulară în membrana lor. Proteina este formată din 1200 de aminocizi, cu o structură repetitivă foarte accentuată. Compus din 1200 de aminoacizi, acest polipeptid comportă repetiţii în mai mult de 100 exemplare. Reducerea numărului de repetiţii în lanţul polipeptidic reduce activitatea glasogenă.
Alte proteine cu structură repetitivă sunt prezente în sângele unor specii de animale din regiunile polare, unde au un efect antigel.
Bacteriile glasogene joacă un rol important în practică, deoarece provoacă îngheţarea în primăvară a numeroase plante, în special pomii fructiferi şi viţa de vie. Stoparea acţiunii negative a acestor bacterii se încearcă pe diferite căi: tratamente chimice cu antibiotice, pulverizări cu apă, fumigaţii, folosirea bacteriofagilor, a microorganismelor antagoniste, aplicarea de polimeri care limitează schimburile termice cu exteriorul etc.
În ultimii ani s-au obţinut prin inginerie genetică suşe neglasogene de Ps. syringae, care, aplicate pe plante, inhibă dezvoltarea speciilor glasogene.
În sens pozitiv, specia Ps. syringae glasogenă este utilizată pentru obţinerea zăpezii artificiale (Snow max).
2. Micoplasmele reprezintă un grup particular de bacterii lipsite de
perete celular, celula fiind delimitată de o membrană lipoproteică cu o compoziţie chimică asemănătoare celulei animale.

65
Sunt considerate cele mai mici organisme capabile de creştere autonomă pe medii acelulare.
Micoplasmele sunt grupate în clasa Mollicutes (moli - moale, pliabil şi cute - piele) cu două genuri: • Mycoplasma – creşterea speciilor din acest gen este dependentă de prezenţa sterolilor în mediu. • Acholeplasma cuprinde specii care se dezvoltă independent de steroli.
Morfologie. Datorită lipsei peretelui celular, micoplasmele au o plasticitate naturală foarte mare, fiind descrise mai multe tipuri morfologice ca: sferice-cocoidale, diplococi, filamentoase.
Cu toate acestea, cultivate în condiţii de mediu bine definite, fiecare specie are o morfologie caracteristică, astfel încât pleomorfismul acestora nu trebuie confundat cu polimorfismul, care implică, obligatoriu, existenţa a mai mult de o formă celulară distinctă în cursul vieţii unui organism.
Micoplasmele sunt Gram-negative, iar dimensiunile lor variază de la 125-250 nm la formele sferice, până la 1-2 μm la cele filamentoase.
Lipsa peretelui celular este răspunzătoare de instabilitatea morfologică şi de alte proprietăţi, cum ar fi: • sensibilitatea osmotică; • rezistenţa la antibioticele care acţionează asupra sintezei peretelui celular (penicilina); • tendinţa de pătrundere şi creştere în profunzimea mediilor; • capacitatea de a traversa filtrele bacteriene.
Multiplicarea micoplasmelor se poate realiza la aceeaşi specie prin diviziune, înmugurire şi fragmentarea filamentelor.
În condiţii naturale, micoplasmele sunt parazite, instalarea lor în diferite organisme fiind uşurată de natura chimică a membranei, similară membranelor gazdei. Micoplasmele produc peste 40 de boli la plante, dar şi la animale şi om.
3. Actinomicetele constituie un grup mare de bacterii filamentoase,
imobile, Gram pozitive, caracterizate printr-o mare varietate morfologică. Multă vreme, actinomicetele au fost considerate ca făcând parte din
categoria ciupercilor, deoarece au particularitatea de a forma hife (filamente) ramificate şi spori de diseminare.

66
Din punct de vedere taxonomic, actinomicetele sunt incluse în divizia Firmicutes, clasa Thallobacteria. În sol, se găsesc mai ales actinomicete ce aparţin familiior Frankiaceae, Nocardiaceae, Streptomycetaceae (cca..70% din total) şi Micromonosporaceae.
Morfologie. Majoritatea actinomicetelor au o formă alungită filamentoasă, cu tendinţă de ramificare, de unde şi aspectul lor de miceliu. Filamentele miceliene au un diametru cuprins între 0,5 şi 2 μm şi prezintă o structură de tip procariot, corespunzând unor celule foarte lungi, multinucleate, fără pereţi transversali.
Pe medii de cultură solide formează colonii dense cu aspect cartilaginos sau cretos, aderente la substrat. Pe aceste colonii se formează sporofori cu spori. Sporoforii sunt diferiţi ca formă: spiralaţi, drepţi sau curbaţi.
Sporii sunt, de asemenea, variaţi ca formă, fiind sferici ovali, alungiţi sau cilindrici. Suprafaţa sporilor este netedă sau prezintă diverse ornamentaţii.
Actinomicetele produc pigmenţi, care dau miceliului aerian culori diferite (galben, portocaliu, roşu, violet până la albastru sau cenuşiu). La unele specii, pigmenţii difuzează în mediu.
Metabolism. O particularitate a fiziologiei actinomicetelor este modul lor de nutriţie omnivor, care le permite să se dezvolte pe substraturi organice dintre cele mai diferite.
În condiţii naturale, pe resturi vegetale, actinomicetele încep să se dezvolte numai după ce bacteriile şi ciupercile au încetat să se mai multiplice, atunci când toate substanţele uşor asimilabile din resturile vegetale au fost descompuse de predecesorii lor.
Capacitatea lor de a degrada substanţele organice dintre cele mai complexe le conferă un rol important în natură, în special în descompunerea substanţelor organice din nămoluri şi sol. În sol, acestea participă la degradarea biologică şi mineralizarea substanţelor organice.
Fig. 51 – Streptomyces sp

67
Importanţa lor practică este legată, în mod deosebit, prin intermediul speciilor genului Streptomyces (Fig. 51), de producerea de substanţe antibiotice (sunt cunoscute peste 500 de antibiotice distincte).
Aproximativ 50 % din tulpinile izolate în natură produc antibiotice. Unele actinomicete produc boli la plante, animale şi om.
2.4.8. Sistematica, nomenclatura şi identificarea bacteriilor
La baza clasificǎrii bacteriilor stau o serie de caractere, care constitue criterii de încadrare a acestora în mai multe categorii taxonomice, având la bazǎ specia. În succesiunea lor, aceste categorii sunt: specia, genul, familia, ordinul, clasa şi diviziunea (încrengătura).
Rangurile taxonomice sunt indicate de câte un sufix: • Ordin (-ales) • Subordin (-ineae) • Familie (-aceae) • Subfamilie (-oidiae)
Unitatea de lucru efectivǎ este tulpina (suşa) bacterianǎ, care este o culturǎ purǎ, constituitǎ dintr-o singurǎ specie, având la origine o colonie, ce provine dintr-o singurǎ bacterie, care s-a multiplicat. Tulpina sau suşa se noteazǎ cu un simbol sau un numǎr şi se pǎstreazǎ în colecţii ştiinţifice.
Cercetǎrile ce stau la baza identificǎrii şi clasificǎrii bacteriilor sunt numeroase şi ţin cont de următoarele aspecte: • caractere morfologice: formă, dimensiune, cilii, capsula şi comportarea tinctorialǎ fatǎ de coloraţia Gram; • caractere culturale: creşterea pe medii de culturǎ lichide sau solide, forma şi tipul coloniei, pigmentaţia etc; • caracterele biochimice se referǎ la capacitatea de utilizare a surselor de carbon şi azot, tipul de respiraţie etc; • caractere antigenice (serologice); • caractere de patogenitate, privind virulenţa şi toxicitatea speciilor patogene; • sensibilitatea faţǎ de bacteriofagii specifici;

68
• posibilitatea de transfer genetic prin bacteriofag sau prin fenomene parasexuale; • raportul guaninǎ (G) + citozinǎ (C) sau raportul G/C % faţǎ de masa genomului. Speciile înrudite au un conţinut de G/C % aproape identic. • caractere ecologice: habitatul natural al microorganismului şi relaţiile lui cu mediul natural.
Dintre încercările de sistematizare a bacteriilor se amintesc cele realizate de Ehrenberg (1839), Lehman şi Neuman (1927), Pribram (1933), Kluyver şi Van Niel (1936), Prevot (1957, 1961), Krasilnicov (1959), Skerman (1967) şi Murray (1974).
Societatea bacteriologilor americani, pe baza lucrărilor lui Buchanan (1916-1918) şi Winslow (1920), a stabilit un sistem de clasificare, prezentat în 1923 de David-Hendricks Bergey în prima ediţie a „Bergey’s Manual of Determinative Bacteriology”. Acest determinator, adoptat de majoritatea microbiologilor, se află la a noua ediţie (1994) şi clasifică bacteriile în baza caracterelor fenotipice comune (caractere morfologice, de cultură, fiziologice, biochimice etc.) în patru divizii şi 35 de secţiuni:
Cel mai nou sistem de clasificare bacteriană a apărut în perioada 2001-
2011 şi este prezentat în cinci volume ale celei de-a doua ediţii din „Bergey’s Manual of Systematic Bacteriology”. Acest sistem clasifică bacteriile după particularităţile genotipice (gradul de omologie al secvenţelor nucleotidice ale ARN ribozomal, gradul de omologie al secvenţelor nucleotidice ale ADN, conţinutul relativ de guanină+citozină (GC%) al ADN purificat etc.).
Divizia Clasa
I. Gracilicutes – bacterii Gram negative
1. Scotobacteria - bacterii non-fotosintetice 2. Anoxyphotobacteria - bacterii fotosintetice anaerobe 3. Oxyphotobacteria - bacterii fotosintetice aerobe (cyanobacterii)
II. Firmicutes – bacterii Gram pozitive
1. Firmibacteria - coci şi bacili 2. Thallobacteria - bacterii filamentoase (actinomicete)
III. Tenericutes 1. Mollicutes - bacterii lipsite de perete celular (micoplasme)
IV. Mendosicutes 1. Archaebacteria – bacterii cu perete celular, lipsit de stratul peptidoglican

69
2.4.9. Rolul bacteriilor în natură Bacteriile care trăiesc în natură joacă un rol important în mineralizarea
compuşilor organici, contribuind, astfel, la realizarea circuitului elementelor în natură (azotul, fosforul, potasiul, calciul, magneziul şi microelementele).
Bacteriile contribuie şi la formarea substanţelor humice, îmbogăţind solul în materie organică.
Prin unele substanţe bacteriene de natură mucilaginoasă, cum sunt polizaharidele capsulare, acestea contribuie la agregarea particulelor de sol, deci la îmbunătăţirea structurii solului.
Unele specii bacteriene au capacitatea de a produce antibiotice, care pot limita dezvoltarea unor grupe de microorganisme din sol, deci menţin sau schimbă echilibrul biologic din sol.
Unele grupe de bacterii trăiesc în simbioză cu plantele superioare, producând nodozităţi şi realizează fixarea azotului atmosferic; altele activează în zona rădăcinii plantelor, contribuind la dezvoltarea acestora şi la sporirea activităţii rizosferice.
În medii specifice, unele grupe de bacterii produc procese fermentative utile, ca fermentaţia amidonului, a pectinelor, fermentaţia lactică, butirică, propionică etc., în timp ce altele au capacitatea de a secreta enzime extracelulare, cu care lizează unele bacterii sau ciuperci.
2.5. Cyanobacteriile (algele albastre-verzi)
Cyanobacteriile (gr. kyanós – albastru) sunt microorganisme procariote, cunoscute, iniţial, sub denumirea de alge verzi-albastre, care folosesc apa ca donator de electroni şi în prezenţa luminii produc oxigen. Mecanismul lor fotosintetic este identic cu cel al algelor eucariote, pigmenţii fotosintetizanţi fiind reprezentaţi numai de clorofila "a" şi deficobiliproteine (alloficocyanina, ficocyanina şi, uneori, ficoeritrina).
Cyanobacteriile sunt strict fotoautotrofe şi nu se pot dezvolta numai în prezenţa substanţelor organice, nu au nevoie de vitamine ca factori de creştere, iar ca sursă de azot folosesc azotaţii, sărurile de amoniu, iar unele

70
specii folosesc azotul molecular, fiind fixatoare de azot (genurile Nostoc şi Anabaena).
Sunt mult răspândite în sol, în apele dulci şi in apele sărate din mări şi oceane. Sunt mai tolerante faţă de condiţiile extreme nefavorabile decât algele eucariote.
2.5.1. Morfologia cyanobacteriilor
Unele cyanobacterii sunt unicelulare, iar altele filamentoase, formând lanţuri de dimensiuni variabile. Forma celulei este specifică sau bacilară, cu dimensiuni cuprinse între 1 şi 60 µm.
2.5.2. Structura cyanobacteriilor Celulele prezintă un perete rigid, multistratificat, asemănător cu cel de
la bacteriile Gram-negative. Peretele celular poate fi acoperit la exterior cu o teacă gelatinoasă sau fibroasă. Cele mai multe cyanobacterii sunt mobile într-un anumit stadiu de dezvoltare, având o mobilitate de tip glisant, asemănându-se cu bacteriile filamentoase glisante de tip Beggiatoa.
În citoplasmă se observă 2-5 saci tilacoizi, între care se află ficobilisomii, implicaţi în procesul de fotosinteză; genomul, alcătuit dintr-o moleculă de ADN dublu catenară; granule de cianoficină, care conţin acid aspartic şi arginină; glicogen; polifosfat şi vacuole cu gaz, ce au rol de flotaţie. Ficobilisomii conţin pigmenţi asimilatori, reprezentaţi, în principal, de ficocyanină, alloficocyanină şi, uneori, de ficoeritrină şi ficoeritrocianină.
Heterochiştii, întâlniţi la speciile din ordinele Nostocales şi Stigonematales, sunt celule vegetative, care au suferit un proces de diferenţiere structurală şi funcţională. Astfel, ei au pierdut capacitatea de a realiza fotosinteza oxigenică, dar conţin enzime specifice procesului de fixare a azotului molecular (nitrogenaza, hidrogenaza). Prezintă un perete celular îngroşat şi o legătură cu celulele vegetative adiacente. În schimbul azotului organic furnizat de către heterochişti, celulele vegetative furnizează metaboliţi organici.
Akineţii reprezintă structuri de rezistenţă, care se formează prin creşterea în dimensiune şi deformarea unor celule vegetative, simultan cu

71
depunerea unor straturi fibrilare dense la exterior. Conservă capacitatea vitală a speciei pentru o perioadă lungă de timp şi oferă protecţie la acţiunea unor factori fizici (temperatură).
2.5.3. Sistematica cyanobacteriilor Cyanobacteriile (Fig. 52) sunt încadrate în Regnul Procaryotae,
Divizia Gracilicutes, Cl. Oxyphotobacteria şi, pe baza caracterelor morfologice, au fost clasificate în cinci ordine: • Chroococca.les cuprinde reprezentanţi unicelulari, grupaţi în asociaţii coloniale. Ex: genurile Chroococcus, Cyanobacterium şi Gloeocapsa. • Pleurocapsales cuprinde specii unicelulare, capabile să formeze spori interni. Ex: genurile Chroococcidiopsis, Pleurocapsa şi Stanieria. • Oscillatoriales cuprinde genuri, în care celulele sunt grupate în asociaţii filamentoase, fără a prezenta, însă, şi celule diferenţiate, numite akineţi şi heterocişti. Ex: genurile Oscillatoria, Plectonema şi Spirulina. • Nostocales cuprinde specii filamentoase cu celule diferenţiate, numite akineţi şi heterocişti. Ex: genurile Anabaena, Nostoc şi Cyanospira. • Stigonematales cuprinde specii filamentoase cu celule diferenţiate, numite akineţi şi heterocişti. Acest ordin, spre deosebire de Ord. Nostocales, include şi cyanobacteriile, care formează trichoame adevărate. Ex: genurile Capsosira, Mastigocladus şi Stigonema.
(a) (b) (c)
Fig. 52 – Cyanobacterii: (a) Chroococcus sp.; (b) Oscillatoria sp.; (c) Anabena sp.

72
2.5.4. Rolul cyanobacteriilor în natură Acestea constituie primele organisme ce populează rocile dezagregate,
participând la procesul de solificare, şi favorizează dezvoltarea în masa lor mucilaginoasă a unor bacterii fixatoare de azot, între care Azotobacter sp. şi Clostridium sp.
Dintre cyanobacterii, mai răspândite în natură sunt genurile Gloeocapsa şi Chroococcus, care sunt organisme unicelulare. Alte grupe sunt filamentoase ca: Oscillatoria şi Lyngbia. Genurile Nostoc şi Anabaena, ce au capacitate de a fixa azotul molecular, trăiesc în solurile umede din orezării, unde asigură N2 necesar plantelor.
2.6. Protozoarele Protozoarele (gr. protos – primul, zoon – animal) sunt organisme
unicelulare de tip eucariot, variate ca formă şi dimensiune, la care se cunoaşte un anumit ciclu evolutiv şi care reprezintă vieţuitoare complete şi independente.
Sunt caracterizate de prezenţa unor organite, numite cili, care participă la locomoţie şi la obţinerea hranei. Hrănirea este exclusiv heterotrofă; trăiesc libere în mediul acvatic (majoritatea trăiesc în ape dulci, dar pot fi întâlnite şi specii de apă sărată), precum şi în soluri umede.
2.6.1. Morfologia şi fiziologia protozoarelor
Protozoarele sunt variate ca formă, iar dimensiunile variază, în funcţie de specie, între 1 şi 2000 µm.
Valorile de temperatură optime pentru creştere şi dezvoltare sunt cuprinse între 18oC şi 25oC, iar chiştii speciilor din Cl. Rhizopoda (Sarcodina) rezistă pâna la 70oC. Protozoarele preferă un pH neutru, dar se pot dezvolta într-un interval mai larg, cuprins între 3,5 şi 9. În raport cu oxigenul, majoritatea protozoarele din sol sunt strict aerobe.
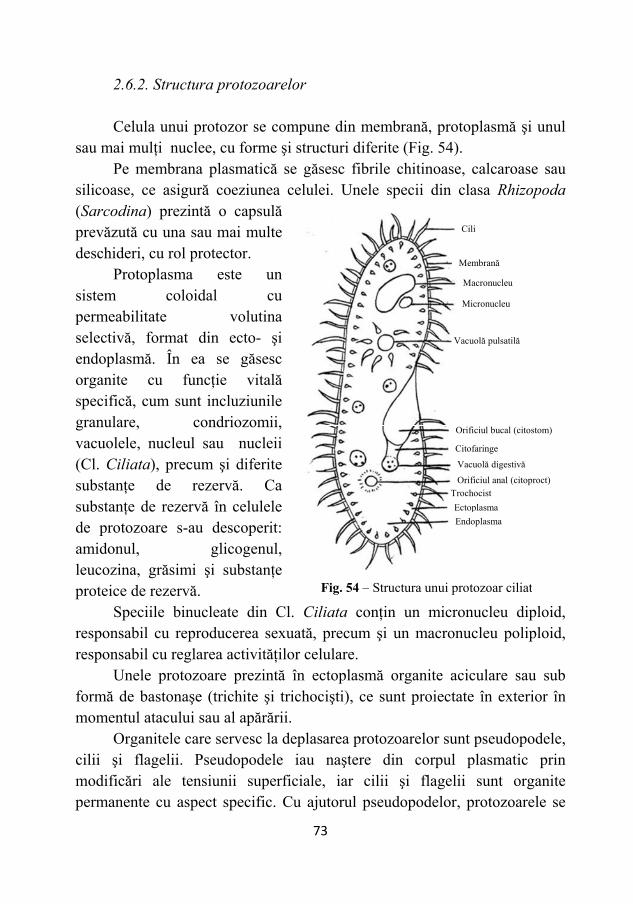
73
2.6.2. Structura protozoarelor Celula unui protozor se compune din membrană, protoplasmă şi unul
sau mai mulţi nuclee, cu forme şi structuri diferite (Fig. 54). Pe membrana plasmatică se găsesc fibrile chitinoase, calcaroase sau
silicoase, ce asigură coeziunea celulei. Unele specii din clasa Rhizopoda (Sarcodina) prezintă o capsulă prevăzută cu una sau mai multe deschideri, cu rol protector.
Protoplasma este un sistem coloidal cu permeabilitate volutina selectivă, format din ecto- şi endoplasmă. În ea se găsesc organite cu funcţie vitală specifică, cum sunt incluziunile granulare, condriozomii, vacuolele, nucleul sau nucleii (Cl. Ciliata), precum şi diferite substanţe de rezervă. Ca substanţe de rezervă în celulele de protozoare s-au descoperit: amidonul, glicogenul, leucozina, grăsimi şi substanţe proteice de rezervă.
Speciile binucleate din Cl. Ciliata conţin un micronucleu diploid, responsabil cu reproducerea sexuată, precum şi un macronucleu poliploid, responsabil cu reglarea activităţilor celulare.
Unele protozoare prezintă în ectoplasmă organite aciculare sau sub formă de bastonaşe (trichite şi trichocişti), ce sunt proiectate în exterior în momentul atacului sau al apărării.
Organitele care servesc la deplasarea protozoarelor sunt pseudopodele, cilii şi flagelii. Pseudopodele iau naştere din corpul plasmatic prin modificări ale tensiunii superficiale, iar cilii şi flagelii sunt organite permanente cu aspect specific. Cu ajutorul pseudopodelor, protozoarele se
Vacuolă pulsatilă
Orificiul bucal (citostom)
Orificiul anal (citoproct)
Vacuolă digestivă
Citofaringe
Micronucleu
Membrană
Cili
Macronucleu
EctoplasmaEndoplasma
Trochocist
Fig. 54 – Structura unui protozoar ciliat

74
deplasează pe substrat solid, iar cu ajutorul cililor şi flagelilor, în mediul lichid.
Resturile nedigerate sunt eliminate la cele cu pseudopode prin orice parte a membranei plasmatice, iar la celelate protozoare prin anusul celular (citoproct).
2.6.3. Reproducerea protozoarelor
Înmulţirea protozoarelor se face pe cale asexuată prin fisiune, sciziparitate sau înmugurire. Reproducerea prin sciziparitate (diviziune directă) presupune împărţirea celulei în două părţi aproape egale, în urma acestui proces rezultând două celule fiice.
Înmulţirea sexuată are loc după mai multe diviziuni simple, prin copulare între gameţi sau, în cazul speciilor din Cl. Ciliata, prin conjugare.
Copularea constă în unirea a doi gameţi identici (isogamie) sau diferiţi (heterogamie) din punct de vedere morfologic, cu formarea unui zigot. În urma contopirii gameţilor iau naştere zigoţii, care, în condiţii neprielnice, se transformă în zigochişti sau chişti de repaus, care au o mare rezistenţă la temperaturi nefavorabile, concentraţii mari de săruri solubile, uscăciune sau acumulare în mediul ambiant al unor factori nocivi. La revenirea condiţiilor favorabile, din zigochişti apar indivizi obişnuiţi.
Conjugarea apare la ciliate şi constă în unirea temporară printr-o punte plasmatică a doi indivizi, realizîndu-se, astfel, un schimb reciproc de nuclee urmat de o contopire a nucleului introdus cu cel preexistent în celulă.
2.6.4. Nutriţia protozoarelor Nutriţia este autotrofă, la speciile care conţin clorofilă (Cl.
Flagellata), şi heterotrofă (osmotrofă sau fagotrofă), în cazul celorlalte protozoare. Speciile heterotrofe pot fi: • saprozoice, utilizează substanţele organice moarte; • holozoice, se hrănesc cu microorganisme (bacterii, alge şi protozoare).
Majoritatea protozoarelor sunt holozoice şi, prin aceasta, pot limita populaţia bacteriană a solului. Amoebele înglobează hrana cu ajutorul extensiunilor plasmatice iar ciliatele îşi procură hrana prin invaginări de

75
diferite forme (citostom, citofaringe). Digestia hranei solide se face în vacuole digestive iar hrana lichidă este absorbită osmotic.
2.6.5. Sistematica protozoarelor
În actualul sistem de clasificare, protozoarele sunt încadrate în Regnul
Protista, Încrengătura Protozoa cu clasele: • Flagellata (Mastigophora) sunt protozoare prevăzute cu flageli ca organite de mişcare (gr. mastigos – flagel). În condiţii nefavorabile, unele specii îşi pot pierde flagelii şi se pot deplasa pe stratul solid printr-o mişcare de curgere (târâre), caracteristică rizopodelor. Flagelatele includ grupe de organisme cu apartenenţă fie la regnul vegetal (fitoflagelate), fie la regnul animal (zooflagelate). Ca reprezentanţi sunt genurile Bodo şi Euglena. • Rhizopoda (Sarcodina) sunt protozoare care au corpul celular nud, cu formă instabilă, care formează chişti. Ele emit pseudopode temporare, care le servesc atât pentru deplasare, cât şi pentru înglobarea hranei. Unele specii pot fi protejate de o cochilie cu una sau mai multe deschideri. Cele mai răspândite genuri sunt Negleria şi Amoeba. • Ciliata (Infusoria) cuprinde protozoare care se mişcă cu ajutorul a numeroşi cili, care sunt excrescenţe protoplasmatice. Ciliatele prezintă cel puţin două nuclee diferite, atât morfologic, cât şi funcţional. Din acest grup, ca reprezentanţi, cităm: Paramoecium, Balantiophorus şi Colpoda. • Sporozoa include organisme unicelulare parazite, caracterizate prin apariţia în timpul ciclului de dezvoltare al stadiului de spor. Ca reprezentanţi sunt genurile Lankesterella şi Schellackia. • Cnidosporidia (Amoebosporidia) cuprinde specii unicelulare parazite.
(a) (b) (c) Fig. 53 – Protozoare: (a) Euglena sp.; (b) Amoeba sp.; (c) Paramoecium sp.

76
2.6.6. Rolul protozoarelor în natură Rolul protozoarelor în sol este secundar faţă de cel al bacteriilor şi
ciupercilor. Ele sunt, în majoritate, prădători, ce se hrănesc cu bacterii şi se găsesc în sol, până la 15 cm adâncime, într-o densitate care variază între câteva sute şi până la 10.000 de exemplare la 1 gram de sol. Ca microprădători, protozoarele contribuie la diminuarea numărului de bacterii în sol.
În acelaşi timp, s-a constatat că, prin moartea lor, protozoarele pun la dispoziţia microorganismelor surse nutritive, care stimulează creşterea numărului acestora în sol.
În unele experienţe cu Azotobacter, în culturile la care s-au adăugat protozoare (Colpidium colpoda), s-a fixat mai mult azot decât la culturile martor. De aici, se trage concluzia că protozoarele, fagocitând bacteriile, forţează colonia să producă mai multe celule tinere, care sunt mai active în fixarea azotului molecular. Pe de altă parte, se presupune că protozoarele ar elimina în mediu de cultură unele substanţe stimulatoare pentru dezvoltarea bacteriilor.
După alţi autori, în afară de acţiunea indirectă asupra biologiei solului, protozoarele ar contribui şi la formarea humusului.
2.7. Diatomeele Diatomee (gr. dia – de-a curezişul; temnein – a tăia) – clasă de alge
fotosintetizante, unicelulare, brune-gălbui, care participă la formarea fitoplanctonului. Prezintă o nutriţie de tip autotrof, iar nucleul este de tip eucariot.
2.7.1. Morfologia diatomeelor
Diatomeele sunt microorganisme solitare sau unite în colonii sub
formă de filament sau fundă (Fragillaria spp.), evantai (Meridion spp.), zig-zag (Tabellaria spp.) sau stea (Asterionella spp.). Multe diatomee secretă o substanţă mucilaginoasă, care acoperă peretele silicios şi care serveşte ca mijloc de ataşare a celulelor în colonii. Substanţa mucilaginoasă poate fi

77
utilizată şi pentru ataşarea diatomeelor de substrat. Celulele mai pot fi unite în colonii prin diferite excrescenţe ale frustulei.
Dimensiunea diatomeelor variază de la un μm la mai mult de 2000 μm, dar cele mai multe se încadrează în intervalul 10 - 100 μm.
2.7.2. Structura diatomeelor O trăsătură caracteristică a algelor diatomee este că acestea sunt
închise într-un perete celular pectocelulozic, format din două teci silicioase (epiteca şi hipoteca), numit frustula (schelet extern). Fiecare teacă este formată dintr-o valvă şi un inel conectival. Epiteca este mai mare comparativ cu hipoteca, astfel că cele două teci se suprapun asemenea unei plăci Petri, îmbinarea realizându-se prin marginile libere ale inelelor conectivale (Fig. 58).
Frustula poate prezenta o serie de elemente structurale: puncte, striuri, coaste, cutări sau loje alungite, deschise în cavitatea celulară. Suprafaţa valvei poate fi reticulată, cu o serie de cavităţi, numite areole. Peretele areolei poate prezenta pori, prin care citoplasma comunică cu exteriorul. La unele specii din Ord. Pennales se observă prezenţa unor fisuri, care permit comunicarea citoplasmei cu mediul extern şi care poartă numele de rafă. La celelalte specii ale ordinului se găseşte o pseudorafă.
Frustulele de diatomee sunt, de obicei, foarte complicate şi variate, iar modelele de ornamentaţie reprezintă un criteriu taxonomic important de clasificare.
Membrana citoplasmatică separă citoplasma de peretele celular (faţa internă a frustulei). Citoplasma diatomeelor prezintă curenţi citoplasmatici, iar în interiorul ei se găsesc organitele citoplasmatice.
Fig. 58 – Frustulă de diatomee (CRPGL, Luxembourg)

78
Nucleul este bine dezvoltat, are o formă sferică sau lenticulară şi prezintă 1-2 nucleoli. Citoplasma conţine cromoplastide (cromatofori) în care, alături de clorofilă (a şi c), se găseşte β-caroten, fucoxantină, lutenină şi diatomină, pigmenţi ce dau o culoare galben-brună. Din punct de vedere filogenetic, se crede că diatomeele fac trecerea spre algele brune, datorită existenţei pigmentului diatomină (feofil), care maschează prezenţa clorofilei.
Diatomeele sunt organisme autotrofe, capabile de sinteza tuturor
constituenţilor celulari prin mijloace proprii, pornind de la surse simple anorganice de carbon şi azot.
Deplasarea diatomeelor cu rafă se poate realiza prin glisare cu ajutorul citoplasmei, care exudă la nivelul acesteia. Speciile care nu prezintă rafa, nu se pot deplasa şi sunt dispersate pasiv de curenţi în straturile de suprafaţă ale oceanelor. Adaptările care favorizează plutirea includ: forma frustulelor şi procesele care măresc raportul arie/volum, formarea coloniilor şi înmagazinarea de grăsimi sau uleiuri în celulă (reduc greutatea specifică totală). Celula diatomeelor nu prezintă flageli, cu excepţia gameţilor flagelaţi din timpul perioadei de reproducere sexuată.
Din punct de vedere chimic, algele diatomee conţin 80-90% SiO2. 2.7.3. Reproducerea diatomeelor
Diatomeele se înmulţesc asexuat prin diviziune binară şi sexuat prin
oogamie, isogamie sau anisogamie (heterogamie).
Fig. 59 – Structura la Melosira spp. (după Margulis şi colab., 1999)

79
Reproducerea prin diviziune binară (Fig. 60) presupune scindarea celor două teci după un plan paralel, astfel încât fiecare dintre cele două celule fiice rezultate va avea câte o teacă de la celula mamă. Cealaltă teacă va fi rapid secretată de protoplast.
Deoarece hipoteca celulei mamă devine epiteca pentru celulă fiică, creşterea numerică a populaţiei de diatomee determină o reducere a dimensiunii medii. Ca urmare a acestui fapt, jumătate din celulele nou formate vor suferi o reducere continuă a dimensiunilor, iar celelalte celule vor avea aceleaşi dimensiuni, comparativ cu celula parentală. Reproducerea asexuată se poate repeta de până la opt ori pe zi, factorii limitativi fiind cantitatea de silice existentă în mediu şi dimensiunea celulei mamă.
După o serie de diviziuni succesive, algele diatomee vor ajunge la o
dimensiune minimă (<1/3 din celula iniţială), caz în care mitoza nu mai poate avea loc. Acest lucru induce reproducerea sexuată, care va restabili dimensiunea maximă a celulei de diatomee.
Reproducerea sexuată (Fig. 61) este diferită la diatomeele centrice (Ord. Centrales) şi penate (Ord. Pennales) şi se realizează prin oogamie, isogamie sau anisogamie (heterogamie). Ciclul de viaţă al diatomeelor este preponderent diploid (2n), stadiul haploid (n) fiind de scurtă durată (în timpul înmulţirii sexuate).
Oogamia este un tip de proces sexuat caracteristic pentru diatomeele centrice, care constă în contopirea gametului femel imobil cu gametul mascul mobil (prezintă un flagel).
Mitoză
Fig. 60 – Reproducerea asexuată la diatomee (după MacDonald, 1869 şi Pfitzer, 1869)

80
În cazul diatomeelor penate, reproducerea sexuată se realizează prin isogamie sau, mai rar, prin anisogamie. Isogamia este o formă de înmulţire sexuată, în care cei doi gameţi care participă la fecundare sunt identici din punct de vedere morfologic, în timp de anisogamia (heterogamie) se realizează prin copulaţia a doi gameţi diferiţ i ca mărime şi formă. În cazul Ord. Pennales, cu excepţia genului Rhabdonema, reproducerea sexuată se realizează prin intermediul unor gameţi imobili, nediferenţiaţi din punct de vedere morfologic.
Reproducerea sexuată este declanşată de reducerea în dimensiune a frustulei. Când celulele vegetative (2n) ajung la dimensiunea minimă, se iniţiază fenomenul de meioză, în urma căruia sunt produşi gameţi masculi şi femeli. După fuziunea celor două nuclee, diferite din punct de vedere sexual, rezultă zigotul, care începe să crească în volum până ajunge la dimensiunea celulei caracteristice pentru specia respectivă. Stadiul final poartă numele de auxospor (gr. auxein – a creşte, sporos – germen) şi este caracterizat prin absenţa frustulei silicioase şi prin capacitatea de a forma o nouă celulă vegetativă prin germinare.
A B
Fig. 61 – Reproducerea sexuată la diatomee: A – reproducerea sexuată la Ord. Centrales; B - reproducerea sexuată la Ord. Pennales (după Hasle and Syvertsen, 1997).

81
2.7.4. Sistematica diatomeelor Whittaker (1969) şi Bayard (1978) plasează diatomeele în Cl.
Bacillariophyceae, alături de algele din Cl. Chrysophyceae (alge maro-aurii) şi Cl. Xanthiophyceae (alge galben-verzi). Cele trei clase sunt încadrate taxonomic în Încr. Chrysophyta, Regnul Protista şi se caracterizează prin formarea de chişti endoplasmatici asemănători, secreţia de silice, acumularea de grăsimi şi existenţa unui perete celular bipartit în anumite stadii de dezvoltare. Alţi cercetători (Woese şi colab., 1990; Margulis şi Schwartz, 2000) subliniază diferenţele existente între clasele enumerate anterior (în special structura frustulei de diatomee) şi au ridicat diatomeele la rang de încrengătura separată, Bacillariophyta (sin. Diatomeae).
O mare parte din sistematica diatomeelor este bazată pe tipurile şi modelele ornamentaţiilor, care apar pe scheletul extern opalin sau frustula. Aceasta frustulă are formă de cutie cu teci (valve) suprapuse, cea mai mare fiind numită epiteca, iar cea mai mică, hipoteca. Structurile cele mai importante sunt reprezentate de pori (areole), arii hialine (clare), spini marginali şi submarginali.
Sunt cunoscute peste 250 genuri de diatomee (70 genuri fosile); acestea cuprind aproximativ 100.000 specii, dintre care peste 10.000 existente. Dintre genurile mai comune menţionăm: Amphipleura, Asterionella, Biddulphia, Coscinodiscus, Cymbella, Diatoma, Diploneis, Fragilaria, Melosira, Meridion, Navicula, Nitzschia, Pinnularia, Surirella şi Thalassiosira.
Diatomeele sunt divizate în două ordine (după Whittaker, 1969 şi Bayard, 1978): • Ord. Centrales prezintă simetrie radiară (centrul structural este
reprezentat de un punct). Ex: Fam. Coscinodiscaceae cuprinde specii cu frustule discoidale,
lenticulare sau globuloase, unite în colonii filamentoase. Celulele se prezintă sub formă de disc sau de tobă, libere sau fixate, solitare sau unite în colonii laxe cu ajutorul unui mucilagiu. Valvele sunt circulare sau eliptice cu relieful ondulat tangenţial sau concentric. Specii marine, salmastre sau dulcicole. Unele se fixează pe diferite suporturi umede sau pe soluri (Fig. 55).

82
• Ord. Pennales prezintă simetrie bilaterală (centrul structural este
reprezentat de o linie). Acest ordin cuprinde:
1. diatomee fără rafă Exemplu : Fam. Fragilariaceae cuprinde diatomee penate de ape
dulci şi marine, ale căror celule sunt izolate sau reunite în colonii. Celulele pot fi alungite, drepte sau curbate, cu valve ornamentate cu striuri sau coaste transversale. Frustula este lipsită de zone intercalare şi de septuri (Fig. 56).
2. diatomee cu rafă Exemplu: Fam. Naviculaceae cuprinde diatomee penate, care la
ambele valve prezintă câte o rafă bine dezvoltată, sub forma unor fisuri. Rafa este situată pe o carenă. Cuprinde 29 de genuri de apă dulce, salmastră
Fig. 56 – Fragilaria capucina var. amphicephala
(http://craticula.ncl.ac.uk)
Fig. 55 – Cyclotella meneghiniana
(www.pae.ugent.be)

83
sau marină. Unele specii se fixează pe soluri umede, pe ziduri sau pe diferite suporturi umede (Fig. 57).
2.7.5. Rolul diatomeelor în natură Se găsesc în toate mediile marine, salmastre şi de apă dulce, dar şi în
sol (adâncimi < 0,25 - 1 m), gheaţă sau ataşate de roci şi alte substraturi din zonele de spargere a valurilor de lângă ape. În oceanele actuale (adâncimi <100 m), diatomeele sunt printre cei mai importanţi producători primari, iar o abundenţă ridicată a diatomeelor este tipică pentru cele mai fertile arii ale oceanelor. Diatomeele pot fi folosite pentru monitorizarea condiţiilor mediului şi în studiul calităţii apei.
2.8. Ciupercile
Ciupercile sunt organisme eucariote, heterotrofe, ubiquiste, extrem de bogate în specii ce prezintă structuri şi caractere biologice foarte diverse, adaptate modurilor de viaţă saprofite, parazite sau simbiotice, fiind lipsite de pigmenţi asimilatori.
2.8.1. Morfologia ciupercilor Aparatul vegetativ al diferitelor grupe taxonomice de ciuperci este
divers constituit (Fig. 62). Astfel, la cele mai primitive ciuperci din clasa Chytridiomycetes, corpul vegetativ este unicelular, sub forma unei celule
Fig. 57 – Navicula hamiltonii (http://craticula.ncl.ac.uk)

84
nude, format numai din citoplasmă şi nucleu şi poartă numele de gimnoplast.
La un alt grup de ciuperci inferioare din cl. Plasmodiophoromycetes, corpul vegetativ este constituit dintr-o masă citoplasmatică mai dezvoltată, în care se găsesc numeroase nuclee neindividualizate în celule, ce poartă numele de plasmodiu.
Un alt tip de aparat vegetativ mai evoluat, denumit sifonoplast, se întâlneşte la ciupercile din clasele Zigomycetes şi Oomycetes, la care acesta se prezintă sub formă de tuburi simple sau ramificate continuie, fără pereţi transversali, care conţin o masă de citoplasmă şi numeroase nuclee, nediferenţiate în celule, denumit şi sifonoplast cenocitic.
La ciupercile superioare din subîncrengăturile Ascomycotina, Basidiomycotina şi Deuteromycotina, aparatul vegetativ poate fi unicelular ca la Fam. Saccharomycetaceae şi poartă numele de dermatoplast, iar la marea majoritate se prezintă sub formă de filamente simple sau ramificate, cu pereţi transversali sau septe, care separă celulele între ele. Filamentele acestea se numesc taluri filamentoase, carora li se mai dă denumirea de micelii, iar ramificaţiilor, aceea de hife miceliene. Septele sau pereţii transversali, ce despart celulele talului filamentos, sunt prevăzute cu un por central, prin care se face comunicarea între celulele vecine, prin masa de citoplasmă.
Cel mai evoluat tip de aparat vegetativ este alcătuit dintr-o împletire de taluri filamentoase, poartă numele de tal masiv şi este caracteristic ciupercilor din subîncrengătura Basidiomycotina şi unor ciuperci din subîncrengătura Ascomycotina. Prin împletirea filamentelor apar ţesuturi false, numite plectenchime, ce pot îndeplini diferite funcţii (de protecţie, de absorbţie, de rezervă etc.).

85
2.8.2. Structura celulară a ciupercilor Celula fungică este formată din două feluri de componente şi anume
din elemente neprotoplasmatice, cum sunt, membrana, vacuomul şi substanţele de rezervă, şi din componente protoplasmatice, ca citoplasma, nucleul etc.
1. Peretele celular, examinat la microscopul optic, apare ca o structură omogenă, însă la o examinare cu microscopul electronic prezintă o structura stratificată.
După Hunsley şi Burnett, spre exemplu, miceliul matur la Neurospora crassa este constituit din mai multe straturi, după cum urmează: stratul
(a) (b)
(e) (f)
(c) (d)
Fig. 62 – Tipuri de aparate vegetative la ciuperci: (a) gimnoplast; (b) plasmodiu; (c) dermatoplast; (d) sifonoplast; (e) tal filamentos; (f) tal masiv (după Lazăr şi colab., 1977).

86
exterior, compus din glucan amorf este urmat de un alt strat reticular din glucan în amestec cu proteină; al treilea strat este constituit, în principal, din proteină, iar cel de-al patrulea este format din proteină, în care sunt cuprinse şi microfibrile de chitină.
2. Membrana citoplasmatică acoperă citoplasma celulelor fungice şi o separă de faţa interioară a peretelui celular. Apare la microscopul electronic tristratificată, are o grosime variabilă şi funcţionează ca o membrană osmotică selectivă.
3. Vacuolele, puse în evidenţă prin colorarea cu roşu neutru, sunt mai numeroase în celulele tinere, dar, cu vârsta, confluează într-o vacuolă mare, ce ocupă centrul celulei. Sunt separate de citoplasmă printr-o membrană simplă, denumită tonoplast, iar în interiorul lor se acumulează substanţe de rezervă sau care rezultă în urma procesului metabolic.
4. Citoplasma celulelor de ciuperci se prezintă sub formă lichidă, hialină, omogenă, refringentă şi, uneori, la maturitate, cu aspect granular. În compoziţia ei intră: lipide, glicogen, acid ribo nucleic, enzime, acizi organici, fosfaţi de fitosterine, zaharuri etc. În citoplasmă, al cărei conţinut în apă este însemnat, se desfăşoară majoritatea proceselor de metabolism; tot aici se găsesc vacuolele, ribozomii, reticolul endoplasmatic, mitocondriile şi anumite vezicule subglobuloase, mici, cu substanţe de rezervă.
5. Nucleul, la ciuperci, este înconjurat de o membrană nucleară dublă, ce prezintă, din loc în loc, orificii. În interior se află nucleoplasma şi unul sau mai mulţi nucleoli. Substanţa caracteristică nucleului este acidul dezoxiribonucleic (ADN) iar numărul de cromozomi este mic şi diferă în funcţie de specie. Având nuclee bine conturate, ciupercile sunt organisme eucariote. Nucleele la ciuperci sunt, de obicei, de formă sferică sau ovoidă şi au dimensiuni relativ mici, între 0,75 µm la Venturia inaequalis, 1-2 µm la Mucoraceae şi pot ajunge până la 20 µm.
La ciupercile inferioare, cu aparatul vegetativ un plasmodiu, în citoplasma acestuia se găsesc mai multe nuclee ca şi în sifonoplastul care nu prezintă septe transversale.
La ciupercile superioare din subîncrengăturile Ascomycotina şi Basidiomycotina se pot găsi în celule un singur nucleu în faza haploidală şi două nuclee în cea dicariotică.

87
6. Reticulul endoplasmatic este alcătuit din două membrane, separate printr-un lumen; el poate fi sub formă tubulară sau sub formă lamelară, mai evident în celulele tinere şi în legătură cu membrana nucleară.
7. Ribozomii, denumiţi şi granulele lui Palade, sunt formaţiuni citoplasmatice, asociate reticului endoplasmatic. Ei sunt formaţi din ARN în proporţie de până la 65 %, numit şi ARN ribozomal (ARNr), şi din 35 % proteină.
8. Aparatul Golgi este prezent în celulele ciupercilor, fiind format din unităţi structurale morfologice, numite dichtiozomi, ce se găsesc în citoplasmă. Are rol în procesul de sinteză a polizaharidelor, participă la procesul de formare şi creştere a membranei celulare şi a vacuolelor, fiind întâlnit, mai ales, la ciupercile Oomycetes.
2.8.3. Organele de fixare şi de absorbţie ale ciupercilor Ciupercile saprofite se fixează pe substrat cu ajutorul unor filamente
ramificate, numite rizoizi, cu ajutorul cărora îşi extrag şi substanţele nutritive.
Ciupercile parazite pătrund în ţesuturile gazdei, dar miceliul rămâne în spaţiile intercelulare trimiţând în celule haustorii care au rolul de a extrage din celule substanţele hrănitoare (Fig. 63).
Rizoizi
Haustori
Fig. 63 – Organe de fixare şi absorbţie la ciuperci

88
Haustorii sunt constituiţi dintr-un gât subţire, format în momentul pătrunderii în celula parazitată, după care urmează o parte mai dilatată-terminală, ce poate avea forme sferice, ca la Cystopus sp., ramificate sau digitate, ca la genurile Peronospora, Erysiphe sau Puccinia.
Unele ciuperci parazite, care se dezvoltă la suprafaţa organelor atacate ale plantei gazdă, se numesc ectoparazite, iar miceliul lor este epifit. Aşa este cazul ciupercilor din Fam. Erysiphaceae, la care miceliul se fixează de substrat cu ajutorul unor formaţiuni ca nişte ventuze, numite apresorii, din care se dezvoltă mai departe micelii fine, ce străbat membrana celulară a gazdei şi formează apoi haustoriile în interiorul celulei parazitate.
În ceea ce priveşte răspândirea miceliului parazit în ţesuturile gazdelor la ciupercile endoparazite, acesta are o dispersie limitată, provocând infecţii localizate, ca în cazul petelor produse de Polystigma rubrum. La alte grupe de agenţi patogeni, infecţia este generalizată, afectând planta în totalitate sau mari părţi din aceasta, ca în cazul atacului de Puccinia recondita sau Erysiphe graminis.
În alte cazuri, spre exemplu la Verticillium dahliae, se produce o infecţie a vaselor conducătoare în zona coletului, provocând o tracheomicoză, iar efectul parazitar este de “veştejire”, ce afectează toată planta.
Un alt aspect ce trebuie menţionat este durata de viaţă a aparatului vegetativ al agentului patogen în gazdă. Din acest punct de vedere, există ciuperci cu dezvoltare anuală, cum este Erysiphe graminis, care, într-o singură perioadă de vegetaţie, îşi desăvârşeşte întreg ciclul biologic.
O a doua categorie o constituie ciupercile bianuale, care se dezvoltă pe plante multianuale sau pe două gazde diferite, cum este Puccinia graminis.
Există şi ciuperci plurianuale, care au ca gazde plante perene, ca pomii fructiferi sau arborii de pădure, pe a căror organe atacate corpii de fructificare durează un număr mai mare de ani. Aşa sunt ciupercile macromicete ce atacă scoarţa şi lemnul ca Stereum purpureum.
La un mare număr de ciuperci din subîncrengăturile Ascomycotina şi Basidiomycotina corpii de fructificare sau unele organe de rezistenţă sunt alcătuiţi din filamente miceliene mai strâns sau mai lax întreţesute, ce alcătuiesc false ţesuturi, denumite plectenchime.

89
Plectenchimele, care constituie taluri masive ale unor ciuperci macromicete, îndeplinesc funcţii diferite şi alcătuiesc false ţesuturi, de creştere, de protecţie, mecanice, de conducere, de rezervă, de excreţie, de mişcare etc.
Falsele ţesuturi de creştere sau formatoare sunt constituite din celule cu caracter embrionar, cu membrana subţire, foarte bogate în citoplasmă, care se divid activ şi îndeplinesc funcţia de creştere a talului.
Falsele ţesuturi de protecţie se găsesc la suprafaţa talului masiv, venind în contact cu mediul extern. Elementele care alcătuiesc aceste ţesuturi au membrana îngroşată, prezentând, uneori, şi anumite excrescenţe, ornamentaţii sau peri.
La ciuperci, se pot deosebi mai multe feluri de ţesuturi de protecţie şi anume: • Derm este o ţesătură formată din celule dispuse perpendicular pe suprafaţa organelor ciupercii. • Cutis este ţesătura de protecţie, ale cărei elemente constitutive sunt dispuse paralel cu suprafaţa organelor ciupercii. • Cortex este o ţesătură de protecţie compactă, dispusă, de regulă, la suprafaţa unor fructificaţii, apotecii, basidiofructe sau a scleroţilor.
Falsele ţesuturi mecanice au rol în susţinerea talului, cum ar fi piciorul ciupercilor macromicetelor, şi sunt constituite din elemente mecanice bine diferenţiate, formate din celule cu membrana puternic îngroşată, lipsite de conţinut, ce sunt comparabile sclereidelor.
Falsele ţesuturi conducătoare se aseamănă şi au acelaşi rol cu ţesuturile de la plantele vasculare. Elementele ce alcătuiesc aceste formaţiuni sunt celule vii mult alungite, cu pereţi transversali subţiri, prevăzuţi cu pori şi care prezintă dilatări în dreptul septului.
Falsele ţesuturi de rezervă au rolul de a înmagazina apa, hidraţii de carbon, substanţele proteice, grăsimile etc..
Celulele în care se depune apa se numesc cistide, dar aceasta, în unele cazuri, se poate găsi şi în spaţiile intercelulare.
Glicogenul şi grăsimile se depun în celule sub formă de picături, iar în membrane se depune hemiceluloza. La ciuperci, se cunoaşte că amidonul nu se găseşte ca substanţă de rezervă. Pe lângă aceste false ţesuturi de rezervă,

90
ca organe speciale, mai au rol în acest proces şi scleroţii, bulbilii şi rizomorfele, care sunt şi organe de rezistenţă.
Falsele ţesuturi de excreţie sunt acelea prin care se face eliminarea unor produse de metabolism sub formă de secreţii sau excreţii.
Din categoria produselor de secreţie amintim enzimele şi hormonii, ce se elimină din corpul ciupercilor în procesul de parazitism.
Substanţele de excreţie, ca oxalatul de calciu, carbonatul de calciu, unele substanţe gomoase, formate în procesul de asimilaţie şi de dezasimilaţie, se elimină permanent din corpul ciupercilor pe măsura formării lor.
Falsele ţesuturi de mişcare pot fi influenţate de un excitant extern şi pot provoca mişcări, denumite tropisme şi tactisme. Pe lângă acestea se mai cunosc la ciuperci şi mişcări autonome, numite nastii.
Între tropisme se pot deosebi: • Geotropismul, care poate fi pozitiv, în cazul organelor de fixare şi de absorbţie, cum sunt rizoizii, sau negativ, în cazul carpoforilor de la Hymenomycetales. • Fototropismul este influenţa produsă de lumină asupra unor organe de ciuperci; el poate fi pozitiv şi se manifestă asupra ascelor de la unele Discomycetes, care au tendinţa de a se orienta către lumină, sau a sporangioforilor de Pilobolus cristalinus, care, de asemenea, manifestă aceeaşi tendinţă. • Chemotropismul reprezintă tendinţa de atragere sau de respingere a unor organisme de ciuperci faţă de unele substanţe chimice. Chemotropism pozitiv manifestă numeroase ciuperci faţă de glucide şi zaharuri. • Hidrotropismul este influenţa umidităţii asupra ciupercilor. Se ştie că, în diferite faze biologice, acestea au nevoie de o atmosferă aproape saturată în vapori de apă sau chiar de picături pentru mişcarea şi germinarea sporilor şi iniţierea procesului de infecţiune. • Haptotropismul reprezintă mişcările specifice pe care le exercită unele ciuperci ce au tendinţa de a apuca sau înlănţui alte filamente, ca în cazul de la Peziza sp.
Tactismele sunt mişcări libere ale sporilor mobili (zoosporilor) ale unor ciuperci sub influenţa unui excintant extern. În funcţie de factorul excitant, putem avea: chemotactism, fototactism şi hidrotactism.

91
Nastiile sunt, de asemenea, mişcări autonome, dar care nu sunt provocate de un excitant extern.
De exemplu, se consideră ca nastii mişcările de deschidere a fructificaţiilor la unele Gasteromycete, a ecidiilor de la Uredinales, precum şi mişcările de eliminare a ascosporilor din asce sau de proiectare a bazidiosporilor de pe bazidii.
2.8.4. Formele de rezistenţă ale ciupercilor În situaţia când condiţiile de viaţă încetează să mai fie favorabile,
ciupercile au posibilitatea de a supravieţui, trecând în forme de rezistenţă, la care miceliul prezintă membrana celulară îngroşată, conţinutul citoplasmei mai bogat în substanţe de rezervă iar cel al apei este mai redus.
În acest fel, ritmul metabolismului devine mai lent şi permite supravieţuirea până la revenirea condiţiilor favorabile. Uneori, apar şi unele forme de rezistenţă cu o anumită structură, cum sunt: gemele, stromele, scleroţii, rizomorfele sau clamidosporii.
1. Gemele sunt forme de rezistenţă ce apar, mai ales, la ciupercile din cl. Oomycetes, la care porţiuni de sifonoplast se înconjoară cu o membrană îngroşată, conţinutul în apă se reduce şi, în acelaşi timp, cel al substanţelor de rezervă sporeşte. Privite la microscop, gemele apar mai strălucitoare, mai refringente decât miceliul vegetativ.
2. Clamidosporii sunt asemănători cu gemele, dar se formează la ciupercile superioare, ca de exemplu la genul Fusarium.
3. Stromele reprezintă formaţiuni uneori de dimensiuni mai mari, rezultate din împletirea miceliilor vegetative, luînd aspectul de crustă, ce se formează pe organele atacate ale plantelor. Stromele adăpostesc adesea forme conidiene sau sexuate de înmulţire şi sunt viu colorate sau în brun întunecat.
4. Scleroţii (Fig. 64) sunt organe de rezistenţă, ce reprezintă o împletire de filamente miceliene diferenţiată, în stratul extern mai dens, paraplectenchimul, şi mai laxă în interior, ce reprezintă prozoplectenchimul. Scleroţii se formează fie în locul ovarelor florilor, fie la exteriorul şi în interiorul altor organe ale plantelor atacate.

92
Din germinarea scleroţilor pot rezulta micelii vegetative, forme conidiene sau, la unele specii, apotecii cu asce şi ascospori.
5. Rizomorfele sunt formaţiuni ce rezultă din înmănuncherea a mai multe filamente miceliene strâns asociate, ce se formează pe părţile subterane ale plantelor atacate.
2.8.5. Înmulţirea ciupercilor
La ciupercile inferioare nu există o diferenţă tranşantă între aparatul vegetativ şi organele de îmnulţire. Astfel, la ciupercile din clasele Chytridiomycetes şi Plasmodiophoromycetes, gimnoplaştii şi, respectiv, plasmodiile capătă rol de înmulţire, transformându-se, pe cale holocarpică, în sporangi, ce conţin spori asexuaţi.
La ciupercile superioare, pe organele vegetative se formează, în mod diferenţiat, organe de reproducere, pe cale eucarpică.
Ciclurile biologice ale diferitelor grupe taxanomice sunt foarte diversificate şi se succed în timp în cadrul alternanţei de faze.
Formele de reproducere sunt denumite cu un termen generic spori, iar procesul de înmulţire se poate realiza pe cale vegetativă asexuată şi sexuată.
I. Înmulţirea vegetativă este o formă nespecializată, ce se realizează prin fragmente de miceliu, ce se diseminează fie pe cale naturală, fie în mod artificial de către om.
Spre exemplu, odată cu lucrările solului, unele fragmente de miceliu sunt desprinse şi purtate cu piesele active ale agregatelor la distanţe uneori importante. Factorii naturali meteorologici au ca efect acelaşi rezultat.
Alteori, unele specii de ciuperci, prin separarea celulelor miceliului, aşa cum s-ar produce deşirarea unui şirag de mărgele, formează spori, cum sunt artrosporii, aleurisporii sau clamidosporii,
Fig. 64 – Scleroţi de Sclerotinia sclerotiorum

93
care contribuie, de asemenea, la diseminare. Artrosporii sunt porţiuni delimitate de miceliu, ce se înconjoară cu o
membrană groasă şi servesc ca organe de propagare. Un asemenea caz se întâlneşte la Neurospora sitophila sau la Oidium lactis.
Aleurisporii se formează terminal pe filamentele miceliene şi se întâlnesc la genurile Aleurisma, Sepedonium etc.
Clamidosporii se formează intercalar prin fragmentarea hifelor ciupercii, sunt prevăzuţi cu perete gros şi rezistent, adeseori cu ornamentaţii specifice.
Dermatoplaştii ciupercilor din Fam. Saccharomycetaceae înmuguresc, iar aceste formaţiuni poartă numele de blastospori (Fig. 65).
II. Înmulţirea asexuată Acest mod de înmulţire se realizează prin organe diferenţiate, ce se
formează în strânsă dependenţă de condiţiile de nutriţie şi de climă, într-un moment determinant din dezvoltarea ciclului biologic al ciupercii. Organele de înmulţire asexuată sunt denumite în termeni generici spori.
Aceştia au rol de înmulţire şi de propagare şi reproduc faza asexuată, putând, uneori, să ia şi rolul de gameţi şi să contribuie la înmulţirea sexuată.
După posibilităţile de mişcare, sporii ciupercilor pot fi de două feluri; • Sporii imobili sau aplanospori sunt lipsiţi de flageli sau cili şi sunt dispersaţi în natură în mod pasiv;
Blastospor
Fig. 65– Înmulţirea vegetativă la drojdia de bere (Saccharomyces cerevisiae)

94
• Sporii mobili sau planospori sunt prevăzuţi cu 1-2 flageli, care asigură dispersarea în mediul acvatic.
Din punct de vedere al modului cum se formează, sporii pot fi de două feluri: • Spori endogeni sau endospori, care se găsesc în interiorul unor organe speciale. Astfel, ciupercile din clasa Zygomycetes formează spori imobili în interiorul unor sporangi. La ciupercile din clasa Oomycetes, sporii sunt mobili şi se formează în zoosporangi caduci, denumiţi, în mod curent, şi false conidii.
Există şi unele cazuri la ciupercile din subîncrengătura Ascomycotina când o categorie de spori imobili se formează în organe speciale - ascele; formarea acestor spori este în legătură cu procesul sexual şi ei se numesc ascospori. • Sporii exogeni sau exosporii se formează la exteriorul unor organe speciale, diferenţiate în funcţie de faza în care se găsesc ciupercile respective. Aşa, unii spori haploidali se formează la capătul unor filamente cu conformaţie deosebită, ce se numesc conidiofori, iar sporii poartă numele de conidii (Fig. 66).
Sporii exogeni, denumiţi basidiospori, se întâlnesc la subîncrengătura Basidiomycotina; ei se formează la exteriorul basidiilor, după procesul de sexualitate (meioza), şi ca şi ascosporii sunt spori haploidali.
III. Înmulţirea sexuată Asupra problemei sexualităţii la ciuperci s-au elaborat încă din secolul
al XIX-lea şi în cursul celui de al XX-lea lucrări, studii importante şi
Spori (conidii)
Spori (conidii)
(a) (b) Fig. 66 – Înmulţire asexuată prin spori la genurile Aspergillus (a) şi Penicillium (b)

95
sinteze, cum sunt cele ale unor autori ca: Clausen (1906), Guillermond (1913), Killian (1921), Kniep (1928), Gäumann (1940), Moreau (1952), Ainsworth şi Sussman (1973) etc.
Ca şi la organismele mai evoluate şi în ciclul de viaţă al ciupercilor există două faze şi anume, o fază haploidală sau haplofaza, reprezentată prin celule cu n cromozomi şi faza diploidă sau diplofaza cu celule cu 2n cromozomi. Trecerea de la faza haploidală la cea diploidă se face prin procesul de fecundaţie (F) iar prin meioză - diviziunea reducătoare (R) - se face trecerea de la diplofază din nou la haplofază.
În procesul de fecundaţie se întâlnesc două etape şi anume: plasmogamia (P) care constă în contopirea citoplasmelor gameţilor şi kariogamia (K) care constă în fuzionarea celor două nuclee. Celulele în care are loc desăvârşirea procesului sexuat prin kariogamie se numesc zeugite, iar acelea în care se produce diviziunea reducătoare - meioza (R) se numesc gonotoconte. Aceste două procese se produc în mod diferit şi anume la ciupercile inferioare din cl. Chytridiomycetes şi Plasmodiophoromycetes kariogamia urmează imediat după plasmogamie, rezultând un nucleu diploid al zigotului.
La aceste ciuperci, diplofaza este variabilă ca durată, după care, prin diviziunea reducătoare, se revine la faza haploidală.
La ciupercile mai evoluate din subîncrengăturile Ascomycotina şi Basidiomycotina, plasmogamia este separată de kariogamie în timp şi spaţiu, între ele intercalându-se o fază intermediară, denumită dikariofaza, ce este reprezentată prin miceliu secundar cu celule cu câte două nuclee.
Această fază intermediară - dikariofaza este foarte scurtă la ciupercile din clasele Zygomycetales şi Oomycetales, este de mai lungă durată la Ascomycotina, fiind reprezentată prin hifele ascogene, şi este de lungă durată la Basidiomycotina unde constituie miceliul secundar şi miceliul terţiar al carpoforilor.
Sexualitatea ciupercilor este caracterizată printr-o particularitate şi anume existenţa multipolarităţii. Din acest punct de vedere, ciupercile pot fi homotalice şi heterotalice, fenomen descoperit de Blakeslee la Mucoraceae. Autorul a demonstrat că ciuperca Sporodinia grandis este homotalică, pentru că poate forma zigoţi în culturi monosporale, ceea ce denotă că organele sexuale + şi - se găsesc pe acelaşi tal.

96
Alte ciuperci, ca Mucor mucedo, Mucor racemosus, Rhizopus nigricans, nu pot forma zigoţi. Pentru a se produce fructificaţii sexuate trebuie puse în contact două culturi provenite din spori de sexe diferite şi atunci zigoţii se formează la zona de confluenţă a miceliilor de sexe diferite. Altfel, ciupercile heterotalice formează numai sporangi ce reprezintă faza asexuată de dezvoltare.
Heterotalismul este un fenomen care nu se întâlneşte numai la ciupercile din subîncrengătura Zygomycotina, ci şi la cele din subîncrengăturile Ascomycotyna şi Basidiomycotina. Acest fenomen se datorează segregării caracterelor sexuale ce are loc atunci când se produce diviziunea reducătoare (meioza) în timpul germinării zigotului, în ască şi în basidie.
Pe lângă bipolaritatea sexuală a numeroase ciuperci se mai cunoaşte şi sexualitatea tetrapolară, aşa cum se întâlneşte la Schizophyllum commune. Acest fenomen nu trebuie confundat cu dimorfismul sexual al ciupercilor inferioare, singurele la care se mai manifestă deosebiri morfologice între sexe.
După Hartmann şi Gäumann, formele sexualităţii la ciuperci sunt de patru tipuri fundamentale, între care se deosebesc amfimixis, automixis, pseudomixis şi apomixis, fiecare dintre acestea cuprinzând mai multe forme:
1. Amfimixis este procesul ce se produce prin copularea a două celule sexuale neînrudite între ele; aici sunt cuprinse formele de procese sexuale: • Merogamia sau gametogamia ce se realizează prin copularea între doi gameţi, ca la ciupercile Synchytrium sau Olpidium. • Gametangiogamia sau gametangia, la care copularea se face între gametangii, ca la ciupercile din clasele Zygomycetes şi Oomycetes sau la ciupercile superioare din subîncrengătura Ascomycotina. • Hologamia la care întreg corpul vegetativ se transformă în organ de înmulţire, aşa cum se întâmplă la ciupercile inferioare.
2. Automixis este procesul ce se produce între două celule sexuale înrudite sau două nuclee sexuale. Aici există două posibilităţi şi anume: • Partenogamia la care copularea are loc între două celule ale filamentului ascogenal, ca la Ascobolus citrinus. • Autogamia la care are loc copularea celor două nuclee, ca la Humaria granulata.

97
3. Pseudomixis la care copularea se face între două celule vegetative; aceasta poate fi de mai multe feluri: • Pseudogamie sau somatogamie, la care are loc copularea a două celule vegetative ale miceliului ca la Corticium serum. • Pedogamia la care copularea are loc între o celulă matură şi una tânără, ca la Saccharomycetaceae. • Adelfogamie la care copularea se produce între celula mamă şi o celulă fiică, aşa cum se întâmplă la Zygosaccharomyces chevalieri.
4. Apomixis la care se produce dispariţia copulării şi celulele sexuale se dezvoltă mai departe pe cale vegetativă; aici sunt două situaţii: • Partenogeneza la care se produce dezvoltarea celulelor sexuale haploide. • Apogamia la care se realizează dezvoltarea celulelor sexuale diploide.
Pentru a ilustra cele mai obişnuite moduri de înmulţire la ciuperci, vom lua câteva cazuri, începând cu cele mai puţin evoluate până la cele superioare.
Astfel, la clasa Chytrydiomycetes, procesul sexual este o merogamie sau gametogamie şi are loc între doi gameţi mobili, identici ca formă, deci izogameţi. La genul Allomyces sunt anisogameţi, deci diferenţiaţi morfologic.
Aceste ciuperci inferioare sunt adaptate la viaţa acvatică sau trăiesc în medii interne ale gazdelor, unde umiditatea este asigurată.
La ciupercile mai evoluate care s-au desprins de mediul acvatic şi s-au adaptat la mediul terestru, se întâlneşte procesul sexual sub formă de gametangiogamie care reprezintă un proces de fecundaţie superior.
Astfel, la subîncrengătura Zygomycotina gametangiile identice (izogamentangii) conţin fiecare mai multe nuclee. Mai întâi are loc copularea între gametangii, iar ulterior nucleele fuzionează câte două; în urma fecundaţiei rezultă un zigot cenocitic cu nuclee diploide (Fig. 67).
Zigot
Fig. 67 – Înmulţire sexuată la Rhizopus sp.
cu formarea zigotului

98
La ciupercile din clasa Oomycetes din Familia Peronosporaceae, gametangii sunt diferenţiaţi din punct de vedere morfologic- heterogametangii. Gametangiul femel - oogonul conţine o masă de ooplasmă multinucleată, iar gametangiul mascul - anteridia, de asemenea, conţine mai multe nuclee. Gametangiile intră în contact printr-un canal de conjugare prin care anteridia îşi varsă conţinutul în oogon. În urma fecundaţiei rezultă un oospor ce poate fi cenocitic, ca la Cystopus bliti, sau conţine un singur nucleu diploid, ca la Pythium de Baryanum.
La ciupercile mai evoluate din subîncrengătura Ascomycotina, procesul sexual se produce tot între gametangii, în care gametangiul femel, numit ascogon, conţine o masă de citoplasmă şi numeroase nuclee, iar gametangiul mascul, denumit anteridie, este, de asemenea, multinucleat. Procesul de copulare între gametangii are loc prin intermediul trichoginului în două etape în care prima este plasmogamia (P), în urma căreia apare faza intermediară - dikariofaza, reprezentată prin miceliu secundar al hifelor ascogene. Kariogamia se produce mai târziu în asca tânără, prin unirea a două nuclee, când apare diplofaza.
Exemplul tipic al acestui mod de înmulţire îl constituie ciuperca Pyronema confluens la care procesul a fost studiat de către Harper (1900).
Treptat, la Ascomycetele superioare se observă o reducere progresivă, în special a anteridiei, rolul acesteia fiind luat de celulele vegetative, ca în cazul de la Taphrina (Fig. 68).
La ciupercile din subîncrengătura Basidiomycotina, ordinele
Ustilaginales, Uredinales şi Hymenomycetales, nu se mai diferenţiază
Fig. 68 – Înmulţirea sexuată la Taphrina deformans cu formarea ascelor libere.

99
gametangii pe miceliul vegetativ haploid şi copularea are loc între celule vegetative sub formă de somatogamie sau pseudogamie.
În concluzie, pe măsură ce ciupercile sunt mai evoluate procesul sexual se simplifică, organele sexuale se reduc şi sunt înlocuite prin celule vegetative care preiau funcţia sexuală, aşa cum se întâmplă la ascomicetele superioare şi, mai ales, la Basidiomycotina.
Simplificarea procesului sexual la ciuperci reprezintă o adaptare la viaţă în mediul terestru precum şi la cel saprofitic sau parasitic din natură.
În funcţie de evoluţie, rezultatul procesului de fecundaţie la ciuperci capătă aspecte morfologice diferenţiate şi denumiri corespunzătoare. Astfel, la cl. Chytridiomycetes şi Plasmodiophoromycetes, produsul de fecundaţie se numeşte zigozoospor, la subîncrengătura Zygomycotina poartă denumirea de zigot, la clasa Oomycetes este denumit oospor, la subîncrengătura Ascomycotina se numeşte ască, la subîncrengătura Basidiomycotina este basidia.
2.8.6. Alternanţa de faze la ciuperci
Haplofaza, dikariofaza şi diplofaza se succed în cursul ciclului de dezvoltare într-o alternanţă de faze care este specifică anumitor grupe de ciuperci. Schema generală a acestei alternanţe, reprezentată după Gäuman, se prezintă după cum urmează. Astfel, ciupercile din subîncrengăturile Mastigomycotina şi Zygomycotina trăiesc în haplofază, sunt deci haplobionte; dikariofaza este aici absentă.
Ciupercile din subîncrengătura Ascomycotina trăiesc, în cea mai mare parte, în haplofază dar prezintă şi dikariofază ce este reprezentată prin filamente ascogene; la acest grup, diplofaza este de scurtă durată.
La ciupercile din subîncrengătura Basidiomycotina, durata dikariofazei este preponderentă asupra haplofazei; acestea se numesc ciuperci haplo-dikariobionte. Tot în această grupă se mai încadrează ciupercile din ordinul Taphrinales.
O categorie aparte o constituie grupa ciupercilor haplodiplobionte, la care dikariofaza este foarte scurtă, iar diplofaza este preponderentă asupra haplofazei. Acest tip de alternanţă de faze se întâlneşte la unele Saccharomycetaceae.

100
Ciupercile din subîncrengătura Deuteromycotina, neavând ciclul biologic complet, sunt lipsite de faza de înmulţire sexuată. Unele, fiind mai atent studiate, sunt trecute în unităţi sistematice superioare, după ce li se descoperă ciclul complet.
2.8.7 Nutriţia ciupercilor Ciupercile sunt organisme vegetale lipsite de pigment trofic, deci nu
sunt capabile de a face sinteza substanţelor nutritive şi, din această cauză, se dezvoltă pe substrat organic viu sau neviu, fiind heterotrofe.
Corpul ciupercilor este constituit din macroelemente, între care sunt: carbonul, oxigenul, hidrogenul, azotul, fosforul, sulful, siliciul, potasiul, sodiul, calciul, magneziul, fierul, manganul etc. De asemenea, în compoziţia corpului lor se mai găsesc şi microelemente ca: borul, florul, aluminiul, titanul, cobaltul, nikelul, zincul ce au rol de catalizatori ai proceselor de metabolism.
Unele elemente, ca mercurul, cuprul, plumbul, argintul etc. sunt nocive pentru ciuperci servind ca baze toxice la prepararea fungicidelor.
Principala sursă de carbon o constituie glucidele şi zaharurile, dintre care, în special, sunt utilizate hexozele şi pentozele. Unele specii de Fusarium pot folosi şi alcoolii ca: alcoolul etilic, metilic, propilic sau butilic.
Alte ciuperci pot descompune unele polizaharide ca: amidonul, celuloza sau hemiceluloza sau pot folosi şi carbonul din unele lipide.
Ca sursă de azot, ciupercile pot utiliza azotul atmosferic, azotul amoniacal, azotul nitros şi nitric sub formă de nitriţi şi nitraţi ca şi azotul organic sub formă de amide, amine, aminoacizi, peptone şi proteine.
Între ciupercile ce pot utiliza azotul atmosferic sunt: Aspergillus niger, Penicillium glaucum, Alternaria tennuis, Rhizopus nigricans, Cladosporium herbarum etc.
Azotul amoniacal sub formă de săruri de amoniu este folosit de unele Saccharomycetaceae, Mucoraceae, precum şi de Botrytis cinerea sau de Aspergillus niger dar unele Mucoraceae pot folosi şi azotul nitric sub formă de nitraţi.

101
În ceea ce priveşte azotul organic, pot fi utilizate de către ciuperci fie ureea, acidul uric sau guanidina, dintre compuşii simpli sau aminoacizii, peptonele şi albuminele sub formă de compuşi mai complecşi.
Ca surse de fosfor, sunt utilizaţi fosfaţii din sol sau surse organice ca lecitina şi fitina.
Sulful este folosit ca produs mineral sub formă de sulfaţi de fier, zinc, amoniu, sodiu etc. sau ca sulf organic din produse ca cisteină, cistină, glutation, peptone etc.
Elementele necesare ciupercilor trebuie utilizate în concentraţii optime, dozele prea mari fiind nocive iar cele prea mici nefiind suficiente pentru desfăşurarea normală a metabolismului.
Ciupercile mai au nevoie pentru edificarea organismului lor şi de unele substanţe stimulatoare de creştere ce intră în compoziţia mediilor nutritive în cantităţi foarte mici, între care sunt vitaminele, hormonii, auxinele, numite şi factori auximoni.
Unele ciuperci sunt capabile să sintetizeze unele vitamine, ca B, B1, B2, B6, C, K, D etc., ele dezvoltându-se bine pe medii artificiale. Alte ciuperci, care nu-şi pot sintetiza aceste vitamine, nu se dezvoltă decât dacă în mediile nutritive se adaugă vitaminele respective.
După modul lor de nutriţie, ciupercile se împart în patru categorii şi anume: • Ciupercile parazite obligate duc o viaţă strict parazitară, neputându-se
dezvolta pe substrate nevii şi nu pot fi, deci, cultivate pe medii artificiale. Din această categorie fac parte Peronosporaceele, Erysiphaceele şi Uredinalele.
• Ciupercile saprofite facultative duc, de obicei, o viaţă parazitară, dar, în anumite condiţii, pot trăi şi pe substraturi nevii, ca în cazul speciilor: Phytophthora infestans, Pythium de baryanum, Endostigme inaequalis.
• Ciupercile parazite facultative se comportă, în mod obişnuit, ca saprofite şi devin parazite în anumite condiţii, cum sunt diferite specii ale genurilor: Fusarium, Verticillium, Cladosporium, Alternaria etc.
• Ciupercile saprofite obligate se dezvoltă exclusiv pe substraturi nevii, cum sunt numeroase specii de Mucoraceae, Aspergillaceae, unele specii din subîncrengătura Ascomycotina şi din Basidiomycotina.

102
2.8.8. Originea şi evoluţia parazitismului la ciuperci
Prin fenomenul de parazitism se înţelege convieţuirea nearmonioasă între două organisme ce sunt unul faţă de altul în stare de dezechilibru biologic, organismul parazit fiind acela ce ia de la gazda sa substanţele nutritive de care are nevoie.
Cu cât relaţiile de parazitism sunt mai evoluate, cu atât raporturile dintre cei doi parteneri sunt mai strânse.
O prezentare sintetică a problemei parazitismului a fost făcută de Kuprevici, în lucrarea sa : “Fiziologia plantei bolnave în legătură cu problemele generale de parazitism”.
Nutriţia saprofită se consideră a fi cea iniţială, iar formele parazite derivă din cele saprofite.
Apariţia parazitismului s-a produs prin modificarea şi adaptarea la modul de hrănire pe substraturi diferite, pe baza variabilităţii sistemului de fermenţi, secretaţi de ciuperci.
La ciupercile saprofite, sistemul de fermenţi este mai bogat, în timp ce la cele parazite sistemul enzimatic este mai restrâns şi mai specializat.
În prezent, se consideră că, la ciuperci, treptele de trecere de la saprofitism la parazitism sunt următoarele: saprofite obligate - parazite facultative - saprofite facultative - parazite obligate.
În acest cadru, se consideră că, în scara parazitismului, ciupercile cu o specializare largă şi cu stadii saprofite în ontogeneză sunt forme inferioare, în timp ce paraziţii obligaţi, cu o specializare îngustă, sunt cele superioare.
Creşterea filamentelor miceliene este strict apicală, comparativ cu creşterea intercalară, cunoscută la alte organisme filamentoase. Apexul hifelor este bogat în vezicule ce conţin hidrolaze şi sintezaze, necesare creşterii în lungime. Acumularea apicală a veziculelor implică o circulaţie acropetală a conţinutului citoplasmatic, afluxul de elemente nutritive realizându-se din regiunile mature în zonele de creştere.
Dacă veziculele purtătoare de enzime de creştere sunt prea numeroase, apar ramificaţii laterale la câţiva μm distanţă de apex. La un moment dat, apare o concurenţă trofică între axul principal şi ramificaţiile principale, secundare sau terţiare, dată, în principal, de conţinutul mai scăzut al mediului în elemente nutritive.

103
2.8.9. Cerinţe faţă de mediu
Temperatura. Majoritatea ciupercilor sunt mezofile, cu un optim de
creştere situat între 25 - 35oC. Unele specii sunt termotolerante (20 - 50oC). Temperatura maximă limită de dezvoltare este de 60 - 62oC.
Ciupercile termofile joacă un rol important în procesele de degradare a compostului.
PH-ul. Majoritatea ciupercilor pot să se dezvolte într-o zonă de pH cuprins între 4,5 şi 8. Optimul fiind situat între 5,5 şi 7,5.
Există specii de ciuperci acidofile cu creştere la un pH inferior lui 4,5 sau acido-tolerante.
Aeraţia.Unele specii de ciuperci sunt microaerofile, altele sunt strict anaerobe, dar majoritatea sunt aerobe.
2.8.10. Sistematica ciupercilor
Ciupercile cunoscute şi descrise până acum pe plan mondial
depăşesc cifra de 100.000 de specii şi mereu apar contribuţii micologice noi, care sporesc acest număr.
Unităţile sistematice cărora aparţin ciupercile încep de la regnul Fungi, Încr. Eumycota şi continuă cu subîncrengătura, clasa, ordinul, familia, genul şi specia. În afară de acestea, la ciupercile parazite mai sunt formele specializate, rasele fiziologice sau biotipurile, acestea din urmă deosebindu-se numai prin unele caractere biologice în legătură strânsă cu specia, cultivarul sau liniile plantelor gazde, obţinute în procesul de ameliorare.
În decursul timpului au fost elaborate mai multe sisteme de clasificare ale unor autori, dintre care menţionăm: Greis (1943), Moreau (1953), Gäumann (1964), Goidanich (1963-1964), Alexopoulos (1966), Robert şi Bouthroyd (1972).
În prezenta lucrare se foloseşte sistemul de clasificare elaborat de Ainsworth, Sparow şi Sussman (1973), după care ciupercile se încadrează în Încr. Eumycota şi subîncr. Mastigomycotina, Zigomycotina, Ascomycotina, Basidiomycotina şi Deuteromycotina.

104
Capitolul 3
INFLUENŢA FACTORILOR ECOLOGICI ASUPRA MICROORGANISMELOR
3.1. Influenţa pH-ului Majoritatea microorganismelor care alcătuiesc micropopulaţia solului
preferă valori de pH cât mai aproape de neutru. Aşa se explică faptul că în solurile cu reacţie neutră se întâlneşte cea mai bogată, mai variată şi, în acelaşi timp, mai valoroasă microfloră.
Din punct de vedere al preferinţei faţă de anumite valori de pH, microorganismele se împart în mai multe grupe: • Microorganismele indiferente. În această categorie intră unele bacterii şi majoritatea ciupercilor care, în culturi pure, se dezvoltă în limite largi de pH. Ex. : Penicilium aculeatum preferă valori de pH cuprinse între 2 şi 6, iar Trichoderma viride între 2 şi 8. • Microorganismele neutrofile preferă valori de pH cât mai aproape de neutru sau uşor bazice. În această grupă intră cel mai mare număr de microorganisme şi cele mai importante specii şi grupe pentru fertilitatea solului. Ex: genurile Azotobacter, Nitrosomonas, Nitrosococcus, Nitrobacter, Nitrococcus, Cytophaga, Bacillus, Streptomyces etc. • Microorganismele acidofile. Acestea au dezvoltarea maximă la valori de pH 2-3 şi cuprind specii bacteriene care oxidează sulful (ex. Thiobacillus thiooxidans) şi fierul (ex. Thiobacillus ferrooxidans). • Microorganismele bazofile au cea mai bună dezvoltare la valori de pH peste 8; în această categorie intră bacteriile ce descompun ureea ( ex. Bacillus ureolitici).
Ca o aplicaţie practică, corectarea reacţiei solurilor acide prin calcarizare stimulează activitatea microorganismelor valoroase şi acest efect este cu atât mai puternic cu cât solul neamendat a avut valori mai scăzute de pH.
Ciupercile microscopice sunt microorganisme care suportă uşor (chiar preferă) reacţia acidă a solului, concurând, astfel, toate celelalte

105
microorganisme din solurile acide. În această situaţie, pH-ul solului este un factor de reglaj al competiţiei pentru hrană.
De asemenea, s-a observat că între conţinutul de substanţe nutritive şi reacţia solului există o corelaţie dovedită prin schimbul de ioni. Acest raport de schimb între ionii de hidrogen produşi de rădăcinile plantelor este echivalent cu bazele absorbite prin schimb cu mineralele argiloase şi coloizii organici. În cadrul descompunerii substanţelor organice, procesul de dezalcalinizare şi formare a acizilor duce la acidifierea solului, care este contracarată prin fenomene de mineralizare, degradare şi unele măsuri de fertilizare. Acidifierea solurilor produce o restructurare a grupelor de microorganisme, favorizând ciupercile şi bacteriile anaerobe.
3.2. Influenţa temperaturii Studiul influenţei temperaturii asupra microorganismelor prezintă
două aspecte importante - cunoaşterea temperaturile cardinale, adică minima (Tm), optima (To) şi maxima (TM), care permit creşterea şi dezvoltarea microorganismelor; - cunoaşterea temperaturile letale, care impiedică dezvoltarea sau produce moartea microorganismelor.
Temperatura este un factor foarte important pentru cantitatea de humus formată în solurile din regiunile umede cu climă temperată. Referindu-ne la dinamica sintezei şi descompunerii humusului, s-a constatat că odată cu scăderea temperaturii scade şi activitatea microbiotei solului.
Limitele optime ale temperaturii pentru microorganismele din sol variază între 20 şi 30ºC, iar din punct de vedere al oscilaţiilor de temperatură bacteriile au un grad foarte ridicat de adaptabilitate.
Se ştie că fiecare specie microbiană are o anumită temperatură optimă de creştere şi anumite limite între care se desfăşoară în condiţii normale viaţa acesteia. În funcţie de adaptarea la factorul termic, microorganismele pot fi: • Microorganisme psihrofile, cu o temperatura optimă de creştere inferioară valorii de 20ºC, au drept caracteristice urmatoarele valori de temperatură: Tm= –10 .... +5ºC; To = 10 – 15ºC şi TM = 20 – 25ºC.

106
Din această grupă fac parte majoritatea bacteriilor din genurile Pseudomonas, Flavobaterium, Alcaligenes, Acromobacter, Staphilococcus şi Salmonella; drojdiile din genurile Candida şi Rhodothorula şi fungii filamentoşi din genurile Alternaria, Cladosporium şi Botrytis.
Bacteriile adaptate la temperaturi scăzute prezintă anumite particularităţi: conţinut ridicat în lipide care intră în structura peretelui celular şi a membranei citoplasmatice; conţinut ridicat în acizi graşi nesaturaţi; mobilitate ridicată; prezintă capacitatea de a produce enzime care sunt mai active la temperaturi scăzute. • Microorganisme mezofile. Temperatura optimă (To) este cuprinsă între 25 şi 40ºC şi nu se dezvoltă în afara limitelor de 15 - 45ºC. În acestă grupă sunt cuprinse marea majoritate a bacteriilor, actinomicetelor şi a drojdiilor fermentative. Ciupercile filamentoase au temperatura de creştere în jurul valorii de 26ºC, iar cea de sporulare de 30ºC. • Microorganisme termofile sunt reprezentate de specii al căror metabolism este intens la temperaturi relativ ridicate. Au drept caracteristice urmatoarele valori de temperatură: Tm = 20 - 30ºC; To = 40 – 60ºC şi TM = 70 – 90ºC. Între bacteriile din această grupă întâlnim bacteriile amonificatoare, celulolitice (Clostridium termocellulolyticum Clostridium termosaccharoliticum, Bacillus cellulosae hydrogenicus şi Methylomonas methanica), amilolitice, sulfato-reductoare şi altele. Dintre fungi, amintim Aspergillus fumigatus şi Penicillium duponti.
Trebuie subliniat că această clasificare este relativă deoarece multe microorganisme se dezvoltă bine în limite largi de temperatură.
Rezistenţa microorganismelor la temperaturi letale sau termorezistenţa acestora se referă la posibilitatea celulelor microbiene de a rezista pentru o perioadă mai scurtă sau lungă de timp la temperaturi de 90 - 100ºC sau peste 100ºC.
3.3. Influenţa apei Activitatea biologică a unui sol tinde să se diminueze odată cu
scăderea conţinutului în apă, dar contrar celor observate la plante, punctul de ofilire (pF = 4,2) nu constituie un prag indicator al încetării activităţii microbiene. Se apreciază că limita superioară oricărei activităţi biologice

107
corespunde la pF= 5,5-5,6. Uscarea solului peste aceste valori antrenează moartea tuturor microorganismelor.
Această particularitate a microorganismelor faţă de plantele superioare se explică, în primul rând, prin presiunea osmotică mai ridicată din celulele microbiene; în al doilea rând, bacteriile din sol nu pierd apă prin transpiraţie ca plantele superioare, iar în al treilea rând, suprafaţa de absorbţie la microorganisme faţă de masa protoplasmatică egală este mai mare ca la plante.
Moartea microorganismelor la umiditate scăzută este determinată de mai mulţi factori, ca durata şi nivelul uscării, tipul microorganismului, tipul de sol şi structura solului.
În funcţie de nevoile de apă, microorganismele se împart în mai multe grupe: • Microorganismele hiperxerofile sunt germeni al căror prag este superior valorii pF = 4,9. Aici, majoritatea microorganismelor sunt cele amonificatoare, amilolitice şi cele care degradează substanţele vegetale complexe. • Microorganismele xerofile sunt cele al căror prag este cuprins între pF = 4,9 şi 4,2. Aici se includ germenii care degradează celuloza, cei care oxidează sulful, o parte dintre microorganismele amilolitice, amonificatorii ureei şi cei care produc nitrificarea sulfatului de amoniu. • Microorganismele hidrofile includ germeni care au pragul pF = 2. Aici se găsesc agenţi care descompun carboximetilceluloza ce fixează azotul atmosferic precum şi o parte dintre nitrificatorii sulfatului de amoniu.
Existenţa pragurilor diferite ridică probleme de ordin practic, deosebit de importante pentru agricultură. Astfel, una dintre consecinţe este predominarea selectivă a unor procese microbiene asupra altora. Aceasta se întâmplă în cazul umidităţii situate în vecinătatea punctului de ofilire, când predominarea proceselor de celuloliză asupra amonificării are drept urmare imobilizarea totală sau parţială a azotului mineral.
De asemenea, în cazul umidităţii scăzute predomină procesele de amonificare, fapt ce antrenează o acumulare de azot amoniacal, remarcată în solurile în curs de uscare.

108
Diferitele grupe de microorganisme au un grad variat de rezistenţă faţă de uscăciune. Astfel, ciupercile şi bacteriile sunt mai sensibile, comparativ cu actinomicetele.
3.4. Influenţa energiei radiante Radiaţiile electromagnetice au un spectru larg, includ radiaţiile
herziene, infraroşii, vizibile, ultraviolete X şi Y ce sunt caracterizate fiecare printr-o anumită lungime de undă. Ele au dublu caracter, de fenomen ondulatoriu şi de fascicol discontinuu de mici cuante de energie.
Iradierea reprezintă un bombardament cu fascicole de cuante de energie iar efectele ei asupra materialului iradiat sunt determinate de absorbţia de către acestea a energiei radiaţiilor şi depind de energia cuantei şi de natura materialului absorbant.
Radiaţiile herziene şi infraroşii cu lungimea de undă mare ( >12.000 Å) au un conţinut de energie slab, astfel încât sunt incapabile să modifice chimic substanţele pe care le absorb şi, ca atare, energia lor este transformată în căldură.
Radiaţiile cuprinse între 12.000 Å (infraroşu apropiat) şi 2.000 Å (ultraviolet îndepărtat) au un conţinut energetic suficient de ridicat, pentru a produce modificări fotochimice ale moleculelor care sunt absorbite.
Radiaţiile cu lungime de undă sub 2.000 Å (X, Y şi cosmice) au un conţinut energetic atât de ridicat, încât provoacă ionizarea moleculelor pe care le întâlnesc, de unde şi denumirea de radiaţii ionizante.
Efectul biologic al radiaţiilor depinde de lungimea lor de undă şi de doza de iradiere. În funcţie de aceşti doi factori, iradierea poate avea două efecte distincte şi anume efectul letal asupra microorganismelor şi efectul mutagen, ce constă în modificarea unor caractere genotipice.
Radiaţiile luminoase acţionează asupra celulelor prin intermediul unor reacţii fotochimice cu constituenţii celulari. Dintre acestea sunt bine cunoscute cele implicate în procesul de fotosinteză şi în fenomenele de fototoxie, ce se întâlnesc la microorganismele fototrofe. Lumina solară directă este bactericidă, datorită nu atât radiaţiilor ultraviolet, cât încălzirii prin radiaţiile infraroşii şi deshidratării celulelor expuse. Radiaţiile

109
ultraviolete provenite de la soare sunt reţinute, în majoritate, de stratul de ozon din partea superioară a atmosferei.
Radiaţiile ultraviolete au o acţiune antimicrobiană netă, ale cărui efect depinde de intensitatea şi durata iradierii.
În cazul unei iradieri slabe şi de scurtă durată, se produc fie alterări limitate la nivelul materialului genetic, care se traduc prin apariţia de mutaţii, materializate prin pierderea unui caracter de lizogenie al celulei bacteriene sau o inactivare reversibilă a unor procese metabolice. Acestea au ca efect blocarea temporară a multiplicării celulare, deoarece, în urma unei iradieri intense, în celulă survin modificări profunde, ireversibile, cu efect letal.
Radiaţiile ultraviolete cuprinse între 2300 şi 2800 Å sunt cele mai active din punct de vedere biologic. Spectrul de acţiune antimicrobiană al acestora seamănă foarte mult cu spectrul de absorbţie al UV de către ADN, ceea ce a sugerat ipoteza cunoscută sub numele de „teoria ţintei”. Conform acesteia, efectul iradierii se exercită, în cea mai mare măsură, prin intermediul ADN-ului. Radiaţiile lovesc direct acest constituent celular şi determină reacţii fotochimice selective cu bazele din structura lui.
Radiaţiile corpusculare, ca razele gamma, dar şi radiaţiile electromagnetice Y şi X produc efecte legate de bombardarea generală a celulei. Când întâlnesc un microorganism, particulele active ”lovesc” succesiv o serie de atomi din structura sa, desprinzând electroni de pe particulele respective. Se creează, în felul acesta, coloane de ionizare intensă, ce determină alterarea sau distrugerea unor substanţe esenţiale pentru celulă şi blocarea a diferite sinteze sau inhibarea procesului de diviziune.
3.5. Activitatea microbiană a diferitelor soluri Variaţia microflorei în funcţie de factorii mai sus amintiţi duce la o
diferenţiere a activităţii biologice a diferitelor tipuri de sol. În ţara noastră, cercetări de peste 20 de ani făcute de Papacostea au
scos în evidenţă existenţa în solurile româneşti a unor ”microorganisme indicatoare”, care sunt caracteristice diferitelor tipuri de sol şi care au permis din acest punct de vedere clasificarea solurilor în trei grupe.

110
1. Soluri cu Mucor rammanianus - de la cele foarte acide (de sub pajiştile alpine) până la cele acide, brune (de sub pădurea de molid). 2. Soluri cu Pseudomonas acidophilus şi Pseudomonas aureofaciens, cum sunt solurile podzolite pseudogleizate. 3. Solurile cu Pseudomonas lemonieri şi Pseudomonas aureofaciens, care sunt formate în climat mai uscat, în zone cu precipitaţii între 400 şi 600 mm pe an.
3.6. Profilul microbian al solului Având în vedere influenţa pe care aeraţia o exercită asupra activităţii
biologice din sol, se constată că numărul genurilor de specii aerobe scade odată cu adâncimea, având loc, în paralel, o creştere a numărului de specii anaerobe şi/sau facultativ anaerobe. Nu putem însă atribui scăderea numărului de germeni numai aeraţiei; această distribuţie reflectă, în mare măsură condiţiile de nutriţie pe care microorganismele le găsesc în sol şi care sunt mai bine satisfăcute în orizonturile de suprafaţă, datorită cantităţii mai mari de humus şi aprovizionării mai frecvente cu resturi organice.

111
Capitolul 4
INTERRELAŢIILE ECOLOGICE ÎNTRE ORGANISME
4.1. Interrelaţiile dintre populaţiile de microorganisme
Răspândirea foarte largă a microorganismelor în natură face ca, frecvent, să se găsească, în acelaşi habitat, mai multe specii între care se stabilesc interrelaţii de complexitate variată.
Aceste interrelaţii între două populaţii microbiene pot fi: • indiferente sau neutrale; • pozitive sau sinergice; • negative sau antagonice.
I. Neutralismul poate fi definit ca relaţie de indiferenţă între două sau mai multe specii, datorită distanţei mari între populaţiile respective sau deosebirilor mari în ceea ce priveşte exigenţelor nutritive, fapt ce le face să nu intre în competiţie pentru acelaşi substrat.
II. Interrelaţiile sinergice sunt relaţii benefice care cuprind o mare varietate de forme de la comensalism (sau metabioză) şi mutualism (sau simbioză nutriţională) până la simbioza propriu-zisă.
1. Comensalismul microbian sau metabioza este caracterizat prin creşterea împreună a două specii de microorganisme într-un tip de relaţie, în care una profită de asociere, pe câtă vreme cealaltă, în aparenţă, nu profită, dar nici nu este influenţată negativ. Interrelaţiile comensalice sunt, la rândul lor, de mai multe feluri, după modul de acţiune a unuia dintre parteneri: • Un microorganism produce un metabolit sau un factor de creştere esenţial pentru un alt microorganism.
Astfel, bacteriile care oxidează nitriţii la nitraţi (genurile Nitrobacter, Nitrospira, Nitrococcus) nu se pot dezvolta în sol decât în prezenţa bacteriilor care oxidează amoniacul la nitriţi (genurile Nitrosomonas, Nitrosococcus, Nitrosospira şi Nitrosolobus) şi care le furnizează substanţa necesară nutriţiei. • Un anumit microorganism poate degrada sau neutraliza un factor microbian activ asupra unei alte specii de microorganism.

112
Biodegradarea bacteriană şi fungică a fenolilor, a acidului benzoic sau a antibioticelor face posibilă dezvoltarea microorganismelor sensibile la acţiunea toxică a acestor substanţe. Unele microorganisme pot să sintetizeze şi să elimine în mediu enzime (panicilinaza) care degradează anumite antibiotice, protejând, în felul acesta, microorganismele penicilino-sensibile. • Un anumit microorganism modifica proprietăţile fizice şi chimice ale habitatului şi creează condiţii de creştere pentru microorganismul asociat.
Spre exemplu, bacteriile aerobe consumă din mediu oxigenul, creând, în acest fel, condiţii favorabile microorganismelor anaerobe. O parte dintre microorganisme alterează mediul din punct de vedere fizic, creând condiţii de creştere pentru alte specii.
Drojdiile osmofile, ce cresc în soluţii concentrate de zahăr, modifică aceste soluţii prin fermentaţii sau oxidări, permiţând, astfel, dezvoltarea microorganismelor neosmofile sau mai puţin osmotolerante. • Un anumit microorganism poate modifica substratul nutritiv, făcându-l accesibil altui microorganism.
Spre exemplu, relaţiile comensale între bacteriile celulozitice şi speciile genului Azotobacter. Primele hidrolizează celuloza până la compuşi intermediari (monozaharide, acizi organici, alcooli), asimilabili de către Azotobacter sp.
2. Mutualismul sau simbioza nutriţională constă în exercitarea de influenţe favorabile bilaterale între specii diferite, ce se dezvoltă simultan pe acelaşi substrat. Acest tip de interrelaţie se deosebeşte de simbioza propriu-zisă prin fapul că nu este obligatorie, permanentă.
El prezintă un tip de interrelaţie nutriţională între două specii de microorganisme, care au nevoie una de alta pentru realizarea unor metaboliţi esenţiali, pe care niciuna dintre specii nu-i poate sintetiza.
Astfel, Mucor ramanianum şi Rhodotorula rubra nu se pot dezvolta separat pe medii lipsite de tiamină (vitamina B1). Dacă sunt însă cultivate împreună, nu necesită adăugarea vitaminei B1 în mediu, deoarece Mucor sp. sintetizează o parte din vitamina B1 şi anume nucleul tiazolic, iar Rhodotorula sp. cealaltă parte – nucleul pirimidinic.
Un alt exemplu este cel referitor la interrelaţiile dintre Lactobacillus arabinosus şi Streptococcus faecalis pe un mediu lipsit de fenilamina (indinspensabilă pentru Lactobacillus) şi acid folic

113
(indinspensabil pentru Streptococcus), când fiecare bacterie sintetizează factorul de creştere necesar celeilalte.
3. Simbioza microbiană Două sau mai multe tipuri de microorganisme, ce se găsesc împreună
în acelaşi mediu, pot avea activităţi combinate sinergic, diferite cantitativ sau calitativ de activităţile aceloraşi organisme, crescute separat pe mediul respectiv.
În urma acestui tip de asociere pot fi sintetizaţi unii compuşi organici cu structură complexă sau pot fi degradate unele substanţe complexe. Fenomene de simbioză pot fi observate în asociaţii de microorganisme de tipul lichenilor, la care se formează anumiţi acizi organici, pigmenţi şi corpi reproducători complecşi, care nu pot fi elaboraţi de niciunul dintre cei doi parteneri consideraţi separat.
III. Relaţii de tip antagonic. În cadrul acestor relaţii, cunoscute sub numele de antagonism microbian sau de relaţii antagonice, una dintre specii se comportă ca agresor şi profită de pe urma asociaţiei cu cealaltă care este prejudiciată. Un microorganism sau o populaţie de microorganisme poate avea o acţiune negativă, de inhibare sau de distrugere a altui microorganism ori micropopulaţii, prin diferite mecanisme: • modificarea condiţiilor de mediu;
De exemplu, din activitatea microflorei solului pot rezulta cantităţi mari de CO2, care inhibă dezvoltarea altor microorganisme sensibile (Rhizobium sp.) şi modifică pH-ul mediului (devine acid). Astfel, sunt eliminate speciile sensibile la pH acid, cum ar fi cele din genurile Azotobacter, Nitrosomonas, Nitrobacter şi unele actinomicete. Pe de altă parte, scăderea potenţialului oxido-reducător elimină din mediu numeroase specii aerobe.
Ca alt exemplu amintim apariţia alcoolului etilic sau a acidului lactic sau butiric în timpul unor procese fermentative, compuşi care provoacă dispariţia celorlalte micoorganisme din mediu fermentat. • competiţie;
În natură, substanţele nutritive se găsesc în cantităţi limitate, iar microorganismele, adesea, intră în competiţie pentru procurarea acestora. Acest tip de interrelaţie poartă numele de competitivitate nutritivă. Când

114
mediul este bogat în substanţe nutritive dar este limitat ca spaţiu, poate să apară şi o competiţie pentru spaţiu.
Capacitatea competitivă a microorganismelor este determinată, în special, de viteza lor de creştere. Astfel, microorganismele cu o creştere rapidă epuizează disponibilităţile nutritive ale mediului şi împiedică dezvoltarea speciei cu creştere lentă. Spre exemplu, unele bacterii nesporulate împiedică dezvoltarea actinomicetelor. • elaborarea unor substanţe toxice sau inhibatoare;
Cele mai multe relaţii antagonice se manifestă prin producerea şi eliberarea în mediu de către o specie microbiană (specie antagonistă) a unor substanţe inhibatoare sau toxice pentru o singură specie sau un grup de specii, situate în imediata lor apropiere. Efectele acestor substanţe pot fi bacteriostatic sau fungistatic şi bactericid sau fungicid bacteriolitic.
În mediile naturale, antagonismul prin antibiotice este foarte frecvent. Microflora solului conţine numeroase microorganisme producătoare de antibiotice: actinomicete (Streptomyces, Nocardia, Micromonospora), bacterii (Bacillus şi Pseudomonas) şi fungi (Penicillium, Aspergillus, Trichoderma, Fusarium, Sclerotinia). • parazitism;
Parazitismul este acel tip de relaţie în care un microorganism este distrus, lizat, de un alt microorganism, care se dezvoltă în interiorul şi pe seama sa. În această relaţie, parazitul este întotdeauna mai mic decât gazda şi rămâne viu cel puţin o scurtă perioadă de timp, multiplicându-se în celula microorganismului gazdă.
Exemplu clasic de parazitism este infecţia unei bacterii cu un bacteriofag virulent, care are ca efect final liza celulei bacteriene. De asemenea, se cunosc bacterii micolitice, care au capacitatea de a elibera chitinaza, o enzimă ce distruge peretele celular al unor ciuperci microscopice, producând moartea acestora. Multiplicarea microorganismului agresor are loc pe seama constituenţilor celulari ai gazdei sale. • prădătorism.
Prădătorismul este o manifestare de agresiune, în care unele microorganisme mobile, lipsite de perete celular rigid, captează, ingerează şi

115
digeră microorganisme de dimensiuni mai mici. Ca exemplu tipic avem protozoarele, care se hrănesc cu bacterii vii.
În lumea microorganismelor, prădătorismul este limitat numai la speciile capabile de fagocitoză. Bacteriile, fungii şi algele sunt excluse din cauza peretelui celular rigid, care împiedică înglobarea altor microorganisme sau macromolecule.
4.2. Interrelaţiile dintre plantele superioare şi microorganismele
din sol Plantele, dezvoltându-se în mediul lor natural (solul) intră în
concurenţă pentru hrană cu microorganismele. Încă din 1895, Caron a emis ideea că prezenţa bacteriilor în sol ar
putea să aibă un efect favorabil asupra plantelor de cultură şi invers, că plantele, la rândul lor, ar avea o acţiune favorabilă asupra florei bacteriene din sol.
4.2.1. Influenţa sistemului radicular al plantelor asupra microflorei
solului Pentru a putea prezenta relaţiile care se stabilesc între sistemul
radicular al plantelor şi microbiota din sol este necesar să definim câteva noţiuni: spermatosferă şi rizosferă.
Spermatosfera este zona de sol care înconjoară seminţele în curs de germinare. Numărul microorganismelor din această zonă (în special bacterii) este mult mai mare, comparativ cu zona cu seminţe aflate în repaus germinativ, deoarece, microorganismele utilizează substanţele eliminate de seminţe în timpul germinării lor şi produc, la rândul lor, fitohormoni, care stimulează germinarea şi apoi creşterea plantei. Ulterior, spermatosfera se transformă în rizosferă.
Rizosfera este zona de sol care înconjoară rădăcinile şi este influenţată de acestea din punct de vedere fizic, chimic şi biologic, datorită acţiunilor specifice dintre microorganismele solului şi plantele superioare.
Analiza probelor de sol recoltate de la diferite distanţe faţă de rădăcină şi chiar de la suprafaţa rădăcinii a demonstrat influenţa favorabilă pe care o

116
are sistemul radicular asupra bacteriilor. Existenţa zonalităţii în răspândirea microorganismelor din sol, în jurul rădăcinilor plantelor, a determinat pe cercetători să împartă rizosfera în mai multe zone.
Astfel, Berezova (1950) distinge în cadrul rizosferei trei zone care se deosebesc între ele prin densitatea microbiană dar şi prin structura speciilor de microorganisme: • Prima zonă, numită rizoplan, rizosfera internă (rizosfera propriu-zisă), zonă radiculară sau zonă radicelară, are o rază de 0-0,5 cm şi cuprinde microorganisme de la suprafaţa rădăcinilor; • A doua zonă, numită zona periradiculară, cuprinde microorganismele ce se găsesc la o distanţă de 0,5 cm faţă de rădăcină; • A treia zonă, numită rizosfera externă, zona rizosferică sau edafosferă, cuprinde microorganisme ce se găsesc la o distanţă mai mare de 0,5 cm faţă de rădăcină.
Această împărţire înfăţişează aspectul cantitativ al efectului de rizosferă dar influenţa rădăcinilor asupra microflorei din sol se manifestă şi prin modificarea raportului între grupele de microorganisme, ceea ce duce la modificarea echilibrului microbian din sol. Prezenţa mai multor microorganisme (în special bacterii) în rizosferă decât în solul nerizosferic (fără rădăcini) se numeşte efect rizosferic. Intensitatea efectului rizosferic la bacterii se exprimă prin raportul R/S (rizosferă/sol nerizosferic). La actinomicete, ciuperci, alge şi protozoare R/S este între 2 şi 3.
Concentraţia speciilor bacteriene din rizosferă poate fi influenţată de mai mulţi factori, dintre care amintim: • specia plantei; • conţinutul solului în substanţe nutritive; • faza de vegetaţie; • distanţa dintre suprafaţa rădăcinii şi particulele de sol; • zonele rădăcinii.
Creşterea plantelor provoacă schimbări în echilibrul dintre grupe de nutriţie. Sunt stimulate microorganisme cu cerinţe nutritive simple şi cele care folosesc aminoacizi; sunt inhibate cele ce depind de substanţe mai complexe, conţinute de extractul de sol. Cook şi Lockhead (1959) constată, de asemenea, că numărul de bacterii care necesită substanţe biologice active

117
este din ce în ce mai mic pe măsură ce se află mai aproape de rădăcină iar bacteriile capabile să le sintetizeze au fost mai numeroase.
Rovira (1956), în scopul de a demonstra că efectul de rizosferă se datorează secreţiilor radiculare, în special, a adăugat solului soluţia nutritivă care conţinea substanţele eliberate în ea a rădăcinilor de mazăre, în timp ce, în paralel, alte probe s-au tratat numai cu soluţia nutritivă simplă. El constatat că stimularera bacteriilor G– sporogene este unul din cele mai stabile criterii de apreciere a efectului de rizosferă.
În rizosfera plantelor predomină bacteriile nesporogene, G-, din genurile Pseudomonas, Flavobacterium, Achromobacter şi Agrobacterium (în rizosfera plantelor leguminoase se regăsesc şi Rhizobium sp., Azotobacter sp., Azospirillum sp.).
4.2.2. Natura secreţiilor radicelare şi influenţa lor asupra
structuriilor taxonomice a microflorei solului Încă de la primele observaţii privind existenţa densităţii diferite a
microorganismelor în jurul rădăcinilor, Hiltner trage concluzia că secreţiile eliberate de plantele leguminoase determină proliferarea altor grupe de microorganisme, comparativ cu cele de la plantele de muştar.
Landergadh şi Stenlind, lucrând cu rădăcini extirpate, au găsit că grâul a secretat o serie de substanţe, ca glucoza, flavonoli şi nucleotide, în timp ce mazărea a secretat numai glucoză şi nucleotide.
Rovira, analizând compoziţia materialului secretat de rădăcinile de mazăre şi ovăz, stabileşte că mazărea elimină prin rădăcini cantităţi mai mari de aminoacizi decât ovăzul; totodată, seria aminoacizilor este mai variată la mazăre, cuprinzând un număr mai mare de compuşi aminici.
Substanţele biologic active sunt, de asemenea, eliminate de rădăcini, contribuind la stimularea înmulţirii în rizosferă a microorganismelor. Au fost puse în evidenţă tiamina (B1), biotina, riboflavina (B2), acidul pantotenic, acidul folic şi acidul nicotinic.
O analiză critică a rezultatelor şi concluziilor la care au ajuns cercetătorii care s-au ocupat cu studiul microbiologic şi chimic al rizosferei plantelor scoate în evidenţă următoarele aspecte:

118
• în jurul rădăcinilor plantelor care vegetează normal există un număr de microorganisme, care este mai mare de 10 ori, comparativ cu numărul celor existente în sol fără rădăcini; • mecanismul stimulării microflorei se bazează pe capacitatea plantelor de a elimina prin rădăcini substanţe cu rol energetic, plastic şi de stimulare a creşterii; • în rizosferă predomină bacteriile G–, nesporogene din genul Pseudomanas; • selecţia în cadrul microflorei rizosferice apare ca un efect al interacţiunii microbiene. Microorganismele cu un potenţial ridicat de sinteză a substanţelor biologic active, dotate cu capacitate mare de înmulţire şi capabile să se hrănească cu produsele eliminate de rădăcini, precum şi cele care în cursul metabolismului lor produc substanţe care inhibă înmulţirea altora, ajungând dominante în structura microbiologică a rizosferei. • secreţiile radiculare ale plantelor sunt influenţate de condiţiile de nutriţie, de condiţiile climatice (umiditatea solului, reacţia mediului, lumină, temperatură, aeraţie etc.).
4.2.3. Vârsta plantelor şi efectul de rizosferă În stadiile timpurii, plantele nu influenţează puternic microflora
rizosferică, acest lucru realizându-se după ce plantele au atins stadiul de dezvoltare maximă.
Krasilnikov (1958) şi alţi autori au găsit că numărul microorganismelor atinge apogeul când plantele sunt în stadiul de creştere activă, mai ales în faza de înflorit.
Diferenţele de rezultate se datorează metodologiei folosite şi modului de interpretare dar, în principiu, efectul de rizosferă creşte odată cu dezvoltarea plantelor şi prezintă fluctuaţii în funcţie de condiţiile de mediu.
4.2.4. Micorizele Sub denumirea de micoriză (gr. mikes – ciupercă, rhiza – rădăcină) se
înţelege o asociaţie de tip benefic între ciupercile filamentoase şi rădăcinile sau rizoizii dicotiledonatelor superioare (plante lemnoase şi ierboase), între

119
ciupercile filamentoase şi rădăcinile unor monocotiledonate (orhidee), precum şi cu muşchii (Bryophyte) şi ferigile (Pteridophyta).
Ciupercile de micoriză se dezvoltă numai în solul din jurul rădăcinii, ajungând cel mult la suprafaţa şi în interiorul celulelor corticale radiculare. Prezenţa micorizelor se exteriorizează prin apariţia pe rădăcinile laterale ale plantei gazdă a unor excrescenţe scurte şi ramificate care înlocuiesc perişorii absorbanţi şi formează o pâslă de miceliu fungic care alcătuieşte, cu timpul, un adevărat manşon periradicular, colorat variabil, începând de la alb până la brun sau negru. Rădăcinle invadate de ciuperci sunt mai scurte decât cele normale şi prezintă o hipertrofie a celulelor corticale.
Plantele furnizează ciupercilor de micoriză substanţe nutritive organice (glucide, vitamine ş.a.) şi acestea favorizează, în schimb, absorbţia apei şi a substanţelor nutritive minerale (P, N, K ş.a.).
Micorizele sunt asociaţii temporare, deoarece ciupercile mor la sfârşitul perioadei de creştere vegetativă a plantei. Dezvoltarea micorizei este determinată de însuşirile fiziologice ale partenerilor simbiozei şi de condiţiile de mediu, precum şi de însuşirile fizice şi chimice ale solului. Ciupercile care constituie micorize au o poziţie sistematică diversificată, putând fi încadrate în următoarele subîncrengături: Mastigomycotina, Ascomycotina, Basidiomycotina sau Deuteromycotina.
După raporturile ce se stabilesc între ciupercă şi plantă, micorizele sunt de trei feluri: • Micorize ectotrofe. Ciuperca emite hife spre exterior (sol) şi spre interior (rădăcină), formând, în acest mod, un manşon micelian în jurul rădăcinilor. Hifele emise spre interior pătrund în stratul cortical al rădăcinilor la nivelul spaţiilor intercelulare, formând o reţea, numită "împletitura lui Harting” (exemple: Boletus, Psalliota, Lactarius, Rusula, Marasmius). Sunt frecvente la arborii din familiile Pinaceae, Betulaceae şi Fagaceae deoarece rădăcinile lor nu prezintă perişori radiculari. • Micorize endotrofe. În acest caz, miceliul ciupercii se dezvoltă aproape exclusiv în interiorul celulelor corticale (scoarţă şi epidermă) ale gazdei, dar pot fi întâlnite şi în spaţiul intercelular. Ele sunt mai frecvente decât cele ectotrofe şi sunt întâlnite la plantele ierboase şi lemnoase ale căror rădăcini prezintă perişori radiculari (Rhizoctonia repens la orhidee şi Phoma sp. la Calluna vulgaris – iarba neagră).

120
Prezenţa ciupercii se manifestă prin tuberizarea rădăcinii, ca la orhidee. De obicei, ciuperca poate fi distinsă la examenul microscopic al rădăcinii, în care formează structuri caracteristice sub formă de „arbusculi” şi umflături terminale – „vezicule”. În aceste structuri se depun grăsimi de rezervă la nivelul celulelor corticale ale rădăcinii.
Pe măsură ce ciuperca se dezvoltă, creşterea în lungime a rădăcinilor infectate este inhibată, iar perişorii absorbanţi dispar. În cele din urmă, ramificaţiile sub formă de arbusculi, situate în grosimea parenchimului cortical, sunt, la rândul lor, digerate de celulele radiculare infectate. Aceasta sugerează că, în timp ce micorizele ectotrofe prezintă particularităţile unei simbioze adevărate, cele endotrofe apar mai curând ca o formă de parazitism controlat (după Pochon). • Micorizele ectoendotrofe sau peritrofe reprezintă o formă de tranziţie, caracterizată prin faptul că, pe lângă aspectul ectotrof, reprezentat de dezvoltarea abundentă a ciupercii în rizosferă şi la suprafaţa rădăcinii, se observă şi pătrunderea hifelor în celulele radiculare.
4.2.5. Interrelaţiile între ciuperci şi plante în cazul micorizelor Prezenţa micorizelor are un efect favorabil asupra dezvoltării
plantelor, fapt semnalat încă din 1886 de Kmenski. Această acţiune se exercită printr-un mecanism complex, constând din mai multe aspecte: • favorizarea absorbţiei apei şi a elementelor minerale de către plantă, deoarece ciupercile de micoriză măresc suprafaţa absorbantă a rădăcinilor, stimulând, astfel, creşterea ramificaţiilor şi împiedicând suberificarea lor; • intensificarea activităţii metabolice generale a plantei, căreia ciuperca îi furnizează azotul sub formă de acid glutamic; spre exemplu, în prezenţa azotului amoniacal, speciile genului Pinus cu micorize au un conţinut dublu de azot faţă de speciile care nu prezintă micorize; aceasta se datorează faptului că sărurile de amoniu pătrund în cantitate mare în ciupercă, de unde trec, apoi, în rădăcinile plantei; • crearea posibilităţii de folosire de către plantă a unor substanţe greu solubile care conţin azot şi fosfor, prin solubilizarea lor de către ciuperci; • absorbţia substanţelor nutritive necesare plantei din sol, prin intermediul hifelor şi transportul lor de către ciupercă în ţesutul cortical al rădăcinii;

121
datorită acestui proces, plantele cu micoriză se dezvoltă mai bine decât cele lipsite de ea. • relaţiile în cazul micorizelor sunt benefice şi pentru ciupercă, deoarece aceasta preia din organismul plantei gazde diferite substanţe nutritive (hidraţi de carbon, vitamine, factori de creştere).

122
Capitolul 5
SOLUL CA MEDIU DE EXISTENŢĂ PENTRU MICROORGANISME
Solul reprezintă stratul superficial afânat al litosferei care, îndelung şi
continuu transformat prin acţiunea formaţiilor vegetale succesive, a agenţilor atmosferici şi a altor factori naturali, în condiţii caracteristice mediului geografic din care face parte, a acumulat, treptat, elementele însuşirii specifice, numite fertilitate, devenind, astfel, capabil să întreţină viaţa plantelor superioare.
5.1. Alcătuirea generală a solului În alcătuirea solului intră trei părţi şi anume:
• partea solidă (minerală şi organică); • partea lichidă sau soluţia solului; • partea gazoasă sau aerul din sol.
I. Partea solidă a solului Partea minerală a solului deţine ponderea cea mai mare (90-95%), ea
provenind din rocile pe care s-a format, prin transformarea acestora sub influenţa proceselor de dezagregare şi alterare.
Dezagregarea este fenomenul fizic de simplă mărunţire a rocilor sub influenţa variaţiilor termice, a apei, vântului şi organismelor. În urma procesului de dezagregare rezultă roci afânate, alcătuite din fragmente şi particule de diferite dimensiuni. Acest complex de dezagregare nu reprezintă încă solul, ci numai o fază premergătoare în formarea acestuia. Totuşi, rocile mărunţite posedă o însuşire nouă faţă de roca iniţială, dură masivă şi anume sunt afânate, ceea ce le face permeabile pentru aer şi apă.
Alterarea este procesul complex de modificare a mineralelor şi rocilor, care se manifestă printr-o serie de procese simple de hidratare, dizolvare, hidroliză, carbonare şi oxido-reducere. Rezultatul alterării constă în faptul că o parte din elementele minerale de nutriţie a plantelor trec în forme

123
asimilabile simple, îndeosebi în săruri minerale. În acelaşi timp, însă, iau naştere şi compuşi minerali complecşi, de natura celor care intră în alcătuirea argilelor (minerale argiloase).
Partea minerală a solului rezultă în urma proceselor de dezagregare şi în timpul solidificării propriu-zise, fiind reprezentată, în principal, prin săruri (cloruri, sulfaţi, carbonaţi, azotaţi, fosfaţi etc.), oxizi şi hidroxizi (fier, mangan, aluminiu, siliciu, etc.), minerale argiloase, care formează argila din sol – principalul coloid mineral al solului, praf sub formă de particule cu diametrul cuprins între 0,2 şi 0,002 mm şi nisip cu particule cu diametrul între 2 şi 0,2 mm. La unele soluri se pot adăuga şi fragmente cu diametrul de peste 2 mm sub formă de pietriş, pietre, bolovani.
Partea organică a solului deţine un procent mai mic decât cea minerală, circa 5-10% în greutate, dar are o importanţă esenţială în formarea, evoluţia şi proprietăţile solului. Aceasta este reprezentată de resturile vegetale şi animale, netransformate sau transformate sub acţiunea microorganismelor. Principalul constituent al părţii organice din sol îl reprezintă humusul, materie organică coloid-dispersă, de culoare brună sau brun-negricioasă.
Humusul este alcătuit din compuşi de resintetizare biochimică a produselor finale de descompunere a materiei organice, acizii humici, la care se pot adăuga produse intermediare de descompunere a materiei organice. Humusul, component specific al solului, format sub acţiunea microorganismelor asupra resturilor organice, constituie principala rezervă de substanţă nutritivă pentru plante; el determină cele mai importante proprietăţi ale solului. Ca o consecinţă a procesului de alterare a părţii minerale a solului şi de descompunere a materiei organice, se pun în libertate o serie de elemente ca: N, P, S, Mg, K, Ca, Fe, Al, Mn, Na etc. Dintre acestea, o parte ajung în soluţia solului, unde sunt asimilate de către plante, iar o altă parte se fixează în sol prin fenomene de reţinere, constituind un rezervor de elemente nutritive.
II. Partea lichidă a solului Partea lichidă sau soluţia solului este reprezentată de apa din sol, în
care se găsesc dizolvate substanţe minerale şi organice în stare de dispersie ionică, moleculară sau coloidală.

124
Importanţa fazei lichide a solului constă în aceea că ea conţine elemente nutritive în formă accesibilă plantelor şi microorganismelor, adică sub formă de săruri solubile, care disociază în anioni şi cationi.
În urma proceselor de reţinere, pe suprafaţa particulelor coloidale ale fazei solide se creează o rezervă de elemente nutritive absorbite, care se eliberează în soluţie, pe măsura consumării lor de către organismele vii. Cantitatea globală de săruri nutritive din soluţia solului este, în general, suficientă pentru satisfacerea cerinţelor trofice ale plantelor superioare şi ale microorganismelor (1,1 – 2,5 g/l).
Din punct de vedere calitativ, în soluţia solului se regăsesc şi numeroase microelemente necesare pentru desfăşurarea în bune condiţii a proceselor vitale din sol: Br, Li, I, Co, Ni, Cr, Pb, V, Zn, Cu, Ag, Au.
Compoziţia soluţiei solului este variabilă şi depinde de condiţiile climatice, de activitatea microorganismelor etc. În ceea ce priveşte soluţia solului, pentru dezvoltarea microorganismelor, o importanţă deosebită o prezintă activitatea osmotică a fazei lichide. În general, presiunea osmotică de 3-6 atmosfere, pe care o posedă celulele majorităţii microorganismelor, este suficientă pentru dezvoltarea normală a acestora în soluri, funcţie de gradul de aprovizionare a acestora.
III. Partea gazoasă a solului este reprezentată de aerul care ocupă
spaţiile libere, lăsate de către apă. Compoziţia aerului din sol se deosebeşte de cea a aerului atmosferic prin raportul cantitativ între gazele obţinute. Astfel, oxigenul se găseşte în concentraţie mai mică, comparativ cu cel din atmosferă, deoarece este consumat în activitatea microorganismelor şi în procesul de respiraţie al rădăcinilor plantelor superioare. Din această cauză, proporţia de oxigen în aerul solului scade, faţă de circa 21% în atmosferă, la numai 10-18%.
Procentul de CO2 în aerul din sol este de cel puţin 10 ori mai mare decât în aerul atmosferic, datorită eliberării sale sub acţiunea de descompunere, exercitată de microorganisme asupra substanţelor organice (0,15-0,65% în sol, faţă de 0,03% în atmosferă).
Cantitatea de azot din aerul solului variază puţin, de la 76 la 79%, faţă de aerul atmosferic, unde este de 79%.

125
Consumul de azot molecular din aerul solului este foarte redus, el fiind folosit numai de către unele microorganisme azotofixatoare. În unele soluri, în special în cele mlăştinoase, în aerul solului se pot găsi şi produsele gazoase ale metabolismului bacterian ca: NH3, CH4, H2S şi H.
Compoziţia aerului din sol nu diferă totuşi prea mult faţă de cea a aerului atmosferic, datorită schimbului permanent, care este în funcţie de permeabilitatea solului, aceasta din urmă fiind determinată de porozitatea necapilară a acestuia.
5.2. Reacţia solului Dezvoltarea normală a microorganismului depinde nu numai de
elementele chimice necesare desfăşurării normale a metabolismul acestora, ci şi de unele condiţii fizico-chimice, dintre care cea mai importantă este reacţia solului.
Majoritatea microorganismelor se dezvoltă optim la un pH apropiat de neutru (pH = 7), dar reacţia solului variază în funcţie de tipul de sol, de la pH = 4 în podzoluri-soluri acide până la pH = 9 în soloneţuri, care sunt soluri sărăturoase. Fiecare tip de sol are o reacţie caracteristică, pe care şi-o menţine datorită capacităţii de tamponare, determinată, în principal, de complexul coloidal al solului. În funcţie de reacţia şi tipul de sol, microorganismele se găsesc răspândite diferenţiat, în funcţie de capacitatea lor de a rezista la reacţii extreme, foarte acide sau alcaline.
5.3. Formarea solului Procesul de formare a solului începe imediat ce roca ia contact cu
mediul extern, cu agenţii fizici şi chimici, precum şi cu acţiunea microorganismelor vii.
Caracteristica principală a solidificării constă în acumularea materiei organice şi a humusului, care determină capacitatea de reţinere a substanţelor nutritive. Elementele chimice eliberate în urma alterării au tendinţa de a fi spălate (levigate) spre structurile mai adânci ale rocii dezagregate şi alterate. Pe de altă parte, organismele vii - plantele superioare şi microorganismele - se opun acestui proces, ducând la reţinerea şi chiar la

126
acumularea substanţelor minerale în partea superioară a solului sub formă de materie organică şi de humus.
În cursul solidificării, se întâlnesc o serie de procese în urma cărora ia naştere solul ca sistem complex şi biologic, în care se desfăşoară permanent fenomene contradictorii, de al căror echilibru depind proprietăţile solului. Astfel, în toate solurile se desfăşoară procesul de alterare a unor minerale primare şi secundare, concomitent cu formarea altor minerale secundare caracteristice; descompunerea resturilor vegetale şi animale paralel cu sinteza acizilor humici; levigarea substanţelor în paralel cu bioacumularea; procesul de acumulare a apei în sol şi pierderea ei prin evaporare şi prin transpiraţia plantelor; absorbţia energiei solare ce duce la încălzirea solului, paralel cu iradierea şi deci pierderea acestei energii.
În felul acesta, intervenţia organismelor vii asupra rocii dezagregate marchează începutul procesului de solidificare şi aduce modificări importante în marele circuit geologic, ce se desfăşoară în natură. Intervenţia organismelor face ca peste marele circuit geologic să se suprapună micul circuit biologic, care este de sens contrar cu primul. Ca rezultat al acestui proces, descompunerile substanţelor organice şi minerale sunt însoţite de intense procese de sinteză organică, efectuate de organismele vii. Astfel, se acumulează diferitele elemente necesare nutriţiei plantelor şi, în special, azotul, element important, care lipseşte din rocile iniţiale.
5.3.1. Compoziţia chimică a solului Raporturile între microorganisme şi sol sunt determinate de
compoziţia chimică a acestuia, în comparaţie cu aceea a litosferei, a rocilor de origine.
Comparând datele din coloanele tabelului prezentat mai jos, constatăm acumularea în soluri a unor elemente biogene esenţiale, cum sunt, în primul rând, carbonul (în litosferă 0,22%, iar în soluri până la 45%), şi azotul (în litosferă inexistent, în soluri până la 1,5%). De asemenea, în solurile destinate agriculturii se acumulează cantităţi importante de magneziu, fosfor şi alte elemente chimice, indispensabile vieţii, şi care, în litosferă, există în cantităţi insuficiente.
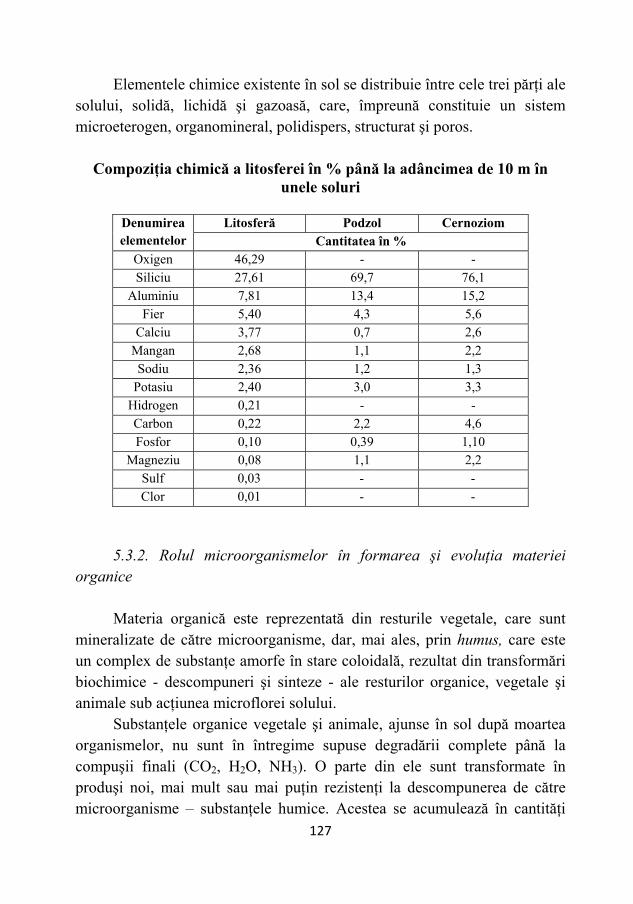
127
Elementele chimice existente în sol se distribuie între cele trei părţi ale solului, solidă, lichidă şi gazoasă, care, împreună constituie un sistem microeterogen, organomineral, polidispers, structurat şi poros.
Compoziţia chimică a litosferei în % până la adâncimea de 10 m în
unele soluri
Litosferă Podzol Cernoziom Denumirea elementelor Cantitatea în %
Oxigen 46,29 - - Siliciu 27,61 69,7 76,1
Aluminiu 7,81 13,4 15,2 Fier 5,40 4,3 5,6
Calciu 3,77 0,7 2,6 Mangan 2,68 1,1 2,2 Sodiu 2,36 1,2 1,3
Potasiu 2,40 3,0 3,3 Hidrogen 0,21 - - Carbon 0,22 2,2 4,6 Fosfor 0,10 0,39 1,10
Magneziu 0,08 1,1 2,2 Sulf 0,03 - - Clor 0,01 - -
5.3.2. Rolul microorganismelor în formarea şi evoluţia materiei
organice Materia organică este reprezentată din resturile vegetale, care sunt
mineralizate de către microorganisme, dar, mai ales, prin humus, care este un complex de substanţe amorfe în stare coloidală, rezultat din transformări biochimice - descompuneri şi sinteze - ale resturilor organice, vegetale şi animale sub acţiunea microflorei solului.
Substanţele organice vegetale şi animale, ajunse în sol după moartea organismelor, nu sunt în întregime supuse degradării complete până la compuşii finali (CO2, H2O, NH3). O parte din ele sunt transformate în produşi noi, mai mult sau mai puţin rezistenţi la descompunerea de către microorganisme – substanţele humice. Acestea se acumulează în cantităţi

128
mari ca rezerve, condiţionând proprietatea cea mai importantă a solului – fertilitatea. În felul acesta, pe lângă acţiunea lor de distrugere a substanţelor organice vegetale şi animale, microorganismele contribuie la formarea unor tipuri noi de substanţe organice. Datorită acestui fenomen, paralel cu dezagregarea enzimatică a substanţelor organice, microorganismele din sol efectuează sinteza unor compuşi noi, care se acumulează temporar în sol, formând adevărate rezerve de C,N, P, S, K etc., absolut necesare vieţii din sol.
Dar, microorganismele solului participă nu numai la formarea substanţelor humice de rezervă, ci contribuie şi la descompunerea lor lentă, prin care se asigură plantelor elementele indispensabile pentru noile sinteze organice.
5.3.3. Teoria microbiologică a formării humusului Procesele de dezagregare şi sinteză, care au drept rezultat formarea
complexelor humice, se datorează acţiunii bacteriilor aerobe şi anaerobe, a actinomicetelor şi a ciupercilor microscopice.
Bacteriile anaerobe acţionează asupra substanţelor organice aflate sub formă de mase dense în profunzimea solului. Procesul este inhibat în momentul când cantitatea de acid ulmic acumulat este atât de mare, încât opreşte dezvoltarea microorganismelor respective şi este reluat după degradarea acestuia. În condiţii naturale, denaturarea acidului ulmic se produce după acţiunea frigului, iarna. Datorită acestui fapt, sinteza anaerobă a compuşilor humici are un caracter periodic, fiind mai activă primăvara şi vara şi scăzând în intensitate spre toamnă şi iarnă.
Acest proces reprezintă forma cea mai importantă de acumulare a substanţelor humice în natură.
Sinteza aerobă a substanţelor humice se realizează în straturile mai superficiale ale solului şi cu condiţia ca acumularea substanţelor organice vegetale să se fi făcut în masa afânată. Ea este efectuată de bacterii, în cazul resturilor de plante ierboase, şi de ciupercile microscopice, atunci când substanţa organică provine din plante lemnoase. În urma acestui proces rezultă acizi humici, care nu omoară microorganismele, deoarece sunt neutralizaţi de NH3, format în paralel.

129
Compoziţia chimică a humusului este variabilă şi heterogenă, neputând fi definită prin nicio formulă chimică; ea constă din acizi ulmici şi fulvici (70-80%) şi din reziduu insolubil (20-30%).
Conţinutul bogat în N al humusului, în jur de 5% cu raportul C : N = 10:1, denotă caracterul său de produs de sinteză. El conţine, în special, acid ulmic (acid humic brun), rezultat din acţiunea bacteriilor anaerobe, şi acid humic (acid humic negru), produs de ciuperci.
Una dintre particularităţile cele mai importante ale substanţelor humice este aceea de a fi mai stabile, în condiţii termodinamice date, decât substanţele naturale din care s-au format. În acelaşi timp, ele sunt într-o permanentă evoluţie; pe de o parte, pornind de la diferite substanţe organice animale şi vegetale se formează substanţe noi sub acţiunea microorganismelor, iar pe de altă parte, complexele humice sintetizate sunt mineralizate de microflora solului.
Datorită acestui proces continuu, formarea humusului se poate afla în diferite stadii de evoluţie, sub formă de: • substanţe prohumice, capabile să agrege particulele de sol; • humus activ, cel mai important pentru fertilitate, care, prin descompunere, formează uşor substanţe nutritive pentru plante şi reprezintă aproximativ 1% din greutatea solului; • humus stabil care cimentează agregatele de sol dar se descompune greu şi nu participă activ la nutriţia plantelor; • humus inactiv, cu capacitate redusă de mineralizare şi fără rol în nutriţia plantelor sau în formarea structurii stabile a agregatelor de sol.
Humusul îndeplineşte numeroase funcţii de importanţă excepţională, din care se poate deduce rolul microorganismelor care participă la sinteza şi descompunerea sa. Astfel, humusul formează în sol rezerve imense de N, C, P, S, K, Fe, care pot fi folosite după mineralizarea prealabilă. După Waksman, cantitatea de substanţe organice sub formă de humus depăşeşte cu mult pe cea prezentă în toate formele vii.
Humusul favorizează nutriţia plantelor superioare, furnizându-le substanţe nutritive, rezultate din descompunerea sa treptată şi lentă de către microorganisme.
Humusul influenţează caracteristicile fizice ale solului, căruia îi conferă structura granulară şi culoarea întunecată, care favorizează absorbţia

130
razelor solare şi determina creşterea temperaturii. De asemenea, reduce vâscozitatea argilelor şi uşurează formarea de agregare care ameliorează aerarea şi drenajul solului, mărind, în acelaşi timp, capacitatea de reţinere a apei din sol.
Humusul influenţează proprietăţile chimice ale solului, prin faptul că menţine echilibrul general între ionii adsorbiţi şi cei dizolvaţi şi dă solului putere tampon.
Prin participarea sa la sinteza şi transformările substanţelor humice, microflora solului îndeplineşte un rol de o importanţă vitală, deoarece, datorită proprietăţilor amintite, humusul conferă solului proprietatea sa de bază – fertilitatea care-l deosebeşte de roca sterilă.
După definiţia dată de Thaer „Humusul este produsul materiei vii şi izvorul acesteia” sau după cum afirmă Waksman, „Humusul este o rezervă şi un stabilizator al vieţii organice pe pământ”.
5.4. Populaţia solului Totalitatea vieţuitoarelor din sol, indiferent de dimensiuni, organizare
celulară, forme, apartenenţă sistematică şi activitate biologică, reprezintă populaţia solului. Cele vizibile cu ochiul liber, datorită dimensiunii lor, sunt studiate încă din cele mai vechi timpuri şi formează macropopulaţia solului. Celelalte organisme formează micropopulaţia solului şi au putut fi observate şi analizate după perfecţionarea mijloacelor de mărire a imaginii.
Macropopulaţia solului este împărţită în două componente majore: 1. Macroflora solului este formată din părţile subterane ale plantelor
superioare (rădăcini, tulpini şi frunze ajunse în sol), care furnizează solului cea mai importantă sursă de materie organică. Rădăcinile plantelor, prin eliminarea a numeroase substanţe (în special enzime) în cadrul procesului de adsorbţie, au un rol important în modificarea concentraţiei soluţiei solului, intervenind, pe această cale, în mod direct în activitatea microorganismelor. Datorită acestor condiţii specifice, numărul de microorganisme aflate în jurul rădăcinilor (rizosferă) depăşeşte cu mult pe cel aflat în restul solului.
2. Macrofauna solului cuprinde o diversitate mare de organisme, dintre care amintim: râme (90-500 kg/ha), furnici, coleoptere, nematozi (circa 10 mii/mp), viermi inelaţi şi diverse vertebrate. Acestea, prin

131
activitatea lor, îmbogăţesc solul în materie organică, ce va fi utilizată ulterior de către populaţiile de microorganisme.
Micropopulaţia solului este reprezentată de microfloră şi microfaună. 1. Microflora solului constituie populaţia cea mai importantă pentru
viaţa solului. Ea este formată din bacterii, cyanobacterii, ciuperci microscopice şi alge.
Activitatea microorganismelor din sol poate fi determinată prin raportarea numărului lor la unitatea de volum, de greutate sau de suprafaţă. Valoarea absolută a unor microorganisme, raportată la un gram de sol, variază între 1 milion-1 miliard de bacterii, între 5000 şi 50000 la ciuperci, între 3000 şi 30000 la actinomicete şi 500 şi 5000 la alge (la adâncimea de 2-5 cm).
2. Microfauna solului este constituită din protozoare monocelulare (între 100-1000 indivizi/g sol), dintre care majoritatea aparţin clasei Flagellata (Mastigophora). Deşi acţiunea lor este cunoscută, rolul protozoarelor în sol nu este pe deplin clarificat.
Numărul microorganismelor din sol şi componenţa lor sistematică constituie un important criteriu practic în aprecierea activităţii biologice a acestora dar pentru o edificarea cât mai exactă a acestei activităţi este necesară şi cunoaşterea activităţii biochimice a microorganismelor care alcătuiesc populaţia solului.

132
Capitolul 6
ROLUL MICROORGANISMELOR ÎN CIRCUITUL MATERIEI ÎN NATURĂ
6.1. Circuitul azotului Azotul este un element esenţial pentru existenţa vieţii în biosferă,
deoarece este inclus în structura tuturor proteinelor şi a acizilor nucleici. Deşi este prezent în natură în cantităţi foarte mari, se găseşte aproape invariabil în forme inaccesibile direct, plantelor şi animalelor.
Circuitul azotului în natură (Fig. 69) se desfăşoară pe parcursul a mai multor etape, care implică desfăşurarea unor multiple activităţi biochimice; unele au loc în anaerobioză, iar altele în prezenţa oxigenului.
Aceste etape sunt: I. Fixarea azotului molecular se poate realiza pe cale abiotică şi biotică.
Pe cale abiotică, fixarea se realizează prin iradieri, descărcări electrice şi prin intermediul precipitaţiilor.
Pe cale biotică, fixarea azotului molecular este un proces lent şi continuu, prin care, sub acţiunea a peste 100 genuri de bacterii diferite, azotul gazos atmosferic este convertit la forme fixe (NH4
-, NO3-, NO2
-), care sunt folosite de plante şi introduse în forme organice, utilizând energia obţinută prin fotosinteză.
Bacteriile care participă la această fixare au fost grupate astfel: • bacterii fixatoare de azot molecular (N2) libere aerobe; • bacterii fixatoare de azot molecular (N2) libere anaerobe; • bacterii fixatoare de azot molecular (N2), care trăiesc în asociaţie cu plantele.
În cazul celui de-al treilea grup, în funcţie de relaţia care se stabileşte între cei doi parteneri, întâlnim patru tipuri de simbioză:
• simbioza Rhizobium – leguminoase; • simbioza Actinomicetae – neleguminoase; • simbioza asociativă; • simbioza foliară.

133
II. Amonificarea constă în mineralizarea azotului proteic până la amoniac (NH3). III. Nitrificarea prezintă două trepte: nitritarea şi nitratarea, în care are loc oxidarea amoniacului (NH3) până la nitriţi (NO2
-) respectiv nitraţi (NO3-).
IV. Denitrificarea este procesul prin care are loc reducerea azotului nitric până la azot molecular, încheindu-se, în acest mod, circuitul azotului.
Toate procesele desfăşurate pe parcursul celor patru etape sunt
mediate numai de bacterii, cu excepţia unei etape premergătoare amonificării, proteoliza, în care pot acţiona şi microfungii.
În timp ce în ecosistemele naturale, circuitul azotului este un proces echilibrat, în sensul că pierderile de azot din sol sunt compensate de
Fig. 69 – Reprezentarea schematică a principalelor etape ale circuitului azotului în natură
(după Delviche, 1977).

134
procesele de fixare, în sistemele supuse procesului de agricultură, pierderile depăşesc ritmul de înlocuire.
Dacă rezervele de azot din sol sunt limitate, fapt ce duce la apariţia unor frecvente dezechilibre între nevoia de azot şi posibilitatea de a o satisface, din contră, rezerva de azot atmosferic, este practic inepuizabilă, calculele arătând că aceasta ar putea asigura necesarul pentru cel puţin 800.000 ani. Din păcate, această imensă cantitate este inaccesibilă în mod direct organismelor vii şi mai cu seamă plantelor superioare.
În cursul ultimilor decenii, datorită nevoilor crescânde de hrană pentru o populaţie în permanentă dezvoltare, cantităţile de îngrăşăminte chimice cu azot aplicate au crescut continuu. Deşi, o perioadă de timp, îngrăşămintele chimice cu azot au îndreptăţit speranţele rezolvării şi pe viitor a satisfacerii nevoilor nutritive pentru populaţie, criza energetică a ultimilor ani şi apariţia unor efecte secundare nedorite au impus reconsiderarea acestei posibilităţi.
Azotul atmosferic este convertit industrial în amoniac (NH3), prin procedeul Haber. Reacţia are loc la temperaturi ridicate de cca.. 500oC şi la o presiune de 300-600 atmosfere, ceea ce presupune un consum ridicat de energie de cca.. 22000 kcal/kg.
În acelaşi timp, excesul de îngrăşăminte chimice cu azot are efecte negative asupra mediului şi, uneori, asupra omului, prin produsele vegetale pe care acesta le consumă.
Creşterea conţinutului în proteină la plante sub influenţa îngrăşămintelor cu azot are loc în limita potenţialului genetic al speciei sau soiului. De aceea, excesul de azot duce la o serie de deranjamente şi la acumularea de azot nitric în plante. Aceste acumulări în plantele furajere sau alimentare provoacă intoxicaţii grave, ce pot duce chiar la moartea individului.
La copii, nitraţii sunt reduşi la nitriţi, care, fiind absorbiţi de sânge, reduc capacitatea acestuia de a transporta oxigenul prin formarea metahemoglobinei.
La adulţi, nitraţii sunt convertiţi la nitrosamină, care, de asemenea, este otrăvitoare.
Folosirea în doze excesive a azotului duce la spălarea acestuia în straturile profunde ale solului şi la îmbogăţirea în nitraţi a pânzei de apă freatică care poate fi utilizată ca apă potabilă sau la adăpatul animalelor. De

135
asemenea, utilizarea intensivă a îngrăşămintelor chimice duce la creşterea conţinutului solului în unele elemente (Zn, Pb, Ni, Cr etc.) care anterior se găseau în forme solubile doar ca urme. Creşterea concentraţiei acestora peste o anumită limită are efecte toxice asupra plantelor şi microorganismelor (Davidescu, 1974).
Datorită crizei energetice şi efectelor secundare ale îngrăşămintelor chimice cu azot, o atenţie tot mai mare se acordă în ultimul timp mecanismelor naturale de fixare a azotului în sol şi, în special, a fixării biologice.
Prin procesele de fixare biologică sunt introduse în sol cantităţi foarte mari de azot. Microorganismele fixatoare de azot posedă un complex enzimatic, denumit nitrogenază, capabil să activeze tripla legătură a moleculei de azot, care poate fi astfel redusă la amoniac.
Procesul se desfăşoară la o temperatură relativ scăzută, de 15-40oC, frecvent întâlnită în sol şi la presiune atmosferică normală. În funcţie de numeroşi factori, dar mai ales de planta de cultură, cantitatea de azot atmosferic fixată biologic variază între limite foarte largi.
Legat de acest aspect, în anul 1971, Hank a sintetizat rezultatele din literatura de specialitate pentru diferite ecosisteme, astfel:
- culturi de câmp 7-28 kg N/ha; - angiosperme şi alte plante neleguminoase 7-114 kg N/ha; - leguminoase 73-865 kg N/ha; - pădure 58-594 kg N/ha; - păşuni 13-99 kg N/ha; - ape 70-250 kg N/ha. Fenomenul de fixare a azotului de către microorgansime a fost pus în
evidenţă în anul 1888 de către Berthelot. Acesta a urmărit, în paralel, variaţia azotului în vase cu sol nesteril şi în vase cu sol steril, constatând că, în solul nesteril, conţinutul în azot creşte iar în vasele cu sol sterilizat nu.
6.1.1. Fixarea azotului molecular
1. Fixarea azotului molecular (N2) de către microorganisme libere aerobe
În anul 1901, Beijerinck reuşeşte izolarea din sol a două specii de bacterii care au capacitatea de fixare aerobă a azotului molecular. Ulterior,

136
cercetările au evidenţiat existenţa şi a altor specii capabile să realizeze acest proces.
Dintre cele mai cunoscute specii putem enumera: - Azotobacter chroococcum; - Azotobacter vinelandii; - Azotobacter agile; - Azotobacter insigne; - Azotobacter macrocytogenes; - Azotobacter mobile; - Azotobacter fluminensis; - Beijerinckia indica; - Derxia gummosa; - Rhodospirillum sp.; - Rhodopseudomonas sp. Dintre acestea, cele mai răspândite şi mai cunoscute sunt cele din
genul Azotobacter. Bacteriile din genul Azotobacter sunt răspândite în natură mai ales în
solurile cultivate, bogate în humus, cu reacţie neutră şi se găsesc rar în solurile acide, podzolice, turboase şi mlăştinoase. Morfologia lor este foarte variabilă, în funcţie de condiţiile de mediu, fapt ce explică descrierea la un moment dat a numerose specii separate, care, ulterior, au fost considerate ca sinonime. Dintre speciile genului Azotobacter, cea mai răspândită şi mai eficace este Azotobacter chroococcum.
Azotobacter chroococcum prezintă o serie de aspecte morfologice considerate ca făcând parte dintr-un ciclu de evoluţie corelat cu vârsta.
În funcţie de vârsta coloniei, celulele apar diferenţiat: 1. Celulele tinere au aspect de bastonaşe groase de 2 x 4 μm, cu
capetele rotunjite, izolate sau în perechi. 2. Celulele adulte sunt sferice sau ovale ca nişte coci mari de 2 x 1,2
μm, conţinând numeroase granulaţii, constituite din lipide, volutină, glicogen etc.
3. Celulele bătrâne, care apar în a 5-a zi de dezvoltare a culturii, sunt aproape sferice şi au dimensiuni mai mici de 1,5 x 1,3 μm. În condiţii nefavorabile de mediu apar chiştii.
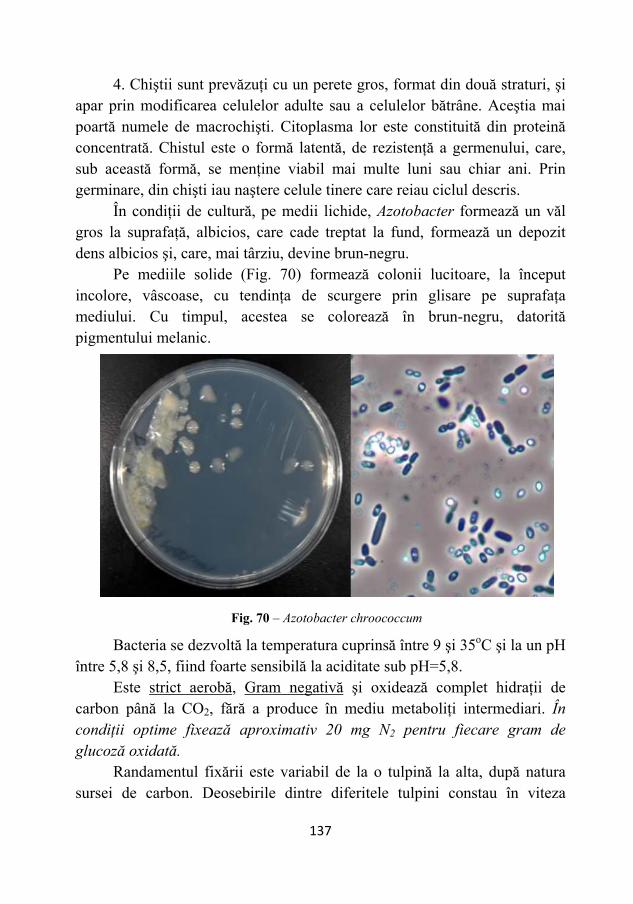
137
4. Chiştii sunt prevăzuţi cu un perete gros, format din două straturi, şi apar prin modificarea celulelor adulte sau a celulelor bătrâne. Aceştia mai poartă numele de macrochişti. Citoplasma lor este constituită din proteină concentrată. Chistul este o formă latentă, de rezistenţă a germenului, care, sub această formă, se menţine viabil mai multe luni sau chiar ani. Prin germinare, din chişti iau naştere celule tinere care reiau ciclul descris.
În condiţii de cultură, pe medii lichide, Azotobacter formează un văl gros la suprafaţă, albicios, care cade treptat la fund, formează un depozit dens albicios şi, care, mai târziu, devine brun-negru.
Pe mediile solide (Fig. 70) formează colonii lucitoare, la început incolore, vâscoase, cu tendinţa de scurgere prin glisare pe suprafaţa mediului. Cu timpul, acestea se colorează în brun-negru, datorită pigmentului melanic.
Bacteria se dezvoltă la temperatura cuprinsă între 9 şi 35oC şi la un pH
între 5,8 şi 8,5, fiind foarte sensibilă la aciditate sub pH=5,8. Este strict aerobă, Gram negativă şi oxidează complet hidraţii de
carbon până la CO2, fără a produce în mediu metaboliţi intermediari. În condiţii optime fixează aproximativ 20 mg N2 pentru fiecare gram de glucoză oxidată.
Randamentul fixării este variabil de la o tulpină la alta, după natura sursei de carbon. Deosebirile dintre diferitele tulpini constau în viteza
Fig. 70 – Azotobacter chroococcum
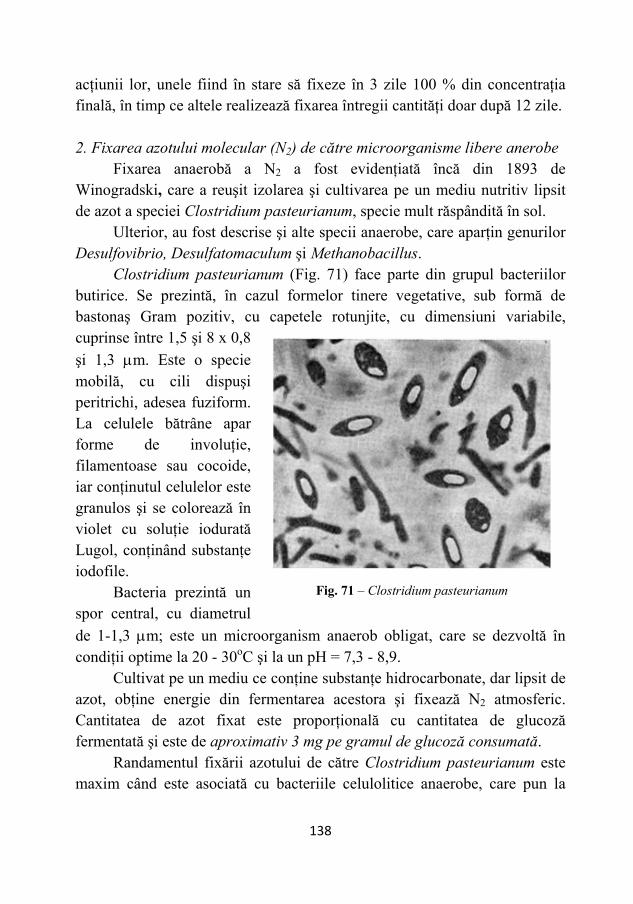
138
acţiunii lor, unele fiind în stare să fixeze în 3 zile 100 % din concentraţia finală, în timp ce altele realizează fixarea întregii cantităţi doar după 12 zile.
2. Fixarea azotului molecular (N2) de către microorganisme libere anerobe
Fixarea anaerobă a N2 a fost evidenţiată încă din 1893 de Winogradski, care a reuşit izolarea şi cultivarea pe un mediu nutritiv lipsit de azot a speciei Clostridium pasteurianum, specie mult răspândită în sol.
Ulterior, au fost descrise şi alte specii anaerobe, care aparţin genurilor Desulfovibrio, Desulfatomaculum şi Methanobacillus.
Clostridium pasteurianum (Fig. 71) face parte din grupul bacteriilor butirice. Se prezintă, în cazul formelor tinere vegetative, sub formă de bastonaş Gram pozitiv, cu capetele rotunjite, cu dimensiuni variabile, cuprinse între 1,5 şi 8 x 0,8 şi 1,3 μm. Este o specie mobilă, cu cili dispuşi peritrichi, adesea fuziform. La celulele bătrâne apar forme de involuţie, filamentoase sau cocoide, iar conţinutul celulelor este granulos şi se colorează în violet cu soluţie iodurată Lugol, conţinând substanţe iodofile.
Bacteria prezintă un spor central, cu diametrul de 1-1,3 μm; este un microorganism anaerob obligat, care se dezvoltă în condiţii optime la 20 - 30oC şi la un pH = 7,3 - 8,9.
Cultivat pe un mediu ce conţine substanţe hidrocarbonate, dar lipsit de azot, obţine energie din fermentarea acestora şi fixează N2 atmosferic. Cantitatea de azot fixat este proporţională cu cantitatea de glucoză fermentată şi este de aproximativ 3 mg pe gramul de glucoză consumată.
Randamentul fixării azotului de către Clostridium pasteurianum este maxim când este asociată cu bacteriile celulolitice anaerobe, care pun la
Fig. 71 – Clostridium pasteurianum

139
dispoziţie hidraţi de carbon solubili, ca glicerina sau manita, care se găsesc în sol în concentraţii mici.
Fixare anaerobă a N2 are un rol esenţial în solurile puţin aerate sau invadate de apă (mlaştini, orezării), unde Clostridium pasteurianum este principalul fixator de azot.
În ceea ce priveşte proporţia dintre cele două specii descrise, Fedorov arată că, în soluri Azotobacter chroococcum a fost găsit numai în 30 % din cazuri, în timp ce Clostridium pasteurianum în 70 %, având deci o preponderenţă mai mare.
3. Bacterii fixatoare de azot molecular (N2), care trăiesc în asociaţie cu plantele • Simbioza Rhizobium – leguminoase
În solurile cultivate cu leguminoase, în afară de fixarea azotului atmosferic (N2) de către bacteriile libere aerobe sau anaerobe, se mai produce şi fixarea de către microorganismele simbiotice care trăiesc în nodozităţile de pe rădăcinile acestor plante.
Deşi formarea de nodozităţi pe rădăcinile plantelor leguminoase se cunoaşte din antichitate, fenomenul era considerat ca un proces patologic, tumoral sau datorat unor înţepături de insecte. Abia în 1865, Voronin a descris nodozităţile şi corpusculii ce-i conţin, iar în 1887, Beijerinck a izolat şi descris bacteriile pe care le produc.
Biologia bacteriilor de nodozităţi Bacteriile care produc nodozităţi la leguminoasele cu care trăiesc în
simbioză aparţin genului Rhizobium care cuprinde mai multe specii distincte, cu capacitatea de a infecta numai anumite plante gazde (Fig. 72), după cum urmează:
- Rhizobium leguminosarum – mazăre, bob, linte; - Rhizobium trifolii – trifoiul; - Bradyrhizobium japonica (sin. Rhizobium japonicum) – soia; - Rhizobium phaseoli – fasolea; - Rhizobium lupini – lupinul; - Rhizobium meliloti – lucerna.

140
Activitatea diferitelor specii de Rhizobium este variabilă de la o specie
la alta, în funcţie de două proprietăţi biologice importante: infecţiozitatea, care reprezintă capacitatea de a produce nodozităţi, şi eficienţa, care se referă la capacitatea de a fixa azotul molecular (N2).
Bacteriile ajung în zona rizosferică la nivelul perişorilor absorbanţi, fiind stimulate de secreţiile radiculare, care au rol chimiotactic pozitiv, după care are loc un proces de recunoaştere a celor doi parteneri. Recunoaşterea se realizează prin intermediul unor substanţe specifice, denumite generic lectine. După legarea ireversibilă are loc deformarea perişorului radicular, care ia formă de cârlig, sub acţiunea unor substanţe eliminate de bacterii.
Ulterior, peretele perişorului absorbant este dizolvat sub acţiunea unor enzime secretate de bacterii şi acestea pătrund în interior, formând un cordon de infecţie care ajunge în celulele parenchimului cortical. Cordonul este format la început dintr-o matrice mucoasă în care se găsesc bacterii şi care, treptat, se acoperă cu o teacă celulozică. Acest manşon este constituit din secreţii ale ambilor parteneri.
Pe măsură ce cordonul infecţios înaintează, bacteriile se multiplică continuu şi sunt eliberate fie prin unele fisuri ale acestuia, fie prin extremitatea sa distală şi se aşază la periferia celulelor unde se multiplică activ. În acelaşi timp, celulele cortexului radicular infectat se multiplică rapid, transmiţând infecţia bacteriană celulelor fiice.
Rezultatul acestei proliferări este nodozitatea, care la început este inclusă în cortexul radicular, apoi îl deformează pe măsură ce creşte şi în
Fig. 72 – Nodozităţi formate de Rhizobium sp. la plantele leguminoase

141
cele din urmă îl rupe. Ea apare ca rezultat al invadării şi multiplicării bacteriilor în celulele gazde şi al acţiunii unor substanţe stimulente de tipul auxinei (acid indolilacetic) secretate de bacterii (Fig. 73).
La leguminoasele erbacee anuale nodulul este o structură efemeră, în timp ce la cele perene durează 5-6 ani, după care suferă modificări degenerative care duc la necrozarea şi detaşarea sa de rădăcină.
S-a constatat că în procesul fixării N2 nu toate nodozităţile sunt la fel
de active, ele împărţindu-se în două categorii: • Nodozităţile "eficiente" sunt relativ puţin numeroase şi se formează,
mai ales, pe rădăcinile principale; sunt mai voluminoase şi au o suprafaţă netedă sau rugoasă. Aceste nodozităţi conţin un pigment roşu, de natură proteică, numit leghemoglobină (hemoglobina leguminoaselor).
Nodozităţile eficiente au o structură caracteristică, fiind constituite din mai multe regiuni:
Perişor absorbant
Bacterii din Rhizobium sp.
Cordon infecţios
Bacteriile se transformă în bacteroizi, iar celulele îşi măresc dimensiunile.
Legarea ireversibilă a bacteriilor de perişorii absorbanţi; deformarea perişorului radicular şi pătrunderea bacteriilor în celule.
Plantă leguminoasă
Nodozitate
Bacteroizi
Apariţia nodozităţilor
Fig. 73 – Fixarea simbiotică a azotului molecular în nodozităţile leguminoaselor
(după Tortora şi colab., 1992).

142
- regiunea periferică este reprezentată de cortexul nodozităţilor, format din mai multe straturi de celule, compacte, cu rol de protecţie;
- regiunea meristematică este situată în regiunea distală a nodozităţii, fiind formată din celule diploide neinfectate şi asigură creşterea nodozităţilor prin diviziune;
- regiunea centrală este alcătuită din celule poliploide, încărcate cu bacteroizi şi prezintă trei zone distincte:
a. zona de invazie, situată sub meristem, formată din celule neinfectate şi celule mărite, infectate;
b. zona simbiotică ocupă porţiunea cea mai mare din nodozităţi şi prezintă două subzone:
- subzona simbiotică timpurie, cu celule ce conţin bacterii care încă mai proliferează; - subzona simbiotică tardivă, conţine celule mărite pline cu bacterii care nu mai proliferează numite bacteroizi. Este zona de fixare activă a azotului molecular (N2).
c. zona de senescenţă, corespunde celulelor vegetale şi bacteroizilor în curs de degenerare sau moarte.
• Nodozităţile "neeficiente" sunt mai numeroase, mai mici, cu suprafaţa netedă şi sunt răspândite pe tot sistemul radicular. Infecţia lor este foarte limitată, conţinând foarte puţine bacterii. Activitatea lor durează numai câteva zile, iar faţă de celelalte sunt nepigmentate.
Mecanismul simbiozei plantă-bacterie Bacteriile simbiotice iau energia necesară asimilării azotului
molecular prin procedee de degradare a substanţelor hidrocarbonate, furnizate de planta gazdă. La rândul lor, bacteriile asimilează N2 atmosferic până la stadiul de acid aspartic, care este preluat de plantă. Aceasta, prin transaminare enzimatică în prezenţa acidului piruvic, sintetizează alanina. În continuare, planta gazdă sintetizează ceilalţi aminoacizi, prin transaminare, pornind de la alanină şi diferiţi acizi cetonici.
Leghemoglobina este singurul reprezentant al hemoglobinelor din regnul vegetal; aceasta apare în stadiul activ al fixării, numai în nodulii eficienţi, aflaţi la maturitate, ca rezultat al simbiozei între bacterii şi leguminoase. În acest sens, bacteriile aduc nucleul "hem", iar planta

143
"proteina". Pigmentul nu poate fi sintetizat separat nici de bacterii, nici de planta gazdă şi fixarea N2 este proporţională cu concentraţia pigmentului din nodozităţi. • Simbioza Frankia sp. - neleguminoase (Actinorizele)
Actinorizele reprezintă asociaţii simbiotice fixatoare de azot molecular (N2) la aproximativ 200 specii de angiosperme, care sunt plante leguminoase caracteristice zonelor cu deficit de azot.
Bacteriile care trăiesc în simbioză cu aceste plante aparţin genului Frankia, fam. Frankiaceae, ord. Actynomycetales. Cele mai cunoscute genuri de plante în România sunt : Hippophae (Fig. 74) şi Elaeagnus.
Formarea nodozităţilor Modul de recunoaştere între cei doi parteneri şi infecţia sunt
asemănătoare ca la simbioza dintre bacteriile genului Rhizobium şi plantele leguminoase.
Evoluţia procesului infecţios determină apariţia unor nodozităţi laterale pe suprafaţa rădăcinilor. Formaţiunile simple originale au o asemănare externă evidentă cu nodozităţile de leguminoase. După 10-15 zile, pe suprafaţa nodulului simplu originar apar lobi noi, situaţi în partea apicală. Repetarea acestui proces de ramificare şi producerea anuală de noi noduli pe rădăcini duce la apariţia unei grupări compacte cu aspect coraloid, fiecare lob reprezentând o nodozitate individuală.
Importanţa deosebită a actinorizelor decurge în primul rând din faptul
că cele mai multe plante gazdă sunt specii pionier. Ele populează terenurile
Fig. 74 – Simbioza Frankia sp. cu rădăcinile de Hippophae rhamnoides.

144
cu deficit mare de azot, terenurile mlăştinoase, solurile sărace, dunele de nisip, pietriş, regiuni subalpine, zone de deşert.
În al doilea rând sunt importante prin larga răspândire a plantelor gazdă, prezente în regiunile arctice, temperate, subtropicale, tropicale şi de la regiunile de şes până la cele subalpine.
• Simbioza asociativă Prima semnalare a fixării azotului de către microorganisme asociative
a fost făcută în anul 1974 (Fig. 75). În România, prima semnalare a fost realizată în anul 1980, de către Hera.
Cercetările efectuate au pus în evidenţă existenţa mai multor specii de bacterii ale genurilor Azospirillum, Bacillus şi Klebsiella, care trăiesc în asociaţie cu rădăcinile unor graminee. Astfel, au fost identificate următoarele simbioze asociative:
- Azospirillum lipoferum - porumb; - Azospirillum brasilense - grâu, orez; - Bacillus macerans - porumb; - Klebsiella pneumoniae - Poa pratensis. Fixarea asociativă este mai răspândită de aproximativ 5 ori în zonele
tropicale faţă de zona temperată. • Simbioza foliară Simbiozele foliare au fost descrise pentru prima dată în anul 1902, de
către Zimmermann, la reprezentanţii a trei familii de plante superioare, răspândite în zonele tropicale şi subtropicale: Rubiaceae, Myrsinaceae (şi Dioscoreaceae. Astfel, frunzele mai multor specii poartă „nodozităţi” superficiale, cuprinzând bacterii. Cei mai mari noduli ating dimensiunile unei gămălii de ac (500 - 2000 µm în diametru), pot fi cilindrici sau sferici şi sunt localizaţi în funcţie de gen şi specie, pe lamină, nervura mediană sau pe peţiol.
Bacteriile pătrund prin ostiolele stomatelor şi se multiplică apoi în mezofil. Aceşti germeni care iniţial au formă de bastonaşe şi devin apoi claviformi sau ramificaţi, au fost denumiţi din 1912, de către Faber, Mycobacterium rubiacearum, iar de către alţi autori, drept specii de Flavobacterium sp. sau de Klebsiella sp.

145
6.1.2. Amonificarea Substanţele organice azotate, reprezentând 99 % din rezervele totale
de N2 din sol, constau din rezerve humice şi din alţi compuşi acumulaţi în mod natural în sol prin procesul de fixare biologică a N2 şi prin cel de degradare a resturilor organice vegetale şi animale, a îngrăşămintelor verzi şi a bălegarului.
Celulele bacteriene reprezintă ele însele o masă de substanţă organică, predominant proteică, de aproximativ 6 t/ha, la care se adaugă circa 20 de tone, reprezentate de restul microflorei şi microfaunei. Dacă acest N2 organic ar rămâne ca atare, rezervele de N2 accesibile plantelor s-ar micşora în fiecare an şi nu ar mai permite creşterea plantelor.
În mod normal, însă, aceste substanţe suferă un proces de mineralizare, la sfârşitul căruia sunt aduse în stare de amoniac (NH3).
Procesul de amonificare propriu-zis este precedat de descompunerea moleculelor proteice prin hidroliză, cu ajutorul proteazelor extracelulare,
Fig. 75 – Simbioză asociativă

146
eliberate de numeroasele specii de microorganisme aerobe şi anaerobe, după formula generală:
Aminoacizii rezultaţi din această degradare pătrund în interiorul
celulelor bacteriene, unde suferă un proces de dezaminare, din care rezultă amoniac (NH3) şi acidul organic corespunzător. Se întâlnesc mai multe tipuri de dezaminare şi anume:
1. Dezaminarea hidrolitică 2. Dezaminarea hidrolitică şi decarboxilare 3. Dezaminarea reducătoare 4. Dezaminarea reducătoare cu decarboxilare (anaerobă) 5. Dezaminarea oxidativă cu decarboxilare 6. Dezaminarea cu desaturare. Din asemenea reacţii, în afară de amoniac (NH3), pot rezulta acizii:
acetic, formic, propionic, butiric, valerianic. În funcţie de condiţiile de mediu şi natura microflorei, aceşti acizi pot fi oxidaţi complet la CO2 şi H2O (în mediul aerob), acumulaţi ca atare, transformaţi în alcooli.
Amonificarea în sens strict poate fi definită, deci, ca un proces biologic în care se eliberează în sol amoniac (NH3), ca rezultat al acţiunii microflorei solului asupra aminoacizilor, rezultaţi din descompunerea substanţelor proteice. În acest sens, nu este inclusă în amonificare eliberarea de NH3 sub acţiunea micoflorei temporare de putrefacţie.
În acest proces, NH3 poate fi refolosit ca atare de o serie întreagă de microorganisme. Cea mai mare parte din el suferă în continuare o transformare absolut necesară pentru viaţa din sol, în forme accesibile plantelor, iar restul poate fi fixat în sol, mai ales în soluri acide sau se evaporă în atmosferă.
Microflora capabilă să producă amonificarea substanţelor proteice este numeroasă, diferită şi acţionează în timp, după cum urmează:
La începutul procesului intervin bacterii obligat aerobe, cum sunt: Bacillus cereus var. micoides, B. subtilis, B. thermoproteolyticus, specii nesporulate, ca Serratia marcescens, Arthrobacter sp. etc. şi specii
proteină peptone peptide aminoaciziproteinaze peptidaze
H2O H2O H2O

147
facultativ anaerobe, cum sunt: Proteus vulgaris, Pseudomonas fluorescens, Escherichia coli, Sarcina lutea etc.
După 2-3 zile, intră în acţiune specii obligat anaerobe, cum sunt: Clostridium putrefaciens, Clostridium perfringens şi unele actinomicete, ca Streptomyces violaceus şi Micromonospora chaleea, care încep să predomine şi fac ca degajarea de NH3 să fie maximă.
Începând cu ziua a 4-a, mediul este invadat de mucegaiuri şi degajarea de amoniac (NH3) descreşte, datorită faptului că acestea folosesc amoniacul pentru sinteza proteinelor proprii şi produc mult acid, care neutralizează amoniacul.
Hidroliza ureei este realizată de un grup numeros de microorganisme, capabile să producă enzima ureaza. În acest grup întâlnim specii din genurile: Achromobacter, Bacillus, Clostridium, Corynebacterium, Pseudomonas, actinomicete şi microfungi filamentoşi. La acestea se adaugă grupul urobacteriilor, caracterizate prin rezistenţa la concentraţii mari de uree şi pH alcalin dar şi prin capacitatea de a elibera cantităţi mari de NH3. Din grupa urobacteriilor fac parte: Bacillus (Urobacillus) pasteuri, Micrococcus ureae, Planosarcina ureae etc.
Activitatea amonificatoare a urobacteriilor este foarte importantă, deoarece ureea conţine 47 % azot, care ar fi altfel neutilizat de plante. Cantitatea de uree eliminată în natură este mare, atingând câteva zeci de milioane de tone pe an. Se estimează că populaţia umană ar elimina în 24 ore circa 15.000 t azot ureic, iar animalele circa 1.500.000 tone. La aceste cantităţi se mai adaugă îngrăşămintele cu uree, aplicate prin tehnologiile actuale sub formă de cianamidă de calciu. Acest compus, fabricat din N2 din aer şi carbură de calciu, nu este asimilabil de către plante. Compusul suferă în sol unele transformări abiotice şi biotice enzimatice până la amoniac.
6.1.3. Nitrificarea Nitrificarea este un proces de oxidare a amoniacului (NH3) şi a altor
forme reduse ale azotului anorganic eliberate în sol în cursul procesului de amonificare în nitraţi, care reprezintă forma asimilabilă pentru majoritatea plantelor verzi.

148
În acelaşi timp însă, nitraţii reprezintă şi forma sub care azotul din sol este pierdut deoarece, fiind solubili, sunt spălaţi de ploaie sau reduşi la NH3, nitriţi sau chiar la azot molecular (N2) prin denitrificare.
Natura biologică a acestui proces a fost demonstrată prin suprimarea sa în urma sterilizării solului prin încălzire la 110oC sau prin tratarea cu vapori de cloroform. În laborator s-a demonstrat că procesul se desfăşoară în două faze: a. Nitritarea – reprezintă procesul de oxidare a amoniacului până la nitriţi (NO2
-).
b. Nitratarea – reprezintă procesul de transformare al nitriţilor (NO2
-) până la nitraţi (NO3
-).
Microorganismele nitrificatoare sunt autotrofe chimiosintetizante, care
îşi procură, prin reacţii chimice oxidative, energia necesară pentru producerea substanţelor celulare proprii.
Ele sunt aerobe stricte, răspândite, practic, în toate tipurile de sol şi folosesc ca sursă de carbon carbonul mineral din CO2 şi carbonaţi, iar ca sursă de azot, amoniacul sau nitriţii.
Între bacteriile nitroase sau nitrit bacterii putem cita: Nitrozomonas europaea, Nitrosococcus oceanus, Nitrosospira briensis şi Nitrosolobus multiformis, care transformă sărurile amoniacale în nitriţi.
Bacteriile nitrice (nitrat bacteriile) frecvent întâlnite sunt: Nitrobacter Winogradskii, Nitrospira gracilis şi Nitrococcus mobilis, care realizează nitratarea ce constă în oxidarea nitriţilor până la nitraţi.
Nitrificarea, care reprezintă a III-a fază a circuitului azotului în natură, este un proces de importanţă excepţională, deoarece aduce substanţele azotate în forma cea mai uşor accesibilă plantelor. De altfel, s-a observat că, în general, numărul microorganismelor nitrificatoare este proporţional cu fertilitatea solului, putând ajunge în solurile fertile până la un milion de bacterii pe gramul de sol. Numărul mare de nitrificatori din solurile fertile, comparativ cu solurile înţelenite, se datorează şi faptului că primele sunt lucrate şi, deci, bine aerate.
NO2- + H2O NO3
- + 2H+ + 2e-
NH3 + O2 NO2- + 3H+ + 2e-

149
6.1.4. Denitrificarea Nitraţii (NO3
-) acumulaţi în sol, ca rezultat al procesului de nitrificare, sunt, în parte, consumaţi de plantele superioare, iar o cantitate variabilă este spălată de apele de infiltrare şi şiroire. Nitraţii pot fi folosiţi şi de microorganisme, fie că sunt asimilaţi în cursul proceselor de sinteză a protoplasmei acestora (asimilarea nitriţilor), fie că sunt reduşi pentru oxidarea unei substanţe organice sau minerale.
Denitrificarea reprezintă un proces care închide circuitul prin întoarcerea azotului molecular (N2) în natură.
Reducerea nitraţilor în procesul de denitrificare se face fie până la N2 sau amoniac (NH3), fie până la stadii intermediare ca nitriţi (NO2
-), acidul hiponitros (HNO3
-) etc. Condiţiile optime de producere ale acestui proces sunt realizate în solurile saturate cu apă şi în structurile profunde în care pot acţiona mai multe grupuri de microorganisme. Astfel, bacteriile denitrificatoare propriu zise, ce reduc nitraţii (NO3
-) la azotul atmosferic (N2), sunt: Pseudomonas stutzeri şi Pseudomonas denitrificans.
Dintre microorganismele din flora generală a solului, capabile să reducă nitraţii (NO3
-) la nitriţi (NO2-), sunt Bacillus megaterium,
Escherichia coli, Pseudomonas aeruginosa etc., precum şi unele bacterii sulfuroase ca: Thiobacillus denitrificans.
Semnificaţia procesului de denitrificare este încă controversată, acesta fiind considerat de unii autori ca un fenomen patologic, inconstant şi relativ rar în condiţii naturale ale solurilor de cultură. Este sigur că reducerea nitraţilor până la N2 gazos reprezintă, pentru solul respectiv, o pierdere reală care poate ajunge până la 120 kg N2/ha/an, cu toate că o parte din N2 degajat poate fi preluat de fixatorii de N2 anaerobi, cum sunt: Clostridium pasteurianum.
În acelaşi timp, reducerea incompletă, până la stadiile intermediare, de nitriţi şi NH3, este mai puţin dăunătoare pentru fertilitatea solului respectiv deoarece NH3 poate fi folosit de unele microorganisme heterotrofe, în timp ce nitriţii sunt preluaţi de nitratbacterii şi reoxidări în nitraţi.

150
6.2. Circuitul carbonului
Dacă a fost relativ uşor să se realizeze o descriere destul de schematică a ciclului azotului în sol, pentru circuitul carbonului nu este la fel de simplu, deoarece, acesta din urmă nu are verigile bine individualizate ca ale primului.
Mai întâi, trebuie să arătăm că sursele de carbon introduse în sol sunt numeroase: carbonul mineral al CO2 atmosferic şi carbonaţii telurici, urmează apoi carbonul organic al vegetalelor şi animalelor sub forme multiple, de la glucide simple până la substanţe polimerizate ca celuloze, sau cu structuri complexe, ca lignina.
Metabolismul acestor forme diverse este mult diferit în sol din mai multe cauze şi anume: • datorită structurii substanţelor carbonate; • datorită florei multiple zimogene, care acţionează asupra acestor substanţe.
Aceste diferenţe se traduc prin viteza de atac diferită şi prin produşii de metabolism intermediari obţinuţi. Dacă produşii finali ai oxidării şi mineralizării carbonului organic sunt în mod constant CO2 şi H2O (şi metanul, CH4, în condiţii de anaerobioză), produsele intermediare sunt foarte variate, prezentându-se sub formă de acizi organici şi zaharuri mai mult sau mai puţin complexe.
De fapt, mineralizarea carbonului organic este, în mod excepţional, realizată într-o singură etapă şi, ca regulă generală, numeroase grupuri de bacterii şi ciuperci intervin în mod succesiv pentru a ajunge la produse simple terminale.

151
Acest ciclu al carbonului (Fig. 76) este complicat încă prin faptul că
substanţele hidrocarbonate sunt totdeauna asociate în proporţie variabilă cu substanţe azotate (proteice), lipide, precum şi cu lignina, cerurile, răşinile, taninurile etc. Proporţia relativă a acestor diverse substanţe modifică în mod profund metabolismul produşilor hidrocarbonaţi.
Metabolismul mai variază şi în funcţie de structura fizică şi chimică a solului, de climă (temperatură, umiditate), de sistemul de cultură etc.
Ultimul factor ce complică situaţia este acela că, în acelaşi timp, când substanţele hidrocarbonate sunt degradate, microorganismele realizează şi sinteza pentru propriul lor corp a unor produşi (ex. hemiceluloza).
Este ca atare dificil de ştiut care sunt produşii intermediari de degradare şi care sunt cei de sinteză. În orice caz, atât unii, cât şi alţii se pot combina pe cale chimică sau fizică între ei, precum şi cu alţi compuşi din sol pentru a forma substanţele de rezervă humică.
Pentru o mai clară prezentare a problemei, se va lua în considerare mai întâi metabolismul carbonului mineral şi apoi al celui organic.
Fig. 76 – Circuitul carbonului în natură (www.wikipedia.com)

152
6.2.1. Metabolismul carbonului mineral Se ştie că majoritatea bacteriilor au nevoie pentru metabolismul
propriu de gazul carbonic (CO2), dar acesta intră în ciclul biologic în cantităţi relativ mici. Există grupări fiziologice de bacterii care nu pot metaboliza decât carbonul mineral, cum sunt autotrofele sau litotrofele, care joacă un anumit rol în sol.
Între acestea, în primul rând, sunt bacteriile ce posedă pigment asemănător cu clorofila (bacterioclorofilă) şi algele care posedă clorofilă şi care sunt organisme fototrofe. Metabolismul acestora este asemănător cu cel al plantelor superioare, energia fiind furnizată de către radiaţiile luminoase.
Sunt apoi bacteriile lipsite de pigment, de tip clorofilian, care îşi iau energia din oxidări ale substanţelor minerale reduse (în mod esenţial NH3 şi SH2); această energie le permite să reducă CO2 şi să metabolizeze carbonul, fiind organisme chimiolitotrofe. Printre ele se află bacteriile nitrificatoare şi unele care intervin în circuitul sulfului şi al fierului. Cu ajutorul carbonului mineral metabolizat cu apa şi cu substanţele minerale luate din sol, aceste autotrofe realizează sinteza protoplasmatică, trăiesc, respiră şi, ca urmare, elimină CO2; în felul acesta, ciclul mineral al carbonului se încheie.
Dacă importanţa teoretică a metabolizării carbonului mineral este foarte mare, din punct de vedere cantitativ nu are decât un rol restrâns, comparativ cu cel al circuitului organic, ce provine de la vegetale şi animale.
6.2.2. Metabolismul carbonului organic Carbonul organic are, în principal, origine vegetală, fiind introdus în
sol prin procese multiple şi sub forme variate. Prin procese naturale este introdus sub formă de ţesuturi vegetale moarte, rădăcini, frunze, care ajung în sol odată cu moartea plantelor. Prin procese artificiale este introdus în sol prin încorporarea unor vegetale parţial metabolizate, sub formă de gunoi natural şi artificial.
Carbonul organic de origine animală este din punct de vedere cantitativ mai puţin important, fiind format din vieţuitoarele faunei telurice după moartea acestora, sub formă de cadavre. În acest caz, descompunerea
153
se realizează prin fenomene de putrefacţie, unde rolul carbonului este foarte slab, datorită raportului C/N foarte scăzut.
Ca atare, se va lua în considerare în mod esenţial compoziţia ţesuturilor vegetale care reprezintă o masă foarte însemnată de materie organică în natură. Plecând de la carbonul mineral atmosferic, de la apă şi sărurile din sol, vegetalele au făcut sinteza constituenţilor celulari. În ceea ce priveşte substanţele carbonate, ţesuturile vegetale sunt constituite din: • glucide ca monoze (hexoze-pentoze); dioze trioze, polioze, amidon, inulină, dextrină, hemiceluloză; • glucozizi ca acizi organici ficşi şi volatili; grăsimi, ceruri şi răşini; lignină. Principalii constituenţi din diferitele produse vegetale sunt prezentaţi în tabelul următor.
Constituenţii ţesuturilor vegetale
Substanţa Celuloza %
Pentozani (hemiceluloze)
Lignina %
Grăsimi şi ceară
Proteine %
Fân 28,50 13,52 28,25 2,00 9,31 Paie de ovăz 25,43 21,33 20,40 2,02 4,30 Paie de grâu 37,27 21,67 21,21 0,67 3,00
Porumb ciocălăi 37,67 31,50 14,70 1,37 2,11 Porumb boabe 30,56 23,54 15,13 0,77 3,50
Mai importantă din punct de vedere agrobiologic decât clasificarea
chimică a produşilor hidrocarbonaţi este aceea care ţine seama de solubilitatea acestora, de care este legată viteza de descompunere din sol. Avem, din acest punct de vedere, clasificarea următoare: • substanţe foarte solubile, între care sunt cuprinse glucidele simple şi acizii organici; • substanţe puţin solubile, cum este hemiceluloza; • substanţe insolubile, cum sunt: amidonul, celuloza, lipidele, cerurile şi răşinile; dintre acestea lignina formează o substanţă aparte, datorită gradului său de rezistenţă la agenţii biologici şi care are tendinţe de a se acumula în sol.
Constituenţii hidrocarbonaţii mai importanţi din punct de vedere cantitativ ai ţesuturilor vegetale, redaţi în % de substanţă uscată, sunt
154
prezentaţi în tabelul următor, din care reiese că această compoziţie este variabilă în funcţie de dezvoltarea plantei, adică de vârstă.
Compoziţia ţesuturilor la secară în funcţie de vârstă
Stadiul de creştere
Celuloză %
Pentoze %
Lignină %
Grăsimi şi ceruri
N total %
30 mm 18,06 16 9,9 2,4 2,5 Înspicare 26,9 21,18 11,8 2,6 1,76 Înflorire 30,59 22,71 18 1,7 1,01
Maturitate 36,26 22,90 17,1 1,26 0,24 În decursul creşterii cerealelor, se produce o sporire a proporţiei de
celuloză, hemiceluloză şi lignină, corelată cu scăderea proteinelor şi, mai ales, a fracţiunilor solubile. Ca urmare, ţesuturile plantelor mature vor fi mai dificil şi mai lent descompuse în sol comparativ cu cele provenite de la plantele tinere.
Concentrarea în substanţe superioare polimerizate este încă şi mai accentuată la plantele lemnoase, unde proporţia medie a acestei componente este următoarea:
- Celuloză = 45 - 55%; - Grăsimi, ceruri şi răşini = 3 - 5%; - Hemiceluloze = 20 - 30%; - Lignină = 25 - 30%; - Proteine = 0,5 - 1%. Lemnul este dificil fermentescibil în sol şi rolul ligninei este, din acest
punct de vedere, important prin îmbogăţirea acestuia cu materie organică. Legăturile acestor produse cu celuloza şi grupările uronice ale hemicelulozelor fac aceste glucide foarte rezistente la acţiunea microbilor din sol. În continuare, vor fi prezentate, succesiv, procesele de degradare biologică în sol ale diverşilor constituenţi hidrocarbonaţi şi ale ţesuturilor vegetale în funcţie de solubilitate, de la glucidele simple, substanţele solubile în apă, hemiceluloza, amidonul, peptina, celuloza, lignina etc.
I. Degradarea glucidelor simple şi a substanţelor solubile în apă După ce resturile de ţesuturi vegetale sunt încorporate în sol, glucidele
şi substanţele solubile sunt primele atacate de microorganisme şi dispar în

155
câteva zile, ajungându-se la produşii finali CO2 şi H2O. Etapele intermediare sunt însă diferite, în funcţie de umiditatea, aeraţia şi temperatura solului care condiţionează intervenţia fie a bacteriilor aerobe, fie a celor anaerobe sau a ciupercilor.
În cazul atacului bacterian aerob se formează, pe parcurs, acizii piruvic, lactic, acetic, acetaldehida, iar ca produşi finali CO2 şi H2O. În cazul intervenţiei ciupercilor aerobe se produc, pe parcurs, acizii gluconic, citric, oxalic, fumaric, succinic etc.
Atacul bacteriilor anaerobe are ca rezultat intermediar formarea de acizi volatili ca acidul lactic, hidrogenul, metanul şi CO2. În urma acţiunii fungilor anaerobi se produc acid fumaric şi alcool.
Aceste procese au fost urmărite în condiţii de laborator dar în natură situaţia este mult mai complexă. Astfel, produşii intermediari formaţi sunt fie metabolizaţi imediat de către alte grupe de microorganisme, cum este cazul la Azotobacter sp., care utilizează alcoolii şi acizii organici pentru sinteza proprie a compuşilor celulari, fie mineralizaţi complet, până la CO2 şi H2O.
În ceea ce priveşte produşii solubili în apă, a fost studiată problema zaharozei care este descompusă de flora zimogenă. Mai întâi, intervine Bacillus megaterium, care formează acid oxi-butiric, şi apoi proliferează genul Pseudomonas, care utilizează acest acid eliberat de primul, producându-se şi aici o reacţie în lanţ.
Reacţiile de degradare sunt, în mod cert, cuplate cu cele de sinteză, căci produsele mai simple rezultate sunt folosite de anumiţi germeni pentru constituirea capsulei gomoase, formată din substanţe poliozide cu greutate moleculară ridicată, de tipul hemicelulozelor.
II. Degradarea hemicelulozelor Cantitatea hemicelulozelor în resturile vegetale variază între 6 şi 30%
din greutatea uscată, în funcţie de specie şi vârsta plantei. Hemicelulozele sunt polizaharide ce însoţesc celuloza în ţesuturile vegetale şi care sunt grupate în pentoze şi hexoze. Din pentoze fac parte xilanul, format din resturi de xiloză, şi arabanul, din resturi de arabinoză. Din hexoze fac parte mananele, constituite din resturi de manoză, glucomananele, alcătuite din

156
resturi de glucoză şi manoză, precum şi galactamanele, formate din resturi de galactoză şi manoză.
Dintre hemiceluloze galactanele sunt mai rezistente la atacul microbian, în timp ce xilanele şi mananele sunt mai repede metabolizate, fiind descompuse de o floră nespecifică.
Viteza de descompunere a acestor substanţe depinde de cantitatea de azot accesibil şi se produce în mod eşalonat şi în ritm diferit. Astfel, în primele 4 zile, din paiele de ovăz, care conţin circa 23% hemiceluloză, se descompun 10% , în următoarele 4 zile alte 8,5% şi numai 4,50% în ultimele 8 zile.
Microorganismele care descompun hemiceluloza se succed într-o anumită ordine, începând cu bacilii nesporulaţi, bacilii sporulaţi şi cocobacilii.
Aproape constant apare şi Azotobacter sp. care nu prezintă specii capabile să atace direct hemicelulozele, dar folosesc produşii intermediari de metabolism, formaţi de celelalte grupe de bacterii. Dintre ciuperci s-au identificat în acest proces Zygorhynchus sp., Cuninghamella sp., Rhizopus sp., Trichoderma sp., Humicola sp., Aspergillus niger şi Aspergillus fumigatus. Actinomicetele au fost identificate, de asemenea, ca fiind active în descompunerea hemicelulozelor.
Unele bacterii polifage sunt capabile să atace şi hemicelulozele. Printre acestea se numără: Alcaligenes sp., Pseudomonas sp., Bacillus circulans şi Sporocytofaga myxococcoides, care este o bacterie celulolitică autentică. Activitatea microflorei hemicelulolitice este foarte variată, în funcţie de tipul de sol şi conţinutul în substanţă organică al ţesuturilor vegetale. Numărul bacteriilor hemicelulolitice este cel mai mare în solurile neutre şi apoi în cele acide.
III. Degradarea amidonului Concentraţia de amidon din ţesuturile vegetale este, în principal,
scăzută, cu excepţia seminţelor (max. 70% din greutatea uscată) şi a tuberculilor de cartof.
În funcţie de natura solului, greu sau uşor, de umiditate şi de temperatură, procesul de amiloliză se desfăşoară în mod diferit. Astfel, în terenurile de cultură mijlocii, bine aerate şi semisaturate în umiditate, la

157
temperatura de 18ºC, intervin în degradarea amidonului unele bacterii şi ciuperci, aparţinând genurilor Aspergillus, Fusarium şi Rhizopus.
În solurile saturate în apă şi cu celelalte condiţii asemănătoare, se găsesc bacterii aerobe şi facultativ anaerobe din genul Bacillus, mai ales B. macerans, B. subtilis şi B. cereus.
În aceste condiţii, fenomenul este mai rapid. În aceleaşi condiţii, dar la o temperatură mai ridicată, până la 33ºC, intervin bacteriile strict anaerobe, de tip Clostridium, şi amiloliza este, în aceste din urmă condiţii, foarte rapidă. Se întâlnesc bacteriile Clostridium butyricus, Clostridium perfringens, dar şi Clostridium pasteurianum, care, pe lângă degradarea amidonului, fixează şi azotul atmosferic.
În lumina acestor condiţii, se pare că amiloliza în sol este rezultatul activităţii unei flore zimogene destul de polifage, cu o dublă constituţie. O microfloră abundentă, puţin specializată, bacteriană şi fungică, care degradează amidonul până la stadiul de dextrină, apoi dextrina este metabolizată de alte microorganisme. O microfloră mai restrânsă şi mai specializată hidrolizează amidonul până la acizi, alcooli şi gaze. În ansamblu, această activitate merge în paralel cu fertilitatea aparentă a solului şi cu bogăţia microflorei totale.
IV. Degradarea pectinei Conţinutul în pectină al ţesuturilor vegetale este cuprins între 10 şi
30%, în funcţie de specie, de organe şi de vârsta plantelor. Pectinele sunt substanţe cu greutate moleculară ridicată, care aparţin grupului poliuronidelor.
Ele formează, în condiţii favorabile, geluri în contact cu apa şi se împart în mai multe grupări: • protopectine – derivaţii ale acidului poligalacturonic, legate prin anumiţi constituenţi cu membrana celulară; • acidul pectic sau pectina propriu-zisă, care este acid poligalacturonic esterificat de alcoolul metilic; • acidul pectic sub formă de acid poligalacturonic neesterificat.
Acestor trei compuşi le corespund enzime specifice ca: protopectinaza, pectinesteraza şi, respectiv, poligalacturonidaza. La

158
descompunerea substanţelor pectice participă unele bacterii, actinomicete şi ciuperci, bacteriile anaerobe fiind agenţii principali de descompunere.
Bacteriile pectinolitice anaerobe sunt reprezentate prin speciile de Clostridium, cum sunt: • Clostridium aurantibutyricum ce atacă atât pectina purificată, cât şi pectina din resturile vegetale. • Clostridium felsineum care atacă numai pectina din resturile vegetale.
Între speciile aerobe de bacterii se încadrează Bacillus polymyxa, B. macerans şi reprezentanţi ai genurilor Pseudomonas, Arthrobacter şi Corynebacterium.
Între actinomicetele pectinolitice se situează genurile Streptomyces şi Micromonospora.
Dintre ciupercile pectinolitice fac parte mai multe genuri ca: Aspergillus, Fusarium, Alternaria, Botrytis, Monilia etc. O altă specie, ciuperca Pullularia pululans, descompune, toamna, pectina din frunzele căzute, uşurând mineralizarea acestora.
V. Descompunerea celulozei Celuloza reprezintă 45-80% din greutatea uscată a resturilor vegetale
şi, ca atare, constituie cea mai importantă parte a substanţelor organice care se întorc în sol.
Fedorov apreciază cantitatea de carbon găsit în sol sub formă de celuloză la 350 milioane tone în compoziţia organelor vii şi la 700 miliarde tone în sol şi turbă.
Celuloza este un polizaharid (C6H10O5), format dintr-un lanţ de 1:4 β D – glucopiranoză (celobioză), care constituie scheletul peretelui celular la plante. Celuloza este foarte rezistentă la degradare, exceptând descompunerea produsă sub acţiunea microorganismelor, datorită căreia plantele ar primi la hectar aproximativ 15 tone CO2 pentru nutriţia lor carbonată.
În acest fel, degradarea şi mineralizarea microbiană a celulozei reprezintă şi din punct de vedere cantitativ unul dintre procesele cele mai importante ce se petrec pe suprafaţa globului.

159
Degradarea biologică a celulozei a fost evidenţiată prima dată de Omelianschi care a studiat fenomenele descompunerii aerobe iar apoi Vinogradscki a stabilit şi rolul deosebit al degradării anaerobe.
Din hidroliza celulozei rezultă glucoză şi celobioză iar celobioza, la rândul ei, în final, este descompusă în glucoză. Aceşti compuşi apar numai excepţional în sol, deoarece procesele biologice merg până la stadiile de combustie incompletă, cu eliberare de acizi şi alcooli sau completă, până la CO2 şi H2O a glucozei.
Microorganismele se dezvoltă direct pe fibrele de celuloză de care se fixează şi produc hidroliza enzimatică a celulozei, în cursul căreia fibrele îşi pierd consistenţa, se gonflează şi se transformă într-o masă cu o consistenţă păstoasă şi gelatinoasă.
A. Degradarea aerobă a celulozei poate fi realizată de bacterii şi ciuperci. Bacteriile celulolitice pot fi puse în evidenţă însămânţând particule de sol pe gel de siliciu impregnat cu o substanţă azotoasă. După câteva zile de incubare la 30ºC apar colonii colorate diferit, în galben, cărămiziu, brun-deschis, roz- portocaliu, verde şi foarte rar brun-deschis şi negru. Culoarea coloniilor variază în funcţie de vârsta culturii şi de specie.
Microorganismele celulolitice pot fi criofile, adică rezistente la temperaturi scăzute, mezofile sau termofile. Cele mai multe dintre acestea sunt mezofile sau termofile şi se împart, la rândul lor, în microorganisme aerobe şi anaerobe. Între microorganismele celulolitice mezofile aerobe sunt cuprinse o serie de bacterii, actinomicete şi ciuperci.
Bacteriile celulolitice sunt reprezentate prin mai multe genuri ce aparţin ord. Cytophagales, genul Cytophaga ce se prezintă sub formă de bastonaşe scurte sau elongate, flexibile sau ca filamente cu dimensiuni de 0,3-0,7 x 5-50 µm, cu capetele bastonaşelor rotunjite sau ascuţite. Celulele şi coloniile sunt colorate în nuanţe de galben, oranj sau roşu. Cea mai răspândită specie este Cytophaga hutchinsonii. Genul Sporocytophaga se prezintă sub formă de celule asemănătoare cu Cytophaga dar formează microchisturi rezistente la uscăciune, specia cea mai comună fiind Sporocytophaga myxococcoides.
Din ord. Myxobacteriales produc descompunerea celulozei speciile Sorangium compositum şi Polyanginum celullosarum.

160
Din fam. Pseudomondaceae face parte şi specia Cellvibrio ochracea – bacterie celulolitică foarte activă.
Se mai cunosc ca bacterii celulolitice şi specii din genurile Cellulomonas, precum şi Bacillus ca B. vagans şi B. soli.
Dintre actinomicetele celulolitice cităm speciile: Streptomyces cellulosae, Streptomyces violaceus, Micromonospora chalcea şi Nocardia cellulans.
Ciupercile celulolitice sunt agenţi activi ai descompunerii celulozei şi, mai ales, a celulozei din materialul lemnos. Dintre cele mai răspândite genuri amintim: Chaetomium, Aspergillus, Penicillium, Myrothecium şi Trichoderma.
În cursul descompunerii aerobe a resturilor vegetale se disting două etape succesive:
• În prima fază se produce descompunerea părţilor vegetale mai uşor atacabile, cu participarea predominantă a speciilor de Cellvibrio şi Myxococcus.
• În a doua fază urmează descompunerea părţilor mai rezistente la degradare, cu predominanţa speciilor Cytophaga şi a ciupercilor microscopice.
Din punct de vedere chimic, celuloliza are loc tot în două etape. La început se produce hidroliza celulozei până la celobioză (C12H22O11) şi alţi produşi mai complecşi, după care, prin oxidarea acestor produşi, se formează, ulterior, acizi uronici.
Dacă hidroliza celulozei este mai puternică, mergând până la formarea glucozei, aceasta este mai departe oxidată până la CO2 şi H2O, trecând prin stadiul de oxiacizi:
Formarea de hidraţi de carbon solubili şi de oxiacizi ca produşi
intermediari de degradare a celulozei are mare importanţă în sol, deoarece stimulează dezvoltarea fixatorilor de N2, care îi folosesc în hrană.
Degradarea aerobă a celulozei se realizează în condiţii optime în solurile bine lucrate, aerate, bogate în azot şi cu climat temperat. În aceste condiţii, bacteriile şi ciupercile celulolitice sunt active până la adâncimea de
(C6H10O5)n + nH2O + O2 R-CHOH-COOH CO2 + H2O + x cal.

161
30 cm, ceea ce corespunde, practic, zonei pentru dezvoltarea culturilor agricole.
B. Degradarea anaerobă a celulozei
Acest proces se realizează, în special, în zonele umede, mai profunde ale solului, când resturile vegetale sunt în mase compacte, dense , neaerate.
Din punct de vedere chimic, ea reprezintă o hidroliză a celulozei sub acţiunea celulazelor şi a celobiazei microbiene, cu formare de glucoză, din descompunerea căreia rezultă acizi volatili, ca formic, butirici, acetic, acizi nevolatili, ca succinic şi lactic, precum şi alcool etilic şi gaze (CO2, H2 şi metan).
Bacteriile celulolitice anaerobe din sol sunt reprezentate prin Bacillus cellulosae hydrogenicus şi Methylomonas methanica, care eliberează metan.
Fermentarea celulozei cu degajare de H2 este însoţită de formarea acidului butiric şi a CO2, după reacţia:
În fermentaţia metanică a celulozei, cantitatea de gaze eliberată
reprezintă aproximativ 1/2 din greutatea celulozei descompuse. Schema generală a reacţiei ar fi următoarea:
Unele specii de bacterii celulolitice anaerobe sunt adaptate să trăiască
în intestinul uman şi al animalelor unde determină descompunerea celulozei din alimente. Bacteriile celulolitice se dezvoltă la temperatura optimă de 55-70ºC şi sunt prezente în medii bogate în celuloză şi azot organic şi mineral, cum sunt bălegarul şi nămolurile orăşeneşti.
Unele bacterii termofile, cum sunt Clostridium termocellus şi Clostridium termocellulolyticum, degradează celuloza cu formare de acizi volatili şi ficşi, alcooli şi gaze, în special metan.
Microorganismele celulolitice mai pot contribui la distrugerea hârtiei în bibliotecile şi depozitele rău aerisite şi umede, provocând pagube însemnate.
(C6H10O5)n + nH2O CH3CH2COOH + CH3COOH + CO2 + CH4
(C6H10O5)n + nH2O CH3CH2COOH + CH3COOH + H2 + CO2 + x cal.

162
VI. Descompunerea ligninei Descompunerea ligninelor are importanţă deosebită pentru circulaţia
carbonului întrucât aceasta reprezintă cea de-a doua sursă a acestui element în natură: 25-30% din corpul plantelor lemnoase şi 15-20% din corpul plantelor ierboase.
Ligninele reprezintă, totodată, complexul vegetal mai rezistent şi, din această cauză, se acumulează continuu în sol, luând parte la formarea humusului. Ligninele sunt substanţe macromoleculare, provenite prin condensarea unor alcooli aromatici cu o structură complexă reticulară care necesită o mare cantitate de energie pentru a fi scindată.
Agenţii principali ai descompunerii ligninei sunt ciupercile din ord. Agaricales, mai ales acelea ce produc „putregaiul alb al lemnului” care pot ataca şi celuloza.
Un alt grup de ciuperci cauzează „putregaiul brun al lemnului” dar acesta nu atacă lignina.
Ciupercile ligninoclastice pot fi grupate în trei tipuri şi anume: • tipul Polyporus versicolor atacă la început lignina şi apoi celuloza; • tipul Armillaria vera atacă la început celuloza apoi lignina; • tipul Ganoderma applanatum atacă simultan, atât lignina, cât şi celuloza.
Lignina se descompune mai repede în condiţii aerobe, în care acţionează ciupercile, şi mai lent în anaerobioză când acţionează mai mult, bacteriile.
VII. Metabolismul taninurilor, lipidelor şi cutinei Taninurile sunt descompuse, în special, de către ciupercile din
genurile Aspergillus şi Penicillium. În cazul când ating o anumită concentraţie în sol, ele au o acţiune antimicrobiană.
Lipidele sunt hidrolizate cu uşurinţă de către microorganisme. Cutina este descompusă de bacterii din genul Pseudomonas în ritm
lent. 6.3. Circuitul sulfului
Circuitul sulfului (Fig. 77) este, alături de cele ale azotului şi
carbonului, unul dintre cele mai importante circuite din punct de vedere
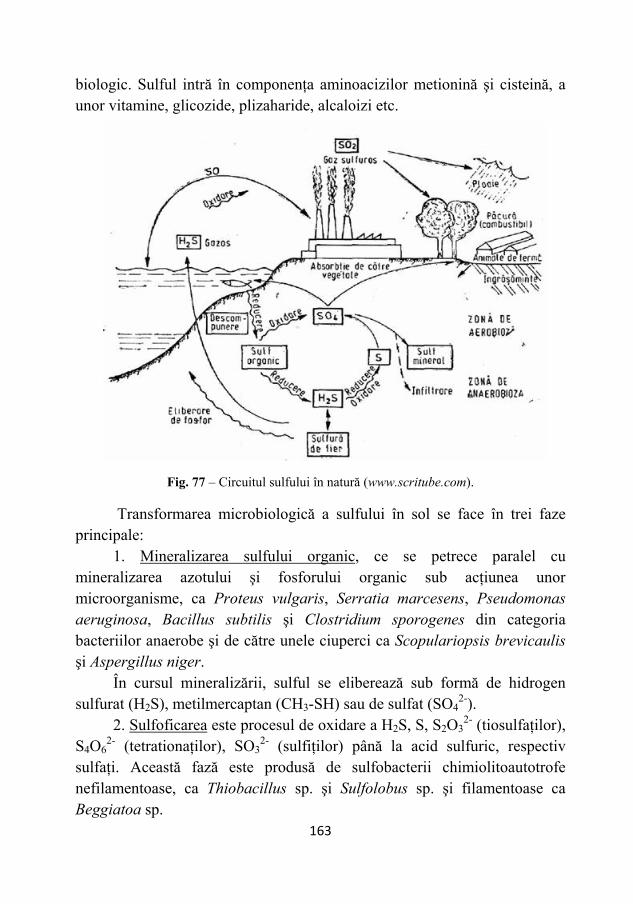
163
biologic. Sulful intră în componenţa aminoacizilor metionină şi cisteină, a unor vitamine, glicozide, plizaharide, alcaloizi etc.
Transformarea microbiologică a sulfului în sol se face în trei faze
principale: 1. Mineralizarea sulfului organic, ce se petrece paralel cu
mineralizarea azotului şi fosforului organic sub acţiunea unor microorganisme, ca Proteus vulgaris, Serratia marcesens, Pseudomonas aeruginosa, Bacillus subtilis şi Clostridium sporogenes din categoria bacteriilor anaerobe şi de către unele ciuperci ca Scopulariopsis brevicaulis şi Aspergillus niger.
În cursul mineralizării, sulful se eliberează sub formă de hidrogen sulfurat (H2S), metilmercaptan (CH3-SH) sau de sulfat (SO4
2-). 2. Sulfoficarea este procesul de oxidare a H2S, S, S2O3
2- (tiosulfaţilor), S4O6
2- (tetrationaţilor), SO32- (sulfiţilor) până la acid sulfuric, respectiv
sulfaţi. Această fază este produsă de sulfobacterii chimiolitoautotrofe nefilamentoase, ca Thiobacillus sp. şi Sulfolobus sp. şi filamentoase ca Beggiatoa sp.
Fig. 77 – Circuitul sulfului în natură (www.scritube.com).

164
Mai este produsă de bacteriile purpurii Chromatium sp. şi bacteriile verzi Chlorobium sp.
Spre exemplu, Thiobacillus thioxidans oxidează sulful elementar la acid sulfuric:
S + 3/2O2 + H2O → H2SO4
Bacteria Beggiatoa sp. oxidează H2S la S:
H2S + 1/2O2 → S + H2O
Sulful se acumulează în celulele microbiene sub formă de incluziuni. În lipsă de H2S, bacteria oxidează sulful din aceste incluziuni până la H2SO4.
Folosirea sulfului elementar poate avea aplicaţii practice pentru îmbunătăţirea solurilor alcaline. Astfel, sulful este oxidat de microorganismele sulfoficatoare până la acid sulfuric, care micşorează pH-ul mediului. Dar sulfoficarea poate avea şi efecte negative din punct de vedere agricol. În solurile inundate se formează cantităţi mari de sulfuri, datorită desulfoficării. În urma drenării şi desecării acestor soluri, sulfurile se oxidează rapid la sulf pe cale chimică, iar acesta la H2SO4, datorită microorganismelor sulfoficatoare. Ca rezultat, pH-ul acestor soluri ajunge la valori de 3,2 - 2,2, care periclitează viaţa plantelor. Pentru a preveni acest efect negativ este necesară aplicarea amendamentelor calcaroase.
3. Desulfoficarea este procesul de reducere dezasimilatorie a sulfaţilor până la H2S. Acest proces are loc în condiţii anaerobe şi se datorează, în primul rând, speciilor din genul Desulfovibrio, precum D. desulfuricans.
Bacteria reduce sulfaţii la H2S cu ajutorul hidrogenului din substanţele organice, ca lactatul şi piruvatul, care servesc atât ca surse de energie dar şi ca sursă de carbon.
Putem trage concluzia că sulfoficarea duce la acidifiere, în timp ce desulfoficarea produce alcalinizarea solului.
În procesul de obţinere a H2S rezultat din desulfoficare, nu se ating concentraţii toxice pentru plante, deoarece Fe2+ din sol îndepărtează H2S sub formă de FeS care precipită. În solurile sărace în Fe2+ şi irigate cu ape bogate în sulfaţi, H2S produs în cantităţi mari produce efecte toxice la orez şi la alte plante.

165
6.4. Circuitul fierului Fierul intră în compoziţia hemienzimelor, a hemoglobinei,
leghemoglobinei, F-S-proteinelor, iar circuitul său cuprinde patru etape microbiene ce se succed astfel:
1. Mineralizarea fierului organic se petrece în paralel cu celelalte procese de mineralizare a substanţelor organice. Microorganismele mineralizatoare atacă partea organică a compuşilor cu fier iar acest element se eliberează sub formă minerală, bi- sau trivalentă. Microorganismele respective pot fi bacterii din genurile Pseudomonas, Bacillus, Corynebacterium, Mycobacterium etc.; mai pot fi actinomicete din genurile Nocardia sau Streptomyces, precum şi ciuperci, toate ca organisme chimioorganoheterotrofe.
Compuşii organici cu Fe din humus pot fi atacaţi de bacterii ce aparţin genurilor Metallogenium, Pedomicrobium şi Seliberia. Pe suprafaţa celulelor respective se depune, sub formă de precipitat, hidroxidul feric Fe(OH)3.
2. Formarea compuşilor organici de fier este un proces opus mineralizării fierului organic. Aici, microorganismele sintetizează compuşi organici solubili, deţinători de fier, bi- sau trivalent.
Fierul poate fi sintetizat de unii compuşi organici ca acidul acetic sau complecşi, ca acizii citrici, acidul 2-ceto-D- gluconic, acizii humici etc.
Microorganismele capabile de formarea compuşilor organici cu fier sunt unele bacterii şi mucegaiuri (ciuperci) precum şi unii licheni. Mineralizarea şi formarea microbiană a compuşilor organici cu Fe joacă un anumit rol în procesul de podzolire a solului. Sub acţiunea microorganismelor, în orizontul A se formează Fe organic solubil care migrează spre orizontul B, unde, sub acţiunea microorganismelor mineralizatoare, se eliberează sub formă anorganică fierul trivalent.
3. Oxidarea fierului bivalent. La aceste procese participă ferobacteriile (bacteriile feroginoase şi siderobacteriile). Acestea sunt organisme acvatice dar unele se pot dezvolta şi în soluri, unde se prezintă sub formă filamentoasă sau nefilamentoasă.

166
La ferobacteriile filamentoase, ca Leptothrix ochracea, celulele individuale sunt incluse într-o teacă organică comună în care se acumulează Fe(OH)3.
Ferobacteriile nefilamentoase cuprind unele bacterii pedunculate, ca Gallionella ferruginea, capsulate, ca Siderocapsa treubii, şi nependulate şi necapsulate, ca Ferrobacillus thiooxidans, la care Fe(OH)3 se depozitează pe penduncul, în capsulă şi, respectiv, pe suprafaţa celulelor.
Aceste bacterii sunt considerate în prezent chimiolitoorganoheterotrofe, cu excepţia lui Thiobacillus ferrooxidans. Acest tiobacil oxidează nu numai sulful elementar, sulfurile şi tiosulfaţii dar şi Fe2+ şi asimilează CO2, dezvoltându-se în medii acide cu un pH = 2 – 6,5. Oxidând pirita (FeS2) sau calcopirita (CuFeS2), Thiobacillus ferrooxidans produce sulfat feric.
Într-o primă etapă, pirita este oxidată la sulfat feros:
2FeS2 + 7O2 + 2H2O → 2FeSO4 + 2H2SO4 + energie
În continuare, sulfatul feros este oxidat la sulfat feric:
2FeSO4 + 1/2 O2 + H2SO4 → Fe2(SO4)3 + H2O + energie
La rândul său, sulfatul feric poate oxida pirita cu forme de sulfat feros şi acid sulfuric:
7Fe2(SO4)3 + FeS2 + S H2O → 15 FeSO4 + S H2SO4
Dezvoltarea lui Thiobacillus ferrooxidans duce la acidifierea mediilor, datorită acidului sulfuric produs. La rândul său, acidul sulfuric poate solubiliza cuprul şi alte metale grele din mineralele ce le conţin. Din această cauză, dezvoltarea speciei Thiobacillus ferrooxidans duce la acumularea de concentraţii fitotoxice pentru plantele de cultură, a acidului sulfuric şi a metalelor grele în haldele de steril luate în cultură.
4. Reducerea fierului trivalent. Acest produs se datorează unor bacterii facultativ sau obligat anaerobe, ca Bacillus circulans, Bacillus polymyxa, Enterobacter aerogenas, Clostridium butyricum etc. Acestea, în prezenţa substanţelor organice, ca glucoza, reduc fierul trivalent (Fe3+) la fierul bivalent (Fe2+), fierul trivalent servind ca acceptor de electroni.

167
Mai sunt capabile de reducerea fierului trivalent şi unele ciuperci din sol. Toate aceste microorganisme fier-reducătoare sunt chimioorganoheterotrofe.
Trebuie să menţionăm că oxidarea bivalent şi reducerea fierului trivalent în sol se pot produce şi fără participarea directă a microorganismelor.
6.5. Circuitul fosforului Fosforul este un element absolut necesar vieţuitoarelor şi intră în
compoziţia acizilor nucleici, a unor proteine, a unor compuşi macroergici (ATP), a fosfolipidelor, a fosfozaharidelor, a fitinei etc.
Circuitul fosforului (Fig. 78) se desfăşoară în mai multe faze, după cum urmează:
1. Mineralizarea fosforului organic se desfăşoară în paralel cu mineralizarea compuşilor organici cu azot (amonificarea), sub acţiunea aceloraşi grupe de microorganisme. Unul dintre microorganismele foarte active în mineralizarea fosforului organic este Bacillus megaterium var. phosphaticus. În urma mineralizării, fosforul se eliberează sub formă de ortofosfat solubil, asimilabil de către plante şi microorganisme; în acelaşi timp, însă, ortofosfatul solubil poate trece în ortofosfaţi de Ca, Fe, Al, Mg, Cu şi Ni, care sunt insolubili.
Compuşii organici cu fosfor reprezintă 15-85% din cantitatea totală de fosfor din diferite soluri.
Între compuşii organici cu fosfor ai solului predomină fitina şi acizii nucleici. Fitina este de origine vegetală, iar acizii organici provin de la microorganisme, plante şi animale. Fitina constituie între 10 şi 80% din cantitatea totală de fosfor organic din soluri. Ea este sarea de Mg şi Ca a acidului fitic, iar descompunerea ei hidrolitică este catalizată de fitoză.
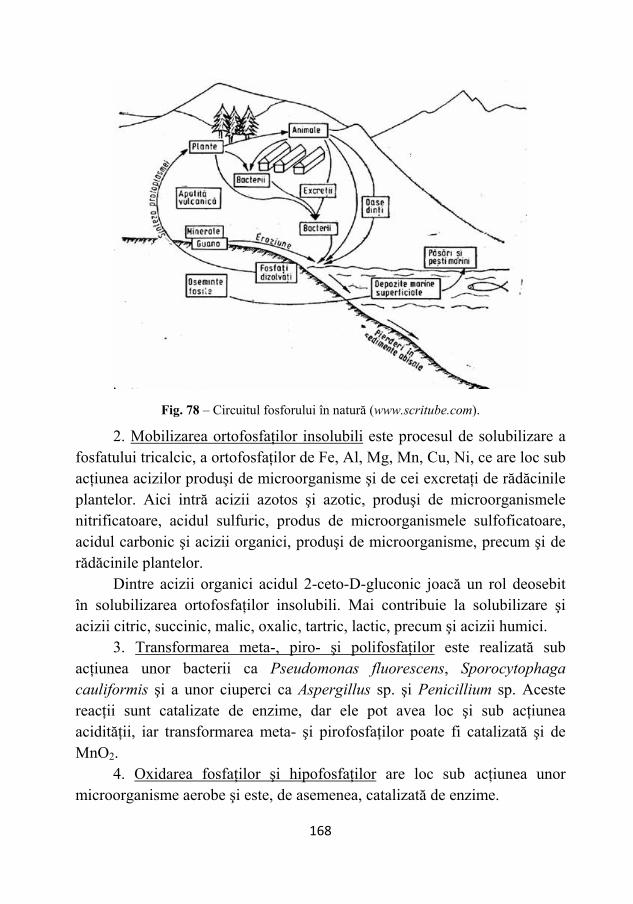
168
2. Mobilizarea ortofosfaţilor insolubili este procesul de solubilizare a
fosfatului tricalcic, a ortofosfaţilor de Fe, Al, Mg, Mn, Cu, Ni, ce are loc sub acţiunea acizilor produşi de microorganisme şi de cei excretaţi de rădăcinile plantelor. Aici intră acizii azotos şi azotic, produşi de microorganismele nitrificatoare, acidul sulfuric, produs de microorganismele sulfoficatoare, acidul carbonic şi acizii organici, produşi de microorganisme, precum şi de rădăcinile plantelor.
Dintre acizii organici acidul 2-ceto-D-gluconic joacă un rol deosebit în solubilizarea ortofosfaţilor insolubili. Mai contribuie la solubilizare şi acizii citric, succinic, malic, oxalic, tartric, lactic, precum şi acizii humici.
3. Transformarea meta-, piro- şi polifosfaţilor este realizată sub acţiunea unor bacterii ca Pseudomonas fluorescens, Sporocytophaga cauliformis şi a unor ciuperci ca Aspergillus sp. şi Penicillium sp. Aceste reacţii sunt catalizate de enzime, dar ele pot avea loc şi sub acţiunea acidităţii, iar transformarea meta- şi pirofosfaţilor poate fi catalizată şi de MnO2.
4. Oxidarea fosfaţilor şi hipofosfaţilor are loc sub acţiunea unor microorganisme aerobe şi este, de asemenea, catalizată de enzime.
Fig. 78 – Circuitul fosforului în natură (www.scritube.com).

169
5. Reducerea ortofosfaţilor este produsă de microorganisme aerobe, reacţia respectivă fiind de natură enzimatică. În prezenţa nitraţilor şi sulfaţilor, reducerea ortofosfaţilor este inhibată, deoarece nitraţii şi sulfaţii sunt utilizaţi mai uşor ca acceptori de electroni.
6.6. Circuitul potasiului Microorganismele eliberează ionii de K+ prin descompunerea sărurilor
de potasiu a acizilor organici (ex. acidul acetic sau acidul butiric). Ionii de K+ reacţionează cu anionii de HO3
-, PO42-, SO4
2-, CO32- şi formează săruri
accesibile plantelor şi microorganismelor sau sunt fixaţi de către coloizii solului.
Microorganismele mai pot elibera potasiul şi prin solubilizarea silicaţilor. Bacteriile care sunt capabile de această acţiune au fost denumite silicatbacterii. Printre acestea este şi Bacillus mucilaginosus var. siliceus, care, după unii autori, ar fi o tulpină de Bacillus circulans. Unele ciuperci, ca Aspergillus niger şi Penicillium nigrescens, pot, de asemenea, elibera K+
din silicaţi. Eliberarea K+ din silicaţi sub acţiunea silicatbacteriilor se datorează
acizilor (H2CO3 şi acizii organici), pe care aceste microorganisme îi produc. Sistemul radicular al plantelor produce, de asemenea, acizi care joacă acelaşi rol.
6.7. Transformările microbiene ale calciului Descompunerea microbiană a compuşilor organici cu calciu duce la
eliberarea Ca2+, care, ca şi K+, poate reacţiona cu anionii minerali, formând săruri solubile şi insolubile sau sunt fixaţi de către coloizii solului. Şi aici, acizii minerali şi organici, produşi fie de microorganisme sau de rădăcini, solubilizează sărurile de Ca insolubile.
6.8. Transformările microbiene ale magneziului Acestea sunt asemănătoare cu cele ale calciului, magneziul activând
unele procese microbiene din sol, îndeosebi nitrificarea şi fixarea azotului atmosferic.

170
6.9. Transformările microbiene ale microelementelor 6.9.1. Manganul Se găseşte în sol în formă bivalentă ionică (Mn2+) la pH mai mic de
5,5 şi sub formă de Mn trivalent şi, mai ales, tetravalent la pH mai mare de 8, datorită autooxidării Mn2+.
Transformările microbiene ale manganului se produc în zona valorilor de pH de 5,5-8. În condiţii aerobe acţionează unele bacterii heterotrofe din genurile Bacillus, Corynebacterium, Metallogenium, actinomicetele din genul Streptomyces, precum şi ciupercile din genurile Curvularia, Cladosporium etc. Aceste microorganisme oxidează Mn2+, precipitându-l sub formă de MnO(OH)2 (hidroxid de mangan trivalent).
Procesul de oxidare a Mn2+ poate duce la apariţia în sol a carenţei de mangan asimilabil şi are ca efect apariţia unor boli de nutriţie la plante.
În condiţii anaerobe, în prezenţa substanţelor organice uşor asimilabile, unele bacterii reduc mangan tetravalent la mangan bivalent; MnO2 serveşte ca acceptor de electroni, înlocuind O2. Totodată, acizii produşi de microorganisme, atât în condiţii aerobe, cât şi anaerobe, deviază echilibrul chimic dintre manganul bivalent şi cel tetravalent în direcţia formării Mn2+.
6.9.2. Seleniul În plante, seleniul este prezent sub formă de selenio-aminoacizi, în
care seleniul înlocuieşte sulful în structura metioninei şi Se-metilcisteină. Ambii Se-aminoacizi din resturile vegetale se mineralizează sub acţiunea microorganismelor din sol. Seleniul apare sub formă de seleniţi (SeO3
2-) şi selenaţi (SeO4
2-). Microorganismele din sol sunt capabile şi de oxidarea seleniului
metalic şi al seleniurilor (Se2-) la selenaţi. O altă transformare microbiană a seleniului în sol este reducerea
seleniţilor şi selenaţilor la seleniul metalic. Această capacitate a fost demonstrată la unele bacterii, cum sunt Clostridium, Corynebacterium şi ciuperci ca Rhizobium, precum şi la unele levuri şi mucegaiuri.

171
Compuşii cu Se (seleniţi, selenaţi, Se-aminoacizi) pot fi metilaţi sub formă de produs gazos, numit dimetilseleină (CH3-Se-CH3). Acest proces poate fi realizat de unele ciuperci, ca Aspergillus, Cephalosporium, Fusarium, Penicillium, ca şi de unele bacterii.
6.9.3. Zincul Microorganismele, în urma descompunerii resturilor vegetale,
eliberează zincul sub formă solubilă (Zn2+). Microflora solului realizează această solubilizare prin acizii minerali şi organici produşi. În sens invers, prin diminuarea acidităţii, microorganismele micşorează solubilitatea zincului. Tiobacilii oxidează sulful din ZnS la acid sulfuric şi eliberează zincul sub formă de Zn2+ care rămâne în soluţie, datorită acidului sulfuric. H2S, produs de microorganisme, îndepărtează zincul prin formarea de ZnS, care precipită.
Insolubilizarea microbiană a zincului provoacă, la pomii fructiferi, o carenţă manifestată prin micşorarea limbului frunzelor, boală ce poate fi prevenită prin tratarea solului cu sulfat de zinc (ZnSO4).
6.9.4. Cuprul Descompunerea microbiană a resturilor vegetale poate duce la
micşorarea concentraţiei cuprului solubil, datorită unei reacţii chimice în care sunt implicaţi produşii microbieni, formaţi în cursul descompunerii.
Tiobacilii pot solubiliza cuprul din minereuri sulfuroase, cum este calcopirita (CuFeS2), iar Thiobacillus ferrooxidans poate oxida atât sulful, cât şi cuprul monovalent din Cu2S. Hidrogenul sulfurat (H2S), format pe cale microbiană, poate precipita Cu2+ sub formă de CuS.
Microorganismele din sol au capacitatea de transformare şi a altor microelemente ca: Mo, V, Co, Ni, Pb, Cd, Si.

172
Capitolul 7
MICROBIOLOGIA FERMENTAŢIILOR
Microbiologia fermentaţiilor studiază caracteristicile biologice şi biochimice, precum şi condiţiile optime de activitate ale microorganismelor care produc transformări chimice în diferite substraturi, cu formarea unor cantităţi relativ mari de substanţe cu valoare utilă sau economică. De asemenea, ea studiază rolul microorganismelor în apariţia unor modificări utile a diferitelor produse alimentare, modalităţile de a dirija această activitate microbiană, ca şi posibilităţile de combatere a microorganismelor dăunătoare pentru evoluţia normală a proceselor fermentative sau pentru buna conservare a produselor alimentare.
Fermentaţiile sunt procese biochimice, determinate de enzimele unor microorganisme, cum sunt bacteriile, levurile, mucegaiurile care au proprietatea de a degrada în condiţii de anaerobioză diferiţi compuşi organici ai carbonului cu formarea de substanţe cu o condiţie chimică mai simplă. Din descompunerea anaerobă a hidraţilor de carbon rezultă o serie întreagă de produşi finali cu 1, 2, 3 sau 4 atomi de carbon, care apar, obişnuit, prin degradarea glucozei până la acid piruvic.
De cele mai multe ori, unul dintre produşii rezultaţi predomină cantitativ şi, ca atare, caracterizează procesul de fermentaţie alcoolică, lactică, butirică, propionică etc.
Termenul generic al fermentaţiei (lat. fervere = a fierbe) a fost ales pentru a indica aspectul de lichid care fierbe, caracteristic pentru faza de intensitate maximă a procesului fermentativ, aceea în care, datorită eliberării CO2 sub formă de bule de gaz, se produce o efervescenţă a mediului lichid, în special în vasele foarte mari (de exemplu, la mustul de struguri care fermentează).
Iniţial, fermentaţiile se produceau sub acţiunea micoflorei "spontane", sălbatice, care se găseşte pe suprafaţa sau în interiorul produsului supus fermentaţiei. Aceste microorganisme sunt slab sau inegal active sau pot fi în amestec cu specii dăunătoare, a căror activitate diminuează calitatea produsului final. În prezent, pentru a se preveni aceste inconvenienţe, se recurge la microorganisme selecţionate, cu ajutorul cărora se obţin produse

173
de calitate superioară, se îmbunătăţeşte randamentul şi se scurtează durata procesului tehnologic.
Microorganismele utilizate în procesele fermentative de tip industrial au următoarele calităţi, ce pot fi astfel sintetizate: • cresc rapid şi pot fi uşor cultivate în cantităţi mari, pe medii ce conţin substanţele organice corespunzătoare; • au capacitatea de a-şi menţine constant proprietăţile fiziologice, în anumite condiţii de cultură, producând uşor şi în cantităţi mari enzimele necesare pentru transformarea substratului în sensul dorit; • realizează aceste modificări, cu formarea unor produşi folositori, în condiţii relativ simple şi puţin costisitoare.
Microorganismele de utilitate industrială sunt astăzi folosite pentru procese ca: • biosinteza a diferite substanţe chimice (alcooli, acid acetic, acid lactic, acid citric, acetonă, antibiotice); • producerea pe cale fermentativă a unor alimente (produse lactate, brânzeturi), a unor băuturi alcoolice (vin, bere) şi a furajelor însilozate; • obţinerea de produse folosite în industria textilelor (fibre de in, cânepă prin topire).
Microbiologia industrială mai studiază şi aplică metodele de combatere a alterărilor microbiene a alimentelor cu ajutorul conservării prin refrigerare, sterilizare, fermentaţii şi alte procese tehnice.
7.1. Fermentaţia alcoolică Este un proces anaerob, prin care glucidele fermentescibile sunt
metabolizate prin reacţii de oxido-reducere sub acţiunea echipamentului enzimatic al drojdiei în produşi principali (alcool etilic şi CO2), produşi secundari (alcooli superiori, acizi, aldehide etc) şi energie.
Alcoolul etilic sau etanolul (CH3-CH2-OH) se obţine prin fermentarea completă, în special de către unele drojdii, a diferite zaharuri, urmată de separarea prin distilare şi purificare a alcoolului obţinut.
Fermentaţia alcoolică este una dintre cele mai importante fermentaţii industriale, deoarece alcoolul este întrebuinţat curent în industria alimentară, chimică, farmaceutică, a parfumurilor şi cauciucului.

174
Materiile prime folosite la fabricarea alcoolului sunt ieftine, abundente în natură şi sunt grupate astfel: • materii prime amidonoase, rezultate din cartof, porumb, orz, secară, care se pot utiliza după transformarea hidrolitică a amidonului din compoziţia lor în zaharuri mai simple, fermentabile de către levuri; • materii prime zaharoase, provenite din sfecla pentru zahăr sau trestia de zahăr, sucurile de fructe, melasa, rezultată ca deşeu de la fabricarea zahărului şi a leşiilor sulfitice, ce rezultă ca deşeu de la fabricarea celulozei, cu conţinut scăzut de 1-5 % zahăr; • materii prime celulolitice, rezultate din industria hârtiei, ca lemnul şi stuful, care, prin fierbere cu acizi sub presiune, se hidrolizează cu formarea de hidraţi de carbon mai simpli, fermentabili.
Înainte de folosirea lor în fermentaţiile industriale, amidonul, celuloza şi hemiceluloza trebuie hidrolizate sau convertite în zaharuri cu molecule relativ simple şi fermentabile. În acest scop, se folosesc fie preparate enzimatice, ca malţul de cereale sau enzime de natură microbiană, provenite de la bacterii şi mucegaiuri, fie anumite substanţe chimice, ca acizii diluaţi sau acţiunea asociată a unora dintre aceşti agenţi.
Microorganismele care produc fermentaţia alcoolică a zaharurilor sunt reprezentate de: • ciuperci, în principal din genul Saccharomyces; • unele bacterii ca: Zymomonas mobilis, Thermobacterium mobile, Bacillus macerans, Sarcina ventriculi, Bacillus acetosetilycus etc.
Fermentaţia alcoolică poate fi spontană, produsă de drojdiile sălbatice, prezente în natură, sau dirijată, prin introducerea în materialul de fermentat a unor suşe de drojdii selecţionate ce au capacitate superioară de fermentaţie.
Temperatura optimă de activitate este de 28-32oC. Tulpinile selecţionate au capacitatea de a suporta concentraţii mari de alcool şi de a consuma cantităţi mari de hidrocarbonate pentru sintezele lor proprii.
Aceste drojdii au un echipament enzimatic foarte bogat, graţie căruia obţin în mediu anaerob energia necesară manifestărilor vitale, prin procesul de fermentaţie alcoolică a hidraţilor de carbon, după relaţia globală:
C6H12O6 → 2 CH3CH2OH + 2 CO2 + 2 ATP (118 kJ/mol)

175
7.1.1. Utilizarea microorganismelor în vinificaţie Vinul este o băutură alcoolică nedistilată, cunoscută din cele mai
vechi timpuri care se obţine prin fermentaţia alcoolică a mustului de struguri. Fermentaţia mustului, obţinut prin zdrobirea strugurilor recoltaţi la maturitate, se face în vase speciale deschise, sub acţiunea drojdiilor din flora naturală sau a unor tulpini selecţionate.
Pe struguri, în must şi, respectiv, în vin, există un număr însemnat de microorganisme, bacterii şi ciuperci, care iau parte la procesul de fermentaţie, având fie un rol pozitiv, în cazul fermentaţiilor normale, fie un rol negativ, de "îmbolnăvire" a vinului, când declanşează unele procese fermentative, care duc la efecte nedorite.
A. Levurile (drojdii) Levurile sunt ciuperci microscopice, care au capacitatea de a produce
fermentaţia caracteristică a mediilor ce conţin zaharuri (levere = a ridica). Ele sunt grupate din punct de vedere sistematic în subîncr. Ascomycotina, fam. Saccharomycetaceae şi în subîncr. Deuteromycotina (Fungi imperfecti), fam. Cryptococca.ceae.
Răspândirea levurilor Levurile sunt larg răspândite în natură, întâlnindu-se în sol, ape, pe
plante, flori, fructe etc. Solul este considerat rezervorul principal al levurilor, unde numărul
lor este în medie de ≈ 250.000 celule pe gramul de sol. În general, se întâlnesc în stratul superficial cuprins între 2 şi 10 cm,
iar în solurile podgoriilor până la 30-40 cm, atât ca forme vegetative, cât şi ca forme de rezistenţă.
Levurile apar pe boabe în timpul maturării acestora şi sunt răspândite de insecte (în special Drosophylla sp.), prin intermediul particulelor de praf şi, nu în ultimul rând, odată cu stropii provocaţi de ploi.
Levurile ce se găsesc pe struguri ce aparţin mai multor specii, proporţia dintre ele fiind diferită în funcţie de mai mulţi factori: condiţiile climatice, starea de sănătate a recoltei, conţinutul în zahăr al mustului etc.

176
Sistematica levurilor Sistemul taxonomic al levurilor utilizat astăzi este rezultatul
dezvoltării şi integrării a trei moduri distincte de abordare. Primul dintre acestea a fost stabilit de Myen Ress în 1870, care a încercat să clasifice levurile, atunci cunoscute, pe baza formei celulelor şi a modului de reproducere. Astfel, Myen Ress a pus bazele utilizării caracterelor morfologice în taxonomia levurilor.
A doua contribuţie fundamentală pentru taxonomie a fost adusă de Emil Christian Hansen în 1883, care a evidenţiat faptul că tulpinile de levuri diferă în ceea ce priveşte capacitatea lor de a fermenta hidraţii de carbon. Practica modernă a utilizării caracterelor fiziologice, pentru diferenţierea speciilor de levuri, a fost facilitată prin această descoperire. Avantajele mult mai importante ale caracterelor fiziologice au fost subliniate de către savanţii olandezi de la şcoala lui Kluyver din Delft: L.M. Stelling-Dekker (1931) şi J. Lödder (1934, 1952).
Al treilea mod a fost iniţiat târziu, în secolul al XX-lea, de către A. Guilliermond, când a început studii asupra reproducerii sexuate şi filogeniei levurilor ascogene.
Aceste moduri distincte de abordare au constituit, de fapt, suportul studiilor clasice asupra geneticii şi al conceptului modern al sexualităţii şi filogeniei levurilor.
De la aparaţia ediţiei a doua a monografiei “The Yeasts, a taxonomic study” (J. Lödder, 1970) au fost înregistrate progrese pe mai multe direcţii, acordându-se un rol primordial în sistematizarea levurilor, caracterelor fiziologice, fie de fermentare, fie de asimilare, rezervând un rol secundar caracterelor morfologice şi extrem de reduse celor ecologice.
O clasificare de referinţă a levurilor, acceptată şi în prezent, este cea a lui N.J.W. Kregger van Rij (1984) care prezintă unele modificări faţă de cea propusă de J. Lödder (1970). Astfel, în clasificarea propusă de J. Lödder sunt cuprinse 39 de genuri şi 349 de specii, în timp ce în clasificarea actuală sunt incluse 60 de genuri şi 500 de specii.
Un studiu complet al evoluţiei principiilor de taxonomie a levurilor a fost făcut de P. Ribereau-Gayon ş.a. şi Kurtman în 1998. Pe baza acestor clasificări şi a unor rezultate proprii, M.K. Grossmann (1999) prezintă un

177
studiu referitor la metodele de identificare a levurilor, în general, şi a celor din genul Saccharomyces, în special.
Multe specii de levuri imperfecte sunt foarte asemănătoare speciilor perfecte şi diferă între ele numai prin capacitatea de a forma spori sexuaţi. Speciile care sporulează sunt indicate ca stare perfectă (telemorfă) a speciilor nesporulate şi acestea din urmă ca stare imperfectă (anamorfă) a celor sporulate. Diferenţierea generică a levurilor imperfecte se bazează pe caracteristici fiziologice, cum ar fi producerea de acid şi asimilarea inositolului.
În prezent, taxonomiştii recunosc două grupe de levuri: • levuri perfecte sau telemorfice care au stadii sexuate distincte, exprimate
fie ca ască, fie ca bazidie; • levuri imperfecte sau anamorfice sunt cele la care lipsesc stadiile sexuate
şi se reproduc numai prin înmugurire sau diviziune. Majoritatea levurilor aparţin acestui ultim grup. S-a demonstrat, totuşi,
că anumite levuri imperfecte pot fi înrudite fie cu levurile ascomicete, fie cu cele bazidiomicete, pe baza unor caracteristici, cum ar fi ultrastructura peretului celular, formarea conidiilor şi compoziţia peretului celular.
În prezent, metodele de identificarea a levurilor presupun o abordare complexă ce vizează tehnici moderne de genetică microbiană, biochimie, biologie moleculară şi imunologie.
Tehnicile de biologie moleculară au făcut posibilă compararea informaţiei inclusă în ADN, arătând că pentru fiecare organism, compoziţia în baze purinice este constantă şi caracteristică. Tehnica hibridării ADN este foarte importantă pentru a explora omologia genetică a multor grupuri de levuri. Aceasta este utilă doar pentru organisme strâns înrudite, aparţinând aceluiaşi gen sau preferabil aceleiaşi specii.
Structura primară a proteinelor enzimatice izofuncţionale poate fi diferită. Datorită acestui fapt pot coexista două levuri cu manifestări metabolice funcţionale identice, asigurate de enzime cu structuri tridimensionale similare, dar diferă în detalii ale secvenţei aminoacizilor.
S-a constatat, de asemenea, existenţa unor diferenţe semnificative atât cantitativ, cât şi calitativ, între genurile de levuri, pornind de la componenţa peretului celular. Implicarea peretului celular în taxonomia levurilor se explică prin participarea unei mari porţiuni a genomului celular la sinteza

178
acestuia, în comparaţie cu relaţiile simple de fermentaţie şi asimilaţie, care sunt sub incidenţa unui număr limitat de gene.
Deoarece, semnificaţia imunologică a antigenilor microbieni depinde de structura stereomoleculară a peretului celular, este evidentă corelaţia dintre rezultatele analizelor chimice ale peretului celular şi cele ale analizelor serologice pentru diferenţierea levurilor.
S-a demonstrat utilitatea criteriilor serologice pentru specii dificil de diferenţiat şi imunitatea din punct de vedere biologic a diviziunii, la numeroase specii a unor suşe similare biologic. Deoarece s-au constatat asemănări în structura antigenică a unor levuri fără nici o corelaţie cu alte trăsături ale acestora, taxonomia realizată exclusiv pe bază serologică nu este acceptată deocamdată. Prin aprofundarea studiilor serologice, acestea vor putea fi luate în considerare pentru elucidarea relaţiilor taxonomice şi filogenetice între levuri.
Prin îmbinarea cunoştinţelor de genetică, biochimie şi biologie celulară au fost stabilite noi tehnici de identificare a levurilor: ECP-ul (Electroforése à Champ Pulsé), ce se bazează pe separarea celor 16 cromozomi ai levurii, PCR-ul (Polymerase Chain Reaction), tehnică bazată pe amplificarea fragmentelor de ADN – specifice, urmată de separarea lor prin electroforeză, RFLP (Restriction Fragment Lenght Polymorphism), ce face, în mod egal, apel la tehnica PCR, dar secvenţele amplificate sunt degradate prin enzime de restricţie, înainte de separarea fragmentelor prin electroforeză şi tehnica AFLP (Amplified Fragment Lenght Polymorphism). Metoda AFLP de identificare a levurilor presupune ca fragmentele ADN ale levurii degradate prin enzime de restricţie, să fie amplificate prin PCR şi în final separate prin electroforeză (A. Popa şi colab., 2004).
Caracterele morfologice generale Celulele drojdiilor măsoară, în medie, 4-14 µm, iar forma poate fi
sferică, ovală sau cilindrică. În schimb, speciile pot fi monomorfe, dimorfe sau polimorfe. Forma şi dimensiunea sunt caractere de gen şi specie şi depind de vârsta acestora, starea fiziologică şi condiţiile de cultivare.
Formele caracteristice ale unor genuri cu importanţă deosebită: • formă ovală - Saccharomyces sp.; • formă sferică - Torulopsis sp.;

179
• formă apiculată – Kloeckera sp..; Hanseniospora sp.; • formă cilidrică - Candida sp.; • formă de sticlă – Saccharomycoides sp.
Cultivate în medii lichide, drojdiile produc tulbureala mediului, formarea de spumă, apoi depunerea de sedimente şi limpezirea supernatantului.
Pe medii solide, se formează colonii cu caractere morfologice specifice, având diametrul cuprins între 0,2 şi 2 cm, cu aspect cremos, lucioase sau mate, în general de culoare alb-crem. Unele formează colonii roşu-portocaliu, datorită pigmenţilor carotenoizi (Rhodotorula sp.).
Structura celulei la levuri Drojdiile prezintă o celulă de tip eucariot, care se diferenţiază puţin
faţă de celula animală (Fig. 79). Comparativ cu celula vegetală, nu prezintă cloroplaste şi înveliş celulozic.
Cunoaşterea organizării celulare la drojdii este necesară pentru a înţelege potenţialul funcţional al diferitelor componente.
1. Peretele celular are o grosime de cca.. 25 nm şi reprezintă 5-15 % din masa uscată a celulelor. Are un aspect lamelar şi este alcătuit din două straturi:
- stratul extern, are o suprafaţă rugoasă şi prezintă în anumite zone cicatrici mugurale; în compoziţia stratului extern predomină mananul cuplat cu proteine; încărcătura electrică este, în mare parte, negativă, cu rol important în stabilirea dispersiilor de drojdii şi în comportarea celulelor în timpul fermentaţiei.
- stratul intern are o suprafaţă ornamentată cu riduri proeminente, constituite din glucan, care formează complexe cu proteine şi asigură rigiditatea sau elasticitatea peretelui celular.
Funcţiile peretelui celular sunt: • sediul unor enzime, cum ar fi invertaza, fosfataza şi permeaza, implicate în biosinteza peretelui celular şi în procesul de transfer al substanţelor; • asigură forma celulei; • asigură protecţia faţă de factorii de mediu; • participă la creşterea şi reproducerea celulară.

180
2. Membrana citoplasmatică (plasmalema) reprezintă un strat lamelar cu o grosime de 8-9 nm, care delimitează protoplastul de faţa internă a peretelui celular. Este construită din două straturi lipidice, în care moleculele de lipide sunt orientate cu partea hidrofobă spre exterior şi cu partea hidrofilă spre interior.
Funcţiile plasmalemei sunt de: • barieră osmotică cu permeabilitate selectivă; • sediu al enzimelor ce reglează procesul de creştere şi înmulţire.
3. Citoplasma este un sistem coloidal cu un conţinut de 75-85 % apă, în care substanţele componente se află sub formă de sol sau gel.
Principalele substanţe organice sunt: • proteinele cu rol structural sau catalitic; • lipidele cu rol plasmatic; • glucidele cu rol energetic; • ARN (mesager, de transfer, ribozomal), dar şi ARNK (killer), implicat în sinteza unor proteine cu acţiune toxică pentru celelalte sensibile; • ADN extracromozomial din plasmide.
4. Incluziunile reprezintă forma sub care se acumulează substanţe de rezervă, în principal sub formă de: • glicogen granule care este metabolizat când celula este în stare de înfometare; • sferozomii care sunt incluziuni lipidice, ce conţin fosfolipide şi acizi graşi nesaturaţi şi care sunt metabolizaţi după liza enzimatică a membranei celulare în condiţii de malnutriţie.
5. Nucleul reprezintă spaţiul în care are loc stocarea, replicarea şi transmiterea informaţiei celulare. Are formă sferică sau ovală şi este protejat de o membrană nucleară cu pori. În interior se află nucleoplasma, iar la exterior se continuă cu reticulul endoplasmatic.
6. Mitocondriile sunt organite care ocupă cca.. 25 % din volumul citoplasmei, fiind în număr de 10-15/celulă. Au rol în ciclul Krebbs şi în sinteza ATP.
7. Vacuolele sunt formaţiuni sferice, care apar în faza staţionară de creştere a celulei, şi conţin apă, în care se solubilizează aminoacizi, proteine, enzime. Are rol de reglare a presiunii osmotice, de menţinere a stabilităţii chimice a citoplasmei, dar şi în hidroliza unor proteine şi lipide.
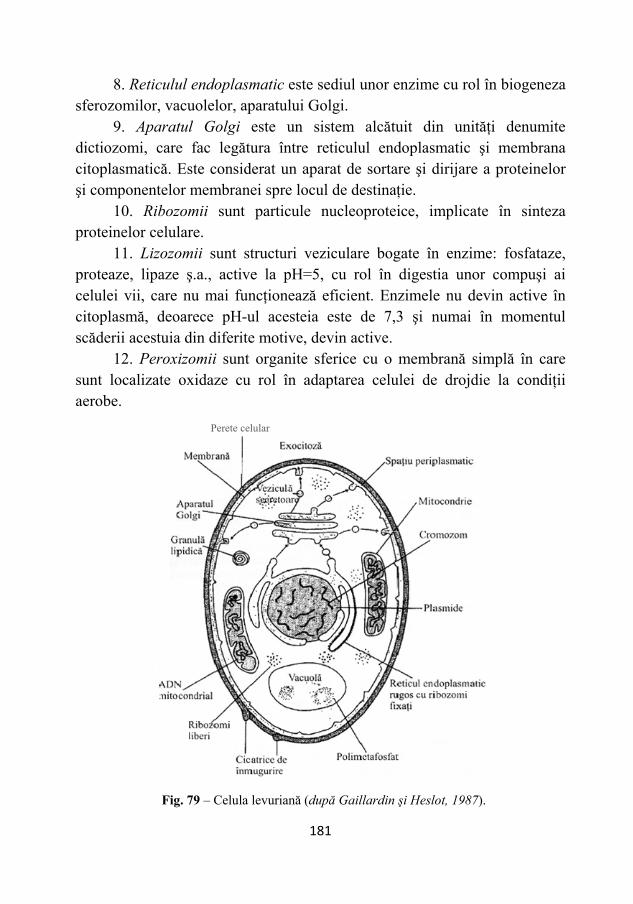
181
8. Reticulul endoplasmatic este sediul unor enzime cu rol în biogeneza sferozomilor, vacuolelor, aparatului Golgi.
9. Aparatul Golgi este un sistem alcătuit din unităţi denumite dictiozomi, care fac legătura între reticulul endoplasmatic şi membrana citoplasmatică. Este considerat un aparat de sortare şi dirijare a proteinelor şi componentelor membranei spre locul de destinaţie.
10. Ribozomii sunt particule nucleoproteice, implicate în sinteza proteinelor celulare.
11. Lizozomii sunt structuri veziculare bogate în enzime: fosfataze, proteaze, lipaze ş.a., active la pH=5, cu rol în digestia unor compuşi ai celulei vii, care nu mai funcţionează eficient. Enzimele nu devin active în citoplasmă, deoarece pH-ul acesteia este de 7,3 şi numai în momentul scăderii acestuia din diferite motive, devin active.
12. Peroxizomii sunt organite sferice cu o membrană simplă în care sunt localizate oxidaze cu rol în adaptarea celulei de drojdie la condiţii aerobe.
Perete celular
Fig. 79 – Celula levuriană (după Gaillardin şi Heslot, 1987).

182
Principalele levuri întâlnite în vin I. Levuri sporogene 1. Saccharomyces ellipsoideus (sin. Sacch. cerevisiae) prezintă
importanţa cea mai mare pentru fermentaţia mustului. Levurile acestei specii au formă elipsoidală sau globuloasă (Fig. 80).
La fermentarea mustului prezintă un randament ridicat, realizând, în medie, 1 ml alcool pur din 1,7 g glucoză consumată, şi rezistă până la concentraţii de 16 % vol. alcool, uneori chiar mai mult.
Temperatura optimă este de 20-25oC şi suportă concentraţii de până la 250-300 mg/l SO2 total.
După declanşarea fermentaţiei, ele pun stăpânire rapid pe mediu, pentru ca la sfârşitul procesului, numărul lor să scadă foarte mult.
2. Saccharomyces oviformis se prezintă sub formă ovală, uneori
rotundă. Această levură este specifică regiunilor calde, unde strugurii realizează concentraţii ridicate în zahăr. Prezintă acelaşi randament în alcool, dar are o putere alcooligenă mai mare decât Sacch. Ellipsoideus, depăşind, uneori 18 % vol. alcool.
Specia devine dominantă spre sfârşitul fermentaţiei şi suportă concentraţii de până la 300 mg/l SO2 total şi 100 mg/l SO2 liber.
Fig. 80 – Saccharomyces ellipsoideus (Sacch. cerevisiae)

183
Este considerată o bună levură de finisaj la prepararea vinurilor seci, obţinute din musturi bogate în zahăr. În schimb, trebuie inactivată sau înlăturată din vinurile demiseci, demidulci şi dulci, întrucât majoritatea accidentelor de refermentare se datorează acestei specii.
3. Zygosaccharomyces bailii. Specie rezistentă la concentraţii mari de SO2, putându-se dezvolta până la concentraţii de 12 % vol. alcool. Provoacă îmbolnăvirea vinurilor demiseci şi dulci sulfitate.
Contaminarea este facilitată de proprietatea ei de a fermenta cu uşurinţă fructoza, lăsând neatinsă glucoza.
Cum majoritatea levurilor fermentează mai întâi glucoza, în vinurile dulci-licoroase zahărul restant este constituit în principal din fructoză.
4. Saccharomyces chevalieri se aseamănă sub raporul activităţii fermentative cu Saccharomyces ellipsoideus dar sunt puţin întâlnite în vinuri.
5. Saccharomyces bayanus este prezentă în toate regiunile viticole, dar se întâlneşte în proporţii foarte mici la fermentarea mustului, când formează proporţii mărite de acizi volatili şi glicoli.
6. Saccharomyces italicus, Sacch. uvarum şi Sacch. florentinus sunt levuri specifice, în general, regiunilor viticole meridionale, au proprietăţi asemănătoare speciei Sacch. ellipsoideus dar cu o fermentare mai lentă decât aceasta.
7. Saccharomyces rosei este o levură prezentă constant pe struguri şi în musturi care fermentează. Are proprietatea de a fermenta lent dar regulat, cantităţi importante de zahăr, fără formarea de acizi volatili. Este folosită pentru obţinerea vinurilor cu un conţinut de până la 12 % vol. alcool, pentru vinuri cu rest de zahăr, precum şi pentru recolte atacate de mană şi putregai cenuşiu, la care obţinerea de vinuri cu aciditate volatilă scăzută este o problemă dificilă.
8. Pichia membranefaciens, P. fermentans şi Hansenula anomala sunt levuri cu putere alcooligenă slabă, încadrate în categoria levurilor patogene, care, în condiţii oxidative, provoacă vinurilor boala denumită "floare". De asemenea, afectează calitatea vinurilor prin proporţii ridicate de esteri, acetat de etil, acetat de amil, pe care îi formează şi care imprimă mirosuri neplăcute.

184
9. Saccharomycoides ludwigii este o levură de dimensiuni mari, de formă apiculată, rar întâlnită pe struguri şi în musturi normale. Excepţional de rezistentă la SO2 (700-1000 mg/l), este specifică musturilor păstrate cu doze mari din acest antiseptic. Este o levură patogenă pentru vinurile demiseci sau dulci, aflate în butoaie sau sticle, cărora le provoacă refermentări.
10. Hanseniospora valbyensis, H. uvarum, H. osmophila. Levură apiculată, specifică zonelor calde, provoacă declanşarea fermentaţiei, aşa cum în regiunile răcoroase acest proces este declanşat de Kloeckera apiculata. Are capacitate mare de a asimila azotaţi.
II. Levuri nesporogene 1. Torulopsis stellata este o levură prezentă pe strugurii stafidiţi, în
urma apariţiei mucegaiului nobil. Prezintă o rezistenţă mai mare decât levurile apiculate la concentraţii de SO2 şi alcool, dar mai scăzută decât Sacch. ellipsoideus.
2. Brettanomyces intermedius. Numele genului se datorează acţiunii unor specii de a imprima berii britanice gust şi aromă caracteristice, în urma unor procese secundare de fermentaţie.
Speciile acestui gen sunt întâlnite în proporţii mici în musturi şi vin. Mai frecvent se întâlnesc în vinurile bolnave de floare, oţetite. Prezenţa lor favorizează formarea de acetat de etil şi alţi compuşi ce dau mirosuri intense, cu iz de şoarece.
3. Candida vini (sin. Candida micoderma, Mycoderma vini). Speciile acestui gen sunt cunoscute sub denumirea de "levuri de floare", producând boala denumită floarea vinului, caracterizată prin formarea unei pelicule la suprafaţa vinurilor slab alcoolice, păstrate în vase incomplet pline.
4. Kloeckera apiculata, cunoscută şi sub denumirea de levură sălbatică, este extrem de răspândită pe toate fructele în regiunea cu climat temperat-răcoros. Prezintă celule sub formă de lămâie, având unul sau ambele capete ascuţite (Fig. 81).
În practică, sunt cunoscute sub denumirea de levuri apiculate. Acestea apar pe bace la faza de pârgă, iar la maturitatea deplină a strugurilor reprezintă 99 % din totalul levurilor existente în acest moment.

185
Ele declanşează fermentaţia mustului, având o capacitate foarte mare de înmulţire, randamentul în alcool este mai scăzut decât al levurilor eliptice. Ele consumă pentru 1 ml alcool pur 2,1 g glucoză.
Puterea alcooligenă este redusă, realizând 4-5 % vol. alcool, concentraţie maximă până la care această specie poate rezista. Prezintă o sensibilitate mai mare decât levurile eliptice la SO2.
5. Rhodotorula mucilaginosa, Rh. glutinis şi Rh. rubra sunt levuri
denumite şi mucilaginoase, deoarece produc mucilagii în musturile şi vinurile tinere, sărace în aciditate, şi formează pigmenţi carotenoizi galbeni până la roşii.
Se întâlnesc în pivniţele de vinuri, în care formează, împreună cu unele bacterii şi alte ciuperci, depuneri mucilaginoase pe pereţii umezi. Deşi sensibile la alcool, prezintă o rezistenţă mare la temperaturi ridicate (50-60oC) şi la acţiunea substanţelor alcaline (fosfat trisodic 1 %). Rezistă la acţiunea acidului boric până la concentraţia de 3 %.
B. Mucegaiurile Ciupercile care formează colonii pufoase, vizibile cu ochiul liber, sunt
denumite în practică mucegaiuri. Mucegaiurile, obişnuit aerobe, sunt foarte răspândite în natură,
întâlnindu-se aproape în toate mediile, datorită capacităţii mari de înmulţire
Fig. 81 – Kloeckera apiculata

186
şi de răspândire, precum şi a rezistenţei mari la condiţiile mai puţin prielnice de mediu.
Sunt frecvent întâlnite în plantaţiile viticole, pe frunze, boabe, în sol, pe utilajele şi pereţii pivniţelor, precum şi la suprafaţa mustului, în primele etape de vinificaţie.
Mucor racemosus produce în plantaţii, pe vreme ploioasă, infecţia strugurilor. În cazul în care se află complet scufundată în must, ciuperca se prezintă sub formă de fragmente de sifonoplast; are capacitatea de a produce fermentarea mustului până la 5-7 % vol. alcool. Mucor circinelloides este o altă specie, care este folosită la fabricarea berii. Mucor piriformis oxidează parţial glucidele, cu formarea acizilor citric, oxalic, gluconic etc.
Rhizopus nigricans este frecventă pe strugurii vătămaţi, în pivniţe sau pe utilajele folosite în vinificaţie, formând pe suprafaţa substratului degradat un mucegai de culoare negricioasă. Prezintă capacitate mică de fermentaţie.
Aspergillus niger se dezvoltă în crame şi pivniţe, pe vase vinicole, dopuri, formând un mucegai negru. Aspergillus glaucus se întâlneşte, mai frecvent, pe suporturi lemnoase din pivniţe.
Penicillium glaucum produce putrezirea boabelor incomplet maturate sau a strugurilor vătămaţi de insecte sau grindină. În cazul în care contaminarea se extinde devreme, se pot produce pierderi de recoltă importante, boabele atacate fiind complet distruse. Când atacă strugurii copţi, mustul obţinut are gust amar. În crame şi pivniţe se dezvoltă pe dopuri, vase vinicole etc., pătrunzând în lemnul doagelor. În timpul fructificării secretă esteri cu gust şi miros caracteristic de mucegai, ce se transmit vinului.
Genul Merulius cuprinde specii extrem de păgubitoare, care se dezvoltă în pivniţe, pe butoaie, căzi, pe rafturi, stelaje şi orice obiect din lemn, pe care se formează un miceliu alb, cu aspect buretos. Speciile genului Merulius formează substanţe cu miros specific neplăcut, unele atacă numai celuloza, altele numai lignina, transformând lemnul într-o masă moale, de culoare brună sau albă.
Merulius lachrymans, denumită popular ciuperca igrasiei sau buretele de casă, formează un mucegai alb-cenuşiu şi filamente izolate sau cordoane compacte.

187
Cladosporium cellare se dezvoltă pe toate substraturile lemnoase, dar şi pe pereţii pivniţelor şi chiar pe ramele de fier, grilaje, sârme etc., datorită capacităţii acesteia de a provoca microcondensarea pe suprafaţa hifelor sale, a moleculelor compuşilor organici şi anorganici în stare de vapori, existenţi în aerul din pivniţă.
În unele pivniţe, acest mucegai se dezvoltă pe pereţi într-un strat gros cu aspect de catifea şi, datorită capacităţilor sale de a purifica aerul şi de a regla umiditatea atmosferică, realizează condiţii deosebite pentru maturarea şi învechirea vinurilor.
Botryotinia fuckeliana prezintă ca formă conidiană Botrytis cinerea şi produce "putregaiul cenuşiu al strugurilor".
Această boală apare la intrarea în pârgă şi este favorizată de crăparea sau rărirea pieliţei boabelor, producţia în unii ani putând fi diminuată cu 60-90 %. Vinurile obţinute din strugurii atacaţi de această ciupercă sunt slab alcoolice, neplăcute la gust, cu miros specific de mucegai şi care se îmbolnăvesc uşor.
În unele toamne secetoase, cu dimineţi nelipsite de rouă, apare aşa- numitul "putregai nobil", care conduce la obţinerea unui must de înaltă calitate. În acest caz, scade mult conţinutul boabelor în apă şi creşte concentraţia în zahăr (de la 200-220 până la 400-500 g/l). Producţia scade mult, uneori cu peste 40 %, dar se obţin vinuri cu o aromă buchet deosebită şi foarte plăcută.
C. Bacteriile vinului Bacteriile prezintă o mare importanţă pentru calitatea vinurilor. Unele
dintre ele, în anumite condiţii, pot contribui prin activitatea lor la
Fig. 82 – Botryotinia fuckeliana

188
îmbunătăţirea acesteia; cele mai multe pot provoca vinurilor transformări nedorite şi chiar îmbolnăviri grave.
În perioada recoltării strugurilor, pe pieliţa boabelor, pe lângă levuri şi mucegaiuri, se găsesc şi bacterii, dar pot fi întâlnite, de asemenea, în crame şi pivniţe, îndeosebi pe suprafaţa internă a butoaielor şi cisternelor în care se fermentează mustul sau se păstrează vinul şi care nu au fost corect curăţate şi întreţinute.
După procesele pe care le declanşează, bacteriile au fost grupate în: I. Bacterii acetice, microorganisme aerobe, care se întâlnesc atât pe strugurii sănătoşi, cât şi pe cei vătămaţi, precum şi în musturi sau vinuri, mai ales când sunt păstrate în vase incomplet pline.
Bacteriile acetice care se dezvoltă în vin formează, la suprafaţa acestuia, un văl sau o peliculă, care, uneori, poate fi alburie, subţire, aproape transparentă, cu reflexe irizante şi care poate urca rapid pe pereţii vasului.
Alteori, acest văl este mai gros, uleios, hidrofob şi se colorează, pe măsura îmbătrânirii, în brun.
Într-un al treilea caz, vălul format atinge mai mulţi milimetri grosime, este vâscos, greu de desfăcut şi, care, cu timpul, cade la fundul vasului, cunoscut şi sub numele de cuib de oţet.
Genul Acetobacter. Acest gen grupează specii cu celule sub formă de
bastonaşe, dispuse izolat, în perechi sau lanţuri, ce sunt imobile sau prevăzute cu cili dispuşi peritriche.
Speciile genului Acetobacter produc oxidarea etanolului până la acid acetic, la un pH de aproximativ 4, precum şi a acidului acetic a lactaţiilor şi aminoacizilor. Ciclul Krebs este prezent şi activ. Se dezvoltă la temperaturi cuprinse între 5 şi 42oC, cu un optim de 30oC.
Speciile cele mai importante sunt: Acetobacter aceti; Acetobacter pasteurianus şi Acetobacter peroxydans.
Genul Gluconobacter cuprinde bacterii asemănătoare din punct de
vedere morfologic cu speciile genului Acetobacter. Singurul criteriu de diferenţiere îl reprezintă flagelaţia, când aceasta există. Astfel, speciile

189
genului Gluconobacter sunt lofotriche (3-8 cili la fiecare pol), faţă de ciliaţia peritriche la Acetobacter.
Produc oxidarea moderată a etanolului până la acidul acetic la un pH=4,5. Temperatura optimă de dezvoltare este 25-30oC, dar variaţia acesteia poate fi cuprinsă între 7 şi 41oC. Fiziologic, se deosebeşte de genul Acetobacter prin următoarele proprietăţi: • ciclul Krebs nu este complet; • nu oxidează acidul acetic şi lactaţii până la CO2.
II. Bacterii lactice
Reprezentanţii acestui grup sunt facultativ-anaerobi şi participă la o serie de transformări în vin şi anume: • transformarea glucidelor în acid lactic şi alţi produşi secundari (glicerol, alcool etilic, acid acetic etc); • fermentaţia malolactică; • degradarea acidului tartric, citric; • degradarea glicerolului şi a altor componenţi.
Genul Pediococcus cuprinde bacterii Gram (+) cu aspect sferic,
grupate sub formă de diplococi sau tetracoci. Sunt nesporulate, imobile, homofermentative. Fermentează glucidele, iar acidul malic numai la pH ridicat. Nu degradează acidul citric şi tartric, din acest motiv nu sunt considerate dăunătoare.
Prezintă două specii importante: - Pediococcus cerevisiae; - Pediococcus pentosacens. Genul Leuconostoc cuprinde bacterii Gram (+) cu celule de formă
sferică, ovoidă sau alungită, dispuse sub forma unor şiraguri de mărgele, ca şi streptococii. Produc fermentaţia malolactică numai în vinuri foarte acide, nesulfitate sau cu un conţinut redus de SO2.
Principalele specii: - Leuconostoc gracile este specia cea mai frecventă în vinuri.
Descompune energic acidul malic şi, în măsură mai mică, acidul citric. - Leuconostoc oinos

190
Genul Lactobacillus grupează bacterii Gram (+) şi cuprinde : - specii homofermentative: Lactobacillus acidophillus, Lactobacillus
helveticus. - specii heterofermentative: Lactobacillus plantarum, Lactobacillus
casei, Lactobacillus brevis. 7.1.2. Utilizarea microorganismelor în industria berii În procesul de fermentaţie a mustului de malţ, obţinut din cariopsele
de orz sau orzoaică, se utilizează două tipuri diferite de levuri: • levuri de fermentaţie inferioară - au proprietatea de a fermenta în condiţii submerse, iar la final levurile se depun la baza recipientului. Ex: Saccharomyces carlsbergensis şi Saccharomyces uvarum. • levuri de fermentaţie superioară - au proprietatea de a fermenta la suprafaţa mustului. Ex: Saccharomyces cerevisiae.
7.2. Fermentaţia lactică Fermentaţia lactică este un fenomen cunoscut încă din antichitate, ce a
fost explicat de L. Pasteur odată cu descoperirea bacteriilor lactice. Acidul lactic rezultat din această fermentaţie este un produs foarte util în medicină, industria alimentară, la tăbăcirea pieilor, ca mordant în industria textilă etc. După natura produşilor finali din fermentaţie, bacteriile lactice (Fig. 83) pot fi împărţite în două grupuri:
I. Bacterii lactice homofermentative – transformă substanţele glucidice şi formează numai acid lactic cu un randament de 90-100%. Dintre acestea, cele mai importante sunt: Lactobacillus delbrueckii ssp. delbrueckii, Lactobacillus delbrueckii ssp. lactis, Lactobacillus delbrueckii ssp. bulgaricus, Lactobacillus helveticus, Lactobacillus acidophilus, Pediococcus cerevisiae, Pediococcus acidi lactici, Streptococcus lactis, Streptococcus salivarius ssp. thermophilus etc.
II. Bacterii lactice heterofermentative – formează cantităţi mici de acid lactic şi cantităţi însemnate de acizi volatili şi CO2. Cele mai obişnuite sunt:

191
Lactobacillus buchneri, Lactobacillus casei, Lactobacillus brevis Lactobacillus sake, Lactobacillus plantarum, Lactobacillus fermenti, Lactobacillus bifidum, Leuconostoc mesenterioides ssp. cremoris, Leuconostoc paracitrovorum ş.a.
Pentru prepararea acidului lactic pe cale fermentativă se folosesc, ca
materie de bază, amidonurile, melasele, cerealele, zerul etc. Amidonul este, în prealabil, zaharificat cu ajutorul malţului din sămânţa de orz germinată artificial. Plămada dulce, care conţine maltoză zaharificată de malţ, se încălzeşte la 80 ̊C pentru omorârea microflorei, după care este însămânţată cu o cultură de bacterii termofile Lactobacillus delbrueckii ssp. delbrueckii.
Fermentaţia durează 8-10 zile, timp în care microorganismele transformă în acid lactic până la 98% din zahărul fermentat. Această fermentaţie se produce printr-un mecanism similar cu cel al fermentaţiei alcoolice, adică tot prin formarea iniţială a unei aldehide (metilglioxal), urmată de transformarea acesteia în acid, realizată pe cale enzimatică în mai multe trepte succesive:
1. C6H6O6 → 2 CH3-CO-CHO + 2H2O glucoză metilglioxal 2. CH3-CO-CHO + 2H2O → 2 CH3-CHOH-COOH metilglioxal acid lactic
Fig. 83 – Lactobacillus sp. (www.wikipedia.com)

192
Pentru a asigura multiplicarea mai departe a microorganismelor,
acidul lactic este neutralizat cu hidroxid de calciu pe măsură ce se formează, după reacţia :
acid lactic lactat de calciu Produsul final este lactatul de calciu. Dacă acidul lactic nu ar fi neutralizat, fermentaţia s-ar opri, întrucât
bacterile lactice nu suportă aciditatea. Din lactat, acidul lactic este separat prin adăugarea de H2SO4 , care precipită calciul sub formă de CaSO4. Soluţia diluată de acid lactic obţinută este evaporată în vid până la concentraţia de 50-80% acid lactic.
Fermentaţia lactică are numeroase aplicaţii practice în industria alimentară, la prepararea laptelui acru, a iaurtului şi kefirului, la conservarea legumelor şi la prepararea nutreţului însilozat.
7.2.1. Rolul microrganismelor în obţinerea produselor lactate Iaurtul este produsul obţinut prin fermentarea laptelui cu o cultură
mixtă de două specii de bacterii lactice, Streptococcus salivarius ssp. thermophilus şi Lactobacillus delbrueckii ssp. bulgaricus în raport de 1:1. Rolul streptococilor şi lactobacililor la obţinerea iaurtului constă în acidifierea laptelui, sinteza de compuşi de aromă, dezvoltarea texturii şi a vâscozităţii.
Bioiaurtul conţine, pe lângă bacteriile specifice, şi Lactobacillus acidophillus şi Bifidobacterium bifidum. Acest produs are efect benefic suplimentar, deoarece are efect de ameliorare a creşterii bacteriilor, care, în

193
mod normal, fac parte din microbiota intestinală şi ar putea ajuta la protecţia faţă de unele boli, cum ar fi, cancerul şi bolile coronariene. Bifidobacteriile se dezvoltă la o temperatură optimă de 36-38oC.
Pentru fabricarea untului se poate utiliza smântână dulce sau fermentată, care este sterilizată prin pasteurizare şi, după răcire, este inoculată cu culturi mixte de bacterii lactice, aromatizante. Dintre acestea fac parte Lactococcus lactis ssp. cremoris, Lactococcus lactis biovar. diacetylactis şi Leuconostoc mesenteroides ssp. cremoris, Lactococcus lactis ssp. lactis.
Pentru obţinerea brânzeturilor se poate folosi lapte crud, în care bacteriile lactice trebuie să reprezinte peste 50 % din totalul microbiotei şi care se pasteurizează, având grijă să nu se producă denaturarea proteinelor din lapte.
După răcire, se face inocularea cu monoculturi sau culturi mixte specifice, iar etapele tehnologice urmăresc să dirijeze activitatea utilă a culturilor starter.
Principalele culturi de bacterii lactice folosite drept culturi de producţie pentru diferite tipuri de brânzeturi în România sunt: • brânză proaspătă de vaci: Lactococcus lactis, Lactococcus lactis var. diacetylactis. • telemea: Lactococcus lactis, Lactobacillus casei. • caşcaval: Lactococcus lactis, Lactobacillus casei, Streptococcus salivarius ssp. thermophilus. • şvaiţer: Streptococcus salivarius ssp. thermophilus, Lactobacillus delbrueckii ssp. bulgaricus, Lactobacillus delbrueckii ssp. lactis, Lactobacillus helveticus, Propionibacterium sp.
7.2.2. Rolul microorganismelor lactice în conservarea alimentelor
murate Una dintre aplicaţiile practice importante ale fermentaţiei lactice este
conservarea alimentelor prin murare. Astfel, varza murată se obţine prin fermentarea naturală a sucului, care conţine zahăr şi care este extras din frunze prin adăugarea de sare 2-3%, proces ce se petrece în mai multe faze:

194
• Faza preliminară se petrece la începutul fermentaţiei, când se multiplică microflora normală de bacterii Gram pozitive cu preponderenţa celor heterofermentative de pe suprafaţa verzei, din vas şi din apă. Preponderentă este în această fază bacteria Leuconostoc mesenteroides, care descompune incomplet zahărul din varză, cu formarea de acizi lactic şi acetic plus diferiţi esteri, care dau un gust şi o aromă particulare.
• Faza primară sau principală începe când cantitatea de acid din mediu atinge 0,7%. Se dezvoltă sub influenţa bacteriilor homofermentative, care produc mari cantităţi de acid lactic; sub influenţa acestor concentraţii, bacteriile îşi încetează acţiunea. Fermentaţia este terminată de Lactobacillus brevis (specie heterofermentativă), care produce acid lactic, alcool etilic, metilen şi CO2 şi care este rezistentă la un pH foarte scăzut. Întregul proces durează 7-8 zile la temperatura optimă de 21°C şi încetează când cantitatea de acid acetic a ajuns la 1,5%.
• Faza secundară se întâlneşte când fermentaţia nu decurge în condiţii normale şi unele bacterii, drojdii şi mucegaiuri produc denaturări şi alterări ale verzei murate ca înmuierea, putrezirea, băloşirea (acoperirea cu un strat vâscos) şi apariţia de pete roşii sau negre, însoţite de un miros dezagreabil.
În cazul castraveţilor muraţi în saramură se petrec trei tipuri de fermentaţii şi anume: • fermentaţia determinată de Lactobacillus plantarum, din care rezultă acid lactic cu efect prezervant; • fermentaţia produsă de Aerobacter sp. cu eliminarea de apă şi CO2; • fermentaţia sub acţiunea unor levuri cu formarea de CO2 şi alcool.
Microorganismele se găsesc pe castraveţi şi în mediul ambiant şi produc fermentarea zaharurilor extrase din soluţie de saramură în 2-6 săptămâni. Unele microorganisme provoacă alterarea castraveţilor muraţi. Bacillus subtilis produce înmuierea, datorită transformării protopectinei în pectină solubilă, şi Bacillus subtilis var. niger produce înnegrirea acestora.
7.2.3. Rolul microrganismelor în producerea furajelor însilozate Însilozarea sau murarea este cel mai bun mijloc de conservare a
furajelor suculente, mai ales a celor verzi, cu păstrarea calităţii nutritive.

195
Procesul se bazează pe fermentaţia lactică în condiţii de anaerobioză a zaharurilor ce intră în constituţia plantelor, cu formarea de CO2, apă şi o cantitate variabilă de acid lactic.
Plantele destinate însilozării sunt recoltate în perioada când produc cea mai mare cantitate de masă verde şi conţin maximum de substanţe nutritive. Masa vegetală este tocată şi aşezată în silozuri construite special sau în gropi, unde se păstrează pentru îndepărtarea aerului.
În prima perioadă, oxigenul rămas în spaţiile din siloz este consumat de celulele vegetale şi este înlocuit cu CO2, care creează condiţii optime pentru fermentaţia anaerobă. Paralel cu acest fenomen şi mai ales pe măsură ce diminuează procesele enzimatice din celulele plantelor supuse conservării, începe dezvoltarea florei epifite, formată din Pseudomonas fluorescens şi a unor bacterii din sol ca Bacillus subtilis, Bacilllus megaterium, Clostridium sporangenes şi altele. Multiplicarea acestor bacterii încetează pe măsura acumulării acidului lactic, produs de bacteriile lactice, prezente şi ele în frunze.
Deoarece, la început, cantitatea naturală de acid lactic este mică, se adaugă în masa însilozată culturi de Lactobacillus delbrueckii ssp. delbrueckii şi Lactobacillus plantarum, precum şi o soluţie de melasă 1% pentru grăbirea fermentaţiei. În ţara noastră există un biopreparat, denumit ‘Lactobacterin’, care este o cultură de Lactobacillus plantarum. După 2-3 săptămâni, când reacţia mediului a atins pH = 4,4, fermentaţia încetează.
Silozul astfel obţinut poate fi păstrat chiar până la 3 ani, cu condiţia să fie ferit de mucegaiurile care descompun acidul lactic.
Dacă însilozarea s-a făcut defectuos, se produce, pe lângă fermentaţia lactică, şi o fermentaţie butirică, şi furajele capătă un miros neplăcut, fiind refuzate de animale.
7.3. Fermentaţia acetică Acest proces este, de fapt, impropriu numit fermentaţie, deoarece
acidul acetic se obţine în condiţii de aerobioză, prin oxidarea alcoolului în prezenţa oxigenului gazos de către bacterii, aparţinând genului Acetobacter.
Deşi oţetul este cunoscut din cele mai vechi timpuri, abia în 1837, Kütying a emis ipoteza că transformarea alcoolului în acid acetic este

196
produsă de microorganisme. Ulterior, în 1868, Pasteur a confirmat ipoteza şi a descris, pentru prima dată, microorganismul Mycoderma aceti.
Dezvoltarea spontană a bacteriilor acetice în timpul păstrării unor produse alcoolizate, ca vinul sau berea, este dăunătoare, deoarece produce oţetirea băuturilor .
Pentru prepararea industrială a acidului acetic se folosesc culturi de bacterii selecţionate, care pot produce până la 10% acid acetic de calitate.
Bacteriile acetice sunt microorganisme aerobe, din genul Acetobacter, care trăiesc la suprafaţa mediilor alcoolizate, formând o peliculă bacteriană groasă (Fig. 84).
Cele mai folosite bacterii în producerea oţetului sunt grupate în patru
categorii, funcţie de mediul din care au fost izolate: I. Bacterii acetice izolate din plămadă:
• Gluconobacter oxidans, izolată din sticle de bere fermentată; • Gluconobacter oxidans ssp. Industrium, izolată din drojdia uscată.
II. Bacterii acetice din bere: • Acetobacter aceti suportă până la 11% alcool şi poate produce 4-6% acid acetic; • Acetobacter pasteurianum suportă 9,5% alcool şi produce 6,2% acid acetic; • Termobacterium aceti este prezentă în depozitele de bere; • Acetobacter kutzingianum este izolată din sake.
III. Bacteriile izolate din vin cuprind câteva specii ca:
Fig. 84 – Acetobacter aceti (www.vinegarman.com)

197
• Acetobacter orleanensis formează un văl subţire, adeseori roz, şi care are tendinţe de a se înălţa pe pereţii vasului. Poate produce 9,3% acid acetic. Oţetul provenit din această specie este parfumat şi limpede. • Acetobacter ascendens este izolată din oţetul de vin; formează un voal ce se rupe uşor; • Acetobacter xylinum este izolată din oţet de vin; suportă alcool şi produce 4,1% acid acetic.
IV. Bacterii acetice fermentative rapide: • Acetobacter acetigenu se prezintă ca celule rotunde, ce nu formează lanţuri şi produce numai 3% acid acetic. • Acetobacter schützenbachii se prezintă sub formă de bastonaşe de 1,6-3,6 µm lungime x 0,3-0,4 µm grosime şi poate produce 11-14% acid acetic. • Acetobacter curvum produce un voal subţire, uşor de rupt, cu numeroase pete albe.
Reacţia probabilă de transformare a alcoolului în acid acetic constă, iniţial, în dehidrogenarea alcoolului cu formare de acetaldehidă; ulterior, acetaldehida fixează o moleculă de apă, formând hidrat de acetaldehidă, iar prin hidrogenare rezultă acid acetic.
În oţetul produs pe cale naturală mai apar urme de diferiţi esteri, ca acetatul de etil şi cantităţi mici de alcool, glicerină, uleiuri volatile, care dau aroma plăcută caracteristică.
Materia primă folosită în producţie este orice produs capabil să producă prin fermentaţie alcool etilic, care, ulterior, este supus fermentaţiei acetice. Se folosesc, deci, vin, bere, sucuri de fructe, siropuri de zahăr şi materiale amidonoase hidrolizate iar calitatea produsului final este în funcţie de compoziţia materiei prime utilizate.
Există mai multe metode de preparare a oţetului: 1. Metoda franceză (orleaneză sau lentă) foloseşte ca materie primă
un amestec de vin cu 4% alcool şi oţet cu 2% acid acetic. Se pune în butoaie de 200 litri jumătate din volum oţet şi 10-15 l vin. După fiecare 7 zile, timp de 4 săptămâni, se adaugă câte 10-15 l vin. Adaosul de oţet are ca scop să împiedice dezvoltarea microflorei normale din mediu.
Producerea oţetului are loc lent, pentru că bacteriile vin în contact cu aerul numai la suprafaţa luchidului; acesta face ca în oţet să se formeze

198
substanţe aromatice, care dau acestuia un “buchet” aparte. După 5-6 săptămâni, se ajunge la concentraţia de 5-6 % acid acetic, se recoltează 10-15 litri oţet şi se adaugă o cantitate egală de vin, după care procesul se repetă.
2. Metoda germană, rapidă, foloseşte un amestec de lichid format din alcool etilic 3% şi oţet 6%, un adaos nutritiv pentru bacterii, care este introdus în butoaie de lemn.
În vas se găseşte talaj fin de lemn de fag, care măreşte suprafaţa de oxidare; se adaugă bacterii selecţionate, ca Acetobacter orleanense, Acetobacter schützenbachii, Acetobacter acetigenum, şi se asigură circulaţia aerului.
Amestecul de alcool-acid acetic este turnat pe la partea superioară a vasului, se filtrează prin stratul de talaj, unde alcoolul se oxidează, iar la partea inferioară se colectează un lichid bogat în acid acetic (9%) şi aproape lipsit de alcool. Pentru ca oxidarea să fie completă, acelaşi lichid se trece de trei ori prin stratul de talaj.

199
BIBLIOGRAFIE SELECTIVĂ
Anghel I., Toma M., Voica C., Cojocaru I. – Biologia şi tehnologia
drojdiilor, Vol. I - 1989, Vol. II - 1991, Vol. III -. Ed. Tehnică, Bucureşti, 1993
Baker H. F., Rosalind M. Ridley – Prion diseases, Ed. Humana Press, New Jersey, USA, 1996
Baker S., Nicklin Jane, Khan N., Killington R. – Microbiology, Ed. Taylor & Francis, Oxford, Marea Britanie, 2007
Buchanan R.E., Gibbons N.E. – Bergey’s manual of determinative Bacteriology, Ed. Wiliams, USA, 1975
Cardon Zoe G., Whitbeck Julie L. – The rhizosphere–an ecological perspective, Ed. Academic Press, San Diego, USA, 2007
Carp-Cărare M. – Microbiologie, Institutul Agronomic „Ion Ionescu de la Brad” Iaşi, 1991
Coman I., Mareş M. – Micologie medicală aplicată, Ed. Junimea, Iaşi, 2000
Constantinescu O. – Metode şi tehnici în micologie, Ed. Ceres, Bucureşti, 1974
Dan Valentina – Microbiologia produselor alimentare, Ed. Alma, Galaţi, Vol. I, II, 1999, 2000
Delfini C. – Scienza e tecnica di microbiologia enologica, Ed. Il Lievito, Italia, 1995
Eliade Gh., Ghinea L., Ştefanic Gh. - Bazele biologice ale fertilităţii solului, Edit. Ceres, Bucureşti, 1983
Gaillardin C., Heslot H.– La levure, Ed. La Recherche, 188 (18) 586-596, 1987
Graham H. Fleet– Wine microbiology and biotechnology, Ed. Harwood Academic Publishers, Switzerland, 1993
Grodzinsky A. M.– Allelopathy in soil sickness, Ed. Scientific Publisher (India), Jodhpur, 2006
Hatman M.– Microbiologie - lucrări practice, Universitatea Agronomică Iaşi, Centrul de multiplicare, Iaşi, 1990

200
Hatman M., Ulea E. – Microbiologie – Curs, Universitatea Agronomică Iaşi, Centrul de multiplicare, Iaşi, 1993
Hughes M.N., Poole R.K., 1989 – Metals and Micro-organisms, Ed. Chapman & Hall, Londra, New York.
Kingsley R.S. – Introductory-Plant biology, Ed. Wm. C. Brown Publishers, U.S.A, 1991
Kiss Şt., Ştefanic Gh., Pasca Daniela, Drăgan-Bularde Zborovschi, Crişan R. - Enzimologia mediului înconjurător, Ed. Ceres, Vol. I - 1991, Vol. II - 1993.
Korn R. – Microbiologia laptelui şi produselor lactate, Ed. Ceres, Bucureşti, 1989
Leclerc H. – Microbiologie, Ed. Doin Deren & Cie, France, pag. 341-531, 1976
Lödder J., Kreger Van Rij N. J. W. – The yeasts, a taxonomic study, Ed. North Holland Publishing Company, Amsterdam, 1952
Lödder J., Kreger Van Rij N. J. W. – The yeasts, a taxonomic study-second edition, Ed. North Holland Publishing Company, Amsterdam, 1970
Lwoff A., Horne R., Tournier P. – A system of viruses. Cold Spring Harb Symp Quant Biol., 27, 51-55, 1962
Madigan M. T., Martinko J. M., Parker J. – Biology of microorganisms, Ed. Prentice Hall International Inc., SUA, 1997
Măzăreanu C. – Microbiologie generală, Ed. Alma Mater, Bacău, 1999
Mihăescu G., Gavrilă L.– Biologia microorganismelor fixatoare de azot, Ed. Ceres, Bucureşti, 1989
Murray P.R., Ellen Jo Baron, Pfaller M.A., Tenover F.C., Yolken R.H. – Manual of Clinical Microbiology, Ed. ASM Press, USA, 1995
Pamfil D. – Microbiologie, Ed. Genesis, Cluj Napoca, 1999 Papacostea P. – Biologia solului, Ed. Ştiinţifică şi Enciclopedică,
Bucureşti, 1976 Popa I. A., Teodorescu C. Şt. – Microbiologia vinului, Ed. Ceres,
Bucureşti, 1990 Priest F., Austin B. – Modern bacterial taxonomy, Ed. Chapman &
Hall, Londra, Marea Britanie, 1993

201
Prusiner S. B. - Prion Biology and Diseases, CSHL Press, Danvers, USA, 2004
Ross F. C. – Introductory microbiology, Editura Charles E. Merrill Publishing Company, USA, 1983
Schlegel H.G. – Allgemeine Mikrobiologie, Ed. Thieme, Germania, 1992
Şesan Tatiana Eugenia, Tănase C. – Mycobiota- Sisteme de clasificare, Ed. Universitaţii „Alexandru Ioan Cuza” Iaşi, 2006
Şesan Tatiana Eugenia, Tănase C. – Ciuperci anamorfe fitopatogene, Ed. Universităţii din Bucureşti, 2007
Tănase C. – Micologie, Ed. Universităţii „Alexandru Ioan Cuza” Iaşi, 2002
Tănase C., Şesan Tatiana Eugenia – Concepte actuale în taxonomia ciupercilor, Ed. Uniersităţii „Alexandru Ioan Cuza”, Iaşi, 2006
Tofan Clemansa – Microbiologie alimentară, Ed. Agir, Bucureşti, 2004
Topală N.D. – Microbiologie generală, vol. I, II. Universitatea Al. I. Cuza, Iaşi, 1978
Tortora G. J., Funke B. R., Case Christine – Microbiology - an introduction, Editura The Benjamin/Cummings Publishing Company, INC., SUA, 1992
Ulea E., Iacob Viorica, Ilişescu Izabela – The lignicole micromycetes of cherry and plum trees from Central Area of Moldavia, USAMV Iaşi, Lucrări Ştiinţifice, Seria Horticultură , 48, 665-670, 2005
Ulea E., Ilisescu Izabela, Mocanu Aliona., Sava D. – The effect of the fertilization with Vinassa Rompak product on the biological activity of the cultivated soils, J. Central European Agr, 7, 297-300, 2006
Ulea E., Lipşa F. D. – Analysis of microbial community on mine tailings from the national park of Călimani mountain (Eastern Carpathians), Research Journal of Agricultural Science, vol. 41 (1), p. 518-522, 2009
Ulea E., Lipşa F.D., Chiriac Irina Paraschiva, Coroi Irina Gabriela – Effects of chlorsulfuron on soil microbial population, Universitatea de Ştiinţe Agricole şi de Medicină Veterinară „Ion Ionescu de la Brad” Iaşi, Lucr. şt. Seria Agronomie, vol. 53(1), p. 157-161, 2010

202
Ulea E., Lipşa F.D., Chiriac Irina Paraschiva, Coroi Irina Gabriela – Acetochlor influence on soil microbial communities, Universitatea de Ştiinţe Agricole şi de Medicină Veterinară „Ion Ionescu de la Brad” Iaşi, Lucr. şt. Seria Agronomie, vol. 53(1), p. 153-157, 2010
Ungureanu Elena, Ulea E., Iacob Viorica – On the biocide properties of some products based on natural aromatic compounds. Cellulose Chemistry and Technology, 42 (7-8), 381-386, 2008
Van Elsas J. D., Jansson Janet, Trevors J. T. – Modern soil microbiology – second edition, CRC Press, SUA, 2007
Whittaker R. H. – New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. Science, 163(863):150-60, 1969
Widmer F., Beffa R. – Aide-mémoire de biochimie et de biologie moléculaire, Editions Médicales Internationales, ISBN: 2- 7430-0175-5, Franţa, 1997
Wood M. – Soil biology. Ed. Blackie, New York, SUA, 1989 Zamfirache Maria-Magdalena, Toma C., 2000 – Simbioza în lumea
vie, Ed. Univ. “Alexandru Ioan Cuza” Iaşi. Zarnea Gh. – Tratat de microbiologie generală, Ed. Academiei
Române, Bucureşti, Vol. I - 1983, Vol. II - 1984, Vol. III - 1986, Vol. IV - 1990,Vol. V - 1994.

Consilier editorial: Vasile VÎNTU Tehnoredactor: Florin Daniel LIPŞA Corector: Gilda DRĂGĂNESCU Bun de tipar: 01.08.2011 Apărut: 2011 Editura: “Ion Ionescu de la Brad” Iaşi
Aleea M. Sadoveanu nr. 3 Tel.: 0232-218300; E-mail: [email protected] ISBN 978-973-147-091-7 PRINTED IN ROMANIA