variabilitatea conŢinutului de amidon Şi fibre la ... · porumb în hrana omului, a animalelor,...
TRANSCRIPT

UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI
MEDICINĂ VETERINARĂ CLUJ-NAPOCA
SCOALA DOCTORALĂ
Ing. TEODORA VERONICA ŞCHIOP
VARIABILITATEA CONŢINUTULUI DE AMIDON ŞI FIBRE
LA POPULAŢII LOCALE ŞI SINTETICE,
LINII CONSANGVINIZATE ŞI HIBRIZI DE PORUMB
REZUMAT AL TEZEI DE DOCTORAT
CONDUCĂTOR ŞTIINŢIFIC
PROF. DR. IOAN HAŞ
CLUJ-NAPOCA
2011

1
CUPRINS
CAPITOLUL I
IMPORTANŢA ECONOMICĂ ŞI RĂSPÂNDIREA GEOGRAFICĂ A
PORUMBULUI………………………………………………………………
9
6
1.1 IMPORTANŢA ECONOMICĂ………………………………………………. 9 6
1.2 RĂSPÂNDIREA GEOGRAFICĂ……………………………………………. 13 7
1.3 PRODUCŢIA DE PORUMB PE PLAN MONDIAL………………………… 14 7
1.4 SITUAŢIA SUPRAFEŢELOR ŞI A PRODUCŢIILOR DE PORUMB ÎN
ROMÂNIA………………………………………………..…………………...
16
8
1.5 UTILIZĂRILE PORUMBULUI……………………………………………… 21
1.6 UTILIZĂRI SPECIFICE ALE AMIDONULUI DE PORUMB....................... 24
CAPITOLUL II
GENETICA ŞI AMELIORAREA CONŢINUTULUI DE AMIDON ............ 28
2.1 GENE CARE CONDIŢIONEAZĂ CONŢINUTUL DE AMIDON LA
PORUMB………………………………………………..…………………….
28
2.2 GENE CARE INFLUENŢEAZĂ DEZVOLTAREA BOABELOR………….. 35
2.3 ANALIZA BIOCHIMICĂ A CONŢINUTULUI DE CARBOHIDRAŢI......... 36
2.4 INTERACŢIUNI ÎNTRE ACUMULAREA COMPONENTELOR
BOBULUI……………………………………………………………………..
42
2.5 STRUCTURA MOLECULARĂ A AMIDONULUI DE PORUMB................ 42
2.5.1 Structura generală a amidonului............................................................. 43
2.5.2 Structura fină a amilozei.......................................................................... 44
2.5.3 Structura fină a amilopectinei.................................................................. 45
2.6 MUTANŢI PENTRU COMPOZIŢIA AMIDONULUI DIN PORUMB.......... 46
2.6.1 Structura amidonului pentru mutanţii cu conţinut ridicat de amiloză 46
2.6.2 Structura amidonului pentru mutanţii cu conţinut ridicat de
amilopectină................................................................................................
48
2.6.3 Structura amidonului pentru alte tipuri de endosperm........................ 48
2.7 ORGANIZAREA GRANULELOR DE AMIDON…………………………… 52
2.8 PROPRIETĂŢI ALE AMIDONULUI DE PORUMB………………………... 55
2.8.1 Proprietăţi generale................................................................................... 55
2.8.2 Proprietăţi ale amidonului din mutanţii bogaţi in amiloză.................... 59
2.8.3 Proprietăţi ale amidonului din mutanţii cu gena wx.............................. 60
2.8.4 Proprietăţi ale amidonului din alte tipuri mutanţi de porumb şi linii
consangvinizte...................................................................................................
61

2
CAPITOLUL III
VARIABILITATEA CONŢINUTULUI DE FIBRE LA PORUMB………… 63
3.1 CONSIDERAŢII GENERALE.......................................................................... 63
3.2 IMPORTANŢA CONŢINUTULUI DE FIBRE LA PORUMB……………… 65
3.3 UTILIZĂRI SPECIFICE ALE FIBRELOR DE PORUMB………………….. 67
CAPITOLUL IV
OBIECTIVELE TEZEI DE DOCTORAT…………………………………… 75 9
4.1 MOTIVAŢIA CERCETĂRII…………………………………………………. 75
4.2 OBIECTIVE....................................................................................................... 77
CAPITOLUL V
CONDIŢIILELE PEDO-CLIMATICE DE DESFĂŞURARE A
EXPERIENŢELOR..............................................................................................
78
CAPITOLUL VI
MATERIALUL BIOLOGIC ŞI METODA DE LUCRU................................. 82 10
6.1 CREAREA LINIILOR CONSANGVINIZATE ISONUCLEARE.................... 82
6.2 TEHNOLOGIA APLICATĂ ÎN CÂMPUL EXPERIMENTAL....................... 86
6.3 PREGĂTIREA MATERIALULUI PENTRU ANALIZE CHIMICE................ 87
6.4 PRELUCRAREA STATISTICĂ A REZULTATELOR EXPERIMENTALE.. 91
CAPITOLUL VII
REZULTATE OBŢINUTE ŞI DISCUŢII........................................................ 95 13
7.1 STUDIUL POPULAŢIILOR LOCALE DIN COLECŢIA SCDA TURDA.... 95 13
7.1.1 Conţinutul de amidon şi fibre la populaţiile locale din colecţia SCDA
Turda..................................................................................................................
95
7.1.2 Studiul relaţiilor dintre constituenţii bobului la populaţiile locale..... 98
7.2 STUDIUL POPULAŢIILOR SINTETICE CREATE LA SCDA TURDA......... 103 14
7.2.1 Conţinutul de amidon şi fibre la populaţiile sintetice create la SCDA
Turda..................................................................................................................
103
7.2.2 Studiul relaţiilor dintre constituenţii bobului la populaţiile sintetice.. 107
7.3 STUDIUL LINIILOR CONSANGVINIZATE CREATE LA SCDA TURDA. 112 15
7.3.1 Conţinutul de amidon şi fibre la liniile consangvinizate create la
SCDA Turda.....................................................................................................
112
7.3.2 Studiul relaţiilor dintre constituenţii bobului la liniile consangvinizate.. 120
7.4 CONŢINUTUL DE AMIDON ŞI FIBRE AL LINIILOR
CONSANGVINIZATE ÎNTR-UN SISTEM GENETIC BALANSAT.............
161
16
7.4.1 Conţinutul în amidon şi fibre la cinci grupe de linii consangvinizate
isonucleare.........................................................................................................
161
7.4.2 Ponderea factorilor genetici localizaţi în citoplasmă şi nucleu în
determinismul conţinutului de amidon şi fibre la un sistem de linii
isonucleare balansat........................................................................................
165

3
7.5 CONŢINUTUL DE AMIDON SI FIBRE AL LINIILOR
CONSANGVINIZATE ÎNTR-UN SISTEM GENETIC NEBALANSAT........
166
18
7.5.1 Conţinutul de amidon al liniilor consangvinizate în sistemul genetic
nebalansat..........................................................................................................
166
7.5.2 Conţinutul de fibre solubile al liniilor consangvinizate într-un sistem
genetic nebalansat.............................................................................................
169
7.6 CONŢINUTUL DE AMIDON SI FIBRE AL LINIILOR
CONSANGVINIZATE FORME PARENTALE ÎN SISTEM DIALEL...........
172
20
7.6.1 Conţinutul de amidon al liniilor consangvinizate forme parentale în
sistem dialel.......................................................................................................
172
7.6.2 Conţinutul de fibre al liniilor consangvinizate forme parentale în
sistem dialel.......................................................................................................
173
7.7 TRANSMITEREA EREDITARĂ A CONŢINUTULUI DE AMIDON ŞI
FIBRE SOLUBILE LA PORUMB ÎNTR-UN SISTEM GENETIC DE LINII
CONSANGVINIZATE ISONUCLEARE.........................................................
174
22
7.7.1 Varianţa conţinutului de amidon în cinci seturi de linii consangvinizate
isonucleare.........................................................................................................
174
7.7.2 Influenţa acţiunilor genice de natură citoplasmatică, nucleară şi
interacţiunilor citoplasmă × nucleu în ereditatea conţinutului de amidon
178
7.7.3 Varianţa conţinutului de fibre solubile în cinci seturi de linii
consangvinizate isonucleare......................................................................
183
7.7.4 Influenţa acţiunilor genice de natură citoplasmatică, nucleară şi
interacţiunilor citoplasmă×nucleu în ereditatea conţinutului de fibre......
187
7.8 EREDITATEA UNOR CARACTERE DE INTERES AGRONOMIC LA
PORUMBUL TIMPURIU STUDIATĂ ÎNTR-UN SISTEM DIALEL DE
TIPUL p(p-1)......................................................................................................
192
7.8.1 Studiul eredităţii greutăţii ştiuletelui....................................................... 192
7.8.2 Studiul eredităţii lungimii ştiuletelui....................................................... 195
7.8.3 Studiul eredităţii numărului de rânduri de boabe pe ştiulete................. 200
7.8.4 Studiul eredităţii capacităţii de producţie............................................... 204
7.8.5 Studiul eredităţii conţinutului de substanţă uscată în boabe la
recoltare......................................................................................................
209
7.9 EREDITATEA CONŢINUTULUI DE AMIDON ŞI FIBRE SOLUBILE LA
UN SISTEM DIALEL DE TIPUL p(p-1) REALIZAT ÎNTRE LINII
CONSANGVINIZATE TIMPURII...................................................................
213
25
7.9.1 Studiul eredităţii conţinutului de amidon............................................... 213 25
7.9.2 Studiul eredităţii conţinutului de fibre.................................................... 221 28
CAPITOLUL VIII
CONCLUZII ŞI RECOMANDĂRI.................................................................. 229 32
BIBLIOGRAFIE............................................................................................. 234 35

4
CUVÂNT ÎNAINTE
Porumbul este una din cele mai importante plante de cultură, datorită
suprafeţelor mari pe care le ocupă, datorită productivităţii ridicate şi a multiplelor sale
intrebuinţări în alimentaţia oamenilor, în zootehnie şi în industrie.
Ameliorarea modernă a porumbului a impus obţinerea unor genotipuri
productive şi de calitate, care să corespundă cerinţelor privind utilizarea boabelor de
porumb în hrana omului, a animalelor, dar şi ca materie primă în industria alimentară,
industria hârtiei, a ambalajelor, industria farmaceutică şi cosmetică, în obţinerea
adezivilor, lacurilor, vopselelor, iar mai recent a biocarburanţilor.
Conţinutul mare în amidon al bobului de porumb a făcut ca în jurul acestui
constituent să se dezvolte o adevărată industruie. Având în vedere numeroasele
utilizări ale amidonului, studiul variabilităţii acestui component dominant al bobului
de porumb capătă o importanţă deosebită. Cunoaşerea conţinutului de amidon din
diferite genotipuri (populaţii locale, populaţii sintetice, soiuri, linii consangvinizate,
hibrizi) este necesară în stabilirea utilizării acestora, luând în considerare si faptul că
acest component este corelat negativ cu conţinutul de proteină şi grăsimi.
Cunoaşterea conţinutului de fibre din bobul de porumb este importantă din
perspectiva utilizării acestora în alimentaţia umană şi ca furaj pentru animale. Fibrele
din porumb sunt un produs secundar al prelucrării industriale umede. Industria
procesatoare a porumbului produce anual circa 4 milioane de tone de fibră de porumb,
cantităţi care sunt valorificate ca nutreţuri, un ingredient ieftin în hrana animalelor.
Fibrele din porumb prezintă potenţial ca materie primă la fabricarea de diferite
produse, fiind disponibilă pe scară largă în ţări mari producătoare de porumb. O
importantă utilizare a fibrelor de porumb se referă la obţinerea uleiului de fibra de
porumb, cu rol important in reducerea colesterolului (SINGH şi colab, 2001). În timp ce
conţinutul de ulei din fibră este de 2%, conţinutul de gumă din aceasta este de 40%
(DONER şi colab. 1998). Poate fi folosită în produsele alimentare ca emulsificator,
agent de îngroşare sau sursă de fibre solubile. În industrie poate fi folosită la obţinerea
adezivilor sau ca agent de îngroşare pentru vopselele pe bază de apă.

5
Prezenta lucrare cuprinde rezultatele cercetărilor efectuate în perioada 2009-
2010, în câmpul experimental şi laboratorul de ameliorare a porumbului de la SCDA
Turda.
Teza de doctorat este structurată pe 8 capitole, şi cuprinde 244 pagini, 100
tabele, 81 figuri, 157 titluri bibliografice.
Teza nu ar fi putut fi realizată fără sprijinul şi îndrumarea distinsului Prof. Dr.
IOAN HAŞ. Dânsului îi adresez cele mai calde mulţumiri pentru suportul ştiinţific,
îndrumarea, atenţia, răbdarea şi sprijinul permanent acordat în aceşti ani.
Mulţumirile mele se îndreapta şi către doamna Ing. Dr. Voichita Haş, şeful
Laboratorului de ameliorare a Porumbului din cadrul SCDA Turda, pentru sprijinul
acordat în realizarea experientelor şi interpretarea rezultatelor, precum şi către întregul
colectiv al Laboratorului de ameliorare a Porumbului din cadrul SCDA Turda, care
mi-au oferit sprijinul lor în vederea realizării experienţelor.
Multumesc colegilor de la Disciplina de Genetica şi Ameliorarea Plantelor
pentru sprijinul acordat pe tot parcursul elaborării tezei de doctorat.
Alese mulţumiri şi recunoştinţă se îndreaptă către soţul meu, către familia mea, cei
care m-au sprijinit moral pe tot parcursul acestor ani de pregătire şi elaborare a tezei.
Vă multumesc !

6
CAPITOLUL I
IMPORTANŢA ECONOMICĂ ŞI RĂSPÂNDIREA GEOGRAFICĂ
A PORUMBULUI
1.1. IMPORTANŢA ECONOMICĂ
Una dintre cele mai importante plante de cultură, porumbul ocupă pe plan
mondial locul trei ca suprafaţă şi primul loc ca producţie, fiind devansat ca suprafaţă
de grâu şi orez. Prin suprafeţele mari pe care le ocupă şi producţiile ce le realizează,
devine un component de baza în asigurarea hranei. Datorită compoziţiei chimice a
tuturor părţilor componente, porumbul pe lângă importanţa alimentară pe care o
prezintă pentru om constituie şi un furaj de bază în hrana animalelor şi o valoroasă
materie primă pentru industrie.
Din producţia mondială de porumb, asa cum statisticile FAO o arată, cea mai
mare parte se foloseşte pentru furajarea animalelor (72%), urmată de consumul în
alimentaţia umană (21%) şi doar 7% fiind destinat industrializării. Ca aliment,
porumbul are şi unele neajunsuri, din care remarcăm cantitatea redusă a unor
aminoacizi esenţiali, cum sunt lizina şi triptofanul, lipsa vitaminelor C şi D şi altele.
Cu toate acestea, porumbul rămâne sursa de bază în furajarea animalelor, sub forma
de nutreţ concentrat, siloz sau masă verde.
Forma sub care porumbul este cel mai des folosit în alimentaţie este forma
procesată prin măcinare uscată sau umedă. Prin măcinare umedă se pot separa
amidonul, proteinele, fibrele şi germenii. Prin măcinare uscată se se separă fracţiunile
de endosperm de diferite dimensiuni (făină, grişuri) de germenii bogaţi în grăsimi, şi
de fibre. Amidonul şi produsele de măciniş stau la baza obţinerii snacks-urilor,
cerealelor pentru micul dejun, siropurilor şi alcoolului. Cel mai mare consumator al
porumbului măcinat este industria bauturilor alcoolice. Produsele de măciniş
constituie o sursă ieftină de carbohidraţi fermentescibili. Prin hidroliză enzimatică
amidonul este transformat în zaharuri, rezultând plămada dulce, produs ce stă la baza
obţinerii băuturilor alcoolice precum berea sau băuturi distilate (votka, whiskey). O

7
recentă utilizare a amidonului este legată de extracţia de bioetanol. Obţinerea
etanolului din materii prime amidonoase (aşa cum este şi porumbul) presupune
transformarea amidonului în zahăr fermentescibil proces ce se realizează sub acţiunea
enzimelor amilolitice. Peste 30% din porumbul cultivat în SUA în 2006 a fost folosit
pentru producţia de bioetanol. Din acest punct de vedere, porumbul tinde să devină
cea mai importantă plantă energetică pentru zona temperată.
Fibrele din porumb sunt un produs secundar al prelucrării industriale umede,
Industrie ce produce anual circa 4 milioane de tone de fibră de porumb. Aceste
cantităţi sunt valorificate ca nutreţuri, un ingredient ieftin în hrana animalelor.
Cercetările recente efectuate asupra fibrei de porumb au arătat că acestea pot fi
folosite pentru extracţia de ulei de fibră de porumb, care are un rol important în
reducerea colesterolului din sange. O importantă utilizare a fibrelor de porumb se
referă şi la obţinerea agenţilor de gelificare, a gumelor. În industrie poate fi folosită la
obţinerea adezivilor sau ca agent de îngroşare pentru vopselele pe bază de apă.
1.2. RĂSPÂNDIREA GEOGRAFICĂ
Din centrul de origine (Mexic, America Centrală, America de Sud), porumbul
s-a răspândit şi este cultivat pe glob în foarte multe condiţii de climă şi sol. Arealul
geografic se întinde între paralelele 42º latitudine sudică (Noua Zeelandă) şi 53º
latitudine nordică (Rusia). Pentru boabe, zona de cultură a porumbului este delimitată
de izoterma de 18º a lunii iulie. Pentru furaj, în emisfera nordică se poate cultiva până
la paralela de 59º în Rusia.
1.3. PRODUCŢIA DE PORUMB PE PLAN MONDIAL
Suprafaţa totală cultivată pe glob a sporit în ultimele decenii, de la 86 milioane
hectare în perioada 1948- 1950 la peste 144,4 milioane hectare în anul 2006 (FAO, 2006).
În anul 2006, în privinţa suprafeţelor cultivate, porumbul ocupă locul al treilea după grâu

8
(216,1 milioane hectare) şi orez (154,3 milioane hectare), iar cele mai mari suprafeţe
cultivate sunt în SUA, China, Brazilia, India, Mexic etc. (VÂRBAN, 2008).
Tabelul 1
Suprafaţa şi producţia de porumb pe continente, 2006 (după Vârban, 2008)
Zona geografică Suprafaţa
(mii ha)
Producţia medie
(kg/ha)
Producţia totală
(mii tone)
AFRICA 26118 1771 46260
AMERICA DE NORD ŞI CENTRALĂ 38772 6027 301654
AMERICA DE SUD 18586 3316 66990
ASIA 47365 4286 203025
EUROPA 13439 5710 76742
OCEANIA 96 5835 558
TOTAL 144376
1.4 SITUAŢIA SUPRAFEŢELOR ŞI A PRODUCŢIILOR DE PORUMB ÎN
ROMÂNIA
În România, porumbul are condiţii favorabile de cultură pe ce mai mare parte din
suprafaţa arabilă. Din punct de vedere ecologic nu există zone în care care cerinţele
porumbului să fie satisfăcute în optimum în fiecare an, având în vedere că în diferite
regiuni ale ţării pot să apară unii factori mai puţin prielnici culturii (temperaturi scăzute în
primăvară, exces de umiditate, secete prelungite în cursul verii etc.).
Zona foarte favorabilă cuprinde câmpia din vestul ţării (de la Carei la
Moraviţa), pe soluri fertile (mai puţin nisipuri şi sărături) şi regim termic favorabil, în
Câmpia Română şi Lunca Dunării. În această zonă predomină solurile fertile, numărul
zilelor cu temperaturi medii zilnice mai mari de 10º C este de 190-200, iar cantitatea
anuală de precipitaţii se ridică la 550-600. Din punct de vedere termic există condiţii
foarte favorabile pentru cultivarea cu precădere a hibrizilor tardivi, iar potenţialul
natural al zonei permite obţinerea unor producţii de peste 6000 kg/ha.

9
CAPITOLUL II
OBIECTIVELE TEZEI DE DOCTORAT
Prin acest proiect de cercetare se urmăreşte realizarea urmatoarelor obiective:
studiul colecţiei de populaţii locale, soiuri, populaţii sintetice privind
conţinutul de amidon şi fibre şi identificarea unor corelaţii cu caractere ale
plantei, ştiuletelui şi boabelor;
identificarea din colectia de linii consangvinizate a genotipurilor
bogate/sărace în amidon si fibre;
evidenţierea rolului citoplasmei în transmiterea ereditară a conţinutului de
amidon şi fibre;
studiul transmiterii ereditare a conţinutului de amidon şi fibre;
identificarea unor genotipuri (populaţii locale, linii consangvinizate) pentru
iniţierea ciclurilor primare pentru selecţie direcţională pentru caracterele
studiate.

10
CAPITOLUL III
MATERIALUL BIOLOGIC ŞI METODA DE LUCRU
Analiza conţinutului de amidon şi fibre s-a realizat la colectia de genotipuri de
porumb al Laboratorului de Ameliorare a Porumbului de la S.C.D.A. Turda. Au fost
cuprinse în studiu următoarele tipuri de germoplasmă:
258 populaţii locale;
60 populaţii sintetice, composite şi soiuri locale;
422 linii consangvinizate create la S.C.D.A. Turda;
un sistem dialel cu 30 de hibrizi simpli şi cele şase forme parentale de tipul
p(p-1);
pentru determinarea conţinutului de amidon şi fibre au fost luate în studiu şi
cinci sisteme ciclice cu linii isonucleare (pe citoplasme diferite).
Liniile consangvinizate elită folosite pentru transferul nucleului pe diferite
provenienţe de citoplasmă, au fost in număr de cinci si sunt prezentate în tabelul 2, iar
cele utilizate ca surse de citoplasmă în tabelul 3.
Tabelul 2
Linii consangvinizate donoare a nucleului
Nr. crt. Linia
consangvinizată Genealogia (germoplasma) Instituţia amelioratoare
1. TC 209 BSSS SCDA Turda
2. TC 243 A 654 X TC 209 SCDA Turda
3. TC 221 Lo3 Berg. X Josenii Bârgăului SCDA Turda
4. TB 367 PI 187 X T 248 I SCDA Turda
5. D 105 Indurat timpuriu european Univ. Hohenheim, Germania
Fiecare linie consangvinizată donoare a nucleului a fost studiată pe şase
provenienţe de citoplasmă, linia donoare de nucleu fiind utilizată ca martor. Pentru
isoliniile nou create denumirea a fost făcută după linia donoare de nucleu, iar
provenienţa citoplasmei a fost trecută alături în paranteză: TC 209 (cit. A 665), TC
243 (cit. T 248), TC 221 (cit. K 1080), TB 367 (cit. K 2051), D 105 (cit. TB 329).
11
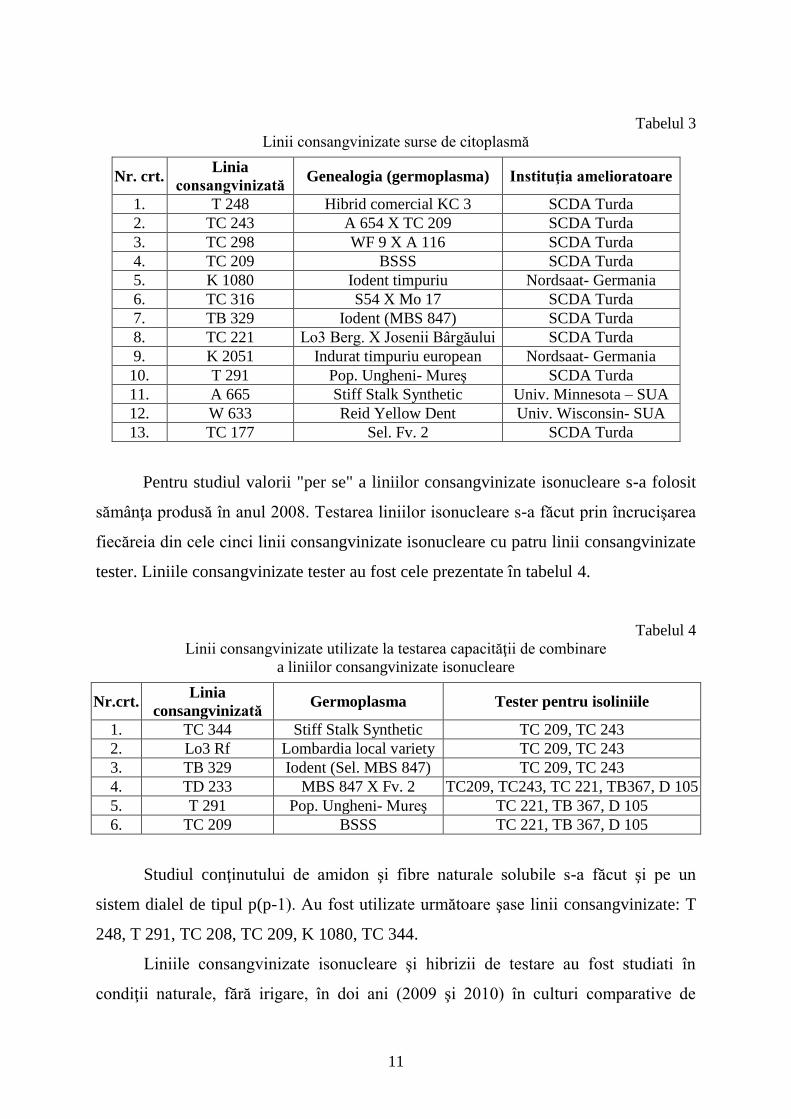
Tabelul 3
Linii consangvinizate surse de citoplasmă
Nr. crt. Linia
consangvinizată Genealogia (germoplasma) Instituţia amelioratoare
1. T 248 Hibrid comercial KC 3 SCDA Turda
2. TC 243 A 654 X TC 209 SCDA Turda
3. TC 298 WF 9 X A 116 SCDA Turda
4. TC 209 BSSS SCDA Turda
5. K 1080 Iodent timpuriu Nordsaat- Germania
6. TC 316 S54 X Mo 17 SCDA Turda
7. TB 329 Iodent (MBS 847) SCDA Turda
8. TC 221 Lo3 Berg. X Josenii Bârgăului SCDA Turda
9. K 2051 Indurat timpuriu european Nordsaat- Germania
10. T 291 Pop. Ungheni- Mureş SCDA Turda
11. A 665 Stiff Stalk Synthetic Univ. Minnesota – SUA
12. W 633 Reid Yellow Dent Univ. Wisconsin- SUA
13. TC 177 Sel. Fv. 2 SCDA Turda
Pentru studiul valorii "per se" a liniilor consangvinizate isonucleare s-a folosit
sămânţa produsă în anul 2008. Testarea liniilor isonucleare s-a făcut prin încrucişarea
fiecăreia din cele cinci linii consangvinizate isonucleare cu patru linii consangvinizate
tester. Liniile consangvinizate tester au fost cele prezentate în tabelul 4.
Tabelul 4
Linii consangvinizate utilizate la testarea capacităţii de combinare
a liniilor consangvinizate isonucleare
Nr.crt. Linia
consangvinizată Germoplasma Tester pentru isoliniile
1. TC 344 Stiff Stalk Synthetic TC 209, TC 243
2. Lo3 Rf Lombardia local variety TC 209, TC 243
3. TB 329 Iodent (Sel. MBS 847) TC 209, TC 243
4. TD 233 MBS 847 X Fv. 2 TC209, TC243, TC 221, TB367, D 105
5. T 291 Pop. Ungheni- Mureş TC 221, TB 367, D 105
6. TC 209 BSSS TC 221, TB 367, D 105
Studiul conţinutului de amidon şi fibre naturale solubile s-a făcut şi pe un
sistem dialel de tipul p(p-1). Au fost utilizate următoare şase linii consangvinizate: T
248, T 291, TC 208, TC 209, K 1080, TC 344.
Liniile consangvinizate isonucleare şi hibrizii de testare au fost studiati în
condiţii naturale, fără irigare, în doi ani (2009 şi 2010) în culturi comparative de

12
orientare la SCDA Turda. Testările de linii isonucleare TC 209 şi TC 243 au fost
studiate în 28 parcele, în 3 repetiţii, în timp ce grupul de linii isonucleare TC 221, TB
367 şi D105 în 21 parcele, în 3 repetiţii. Fiecare parcelă a fost formată de câte 2
rânduri de 5 m lungime, cu 70 cm distanţa între rânduri. Distanta între plante a fost de
23,7 cm, rezultând 23 plante pe rând. Densitatea de plante la hectar a fost de 60.000.
După recoltare, ştiuleţii au fost uscaţi la temperatura de 38-40°C; după
desprinderea de pe rahis s-a constituit proba medie. Din proba medie s-a cântărit o
probă de 50g; această probă a fost măcinată cu moara de laborator, cu cel mult 48 de
ore înainte de efectuarea analizelor. Făina rezultată a fost folosită la determinarea
conţinutului boabelor în proteine, grăsimi, amidon, fibre şi cenuşă. Determinările au
fost făcute cu ajutorul analizorului NIR INSTALAB 600. S-au folosit probe provenite
din autopolenizare, şi probe provenite de la ştiuleţi polenizaţi liber.
Studiul variabilităţii conţinutului de amidon şi fibre s-a analizat cu ajutorul
analizei sirului de variaţie. S-au studiat corelaţiile dintre componentele chimice ale
bobului, precum şi legătura între unele caractere ale bobului şi stiuletelui şi cele două
componente. În vederea evaluǎrii variabilitǎţii colecţiei în privinţa însuşirilor de calitate,
s-a recurs la studiul parametrilor variabilitǎţii pentru caracterele analizate.
Caracterizarea variabilitǎţii s-a fǎcut prin calculul mediei ( x ), abaterii standard a
mediei (s), amplitudinii de variaţie şi a coeficientului de variaţie (s%), după modelul
prezentat de ARDELEAN (2006).

13
CAPITOLUL IV
REZULTATE OBŢINUTE ŞI DISCUŢII
4.1. STUDIUL POPULAŢIILOR LOCALE DIN COLECŢIA SCDA TURDA
Luând în considerare conţinutul de amidon al tuturor celor 258 de genotipuri de
porumb luate în studiu, se observă că amplitudinea de variaţie a acestuia a avut valori
cuprinse între 59,3% şi 68,9%, cu o medie de 64,9%; abaterea standard de 1,9% arată
că majoritatea genotipurilor se grupează în jurul mediei, iar valoarea mică a
coeficientului de variabilitate (2,93) evidenţiază o variabilitate redusă a conţinutului
de amidon în rândul populaţiilor luate în studiu (tabelul 5).
În ceea ce priveste conţinutul de fibre, se observă că amplitudinea de variaţie a
acestuia a avut valori cuprinse între 3,3% şi 7,3%, cu o medie de 5,27%; valoarea
coeficientului de variabilitate de 15,05 arată o variabilitate mijlocie a conţinutului de
fibre în rândul populaţiilor analizate; abaterea standard a fost de 0,79%, fapt care arată
că majoritatea genotipurilor se grupează în jurul mediei, lucru întărit şi de valoarea
mică a varianţei, de 0,63 (tabel 5).
Tabelul 5
Analiza statistică a conţinutului de amidon şi fibre la un set populaţii locale de porumb
Parametrul statistic
Amidon Fibre
Media % 64,90 5,27
Eroarea standard a diferenţei 0,12 0,05
Abaterea standard 1,90 0,79
Varianţa 3,61 0,63
Amplitudinea diferenţelor 9,60 4,00
Valoarea minimă 59,30 3,30
Valoarea maximă 68,90 7,30
Nivel semnificaţie abateri P=95% 0,23 0,10
Coeficientul de variabilitate 2,93 15,05
Numărul de cazuri 258 258
14
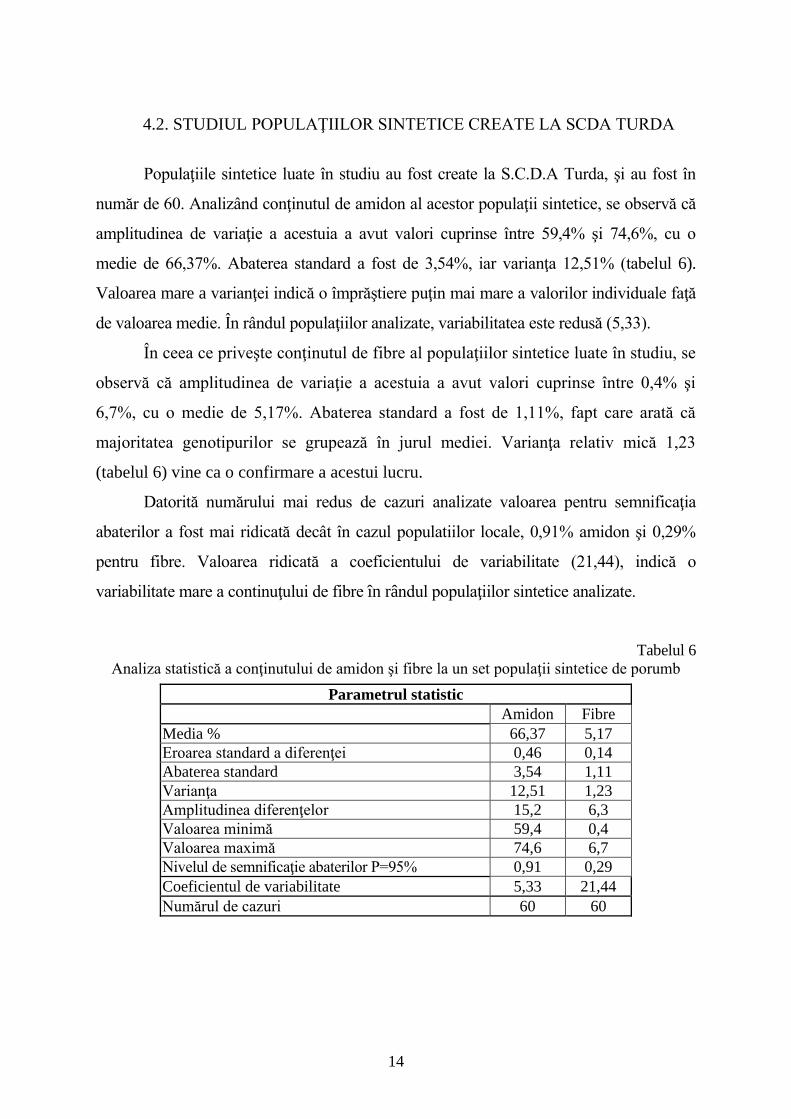
4.2. STUDIUL POPULAŢIILOR SINTETICE CREATE LA SCDA TURDA
Populaţiile sintetice luate în studiu au fost create la S.C.D.A Turda, şi au fost în
număr de 60. Analizând conţinutul de amidon al acestor populaţii sintetice, se observă că
amplitudinea de variaţie a acestuia a avut valori cuprinse între 59,4% şi 74,6%, cu o
medie de 66,37%. Abaterea standard a fost de 3,54%, iar varianţa 12,51% (tabelul 6).
Valoarea mare a varianţei indică o împrăştiere puţin mai mare a valorilor individuale faţă
de valoarea medie. În rândul populaţiilor analizate, variabilitatea este redusă (5,33).
În ceea ce priveşte conţinutul de fibre al populaţiilor sintetice luate în studiu, se
observă că amplitudinea de variaţie a acestuia a avut valori cuprinse între 0,4% şi
6,7%, cu o medie de 5,17%. Abaterea standard a fost de 1,11%, fapt care arată că
majoritatea genotipurilor se grupează în jurul mediei. Varianţa relativ mică 1,23
(tabelul 6) vine ca o confirmare a acestui lucru.
Datorită numărului mai redus de cazuri analizate valoarea pentru semnificaţia
abaterilor a fost mai ridicată decât în cazul populatiilor locale, 0,91% amidon şi 0,29%
pentru fibre. Valoarea ridicată a coeficientului de variabilitate (21,44), indică o
variabilitate mare a continuţului de fibre în rândul populaţiilor sintetice analizate.
Tabelul 6
Analiza statistică a conţinutului de amidon şi fibre la un set populaţii sintetice de porumb
Parametrul statistic
Amidon Fibre
Media % 66,37 5,17
Eroarea standard a diferenţei 0,46 0,14
Abaterea standard 3,54 1,11
Varianţa 12,51 1,23
Amplitudinea diferenţelor 15,2 6,3
Valoarea minimă 59,4 0,4
Valoarea maximă 74,6 6,7
Nivelul de semnificaţie abaterilor P=95% 0,91 0,29
Coeficientul de variabilitate 5,33 21,44
Numărul de cazuri 60 60
15
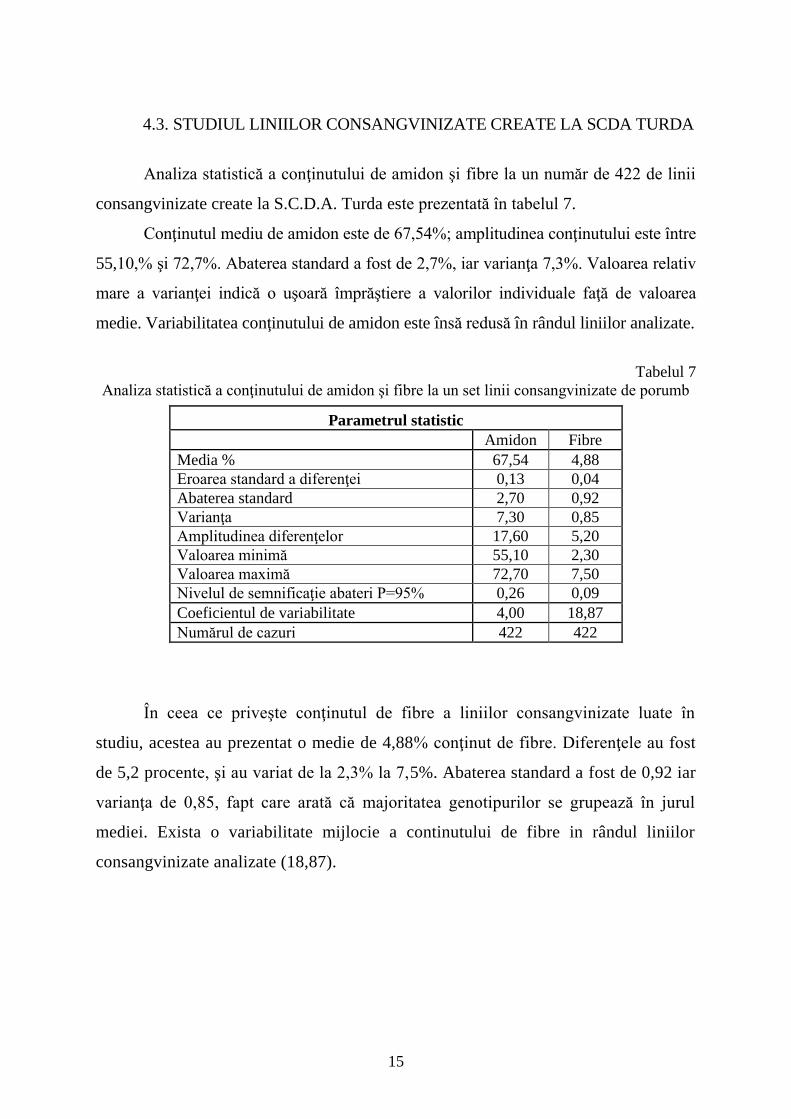
4.3. STUDIUL LINIILOR CONSANGVINIZATE CREATE LA SCDA TURDA
Analiza statistică a conţinutului de amidon şi fibre la un număr de 422 de linii
consangvinizate create la S.C.D.A. Turda este prezentată în tabelul 7.
Conţinutul mediu de amidon este de 67,54%; amplitudinea conţinutului este între
55,10,% şi 72,7%. Abaterea standard a fost de 2,7%, iar varianţa 7,3%. Valoarea relativ
mare a varianţei indică o uşoară împrăştiere a valorilor individuale faţă de valoarea
medie. Variabilitatea conţinutului de amidon este însă redusă în rândul liniilor analizate.
Tabelul 7
Analiza statistică a conţinutului de amidon şi fibre la un set linii consangvinizate de porumb
Parametrul statistic
Amidon Fibre
Media % 67,54 4,88
Eroarea standard a diferenţei 0,13 0,04
Abaterea standard 2,70 0,92
Varianţa 7,30 0,85
Amplitudinea diferenţelor 17,60 5,20
Valoarea minimă 55,10 2,30
Valoarea maximă 72,70 7,50
Nivelul de semnificaţie abateri P=95% 0,26 0,09
Coeficientul de variabilitate 4,00 18,87
Numărul de cazuri 422 422
În ceea ce priveşte conţinutul de fibre a liniilor consangvinizate luate în
studiu, acestea au prezentat o medie de 4,88% conţinut de fibre. Diferenţele au fost
de 5,2 procente, şi au variat de la 2,3% la 7,5%. Abaterea standard a fost de 0,92 iar
varianţa de 0,85, fapt care arată că majoritatea genotipurilor se grupează în jurul
mediei. Exista o variabilitate mijlocie a continutului de fibre in rândul liniilor
consangvinizate analizate (18,87).
16
4.4 CONŢINUTUL DE AMIDON ŞI FIBRE AL LINIILOR
CONSANGVINIZATE ÎNTR-UN SISTEM GENETIC BALANSAT
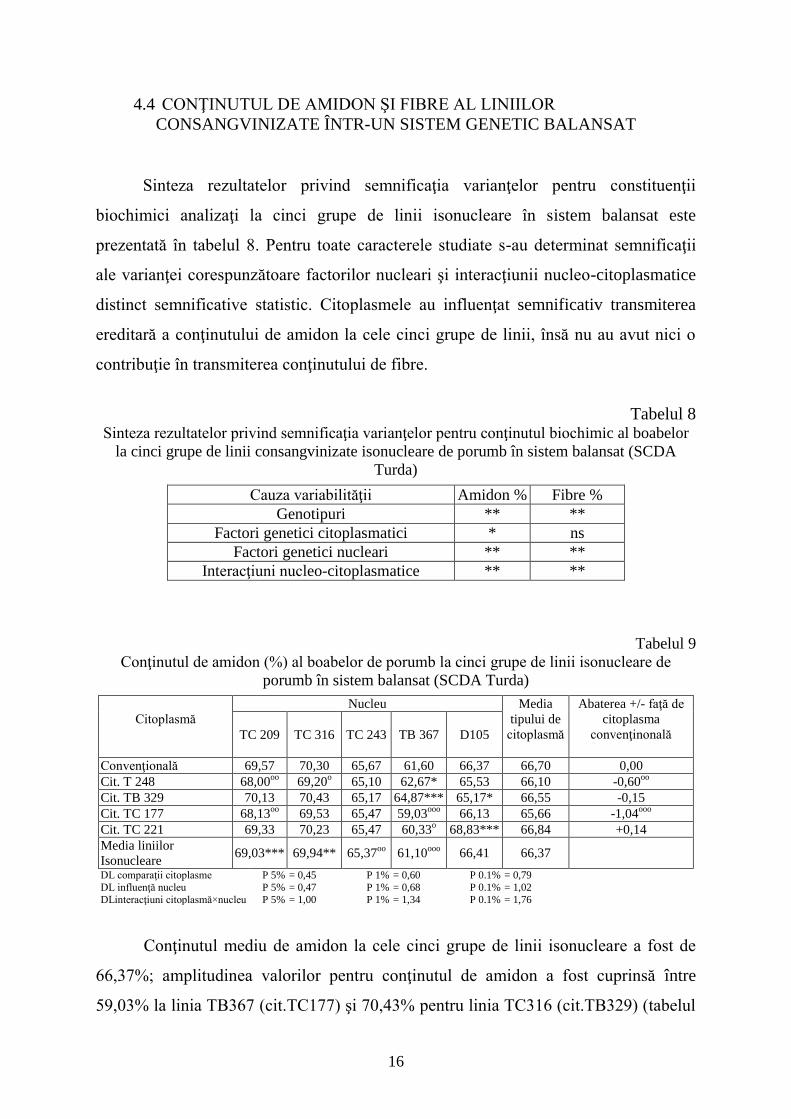
Sinteza rezultatelor privind semnificaţia varianţelor pentru constituenţii
biochimici analizaţi la cinci grupe de linii isonucleare în sistem balansat este
prezentată în tabelul 8. Pentru toate caracterele studiate s-au determinat semnificaţii
ale varianţei corespunzătoare factorilor nucleari şi interacţiunii nucleo-citoplasmatice
distinct semnificative statistic. Citoplasmele au influenţat semnificativ transmiterea
ereditară a conţinutului de amidon la cele cinci grupe de linii, însă nu au avut nici o
contribuţie în transmiterea conţinutului de fibre.
Tabelul 8 Sinteza rezultatelor privind semnificaţia varianţelor pentru conţinutul biochimic al boabelor
la cinci grupe de linii consangvinizate isonucleare de porumb în sistem balansat (SCDA
Turda)
Cauza variabilităţii Amidon % Fibre %
Genotipuri ** **
Factori genetici citoplasmatici * ns
Factori genetici nucleari ** **
Interacţiuni nucleo-citoplasmatice ** **
Tabelul 9
Conţinutul de amidon (%) al boabelor de porumb la cinci grupe de linii isonucleare de
porumb în sistem balansat (SCDA Turda)
Citoplasmă
Nucleu Media
tipului de
citoplasmă
Abaterea +/- faţă de
citoplasma
convenţinonală
TC 209 TC 316 TC 243 TB 367 D105
Convenţională 69,57 70,30 65,67 61,60 66,37 66,70 0,00
Cit. T 248 68,00oo
69,20o
65,10 62,67* 65,53 66,10 -0,60oo
Cit. TB 329 70,13 70,43 65,17 64,87*** 65,17* 66,55 -0,15
Cit. TC 177 68,13oo
69,53 65,47 59,03ooo
66,13 65,66 -1,04ooo
Cit. TC 221 69,33 70,23 65,47 60,33o
68,83*** 66,84 +0,14
Media liniilor
Isonucleare 69,03*** 69,94** 65,37
oo 61,10
ooo 66,41 66,37
DL comparaţii citoplasme P 5% = 0,45 P 1% = 0,60 P 0.1% = 0,79
DL influenţă nucleu P 5% = 0,47 P 1% = 0,68 P 0.1% = 1,02
DLinteracţiuni citoplasmă×nucleu P 5% = 1,00 P 1% = 1,34 P 0.1% = 1,76
Conţinutul mediu de amidon la cele cinci grupe de linii isonucleare a fost de
66,37%; amplitudinea valorilor pentru conţinutul de amidon a fost cuprinsă între
59,03% la linia TB367 (cit.TC177) şi 70,43% pentru linia TC316 (cit.TB329) (tabelul
17
9). Doar media citoplasmei TC 221 a înregistrat o abatere în sens pozitiv faţă de media
citoplasmei convenţionale, deşi în patru din cele cinci grupe analizate a determinat
reducerea conţinutului de amidon.
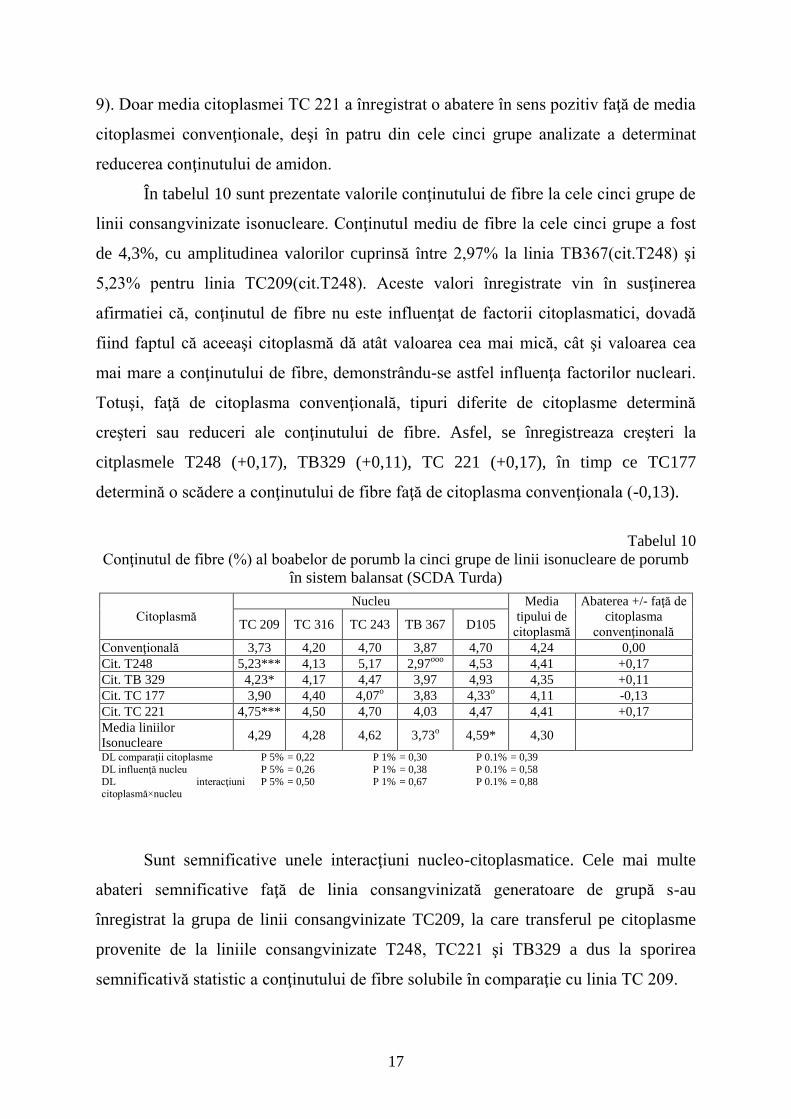
În tabelul 10 sunt prezentate valorile conţinutului de fibre la cele cinci grupe de
linii consangvinizate isonucleare. Conţinutul mediu de fibre la cele cinci grupe a fost
de 4,3%, cu amplitudinea valorilor cuprinsă între 2,97% la linia TB367(cit.T248) şi
5,23% pentru linia TC209(cit.T248). Aceste valori înregistrate vin în susţinerea
afirmatiei că, conţinutul de fibre nu este influenţat de factorii citoplasmatici, dovadă
fiind faptul că aceeaşi citoplasmă dă atât valoarea cea mai mică, cât şi valoarea cea
mai mare a conţinutului de fibre, demonstrându-se astfel influenţa factorilor nucleari.
Totuşi, faţă de citoplasma convenţională, tipuri diferite de citoplasme determină
creşteri sau reduceri ale conţinutului de fibre. Asfel, se înregistreaza creşteri la
citplasmele T248 (+0,17), TB329 (+0,11), TC 221 (+0,17), în timp ce TC177
determină o scădere a conţinutului de fibre faţă de citoplasma convenţionala (-0,13).
Tabelul 10
Conţinutul de fibre (%) al boabelor de porumb la cinci grupe de linii isonucleare de porumb
în sistem balansat (SCDA Turda)
Citoplasmă
Nucleu Media
tipului de
citoplasmă
Abaterea +/- faţă de
citoplasma
convenţinonală TC 209 TC 316 TC 243 TB 367 D105
Convenţională 3,73 4,20 4,70 3,87 4,70 4,24 0,00
Cit. T248 5,23*** 4,13 5,17 2,97ooo
4,53 4,41 +0,17
Cit. TB 329 4,23* 4,17 4,47 3,97 4,93 4,35 +0,11
Cit. TC 177 3,90 4,40 4,07o 3,83 4,33
o 4,11 -0,13
Cit. TC 221 4,75*** 4,50 4,70 4,03 4,47 4,41 +0,17
Media liniilor
Isonucleare 4,29 4,28 4,62 3,73
o 4,59* 4,30
DL comparaţii citoplasme P 5% = 0,22 P 1% = 0,30 P 0.1% = 0,39
DL influenţă nucleu P 5% = 0,26 P 1% = 0,38 P 0.1% = 0,58
DL interacţiuni citoplasmă×nucleu
P 5% = 0,50 P 1% = 0,67 P 0.1% = 0,88
Sunt semnificative unele interacţiuni nucleo-citoplasmatice. Cele mai multe
abateri semnificative faţă de linia consangvinizată generatoare de grupă s-au
înregistrat la grupa de linii consangvinizate TC209, la care transferul pe citoplasme
provenite de la liniile consangvinizate T248, TC221 şi TB329 a dus la sporirea
semnificativă statistic a conţinutului de fibre solubile în comparaţie cu linia TC 209.
18
4.5 CONŢINUTUL DE AMIDON SI FIBRE AL LINIILOR
CONSANGVINIZATE ÎNTR-UN SISTEM GENETIC NEBALANSAT
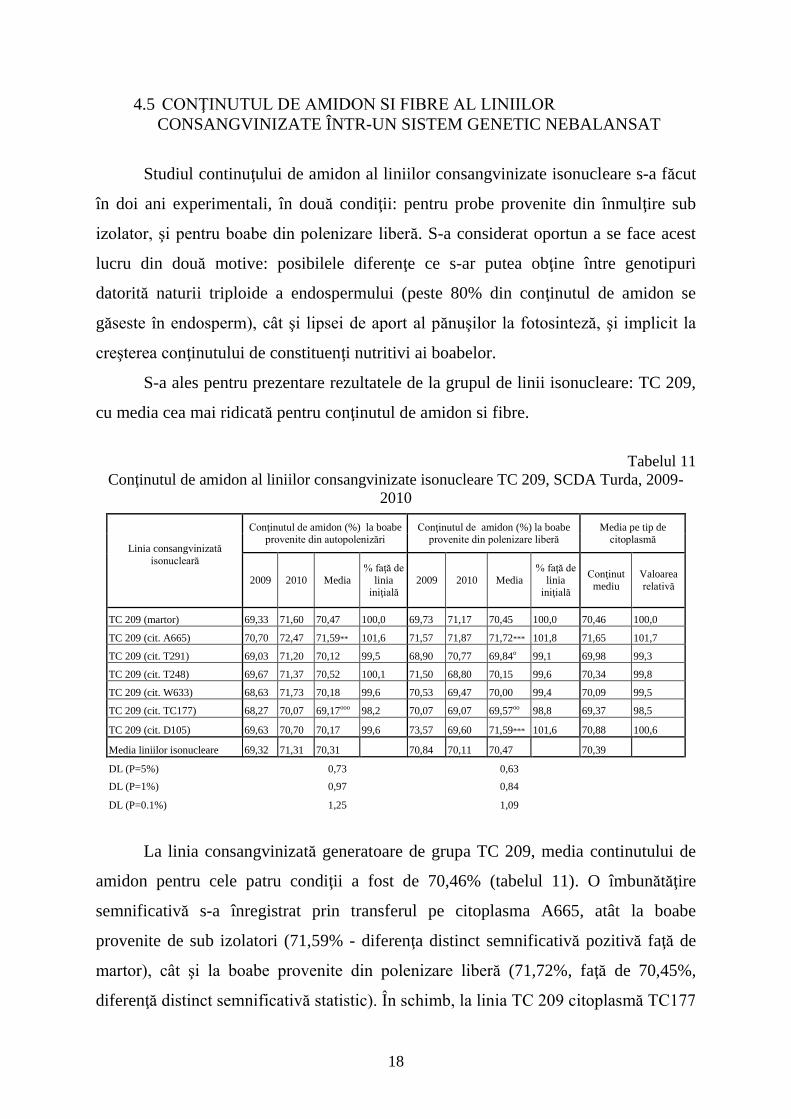
Studiul continuţului de amidon al liniilor consangvinizate isonucleare s-a făcut
în doi ani experimentali, în două condiţii: pentru probe provenite din înmulţire sub
izolator, şi pentru boabe din polenizare liberă. S-a considerat oportun a se face acest
lucru din două motive: posibilele diferenţe ce s-ar putea obţine între genotipuri
datorită naturii triploide a endospermului (peste 80% din conţinutul de amidon se
găseste în endosperm), cât şi lipsei de aport al pănuşilor la fotosinteză, şi implicit la
creşterea conţinutului de constituenţi nutritivi ai boabelor.
S-a ales pentru prezentare rezultatele de la grupul de linii isonucleare: TC 209,
cu media cea mai ridicată pentru conţinutul de amidon si fibre.
Tabelul 11
Conţinutul de amidon al liniilor consangvinizate isonucleare TC 209, SCDA Turda, 2009-
2010
Linia consangvinizată
isonucleară
Conţinutul de amidon (%) la boabe provenite din autopolenizări
Conţinutul de amidon (%) la boabe provenite din polenizare liberă
Media pe tip de citoplasmă
2009 2010 Media
% faţă de
linia iniţială
2009 2010 Media
% faţă de
linia iniţială
Conţinut
mediu
Valoarea
relativă
TC 209 (martor) 69,33 71,60 70,47 100,0 69,73 71,17 70,45 100,0 70,46 100,0
TC 209 (cit. A665) 70,70 72,47 71,59** 101,6 71,57 71,87 71,72*** 101,8 71,65 101,7
TC 209 (cit. T291) 69,03 71,20 70,12 99,5 68,90 70,77 69,84o 99,1 69,98 99,3
TC 209 (cit. T248) 69,67 71,37 70,52 100,1 71,50 68,80 70,15 99,6 70,34 99,8
TC 209 (cit. W633) 68,63 71,73 70,18 99,6 70,53 69,47 70,00 99,4 70,09 99,5
TC 209 (cit. TC177) 68,27 70,07 69,17ooo 98,2 70,07 69,07 69,57oo 98,8 69,37 98,5
TC 209 (cit. D105) 69,63 70,70 70,17 99,6 73,57 69,60 71,59*** 101,6 70,88 100,6
Media liniilor isonucleare 69,32 71,31 70,31 70,84 70,11 70,47 70,39
DL (P=5%) 0,73 0,63
DL (P=1%) 0,97 0,84
DL (P=0.1%) 1,25 1,09
La linia consangvinizată generatoare de grupa TC 209, media continutului de
amidon pentru cele patru condiţii a fost de 70,46% (tabelul 11). O îmbunătăţire
semnificativă s-a înregistrat prin transferul pe citoplasma A665, atât la boabe
provenite de sub izolatori (71,59% - diferenţa distinct semnificativă pozitivă faţă de
martor), cât şi la boabe provenite din polenizare liberă (71,72%, faţă de 70,45%,
diferenţă distinct semnificativă statistic). În schimb, la linia TC 209 citoplasmă TC177
19
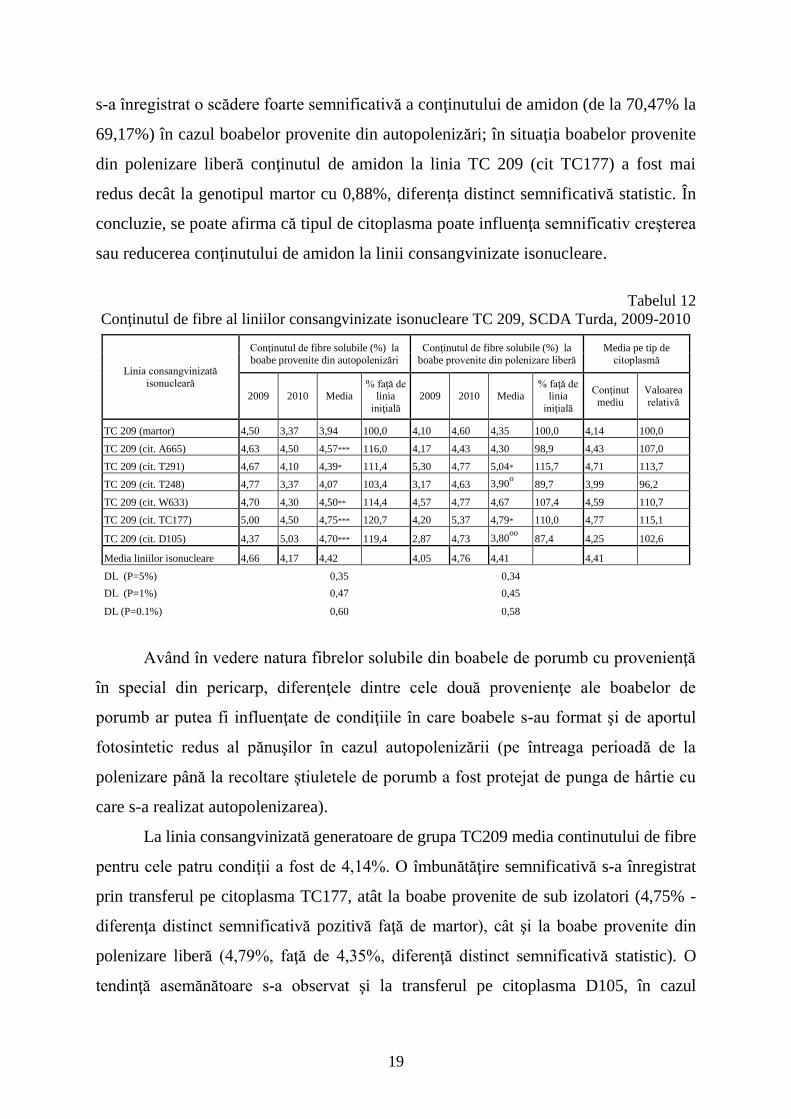
s-a înregistrat o scădere foarte semnificativă a conţinutului de amidon (de la 70,47% la
69,17%) în cazul boabelor provenite din autopolenizări; în situaţia boabelor provenite
din polenizare liberă conţinutul de amidon la linia TC 209 (cit TC177) a fost mai
redus decât la genotipul martor cu 0,88%, diferenţa distinct semnificativă statistic. În
concluzie, se poate afirma că tipul de citoplasma poate influenţa semnificativ creşterea
sau reducerea conţinutului de amidon la linii consangvinizate isonucleare.
Tabelul 12
Conţinutul de fibre al liniilor consangvinizate isonucleare TC 209, SCDA Turda, 2009-2010
Linia consangvinizată
isonucleară
Conţinutul de fibre solubile (%) la
boabe provenite din autopolenizări
Conţinutul de fibre solubile (%) la
boabe provenite din polenizare liberă
Media pe tip de
citoplasmă
2009 2010 Media % faţă de
linia
iniţială
2009 2010 Media % faţă de
linia
iniţială
Conţinut
mediu
Valoarea
relativă
TC 209 (martor) 4,50 3,37 3,94 100,0 4,10 4,60 4,35 100,0 4,14 100,0
TC 209 (cit. A665) 4,63 4,50 4,57*** 116,0 4,17 4,43 4,30 98,9 4,43 107,0
TC 209 (cit. T291) 4,67 4,10 4,39* 111,4 5,30 4,77 5,04* 115,7 4,71 113,7
TC 209 (cit. T248) 4,77 3,37 4,07 103,4 3,17 4,63 3,90o 89,7 3,99 96,2
TC 209 (cit. W633) 4,70 4,30 4,50** 114,4 4,57 4,77 4,67 107,4 4,59 110,7
TC 209 (cit. TC177) 5,00 4,50 4,75*** 120,7 4,20 5,37 4,79* 110,0 4,77 115,1
TC 209 (cit. D105) 4,37 5,03 4,70*** 119,4 2,87 4,73 3,80oo
87,4 4,25 102,6
Media liniilor isonucleare 4,66 4,17 4,42 4,05 4,76 4,41 4,41
DL (P=5%) 0,35 0,34
DL (P=1%) 0,47 0,45
DL (P=0.1%) 0,60 0,58
Având în vedere natura fibrelor solubile din boabele de porumb cu provenienţă
în special din pericarp, diferenţele dintre cele două provenienţe ale boabelor de
porumb ar putea fi influenţate de condiţiile în care boabele s-au format şi de aportul
fotosintetic redus al pănuşilor în cazul autopolenizării (pe întreaga perioadă de la
polenizare până la recoltare ştiuletele de porumb a fost protejat de punga de hârtie cu
care s-a realizat autopolenizarea).
La linia consangvinizată generatoare de grupa TC209 media continutului de fibre
pentru cele patru condiţii a fost de 4,14%. O îmbunătăţire semnificativă s-a înregistrat
prin transferul pe citoplasma TC177, atât la boabe provenite de sub izolatori (4,75% -
diferenţa distinct semnificativă pozitivă faţă de martor), cât şi la boabe provenite din
polenizare liberă (4,79%, faţă de 4,35%, diferenţă distinct semnificativă statistic). O
tendinţă asemănătoare s-a observat şi la transferul pe citoplasma D105, în cazul
20
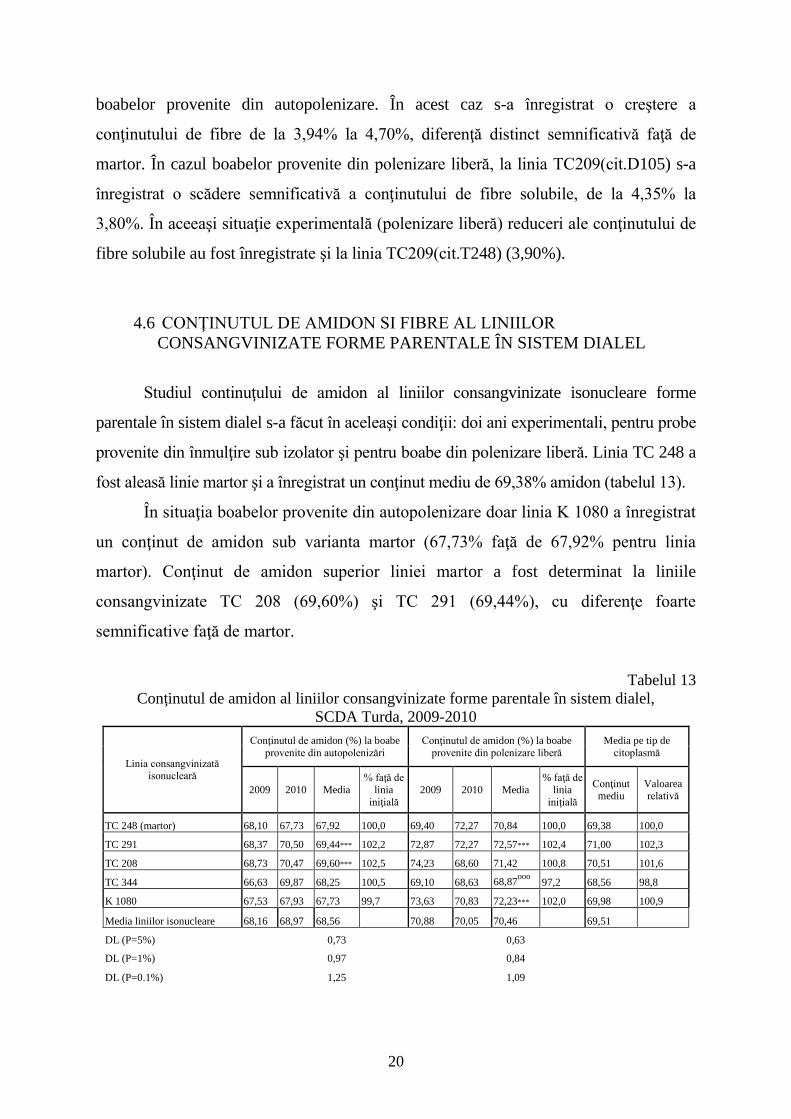
boabelor provenite din autopolenizare. În acest caz s-a înregistrat o creştere a
conţinutului de fibre de la 3,94% la 4,70%, diferenţă distinct semnificativă faţă de
martor. În cazul boabelor provenite din polenizare liberă, la linia TC209(cit.D105) s-a
înregistrat o scădere semnificativă a conţinutului de fibre solubile, de la 4,35% la
3,80%. În aceeaşi situaţie experimentală (polenizare liberă) reduceri ale conţinutului de
fibre solubile au fost înregistrate şi la linia TC209(cit.T248) (3,90%).
4.6 CONŢINUTUL DE AMIDON SI FIBRE AL LINIILOR
CONSANGVINIZATE FORME PARENTALE ÎN SISTEM DIALEL
Studiul continuţului de amidon al liniilor consangvinizate isonucleare forme
parentale în sistem dialel s-a făcut în aceleaşi condiţii: doi ani experimentali, pentru probe
provenite din înmulţire sub izolator şi pentru boabe din polenizare liberă. Linia TC 248 a
fost aleasă linie martor şi a înregistrat un conţinut mediu de 69,38% amidon (tabelul 13).
În situaţia boabelor provenite din autopolenizare doar linia K 1080 a înregistrat
un conţinut de amidon sub varianta martor (67,73% faţă de 67,92% pentru linia
martor). Conţinut de amidon superior liniei martor a fost determinat la liniile
consangvinizate TC 208 (69,60%) şi TC 291 (69,44%), cu diferenţe foarte
semnificative faţă de martor.
Tabelul 13
Conţinutul de amidon al liniilor consangvinizate forme parentale în sistem dialel,
SCDA Turda, 2009-2010
Linia consangvinizată isonucleară
Conţinutul de amidon (%) la boabe
provenite din autopolenizări
Conţinutul de amidon (%) la boabe
provenite din polenizare liberă
Media pe tip de
citoplasmă
2009 2010 Media
% faţă de
linia
iniţială
2009 2010 Media
% faţă de
linia
iniţială
Conţinut mediu
Valoarea relativă
TC 248 (martor) 68,10 67,73 67,92 100,0 69,40 72,27 70,84 100,0 69,38 100,0
TC 291 68,37 70,50 69,44*** 102,2 72,87 72,27 72,57*** 102,4 71,00 102,3
TC 208 68,73 70,47 69,60*** 102,5 74,23 68,60 71,42 100,8 70,51 101,6
TC 344 66,63 69,87 68,25 100,5 69,10 68,63 68,87ooo
97,2 68,56 98,8
K 1080 67,53 67,93 67,73 99,7 73,63 70,83 72,23*** 102,0 69,98 100,9
Media liniilor isonucleare 68,16 68,97 68,56 70,88 70,05 70,46 69,51
DL (P=5%) 0,73 0,63
DL (P=1%) 0,97 0,84
DL (P=0.1%) 1,25 1,09
21
În situaţia boabelor provenite din polenizare liberă conţinutul de amidon a fost
superior condiţiilor de autopolenizare, cu o medie de 70,84% amidon la linia martor.
În acest caz, linia care a prezentat o valoare inferioara a conţinutului de amidon faţă de
linia martor a fost TC 344 (68,87%, diferenţă foarte semnificativă faţă de martor).
Celelalte linii analizate au prezentat valori superioare ale conţinutului de amidon faţă
de linia martor, însă cele care s-au remarcat cu conţinut ridicat au fost TC 291
(72,57%) şi K 1080 (72,23%), cu diferenţe foarte semnificative faţă de linia martor.
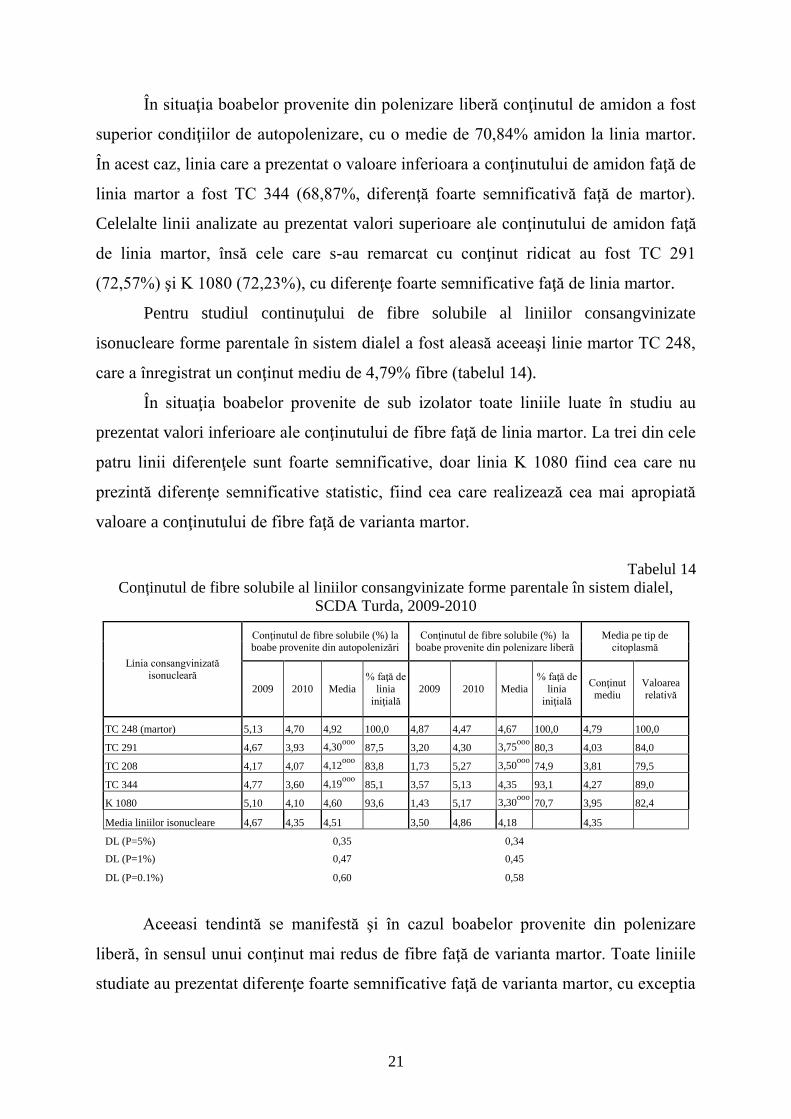
Pentru studiul continuţului de fibre solubile al liniilor consangvinizate
isonucleare forme parentale în sistem dialel a fost aleasă aceeaşi linie martor TC 248,
care a înregistrat un conţinut mediu de 4,79% fibre (tabelul 14).
În situaţia boabelor provenite de sub izolator toate liniile luate în studiu au
prezentat valori inferioare ale conţinutului de fibre faţă de linia martor. La trei din cele
patru linii diferenţele sunt foarte semnificative, doar linia K 1080 fiind cea care nu
prezintă diferenţe semnificative statistic, fiind cea care realizează cea mai apropiată
valoare a conţinutului de fibre faţă de varianta martor.
Tabelul 14
Conţinutul de fibre solubile al liniilor consangvinizate forme parentale în sistem dialel,
SCDA Turda, 2009-2010
Linia consangvinizată isonucleară
Conţinutul de fibre solubile (%) la boabe provenite din autopolenizări
Conţinutul de fibre solubile (%) la boabe provenite din polenizare liberă
Media pe tip de citoplasmă
2009 2010 Media
% faţă de
linia
iniţială
2009 2010 Media
% faţă de
linia
iniţială
Conţinut mediu
Valoarea relativă
TC 248 (martor) 5,13 4,70 4,92 100,0 4,87 4,47 4,67 100,0 4,79 100,0
TC 291 4,67 3,93 4,30ooo
87,5 3,20 4,30 3,75ooo
80,3 4,03 84,0
TC 208 4,17 4,07 4,12ooo
83,8 1,73 5,27 3,50ooo
74,9 3,81 79,5
TC 344 4,77 3,60 4,19ooo
85,1 3,57 5,13 4,35 93,1 4,27 89,0
K 1080 5,10 4,10 4,60 93,6 1,43 5,17 3,30ooo
70,7 3,95 82,4
Media liniilor isonucleare 4,67 4,35 4,51 3,50 4,86 4,18 4,35
DL (P=5%) 0,35 0,34
DL (P=1%) 0,47 0,45
DL (P=0.1%) 0,60 0,58
Aceeasi tendintă se manifestă şi în cazul boabelor provenite din polenizare
liberă, în sensul unui conţinut mai redus de fibre faţă de varianta martor. Toate liniile
studiate au prezentat diferenţe foarte semnificative faţă de varianta martor, cu exceptia
22
liniei TC 344, care a înregistrat un conţinut de fibre apropiat de cel al liniei martor
(4,35% faţă de 4,67%, diferenţă nesemnificativă statistic).
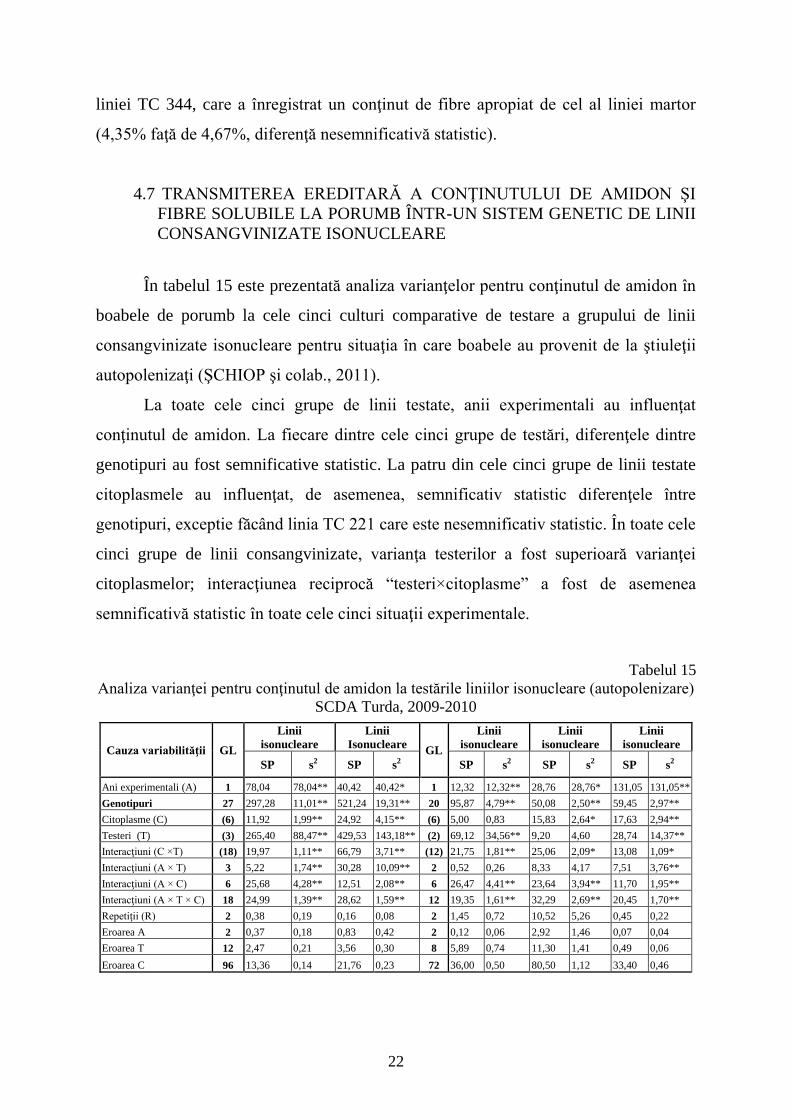
4.7 TRANSMITEREA EREDITARĂ A CONŢINUTULUI DE AMIDON ŞI
FIBRE SOLUBILE LA PORUMB ÎNTR-UN SISTEM GENETIC DE LINII
CONSANGVINIZATE ISONUCLEARE
În tabelul 15 este prezentată analiza varianţelor pentru conţinutul de amidon în
boabele de porumb la cele cinci culturi comparative de testare a grupului de linii
consangvinizate isonucleare pentru situaţia în care boabele au provenit de la ştiuleţii
autopolenizaţi (ŞCHIOP şi colab., 2011).
La toate cele cinci grupe de linii testate, anii experimentali au influenţat
conţinutul de amidon. La fiecare dintre cele cinci grupe de testări, diferenţele dintre
genotipuri au fost semnificative statistic. La patru din cele cinci grupe de linii testate
citoplasmele au influenţat, de asemenea, semnificativ statistic diferenţele între
genotipuri, exceptie făcând linia TC 221 care este nesemnificativ statistic. În toate cele
cinci grupe de linii consangvinizate, varianţa testerilor a fost superioară varianţei
citoplasmelor; interacţiunea reciprocă “testeri×citoplasme” a fost de asemenea
semnificativă statistic în toate cele cinci situaţii experimentale.
Tabelul 15
Analiza varianţei pentru conţinutul de amidon la testările liniilor isonucleare (autopolenizare)
SCDA Turda, 2009-2010
Cauza variabilităţii GL
Linii
isonucleare
Linii
Isonucleare GL
Linii
isonucleare
Linii
isonucleare
Linii
isonucleare
SP s2 SP s2 SP s2 SP s2 SP s2
Ani experimentali (A) 1 78,04 78,04** 40,42 40,42* 1 12,32 12,32** 28,76 28,76* 131,05 131,05**
Genotipuri 27 297,28 11,01** 521,24 19,31** 20 95,87 4,79** 50,08 2,50** 59,45 2,97**
Citoplasme (C) (6) 11,92 1,99** 24,92 4,15** (6) 5,00 0,83 15,83 2,64* 17,63 2,94**
Testeri (T) (3) 265,40 88,47** 429,53 143,18** (2) 69,12 34,56** 9,20 4,60 28,74 14,37**
Interacţiuni (C ×T) (18) 19,97 1,11** 66,79 3,71** (12) 21,75 1,81** 25,06 2,09* 13,08 1,09*
Interacţiuni (A × T) 3 5,22 1,74** 30,28 10,09** 2 0,52 0,26 8,33 4,17 7,51 3,76**
Interacţiuni (A × C) 6 25,68 4,28** 12,51 2,08** 6 26,47 4,41** 23,64 3,94** 11,70 1,95**
Interacţiuni (A × T × C) 18 24,99 1,39** 28,62 1,59** 12 19,35 1,61** 32,29 2,69** 20,45 1,70**
Repetiţii (R) 2 0,38 0,19 0,16 0,08 2 1,45 0,72 10,52 5,26 0,45 0,22
Eroarea A 2 0,37 0,18 0,83 0,42 2 0,12 0,06 2,92 1,46 0,07 0,04
Eroarea T 12 2,47 0,21 3,56 0,30 8 5,89 0,74 11,30 1,41 0,49 0,06
Eroarea C 96 13,36 0,14 21,76 0,23 72 36,00 0,50 80,50 1,12 33,40 0,46

23
Anii experimentali influenţează rolul citoplasmei şi interactiunea “citoplasmă ×
tester”; de asemenea, acţiunile genice transmise cu ajutorul genelor localizate în
nucleul testerilor este influenţată de anii experimentali pentru trei din liniile
isonucleare testate: TC 209, TC 243 şi D 105.
În figura 1 este prezentată ponderea factorilor implicaţi în varianţa conţinutului
de amidon la fiecare din cele cinci grupe de testare, în situaţia de autopolenizare.
Calculul ponderii factorilor implicaţi în varianţa conţinutului de amidon s-a făcut după
modelul propus de LEIN, citat de CEAPOIU (1969). Ponderea varianţei citoplasmelor
este cuprinsă între 4% (TC 209) şi 32% (TB 367), a varianţei testerilor între 18% (TB
367) şi 89% (TC 209), iar a interacţiunii “citoplasme×testeri” între 7% (TC 209) şi 50%
(TB 367).
TC 209 C
4%
T
89%
CxT
7%
TC 243C
5%
T
82%
CxT
13%
TC 221C
5%
T
72%
CxT
23%
TB 367C
32%
T
18%
CxT
50%
D 105C
30%
T
48%
CxT
22%
Figura 1. Ponderea factorilor implicaţi în variabilitatea conţinutului de amidon al boabelor de
porumb la cinci grupe de linii consangvinizate isonucleare (autopolenizare), SCDA Turda
Analiza varianţelor pentru conţinutul de fibre solubile în boabele de porumb la
cele cinci culturi comparative de testare a grupului de linii consangvinizate isonucleare
pentru situaţia în care boabele au provenit de la stiuleţii autopolenizaţi sunt prezentate în
tabelul 16.
În patru din cele cinci grupe de linii testate, anii experimentali au influenţat
conţinutul de fibre. La fiecare dintre cele cinci grupe de testări, diferenţele dintre
genotipuri au fost semnificative statistic. Tipurile de citoplasme au influenţat, de
asemenea, semnificativ statistic diferenţele între genotipuri. În toate cele cinci grupe
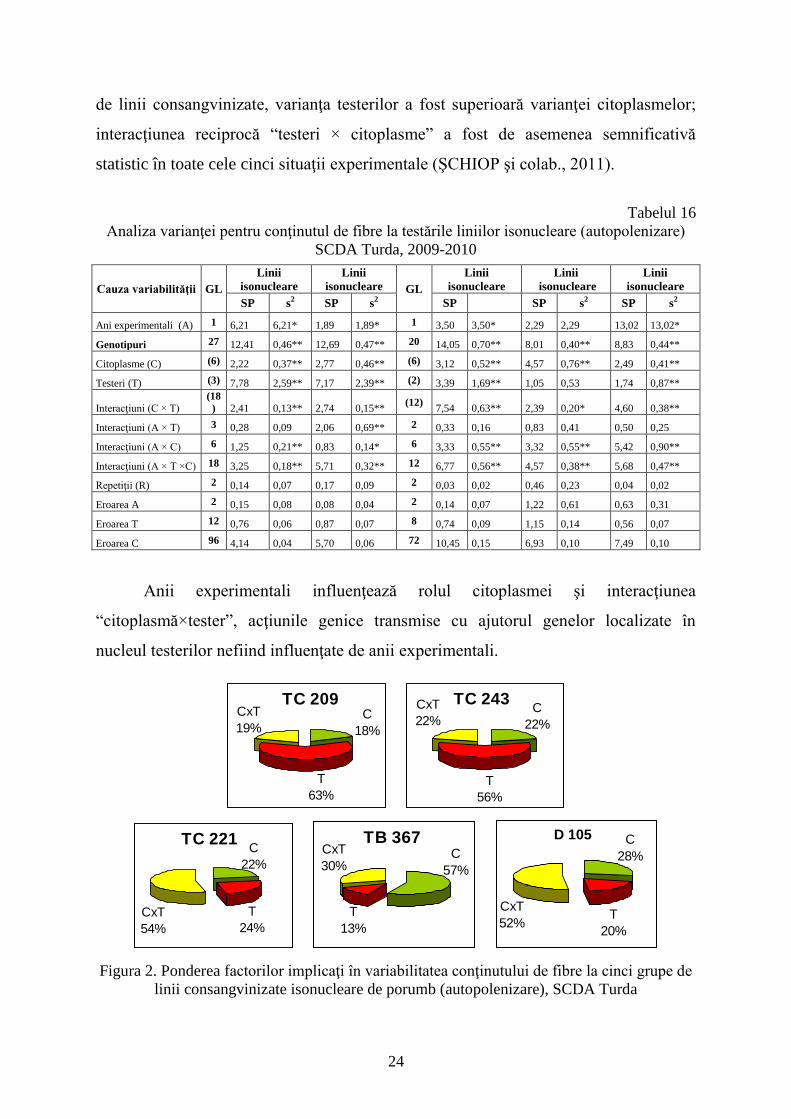
24
de linii consangvinizate, varianţa testerilor a fost superioară varianţei citoplasmelor;
interacţiunea reciprocă “testeri × citoplasme” a fost de asemenea semnificativă
statistic în toate cele cinci situaţii experimentale (ŞCHIOP şi colab., 2011).
Tabelul 16
Analiza varianţei pentru conţinutul de fibre la testările liniilor isonucleare (autopolenizare)
SCDA Turda, 2009-2010
Cauza variabilităţii GL
Linii
isonucleare
Linii
isonucleare GL
Linii
isonucleare
Linii
isonucleare
Linii
isonucleare
SP s2 SP s2
SP SP s2 SP s2
Ani experimentali (A) 1 6,21 6,21* 1,89 1,89* 1 3,50 3,50* 2,29 2,29 13,02 13,02*
Genotipuri 27 12,41 0,46** 12,69 0,47** 20 14,05 0,70** 8,01 0,40** 8,83 0,44**
Citoplasme (C) (6) 2,22 0,37** 2,77 0,46** (6) 3,12 0,52** 4,57 0,76** 2,49 0,41**
Testeri (T) (3) 7,78 2,59** 7,17 2,39** (2) 3,39 1,69** 1,05 0,53 1,74 0,87**
Interacţiuni (C × T) (18
) 2,41 0,13** 2,74 0,15** (12)
7,54 0,63** 2,39 0,20* 4,60 0,38**
Interacţiuni (A × T) 3 0,28 0,09 2,06 0,69** 2 0,33 0,16 0,83 0,41 0,50 0,25
Interacţiuni (A × C) 6 1,25 0,21** 0,83 0,14* 6 3,33 0,55** 3,32 0,55** 5,42 0,90**
Interacţiuni (A × T ×C) 18 3,25 0,18** 5,71 0,32** 12 6,77 0,56** 4,57 0,38** 5,68 0,47**
Repetiţii (R) 2 0,14 0,07 0,17 0,09 2 0,03 0,02 0,46 0,23 0,04 0,02
Eroarea A 2 0,15 0,08 0,08 0,04 2 0,14 0,07 1,22 0,61 0,63 0,31
Eroarea T 12 0,76 0,06 0,87 0,07 8 0,74 0,09 1,15 0,14 0,56 0,07
Eroarea C 96 4,14 0,04 5,70 0,06 72 10,45 0,15 6,93 0,10 7,49 0,10
Anii experimentali influenţează rolul citoplasmei şi interacţiunea
“citoplasmă×tester”, acţiunile genice transmise cu ajutorul genelor localizate în
nucleul testerilor nefiind influenţate de anii experimentali.
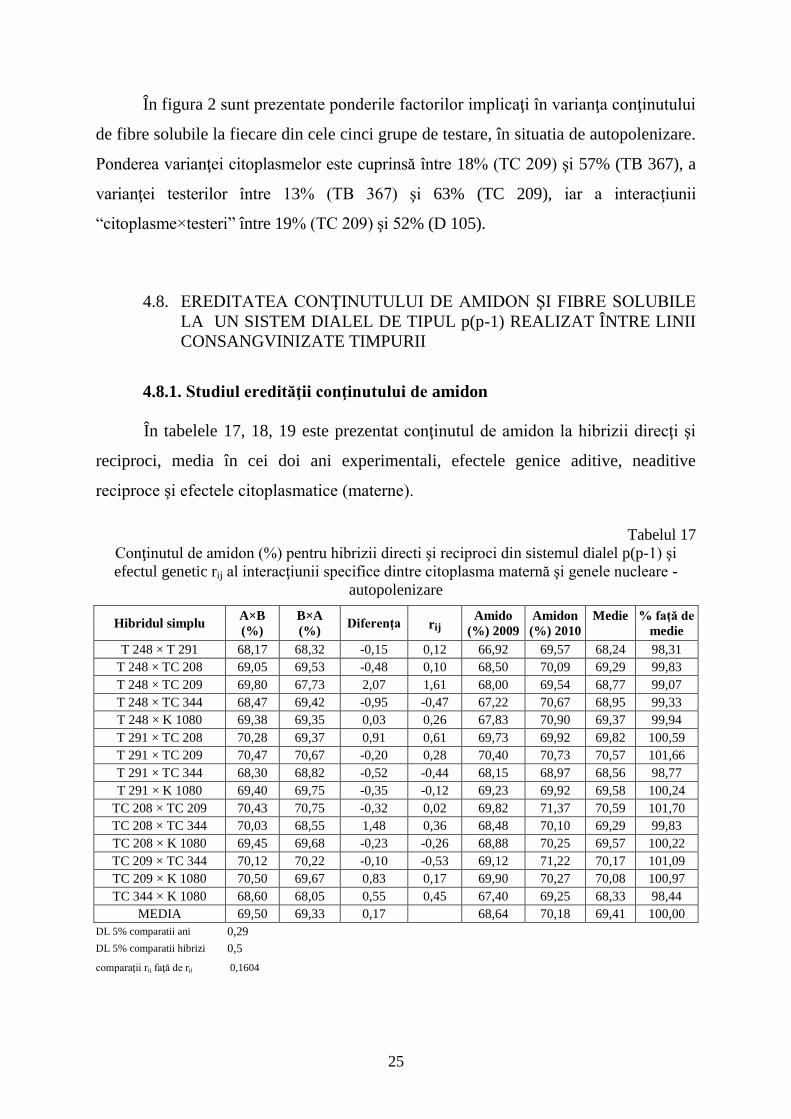
TC 209CxT
19%
T
63%
C
18%
TC 243C
22%
T
56%
CxT
22%
TC 221C
22%
T
24%
CxT
54%
TB 367C
57%
T
13%
CxT
30%
D 105 C
28%
T
20%
CxT
52%
Figura 2. Ponderea factorilor implicaţi în variabilitatea conţinutului de fibre la cinci grupe de
linii consangvinizate isonucleare de porumb (autopolenizare), SCDA Turda
25
În figura 2 sunt prezentate ponderile factorilor implicaţi în varianţa conţinutului
de fibre solubile la fiecare din cele cinci grupe de testare, în situatia de autopolenizare.
Ponderea varianţei citoplasmelor este cuprinsă între 18% (TC 209) şi 57% (TB 367), a
varianţei testerilor între 13% (TB 367) şi 63% (TC 209), iar a interacţiunii
“citoplasme×testeri” între 19% (TC 209) şi 52% (D 105).
4.8. EREDITATEA CONŢINUTULUI DE AMIDON ŞI FIBRE SOLUBILE
LA UN SISTEM DIALEL DE TIPUL p(p-1) REALIZAT ÎNTRE LINII
CONSANGVINIZATE TIMPURII
4.8.1. Studiul eredităţii conţinutului de amidon
În tabelele 17, 18, 19 este prezentat conţinutul de amidon la hibrizii direcţi şi
reciproci, media în cei doi ani experimentali, efectele genice aditive, neaditive
reciproce şi efectele citoplasmatice (materne).
Tabelul 17
Conţinutul de amidon (%) pentru hibrizii directi şi reciproci din sistemul dialel p(p-1) şi
efectul genetic rij al interacţiunii specifice dintre citoplasma maternă şi genele nucleare -
autopolenizare
Hibridul simplu A×B
(%)
B×A
(%) Diferenţa rij
Amido
(%) 2009
Amidon
(%) 2010
Medie
% faţă de
medie
T 248 × T 291 68,17 68,32 -0,15 0,12 66,92 69,57 68,24 98,31
T 248 × TC 208 69,05 69,53 -0,48 0,10 68,50 70,09 69,29 99,83
T 248 × TC 209 69,80 67,73 2,07 1,61 68,00 69,54 68,77 99,07
T 248 × TC 344 68,47 69,42 -0,95 -0,47 67,22 70,67 68,95 99,33
T 248 × K 1080 69,38 69,35 0,03 0,26 67,83 70,90 69,37 99,94
T 291 × TC 208 70,28 69,37 0,91 0,61 69,73 69,92 69,82 100,59
T 291 × TC 209 70,47 70,67 -0,20 0,28 70,40 70,73 70,57 101,66
T 291 × TC 344 68,30 68,82 -0,52 -0,44 68,15 68,97 68,56 98,77
T 291 × K 1080 69,40 69,75 -0,35 -0,12 69,23 69,92 69,58 100,24
TC 208 × TC 209 70,43 70,75 -0,32 0,02 69,82 71,37 70,59 101,70
TC 208 × TC 344 70,03 68,55 1,48 0,36 68,48 70,10 69,29 99,83
TC 208 × K 1080 69,45 69,68 -0,23 -0,26 68,88 70,25 69,57 100,22
TC 209 × TC 344 70,12 70,22 -0,10 -0,53 69,12 71,22 70,17 101,09
TC 209 × K 1080 70,50 69,67 0,83 0,17 69,90 70,27 70,08 100,97
TC 344 × K 1080 68,60 68,05 0,55 0,45 67,40 69,25 68,33 98,44
MEDIA 69,50 69,33 0,17 68,64 70,18 69,41 100,00
DL 5% comparatii ani 0,29
DL 5% comparatii hibrizi 0,5
comparaţii rij faţă de rji 0,1604
26
Conţinutul de amidon a fost cuprins între 68,24% şi 70,59%; conţinutul cel mai
ridicat s-a înregistrat la combinaţia hibridă TC208×TC209, iar cel mai scăzut la
hibridul T248×T291. Anul experimental 2010 a fost mai favorabil acumulării
amidonului în boabe, diferenţa faţă de anul 2009 de 1,54% fiind semnificativă
statistic. Amplitudinea conţinutului în anul favorabil a fost cuprinsă între 68,97% la
hibridul T291×TC344 şi 71,37% la hibridul TC208×TC209, deci 2,4 unităţi
procentuale. În anul 2009, mai puţin favorabil acumulării amidonului în bob,
amplitudinea conţinutului a fost mai ridicată, între 66,92% la hibridul T248×T291 şi
70,40% la hibridul T291×TC209, deci 3,48 unităţi procentuale.
Între hibrizii direcţi şi reciproci s-au înregistrat, în cazul unora dintre ei,
diferenţe semnificative statistic: T248×TC209 (+2,07), TC208×TC344 (+1,48%),
T291×TC208 (+0,91%), TC209×K1080 (0,83%). Hibrizii reciproci au avut conţinut
semnificativ statistic mai ridicat, în cazul următorilor hibrizi : T248×TC344 (-0,95%),
T291×TC344 (-0,52%).
Diferenţele mari între hibrizii direcţi şi reciproci sunt explicabile prin valorile
ridicate ale interacţiunilor reciproce, nuclear-citoplasmatice (+1,61 la combinaţia hibridă
T248 ×TC209, +0,61 la combinaţia hibridă T291×TC208, +0,45 la combinaţia hibridă
TC344×K1080, -0,53 la combinaţia hibridă TC209×TC344, -0,47 la combinaţia hibridă
T248×TC344, -0,44 la combinaţia hibridă T291×TC344).
Tabelul 18
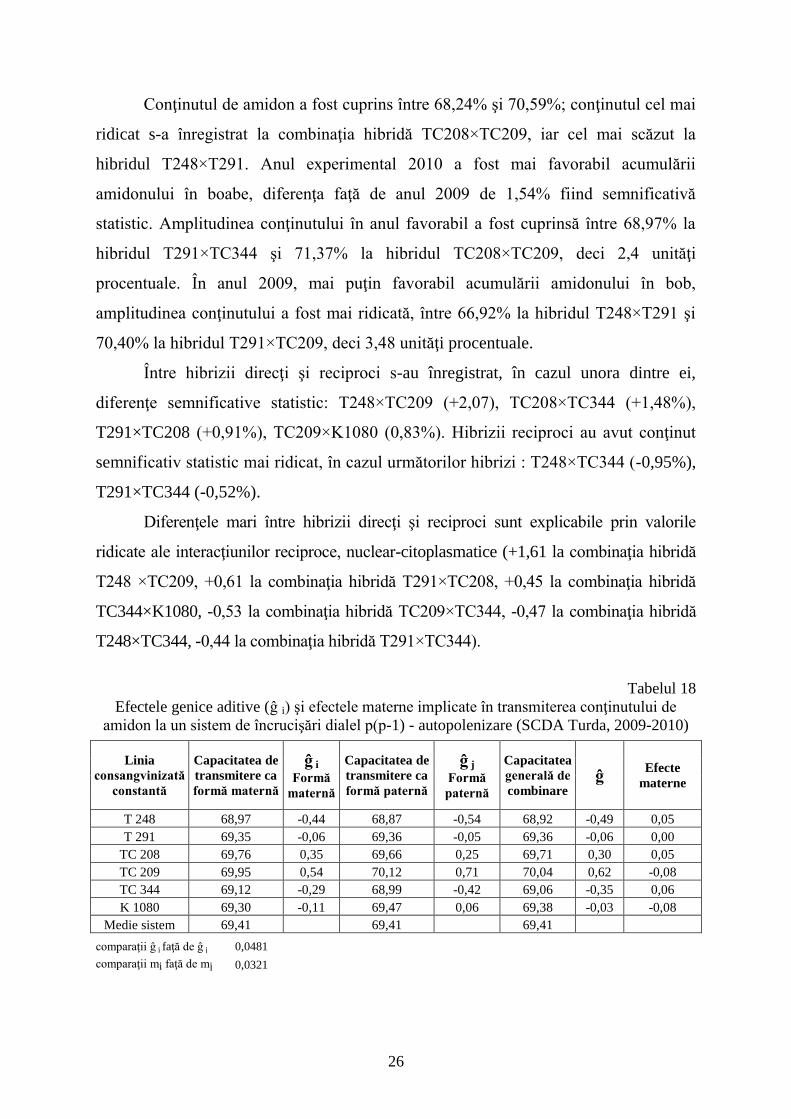
Efectele genice aditive (ĝ i) şi efectele materne implicate în transmiterea conţinutului de
amidon la un sistem de încrucişări dialel p(p-1) - autopolenizare (SCDA Turda, 2009-2010)
Linia
consangvinizată
constantă
Capacitatea de
transmitere ca
formă maternă
ĝ i Formă
maternă
Capacitatea de
transmitere ca
formă paternă
ĝ j Formă
paternă
Capacitatea
generală de
combinare ĝ
Efecte
materne
T 248 68,97 -0,44 68,87 -0,54 68,92 -0,49 0,05
T 291 69,35 -0,06 69,36 -0,05 69,36 -0,06 0,00
TC 208 69,76 0,35 69,66 0,25 69,71 0,30 0,05
TC 209 69,95 0,54 70,12 0,71 70,04 0,62 -0,08
TC 344 69,12 -0,29 68,99 -0,42 69,06 -0,35 0,06
K 1080 69,30 -0,11 69,47 0,06 69,38 -0,03 -0,08
Medie sistem 69,41 69,41 69,41
comparaţii ĝ i faţă de ĝ j 0,0481
comparaţii mi faţă de mj 0,0321
27
Efectele materne au avut valori reduse cuprinse între 0,06 la TC344 şi -0,08 la
TC209 şi K1080; se poate afirma că importanţa interacţiunilor reciproce nuclear-
citoplasmatice este mai ridicată decât efectele materne (citoplasmatice). Efectele genice
aditive sunt destul de ridicate, cuprinse între -0,49 pentru linia consangvinizată T248 şi
+0,62 pentru TC209. Sensul şi valorile absolute ale efectelor aditive calculate pentru
liniile consangvinizate în funcţie de poziţia în hibrid (forma maternă sau paternă) sunt
asemănătoare, indicând faptul că în transmiterea acestui caracter, genele cu poziţionare
nucleară sunt importante.
Acelaşi lucru este relevat şi de valorile efectelor genice neaditive, cu valori destul
de importante, cuprinse între -0,78 (T248×TC209) şi +0,59 (T291×TC209).
Tabelul 19
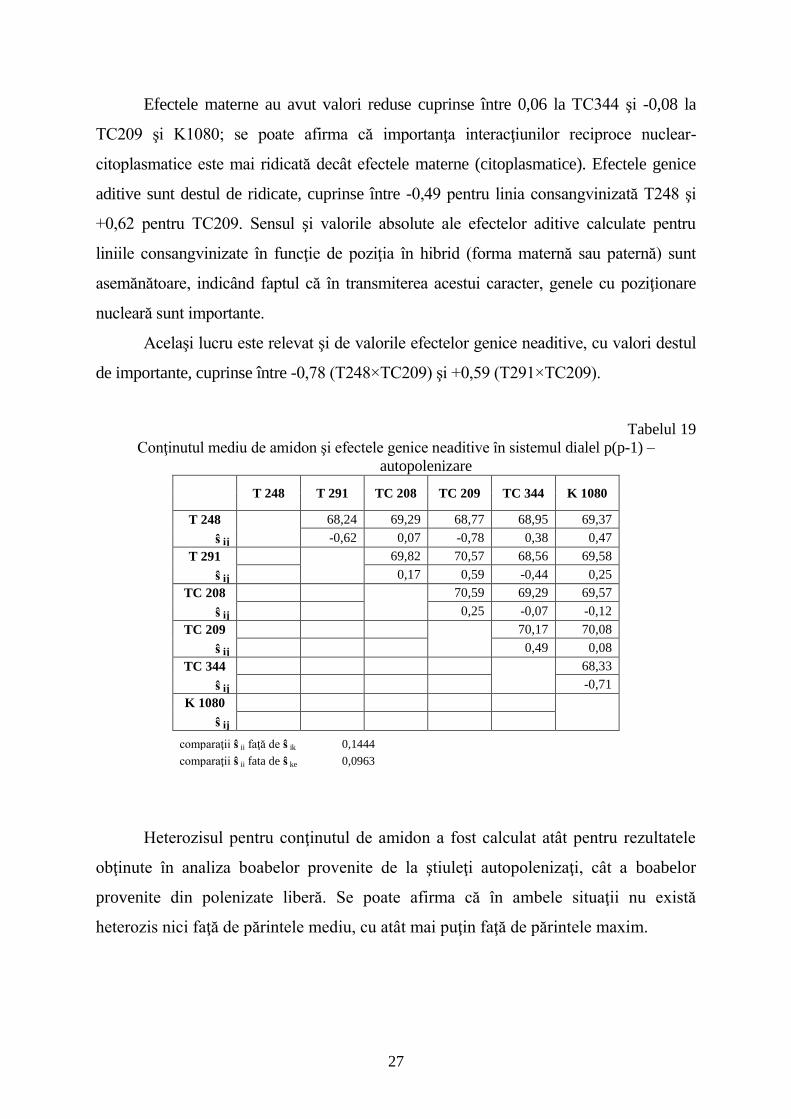
Conţinutul mediu de amidon şi efectele genice neaditive în sistemul dialel p(p-1) –
autopolenizare
T 248 T 291 TC 208 TC 209 TC 344 K 1080
T 248 68,24 69,29 68,77 68,95 69,37
ŝ ij -0,62 0,07 -0,78 0,38 0,47
T 291 69,82 70,57 68,56 69,58
ŝ ij 0,17 0,59 -0,44 0,25
TC 208 70,59 69,29 69,57
ŝ ij 0,25 -0,07 -0,12
TC 209 70,17 70,08
ŝ ij 0,49 0,08
TC 344 68,33
ŝ ij -0,71
K 1080
ŝ ij
comparaţii ŝ ij faţă de ŝ ik 0,1444
comparaţii ŝ ij fata de ŝ ke 0,0963
Heterozisul pentru conţinutul de amidon a fost calculat atât pentru rezultatele
obţinute în analiza boabelor provenite de la ştiuleţi autopolenizaţi, cât a boabelor
provenite din polenizate liberă. Se poate afirma că în ambele situaţii nu există
heterozis nici faţă de părintele mediu, cu atât mai puţin faţă de părintele maxim.
28
Tabel 20
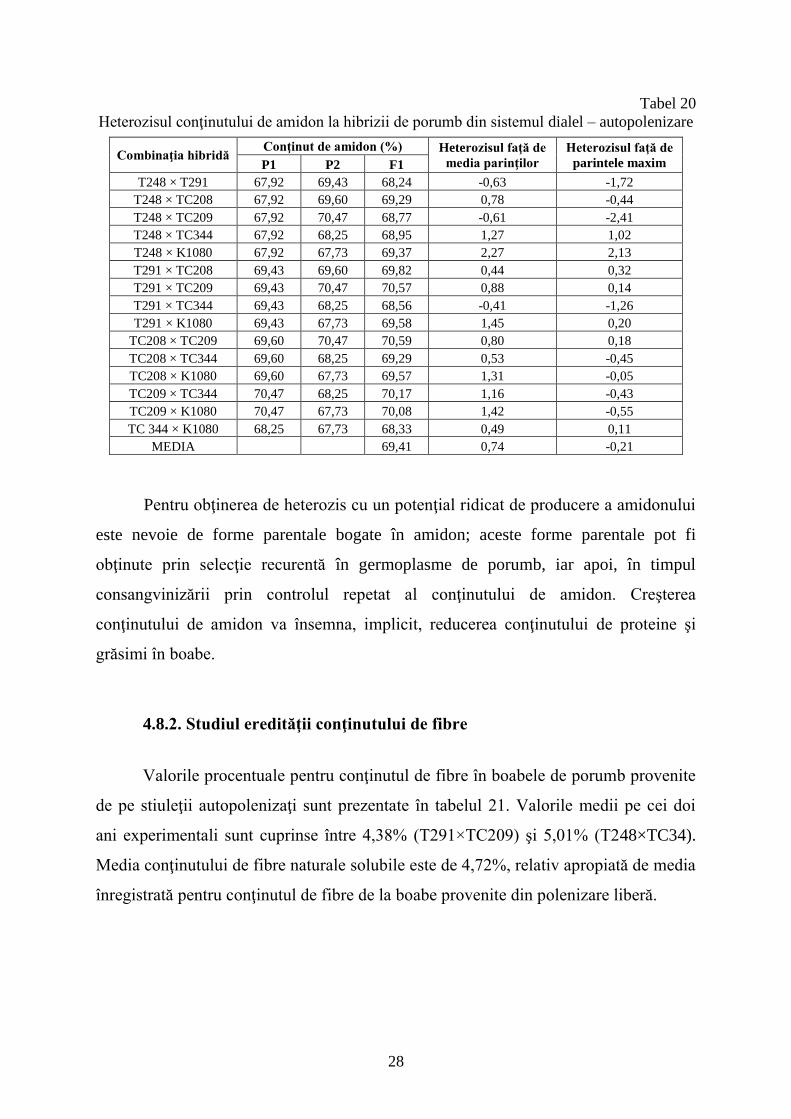
Heterozisul conţinutului de amidon la hibrizii de porumb din sistemul dialel – autopolenizare
Combinaţia hibridă Conţinut de amidon (%) Heterozisul faţă de
media parinţilor
Heterozisul faţă de
parintele maxim P1 P2 F1
T248 × T291 67,92 69,43 68,24 -0,63 -1,72
T248 × TC208 67,92 69,60 69,29 0,78 -0,44
T248 × TC209 67,92 70,47 68,77 -0,61 -2,41
T248 × TC344 67,92 68,25 68,95 1,27 1,02
T248 × K1080 67,92 67,73 69,37 2,27 2,13
T291 × TC208 69,43 69,60 69,82 0,44 0,32
T291 × TC209 69,43 70,47 70,57 0,88 0,14
T291 × TC344 69,43 68,25 68,56 -0,41 -1,26
T291 × K1080 69,43 67,73 69,58 1,45 0,20
TC208 × TC209 69,60 70,47 70,59 0,80 0,18
TC208 × TC344 69,60 68,25 69,29 0,53 -0,45
TC208 × K1080 69,60 67,73 69,57 1,31 -0,05
TC209 × TC344 70,47 68,25 70,17 1,16 -0,43
TC209 × K1080 70,47 67,73 70,08 1,42 -0,55
TC 344 × K1080 68,25 67,73 68,33 0,49 0,11
MEDIA 69,41 0,74 -0,21
Pentru obţinerea de heterozis cu un potenţial ridicat de producere a amidonului
este nevoie de forme parentale bogate în amidon; aceste forme parentale pot fi
obţinute prin selecţie recurentă în germoplasme de porumb, iar apoi, în timpul
consangvinizării prin controlul repetat al conţinutului de amidon. Creşterea
conţinutului de amidon va însemna, implicit, reducerea conţinutului de proteine şi
grăsimi în boabe.
4.8.2. Studiul eredităţii conţinutului de fibre
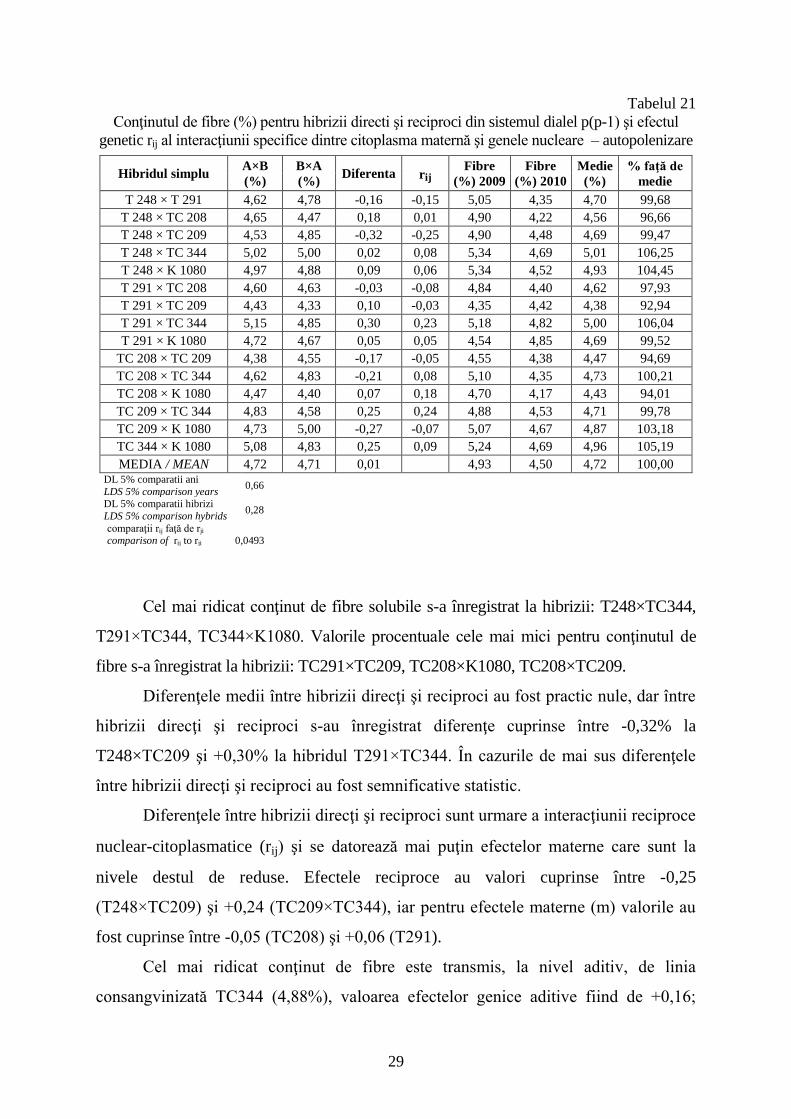
Valorile procentuale pentru conţinutul de fibre în boabele de porumb provenite
de pe stiuleţii autopolenizaţi sunt prezentate în tabelul 21. Valorile medii pe cei doi
ani experimentali sunt cuprinse între 4,38% (T291×TC209) şi 5,01% (T248×TC34).
Media conţinutului de fibre naturale solubile este de 4,72%, relativ apropiată de media
înregistrată pentru conţinutul de fibre de la boabe provenite din polenizare liberă.
29
Tabelul 21
Conţinutul de fibre (%) pentru hibrizii directi şi reciproci din sistemul dialel p(p-1) şi efectul
genetic rij al interacţiunii specifice dintre citoplasma maternă şi genele nucleare – autopolenizare
Hibridul simplu A×B
(%)
B×A
(%) Diferenta rij
Fibre
(%) 2009
Fibre
(%) 2010
Medie
(%)
% faţă de
medie
T 248 × T 291 4,62 4,78 -0,16 -0,15 5,05 4,35 4,70 99,68
T 248 × TC 208 4,65 4,47 0,18 0,01 4,90 4,22 4,56 96,66
T 248 × TC 209 4,53 4,85 -0,32 -0,25 4,90 4,48 4,69 99,47
T 248 × TC 344 5,02 5,00 0,02 0,08 5,34 4,69 5,01 106,25
T 248 × K 1080 4,97 4,88 0,09 0,06 5,34 4,52 4,93 104,45
T 291 × TC 208 4,60 4,63 -0,03 -0,08 4,84 4,40 4,62 97,93
T 291 × TC 209 4,43 4,33 0,10 -0,03 4,35 4,42 4,38 92,94
T 291 × TC 344 5,15 4,85 0,30 0,23 5,18 4,82 5,00 106,04
T 291 × K 1080 4,72 4,67 0,05 0,05 4,54 4,85 4,69 99,52
TC 208 × TC 209 4,38 4,55 -0,17 -0,05 4,55 4,38 4,47 94,69
TC 208 × TC 344 4,62 4,83 -0,21 0,08 5,10 4,35 4,73 100,21
TC 208 × K 1080 4,47 4,40 0,07 0,18 4,70 4,17 4,43 94,01
TC 209 × TC 344 4,83 4,58 0,25 0,24 4,88 4,53 4,71 99,78
TC 209 × K 1080 4,73 5,00 -0,27 -0,07 5,07 4,67 4,87 103,18
TC 344 × K 1080 5,08 4,83 0,25 0,09 5,24 4,69 4,96 105,19
MEDIA / MEAN 4,72 4,71 0,01 4,93 4,50 4,72 100,00 DL 5% comparatii ani
LDS 5% comparison years 0,66
DL 5% comparatii hibrizi
LDS 5% comparison hybrids 0,28
comparaţii rij faţă de rji
comparison of rij to rji 0,0493
Cel mai ridicat conţinut de fibre solubile s-a înregistrat la hibrizii: T248×TC344,
T291×TC344, TC344×K1080. Valorile procentuale cele mai mici pentru conţinutul de
fibre s-a înregistrat la hibrizii: TC291×TC209, TC208×K1080, TC208×TC209.
Diferenţele medii între hibrizii direcţi şi reciproci au fost practic nule, dar între
hibrizii direcţi şi reciproci s-au înregistrat diferenţe cuprinse între -0,32% la
T248×TC209 şi +0,30% la hibridul T291×TC344. În cazurile de mai sus diferenţele
între hibrizii direcţi şi reciproci au fost semnificative statistic.
Diferenţele între hibrizii direcţi şi reciproci sunt urmare a interacţiunii reciproce
nuclear-citoplasmatice (rij) şi se datorează mai puţin efectelor materne care sunt la
nivele destul de reduse. Efectele reciproce au valori cuprinse între -0,25
(T248×TC209) şi +0,24 (TC209×TC344), iar pentru efectele materne (m) valorile au
fost cuprinse între -0,05 (TC208) şi +0,06 (T291).
Cel mai ridicat conţinut de fibre este transmis, la nivel aditiv, de linia
consangvinizată TC344 (4,88%), valoarea efectelor genice aditive fiind de +0,16;
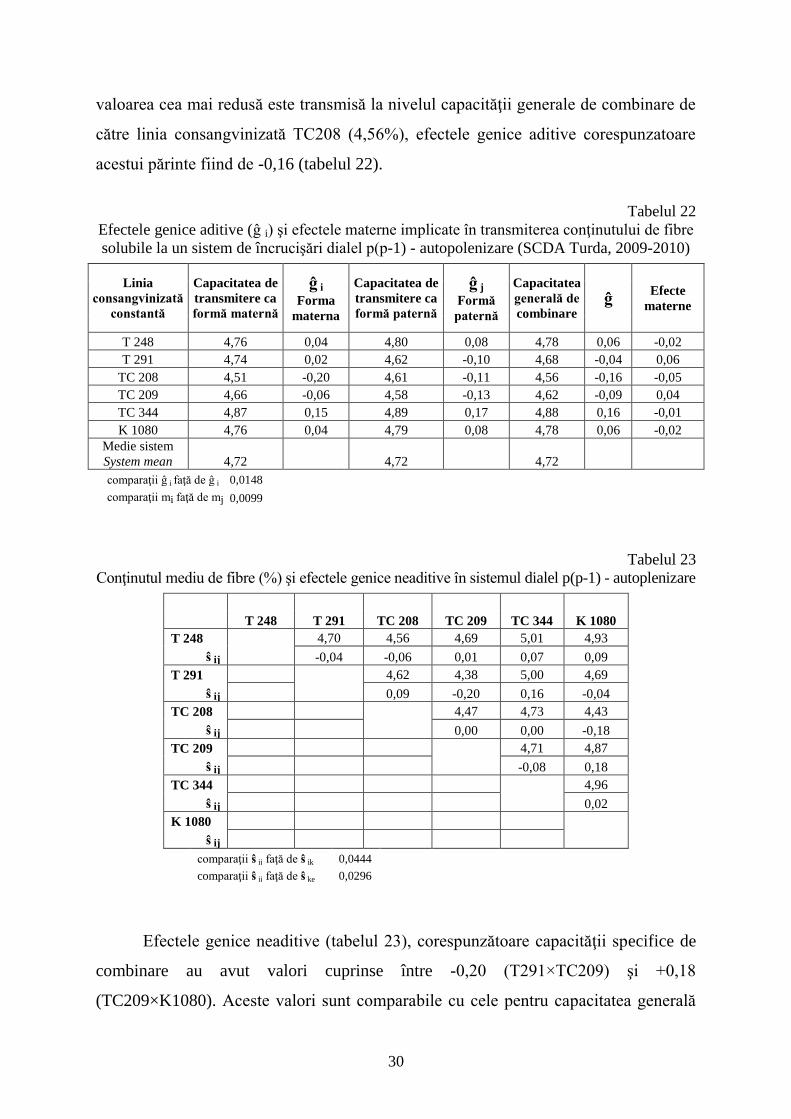
30
valoarea cea mai redusă este transmisă la nivelul capacităţii generale de combinare de
către linia consangvinizată TC208 (4,56%), efectele genice aditive corespunzatoare
acestui părinte fiind de -0,16 (tabelul 22).
Tabelul 22
Efectele genice aditive (ĝ i) şi efectele materne implicate în transmiterea conţinutului de fibre
solubile la un sistem de încrucişări dialel p(p-1) - autopolenizare (SCDA Turda, 2009-2010)
Linia
consangvinizată
constantă
Capacitatea de
transmitere ca
formă maternă
ĝ i Forma
materna
Capacitatea de
transmitere ca
formă paternă
ĝ j Formă
paternă
Capacitatea
generală de
combinare ĝ
Efecte
materne
T 248 4,76 0,04 4,80 0,08 4,78 0,06 -0,02
T 291 4,74 0,02 4,62 -0,10 4,68 -0,04 0,06
TC 208 4,51 -0,20 4,61 -0,11 4,56 -0,16 -0,05
TC 209 4,66 -0,06 4,58 -0,13 4,62 -0,09 0,04
TC 344 4,87 0,15 4,89 0,17 4,88 0,16 -0,01
K 1080 4,76 0,04 4,79 0,08 4,78 0,06 -0,02
Medie sistem
System mean 4,72 4,72 4,72
comparaţii ĝ i faţă de ĝ j 0,0148
comparaţii mi faţă de mj 0,0099
Tabelul 23
Conţinutul mediu de fibre (%) şi efectele genice neaditive în sistemul dialel p(p-1) - autoplenizare
T 248 T 291 TC 208 TC 209 TC 344 K 1080
T 248 4,70 4,56 4,69 5,01 4,93
ŝ ij -0,04 -0,06 0,01 0,07 0,09
T 291 4,62 4,38 5,00 4,69
ŝ ij 0,09 -0,20 0,16 -0,04
TC 208 4,47 4,73 4,43
ŝ ij 0,00 0,00 -0,18
TC 209 4,71 4,87
ŝ ij -0,08 0,18
TC 344 4,96
ŝ ij 0,02
K 1080
ŝ ij
comparaţii ŝ ij faţă de ŝ ik 0,0444
comparaţii ŝ ij faţă de ŝ ke 0,0296
Efectele genice neaditive (tabelul 23), corespunzătoare capacităţii specifice de
combinare au avut valori cuprinse între -0,20 (T291×TC209) şi +0,18
(TC209×K1080). Aceste valori sunt comparabile cu cele pentru capacitatea generală
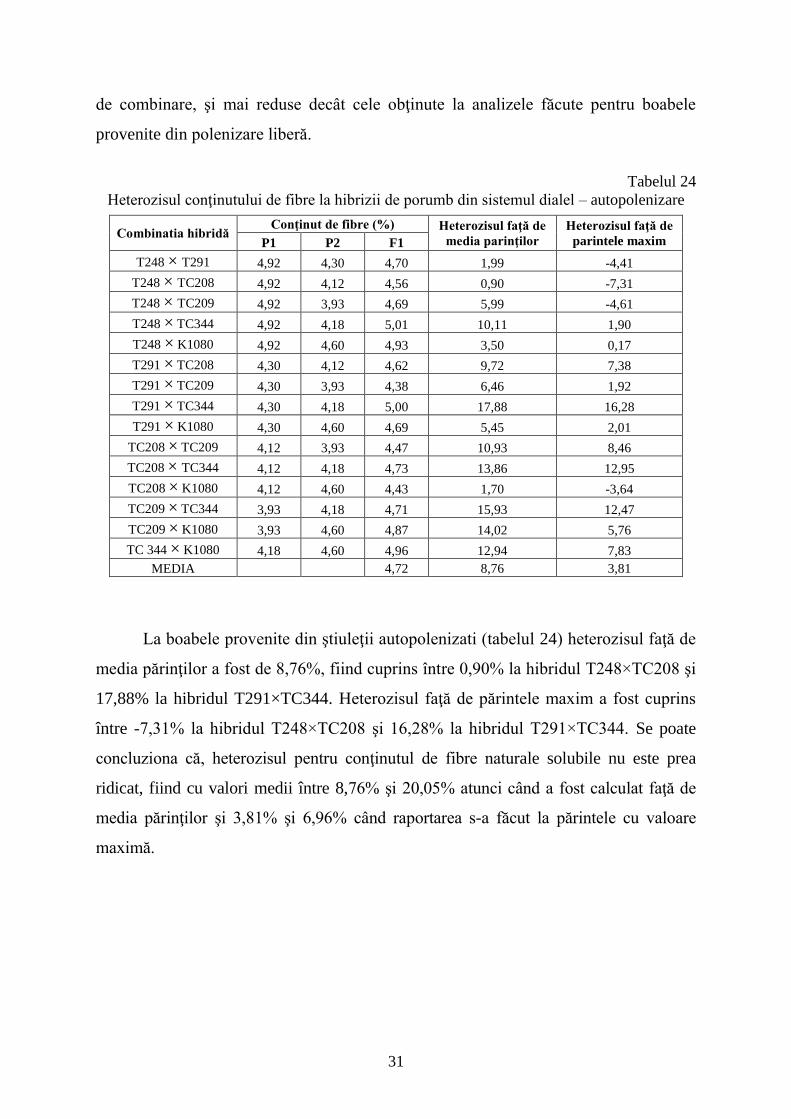
31
de combinare, şi mai reduse decât cele obţinute la analizele făcute pentru boabele
provenite din polenizare liberă.
Tabelul 24
Heterozisul conţinutului de fibre la hibrizii de porumb din sistemul dialel – autopolenizare
Combinatia hibridă Conţinut de fibre (%) Heterozisul faţă de
media parinţilor
Heterozisul faţă de
parintele maxim P1 P2 F1
T248 × T291 4,92 4,30 4,70 1,99 -4,41
T248 × TC208 4,92 4,12 4,56 0,90 -7,31
T248 × TC209 4,92 3,93 4,69 5,99 -4,61
T248 × TC344 4,92 4,18 5,01 10,11 1,90
T248 × K1080 4,92 4,60 4,93 3,50 0,17
T291 × TC208 4,30 4,12 4,62 9,72 7,38
T291 × TC209 4,30 3,93 4,38 6,46 1,92
T291 × TC344 4,30 4,18 5,00 17,88 16,28
T291 × K1080 4,30 4,60 4,69 5,45 2,01
TC208 × TC209 4,12 3,93 4,47 10,93 8,46
TC208 × TC344 4,12 4,18 4,73 13,86 12,95
TC208 × K1080 4,12 4,60 4,43 1,70 -3,64
TC209 × TC344 3,93 4,18 4,71 15,93 12,47
TC209 × K1080 3,93 4,60 4,87 14,02 5,76
TC 344 × K1080 4,18 4,60 4,96 12,94 7,83
MEDIA 4,72 8,76 3,81
La boabele provenite din ştiuleţii autopolenizati (tabelul 24) heterozisul faţă de
media părinţilor a fost de 8,76%, fiind cuprins între 0,90% la hibridul T248×TC208 şi
17,88% la hibridul T291×TC344. Heterozisul faţă de părintele maxim a fost cuprins
între -7,31% la hibridul T248×TC208 şi 16,28% la hibridul T291×TC344. Se poate
concluziona că, heterozisul pentru conţinutul de fibre naturale solubile nu este prea
ridicat, fiind cu valori medii între 8,76% şi 20,05% atunci când a fost calculat faţă de
media părinţilor şi 3,81% şi 6,96% când raportarea s-a făcut la părintele cu valoare
maximă.

32
CAPITOLUL V
CONCLUZII SI RECOMANDARI
1. Conţinutul de amidon al celor 258 de populaţii locale din colecţia SCDA Turda
a avut o medie de 64,9%, şi prezintă o variabilitate redusă, valoarea coeficientului de
variabilitate fiind de 2,93. În ceea ce priveste conţinutul de fibre al populatiilor locale,
acesta a avut o medie de 5,27%, iar abaterea standard a fost de 0,79%, fapt care arată că
majoritatea genotipurilor se grupează în jurul mediei, lucru întărit şi de valoarea mică a
varianţei, 0,63. Populaţiile locale prezintă o variabilitate mijlocie a conţinutului de fibre,
dată de valoarea coeficientului de variabilitate de 15,05.
2. Populaţiile sintetice create la S.C.D.A Turda au prezentat un o medie a
conţinutulului de amidon de 66,37%. Varianţa calculată a fost de 12,51%, ceea ce
reflectă o dispersie a valorilor individuale faţă de medie. Variabilitatea conţinutului de
amidon în rândul populaţiilor studiate este redusa (5,33%). În ceea ce priveşte
conţinutul de fibre al populaţiilor sintetice analizate, acestea au prezentat o
variabilitate ridicata, valoarea coeficientului de variabilitate fiind de 21,44. Valoarea
medie a conţinutului de fibre în rândul populaţiilor sintetice a fost de 5,17%.
3. La liniile consangvinizate create la SCDA Turda, conţinutul de amidon a
prezentat o variabilitate redusă (4,0%). Conţinutul mediu determinat este de 67,54%.
Conţinutul mediu de fibre determinat la liniile consangvinizate analizate a fost de
4,88%, şi a prezentat o variabilitate mijlocie a continutului de fibre in rândul (18,87).
4. În analiza conţinutului de amidon şi fibre la liniile consangvinizate
isonucleare în sistem balansat valoarea medie determinată pentru conţinutul de amidon
a fost de 66,37%, iar pentru fibre de 4,3%. Între genotipurile testate în toate cele cinci
situaţii experimentale diferenţele au fost distinct semnificative statistic.
5. Citoplasmele au influenţat semnificativ transmiterea ereditară a conţinutului de
amidon la cele cinci grupe de linii în sistem balansat, însă conţinutul de fibre nu este
influenţat de factorii citoplasmatici, dovadă fiind faptul că aceeaşi citoplasmă dă atât
valoarea cea mai mică, cât şi valoarea cea mai mare a conţinutului de fibre,
demonstrându-se astfel influenţa factorilor nucleari.

33
6. În analiza liniilor consangvinizate în sistem nebalansat, conţinutul mediu de
amidon a fost cuprins între 62,82% (pentru linia generatoare de grupă TB 367), şi
70,46% (linia generatoare de grupă TC209). Se poate afirma că tipul de citoplasmă
poate influenţa semnificativ creşterea sau reducerea conţinutului de amidon la linii
consangvinizate isonucleare.
7. Valorile medii determinate pentru fibre în analiza liniilor consangvinizate în
sistem nebalansat au fost cuprinse între 3,36% (pentru linia generatoare de grupă TC
221) şi 4,14% (la linia generatoare de grupă TC 209).
8. În studiul continuţului de amidon şi fibre al liniilor consangvinizate forme
parentale în sistemul dialel cu hibrizi direcţi şi reciproci, nu s-au înregistrat diferenţe
semnificative între cele două situaţii experimentale de polenizare liberă şi
autopolenizare.
9. Conţinut ridicat de amidon la liniile consangvinizate forme parentale în
sistem dialel a fost determinat în situaţia de autopolenizare la liniile consangvinizate
TC 208 (69,60%) şi TC 291 (69,44%), cu diferenţe foarte semnificative faţă de
martor, iar în situaţia de polenizare liberă la liniile TC 291 (72,57%) şi K 1080
(72,23%), cu diferenţe foarte semnificative faţă de martor.
10. În ceea ce priveste continutul de fibre la liniile consangvinizate forme
parentale în sistemul dialel, toate liniile luate în studiu au prezentat valori inferioare
ale conţinutului de fibre faţă de linia martor, cu diferenţe foarte semnificative atât în
cazul boabelor provenite de sub isolator, cât şi în situaţia de polenizare liberă.
11. În determinismul genetic al conţinutului de amidon şi fibre solubile la
porumb într-un sistem genetic de linii consangvinizate isonucleare sunt implicate atât
acţiunile genice nucleare (datorate testerilor), acţiunile genice nucleo-citoplasmatice,
cat şi factorii genetici localizaţi în citoplasme.
12. În transmiterea conţinutului de amidon sunt importante efectele genice
aditive, efectele genice neaditive şi mai puţin importante efectele genice materne.
13. La realizarea formulelor pentru hibrizii bogaţi în amidon este necesar a se
include în combinaţiile hibride forme parentale care să transmită la nivel aditiv
conţinut ridicat de amidon, iar datorită efectelor reciproce a interacţiunii nuclear-
citoplasmatice este absolut obligatorie testarea încrucişărilor directe şi reciproce.

34
14. Pentru obţinerea de hibrizi cu un potenţial ridicat de producere a amidonului
este nevoie de forme parentale bogate în amidon; aceste forme parentale pot fi
obţinute prin selecţie recurentă în germoplasme de porumb, iar apoi, în timpul
consangvinizării prin controlul repetat al conţinutului de amidon. Creşterea
conţinutului de amidon va însemna, implicit, reducerea conţinutului de proteine şi
grăsimi în boabe.
15. În condiţionarea genetică a conţinutului de fibre în boabe provenite de la
stiuleţi autopolenizaţi sunt importante, în egală măsură, efectele genice aditive şi cele
neaditive; în realizarea formulelor de hibridare trebuie să se tină seama de efectele
interacţiunii nuclear-citoplasmatice, efectele materne fiind mai reduse.
16. Recomandăm ca în cazul unor cercetări care necesită mai multă acurateţe
experimentală analizele să se efectueze pe probe provenite din ştiuleţii autopolenizaţi,
indiferent dacă este vorba de hibrizi sau linii consangvinizate.

35
BIBLIOGRAFIE SELECTIVĂ
1. ARDELEAN M., 2006, Principii ale metodologiei cercetării agronomice şi
medical veterinare, Ed. AcademicPres;
2. CEAPOIU N., 1969, Istoricul ameliorării plantelor în România, Probl. genet. teor.
aplic. I: 1-50.
3. DONER L.W., H.K. CHAU, M.L. FISHMAN, K.B. HICKS, 1998, An Improved
Process for Isolation of Corn Fiber Gum, Cereal Chem., 75 (4): 408-411;
4. HAŞ I, CAMELIA CHICINAŞ (RACZ), VOICHIŢA HAŞ, CORNELIA STAN,
TEODORA ŞCHIOP, I. D. COSTE, N. TRITEAN, Rolul diversităţii
citoplasmatice asupra comportării liniilor şi încrucişărilor acestora, 2011
5. HAŞ I., 2004, Heterozisul la porumb, În: Butnaru Gallia, I. Căbulea, M. Cristea,
I. Haş, Voichiţa Haş, Dana Malschi, Felicia Mureşan, Elena Naghy, T. Perju, T.
Sarca, Vasilichia Sarca, D. Scurtu, Porumbul- studiu monografic, Ed. Academiei
Române, Bucureşti;
6. SINGH V., P. YANG, R.A. MOUREAU, K.B. HICKS, S.R. ECKHOFF, 2001,
Effect of harvest moisture content and ambient air drying on mayze fiber oil yield
and its phytosterol composition, în Starch 53, 653-658;
7. ŞCHIOP TEODORA, I. HAŞ, Voichiţa HAŞ, I. COSTE, Camelia RACZ, N.
TRITEAN, 2011, Valoarea fenotipică şi genetică a unor linii consangvinizate
isonucleare de porumb. Studiul fenotipic si genetic al continutului de amidon,
Analele I.C.C.P.T., Fundulea vol. 78;
8. ŞCHIOP TEODORA, I. HAŞ, Voichiţa HAŞ, I. COSTE, Camelia RACZ, N.
TRITEAN, 2011, Gene actions, cytoplasmic actions and cytoplasmic-nuclear
interactions involved in the determination of fiber content in a series of isonuclear
maize lines, Research Journal of Agricultural Science, 43 (II), Timisoara;
9. VÂRBAN D. I., 2008, Culturi de câmp, Ed. Risoprint Cluj-Napoca;
10. *** Anuarul Statistic al României pe anul 2007.
11. www.faostat.fao.org
12. www.dickey-john.com/product/instalab.600/