privind implementarea proiectului pe perioada iulie 2012...
TRANSCRIPT

1
Raport stiintific
privind implementarea proiectului pe perioada iulie 2012-decembrie 2015
Proiectului de cercetare PN-II-ID-PCCE-2011-2-0024, a inceput la data 1/07/2012 cu titlul “Suprarasucirea ADN-ului indusa transcriptional orienteaza selectiv in cis recombinarea V(D)J”avandu-l Director de Proiect pe Dr.Ciubotaru Mihai. Prezentul raport stiintific sintetic intermediar descrie evolutia proiectului pe toata perioada desfasurarii sale iulie2012-decembrie 2015.
Rezumatul proiectului
Recombinarea somatica asambleaza si diversifica genele receptorilor antigenici, ai limfocitelor B si T(1). Pentru initierea recombinarii recombinaza RAG(Recombination Activating Gene proteins 1 &2 ) se leaga de 2 situs-uri complementare de ADN (12 cu 23-RSSs Recombination Signal Sequence), printr-un proces de formare a complexului imperecheat (PC) sau sinapsa. Sinapsa este urmata de clivaj si apoi de repararea fragmentelor dublu catenare, proces care fiziologic se produce numai intre segmente genice localizate intracromozomial. Sinapsa mentine integritatea genomului, prevenind jonctiunile intercromozomiale implicate in fiziopatologia limfoamelor B sau T la copii. Acest proiect are drept scop studierea mecanismelor biochimice si a factorilor ce regleaza sinapsa, contribuind astfel la intelegerea modului in care sinapsarea defectuoasa duce la translocatii. Aproape 50% din cancerele la copii apar in celule limfoide si aproximativ 40% din aceste boli maligne se datoreaza translocatiilor aparute in urma recombinarii defectuoase. Ne propunem sa testam modul in care suprarasucirea ADN-ului stimuleaza formarea PC-ului si vom identifica conformatia topologica optima a RSS-urilor in sinapsa. Vom determina cum transcrierea ce afecteaza local suprarasucirea ADN-ului, directioneaza recombinarea in cis. Folosind ADN circular cu topologie cunoscuta vom studia mecanismul prin care RAG discerne intre situs-uri de ADN distinct suprarasucite. Rezultatele acestor studii isi gasesc aplicabilitatea in design-ul si folosirea compusilor metaloelicoidali in profilaxia bolilor mentionate.
OBJECTIVUL D2A (identificat in propunerea de proiect la pg. 25 cu subobiectivele D3A1 si D3A2 )
D3A.Identificarea mecanismului prin care suprarasucirea ADN-ului activeaza cataliza recombinazei
RAG
D3A Purificarea proteineinelor RAG1, RAG2 si HMGB1.
Majoritatea studiilor in vitro au folosit domeniile esentiale "core" ale RAG1 (secventa de aminoacizi 384-1008 din totalul de 1040aa)(2) si RAG2 murin (regiunea 1-383 din 527aa), (3) care sunt mult mai solubile decat proteinele intregi si mai lesne de purificat. RAG1 core interactioneaza direct cu elementele nonamer, si heptamer ale RSSs si joaca un rol central in legarea ADN-ului (4). Deasemenea, RAG1 interactioneaza cu RAG2 si cu el insusi dimerizand si toate resturile aminoacidice cu rol catalitic cunoscute pana in prezent sunt localizate in structura sa (D600, D708, E962). Mult mai putin se stie despre proteina RAG2 core. Desi este esentiala pentru sinapsa si cataliza, studiile de mutageneza intreprinse pana in prezent nu au identificat inca pozitii critice de aminoacizi pentru nici una din aceste functii (4,5). RAG mediaza formarea complexului sinaptic, astfel controland atat selectivitatea (segmentele genice alese pentru recombinare), cit si cea mai mare parte a diversitatii raspunsului imun dobandit (conferita de continutul segmentelor genice unite). Desi RAG leaga eficient RSS-uri in mod individual formand complexul unic (single complexes (SC)), pentru sinapsa are nevoie aditional de prezenta proteinelor HMGB1 sau 2 (high mobility group proteins)(5). Acestea sunt proteine ce indoaie nespecific ADN-ul avand un rol arhitectural in modelarea
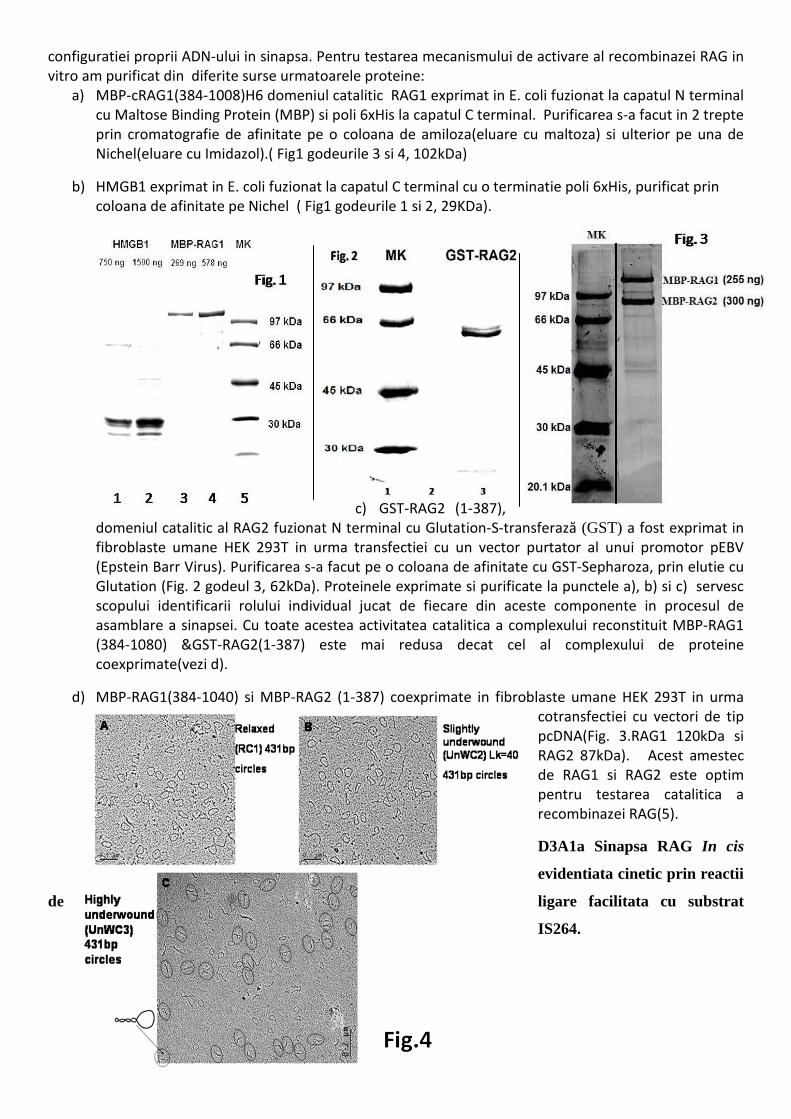
2
configuratiei proprii ADN-ului in sinapsa. Pentru testarea mecanismului de activare al recombinazei RAG in vitro am purificat din diferite surse urmatoarele proteine:
a) MBP-cRAG1(384-1008)H6 domeniul catalitic RAG1 exprimat in E. coli fuzionat la capatul N terminal cu Maltose Binding Protein (MBP) si poli 6xHis la capatul C terminal. Purificarea s-a facut in 2 trepte prin cromatografie de afinitate pe o coloana de amiloza(eluare cu maltoza) si ulterior pe una de Nichel(eluare cu Imidazol).( Fig1 godeurile 3 si 4, 102kDa)
b) HMGB1 exprimat in E. coli fuzionat la capatul C terminal cu o terminatie poli 6xHis, purificat prin coloana de afinitate pe Nichel ( Fig1 godeurile 1 si 2, 29KDa).
c) GST-RAG2 (1-387), domeniul catalitic al RAG2 fuzionat N terminal cu Glutation-S-transferază (GST) a fost exprimat in fibroblaste umane HEK 293T in urma transfectiei cu un vector purtator al unui promotor pEBV (Epstein Barr Virus). Purificarea s-a facut pe o coloana de afinitate cu GST-Sepharoza, prin elutie cu Glutation (Fig. 2 godeul 3, 62kDa). Proteinele exprimate si purificate la punctele a), b) si c) servesc scopului identificarii rolului individual jucat de fiecare din aceste componente in procesul de asamblare a sinapsei. Cu toate acestea activitatea catalitica a complexului reconstituit MBP-RAG1 (384-1080) &GST-RAG2(1-387) este mai redusa decat cel al complexului de proteine coexprimate(vezi d).
d) MBP-RAG1(384-1040) si MBP-RAG2 (1-387) coexprimate in fibroblaste umane HEK 293T in urma cotransfectiei cu vectori de tip pcDNA(Fig. 3.RAG1 120kDa si RAG2 87kDa). Acest amestec de RAG1 si RAG2 este optim pentru testarea catalitica a recombinazei RAG(5).
D3A1a Sinapsa RAG In cis
evidentiata cinetic prin reactii
de ligare facilitata cu substrat
IS264.
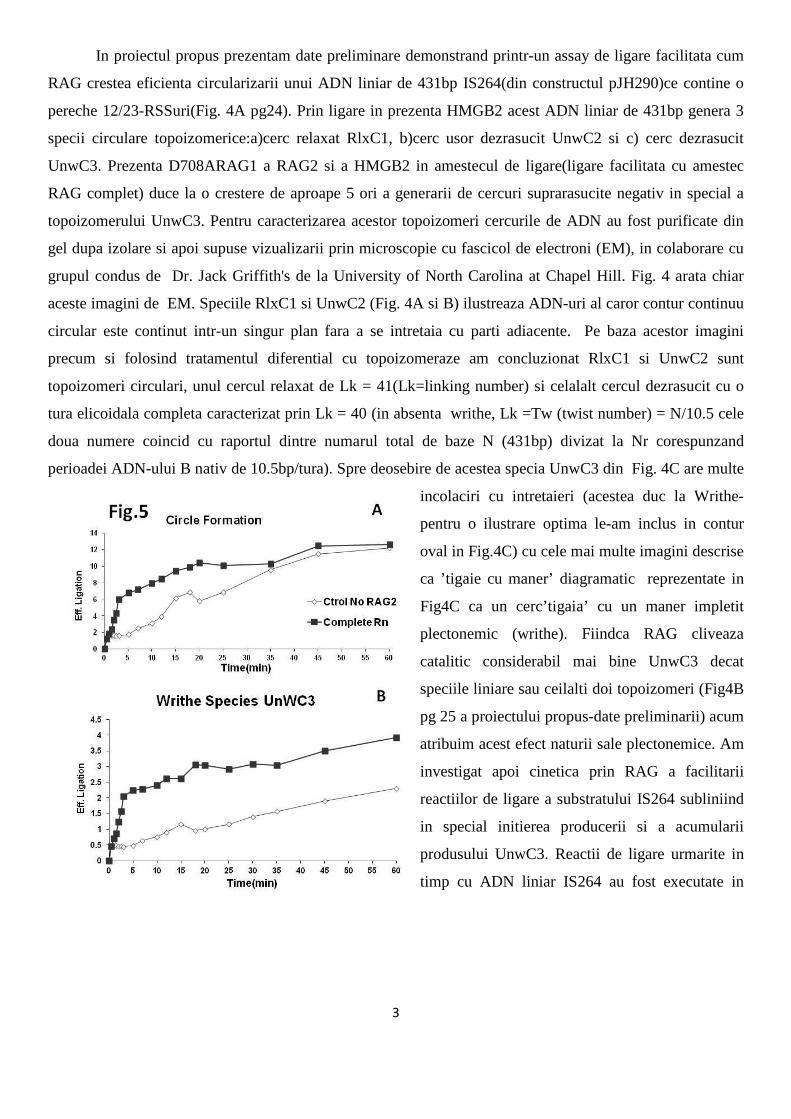
3
In proiectul propus prezentam date preliminare demonstrand printr-un assay de ligare facilitata cum
RAG crestea eficienta circularizarii unui ADN liniar de 431bp IS264(din constructul pJH290)ce contine o
pereche 12/23-RSSuri(Fig. 4A pg24). Prin ligare in prezenta HMGB2 acest ADN liniar de 431bp genera 3
specii circulare topoizomerice:a)cerc relaxat RlxC1, b)cerc usor dezrasucit UnwC2 si c) cerc dezrasucit
UnwC3. Prezenta D708ARAG1 a RAG2 si a HMGB2 in amestecul de ligare(ligare facilitata cu amestec
RAG complet) duce la o crestere de aproape 5 ori a generarii de cercuri suprarasucite negativ in special a
topoizomerului UnwC3. Pentru caracterizarea acestor topoizomeri cercurile de ADN au fost purificate din
gel dupa izolare si apoi supuse vizualizarii prin microscopie cu fascicol de electroni (EM), in colaborare cu
grupul condus de Dr. Jack Griffith's de la University of North Carolina at Chapel Hill. Fig. 4 arata chiar
aceste imagini de EM. Speciile RlxC1 si UnwC2 (Fig. 4A si B) ilustreaza ADN-uri al caror contur continuu
circular este continut intr-un singur plan fara a se intretaia cu parti adiacente. Pe baza acestor imagini
precum si folosind tratamentul diferential cu topoizomeraze am concluzionat RlxC1 si UnwC2 sunt
topoizomeri circulari, unul cercul relaxat de Lk = 41(Lk=linking number) si celalalt cercul dezrasucit cu o
tura elicoidala completa caracterizat prin Lk = 40 (in absenta writhe, Lk =Tw (twist number) = N/10.5 cele
doua numere coincid cu raportul dintre numarul total de baze N (431bp) divizat la Nr corespunzand
perioadei ADN-ului B nativ de 10.5bp/tura). Spre deosebire de acestea specia UnwC3 din Fig. 4C are multe
incolaciri cu intretaieri (acestea duc la Writhe-
pentru o ilustrare optima le-am inclus in contur
oval in Fig.4C) cu cele mai multe imagini descrise
ca ’tigaie cu maner’ diagramatic reprezentate in
Fig4C ca un cerc’tigaia’ cu un maner impletit
plectonemic (writhe). Fiindca RAG cliveaza
catalitic considerabil mai bine UnwC3 decat
speciile liniare sau ceilalti doi topoizomeri (Fig4B
pg 25 a proiectului propus-date preliminarii) acum
atribuim acest efect naturii sale plectonemice. Am
investigat apoi cinetica prin RAG a facilitarii
reactiilor de ligare a substratului IS264 subliniind
in special initierea producerii si a acumularii
produsului UnwC3. Reactii de ligare urmarite in
timp cu ADN liniar IS264 au fost executate in

4
paralel folosind fie un amestec RAG complet (HMGB2∆C, MBPcR1D708A si GST-RAG2), fie cu
amestecuri control executate in conditii similare dar in absenta GST-RAG2(no RAG2 control). Amestecurile
de reactie au fost separate prin migrare pe geluri native PAGE (similar cu reactiile descrise in Fig. 4A pg 24
a propunerii de proiect) si speciile produsi de reactie marcate radioactiv cuantificate prin PhosphorImager.
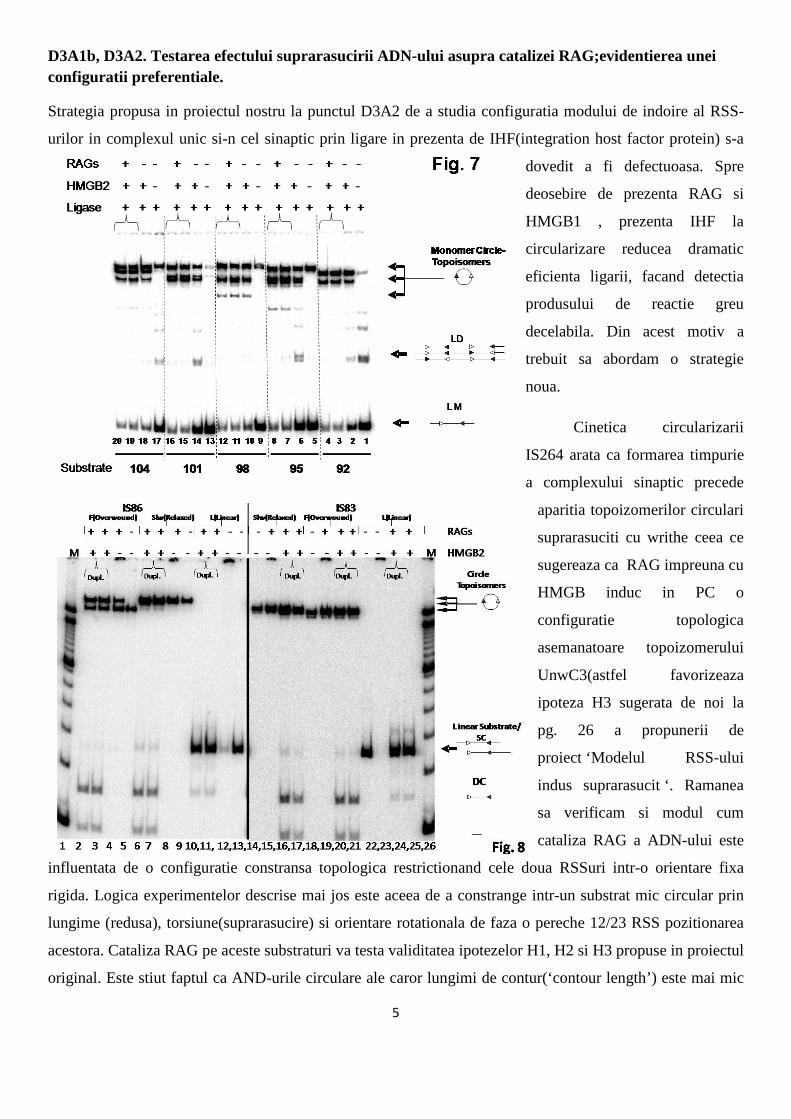
Fig. 5A ilustreaza Eficienta Ligarii (Lig. Eff. = Lig. Product/Total linear DNA substrate) in timp cumulativa
a tuturor cercurilor formate in 60 mins., in timp ce Fig. 5B, ilustreaza eficienta de producere prin ligare a
UnwC3. Curbele cu patrate negre desemneaza reactiile cu amestec RAG total iar cele ale reactiilor control
sunt ilustrate cu romburi deschise (no
RAG2). RAG creste considerabil rata si
eficienta de generare a cercurilor in primele 5
minute ale reactiei si efectul satureaza in
platou in urmatoarele 20 minute. Efectele
descrise capata aspectul cel mai dramatic
pentru UnwC3 a carei generare depinde de
prezenta RAG complet chiar si dupa 60 min.
de ligare. Aceste rezultate sugereaza fara
echivoc ca RAG nu se leaga in mod pasiv de
ADN-ul suprarasucit cu writhe numai
stabilizand formarea acestor specii in cursul ligarii (caz in care UnwC3 ar fi trebuit sa fie in mod similar
generat in reactiile cu amestec RAG complet precum si-n cele de control macar la inceputul reactiei),ci
dimpotriva arata ca RAG induce activ writhe. Pe langa efectul de imperechere a secventelor consens 12/23-
RSSs, sinapsa prin RAG induce impletirea ADN-ului ce flancheaza RSS-urile. Acest rezultat a fost
neasteptat pentru noi. In concluzie impletirea negativa a ADN-ului este facilitata prin sinapsa RAG in cazul
substratului cu orientare deletionala iar acest tip de substrat cu suprarasucire negativa plectonemica este
folosit catalitic preferential de RAG. Asa cum aratam in Fig. 4A a propunerii noastre de proiect (pg. 24),
ligarea constructului IS264 genereaza pe langa produsii monomerici circulari dimeri si trimeri liniari produsi
care presupun sinapsa in trans prin RAG. In experimentul de cinetica pe constructul IS264 am cuantificat si
cantitatile infime de multimeri geerate prin sinapsa in trans de RAG rezultatele fiind prezentate in Fig. 6. In
primul rand notam eficientele reduse de ligare ilustrate pentru acesti produsi (nivelele de ligare in reactiile
cu amestec RAG sunt de aproape douazeci de ori mai reduse la saturatie decat cele de formare de cercuri
monomerice si de cinci ori mai reduse decat cele de formare a topoizomerului UnwC3). Totusi, foarte
timpuriu ligarea este facilitata prin sinapsa RAG in trans dar in mod remarcabil dupa 13 minute prezenta in
amestecul de ligare a RAG-HMGB are un efect inhibitor, foarte probabil datorat folosirii substratului indus
suprarasucit prevalent in reactiile de ciclizare monomerice.
5
D3A1b, D3A2. Testarea efectului suprarasucirii ADN-ului asupra catalizei RAG;evidentierea unei configuratii preferentiale.
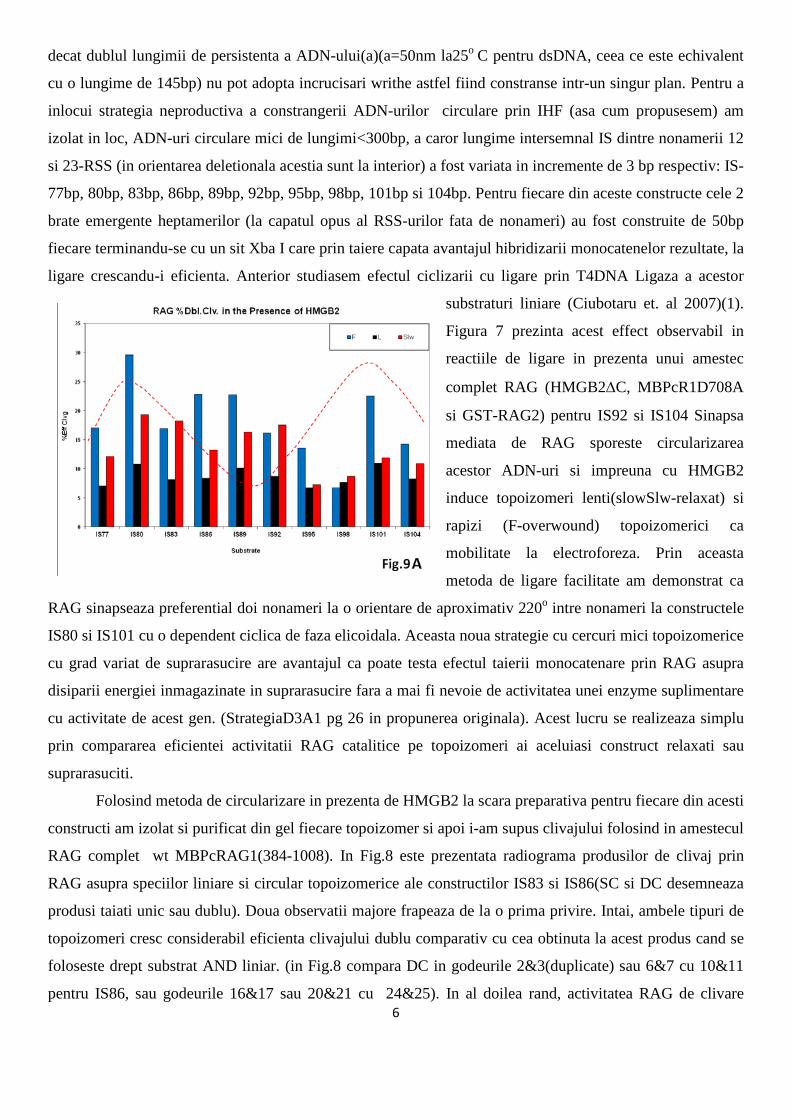
Strategia propusa in proiectul nostru la punctul D3A2 de a studia configuratia modului de indoire al RSS-
urilor in complexul unic si-n cel sinaptic prin ligare in prezenta de IHF(integration host factor protein) s-a
dovedit a fi defectuoasa. Spre
deosebire de prezenta RAG si
HMGB1 , prezenta IHF la
circularizare reducea dramatic
eficienta ligarii, facand detectia
produsului de reactie greu
decelabila. Din acest motiv a
trebuit sa abordam o strategie
noua.
Cinetica circularizarii
IS264 arata ca formarea timpurie
a complexului sinaptic precede
aparitia topoizomerilor circulari
suprarasuciti cu writhe ceea ce
sugereaza ca RAG impreuna cu
HMGB induc in PC o
configuratie topologica
asemanatoare topoizomerului
UnwC3(astfel favorizeaza
ipoteza H3 sugerata de noi la
pg. 26 a propunerii de
proiect ‘Modelul RSS-ului
indus suprarasucit ‘. Ramanea
sa verificam si modul cum
cataliza RAG a ADN-ului este
influentata de o configuratie constransa topologica restrictionand cele doua RSSuri intr-o orientare fixa
rigida. Logica experimentelor descrise mai jos este aceea de a constrange intr-un substrat mic circular prin
lungime (redusa), torsiune(suprarasucire) si orientare rotationala de faza o pereche 12/23 RSS pozitionarea
acestora. Cataliza RAG pe aceste substraturi va testa validitatea ipotezelor H1, H2 si H3 propuse in proiectul
original. Este stiut faptul ca AND-urile circulare ale caror lungimi de contur(‘contour length’) este mai mic
6
decat dublul lungimii de persistenta a ADN-ului(a)(a=50nm la25o C pentru dsDNA, ceea ce este echivalent
cu o lungime de 145bp) nu pot adopta incrucisari writhe astfel fiind constranse intr-un singur plan. Pentru a
inlocui strategia neproductiva a constrangerii ADN-urilor circulare prin IHF (asa cum propusesem) am
izolat in loc, ADN-uri circulare mici de lungimi<300bp, a caror lungime intersemnal IS dintre nonamerii 12
si 23-RSS (in orientarea deletionala acestia sunt la interior) a fost variata in incremente de 3 bp respectiv: IS-
77bp, 80bp, 83bp, 86bp, 89bp, 92bp, 95bp, 98bp, 101bp si 104bp. Pentru fiecare din aceste constructe cele 2
brate emergente heptamerilor (la capatul opus al RSS-urilor fata de nonameri) au fost construite de 50bp
fiecare terminandu-se cu un sit Xba I care prin taiere capata avantajul hibridizarii monocatenelor rezultate, la
ligare crescandu-i eficienta. Anterior studiasem efectul ciclizarii cu ligare prin T4DNA Ligaza a acestor
substraturi liniare (Ciubotaru et. al 2007)(1).
Figura 7 prezinta acest effect observabil in
reactiile de ligare in prezenta unui amestec
complet RAG (HMGB2∆C, MBPcR1D708A
si GST-RAG2) pentru IS92 si IS104 Sinapsa
mediata de RAG sporeste circularizarea
acestor ADN-uri si impreuna cu HMGB2
induce topoizomeri lenti(slowSlw-relaxat) si
rapizi (F-overwound) topoizomerici ca
mobilitate la electroforeza. Prin aceasta
metoda de ligare facilitate am demonstrat ca
RAG sinapseaza preferential doi nonameri la o orientare de aproximativ 220o intre nonameri la constructele
IS80 si IS101 cu o dependent ciclica de faza elicoidala. Aceasta noua strategie cu cercuri mici topoizomerice
cu grad variat de suprarasucire are avantajul ca poate testa efectul taierii monocatenare prin RAG asupra
disiparii energiei inmagazinate in suprarasucire fara a mai fi nevoie de activitatea unei enzyme suplimentare
cu activitate de acest gen. (StrategiaD3A1 pg 26 in propunerea originala). Acest lucru se realizeaza simplu
prin compararea eficientei activitatii RAG catalitice pe topoizomeri ai aceluiasi construct relaxati sau
suprarasuciti.
Folosind metoda de circularizare in prezenta de HMGB2 la scara preparativa pentru fiecare din acesti
constructi am izolat si purificat din gel fiecare topoizomer si apoi i-am supus clivajului folosind in amestecul
RAG complet wt MBPcRAG1(384-1008). In Fig.8 este prezentata radiograma produsilor de clivaj prin
RAG asupra speciilor liniare si circular topoizomerice ale constructilor IS83 si IS86(SC si DC desemneaza
produsi taiati unic sau dublu). Doua observatii majore frapeaza de la o prima privire. Intai, ambele tipuri de
topoizomeri cresc considerabil eficienta clivajului dublu comparativ cu cea obtinuta la acest produs cand se
foloseste drept substrat AND liniar. (in Fig.8 compara DC in godeurile 2&3(duplicate) sau 6&7 cu 10&11
pentru IS86, sau godeurile 16&17 sau 20&21 cu 24&25). In al doilea rand, activitatea RAG de clivare

7
monocatenara intrinseca converteste topoizomerii F suprarasuciti migrand rapid in topoizomeri Slw chiar in
absenta sinapsei(vezi interconversia din godeurile 4 si 19 cu reactiile fara HMGB2). Reactii similare cu cele
prezentate in Fig. 8 s-au obtinut in alte trei experimente individuale pentru fiecare tip de substrat ADN si
acelasi tip de observatii s-au facut pentru toti cei zece constructi. Inainte insa de a discuta rezultatele de
dublu clivaj cumulativ obtinute pentru toti cei zece constructi, vom incerca sa revizuim cele trei ipoteze (H1-
3) ale proiectului propus initial (pg.26) si vom trece in revista ipotezele de lucru atunci formulate asupra
acestor experimente. H1 " modelul ADN-ului tip resort elastic” presupune ca intreaga energie de
suprarasucire este convertita in energie mecanica necesara desfacerii ADN-ului la treapta a IIa de formare a
acului de par. Daca aceasta ipoteza ar fi adevarata deoarece taierea monocatenara indeparteaza complet
energia torsionala de suprarasucire la treapta premergatoare formarii acului de par, H1 prezice ca n-ar trebui
sa existe nici o diferenta intre eficientele de dublu clivaj obtinute la topoizomerul Slw fata de cel F al
aceluiasi construct (cum am aratat anterior clivajul monocatenar ii interconverteste inainte de taierea dublu
catenara). H2 "Modelul complexului proteina-ADN elastic” propune conversia intregii energii de
suprarasucire intr-o reconfigurare proteica in timpul tranzitiei de la SC la PC, chiar inaintea formarii acului
de par. Asadar H2 prezice eficiente de clivaj mai crescute in mod consistent la toate reactiile folosind
substrat F fata de cele folosind topoizomeri Slw pentru acelasi construct, in toate cele zece cazuri. H3
"Modelul RSS-urilor configurate prin suprarasucire " potrivit careia RAG necesita la cataliza o anume
configuratie a ADN-ului RSS mult mai usor de indus in complexul cu RAG-HMGB daca ADN-ul porneste
de la starea suprarasucita decat de la cea relaxata(bariera energiei de activare mai redusa). Orientarea fazei
elicoidale optime a perechilor 12/23-RSS determinata in assay-ul de circularizare prin legare ar trebui sa
favorizeze si clivajul cuplat prin RAG. Histograma din Fig. 9 ilustreaza rezultatele cumulative de clivaj
dublu obtinute cu amestec complet activ RAG pentru toti constructii; cu negru pentru substratul liniar, cu
rosu Slw- cercuri relaxate si in albastru F-topoizomeri suprarasuciti. Cu rosu linia punctata denota aspectul
rezultat din eficientele de legare ale acestor constructi/(formarea sinapsei) anterior publicate (Ciubotaru et.
al. 2007)(1). Notabil toate substratele sub forma de ADN liniar sunt clivate cu eficienta scazuta similara de
5-8% la toti constructii. In mod neasteptat insa substratele circulare cuprinse intre IS77 si IS92 sunt in
general mai bine dublu clivate de RAG decat constructii cu distante mai mari inter-RSS(de la IS95 la
IS104). In mod considerabil pentru toti constructii, topoizomerii circulari sunt sensibil mai bine clivate decat
variantele lor liniare. Pentru trei dintre constructi RAG nu discerne intre variantele lor de topoizomeri F sau
Slw (IS83, IS92, IS98), dar in cazul celorlalti 7 constructi topoizomerii F sunt substrate mai bune pentru
cataliza RAG. Aceasta observatie invalideaza ambele ipoteze H1 si H2 care asa cum am afirmat anterior fie
presupun echivalenta la clivaj a tuturor topoizomerilor (H1) sau presupun preferinta clar catalitica a
topoizomerilor suprarasuciti F fata de cei Slw (H2). Un alt argument pledand serios impotriva mecanismelor
propuse de ipotezele H1 si H2 este gradul mare de variabilitate observat la eficienta de clivaj intre diferitii
constructi (H1 si H2 prezic eficienta asemanatoare intre aceleasi forme ale diferitilor constructi). Desi
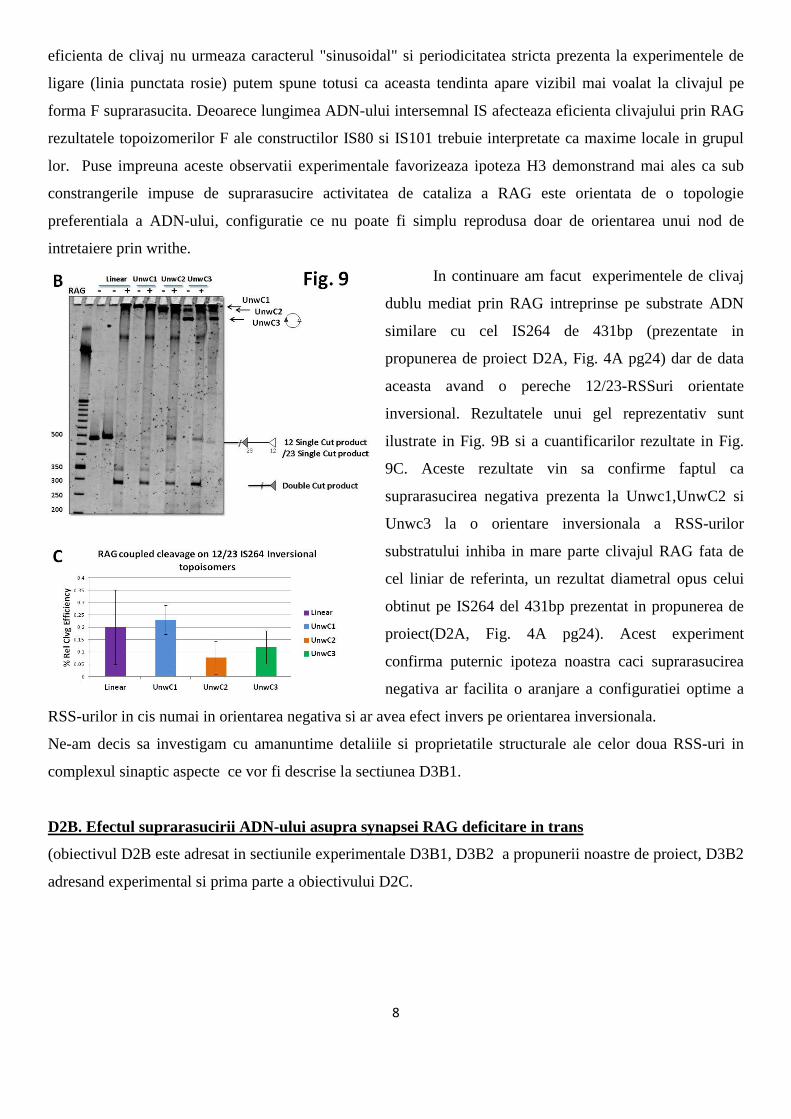
8
eficienta de clivaj nu urmeaza caracterul "sinusoidal" si periodicitatea stricta prezenta la experimentele de
ligare (linia punctata rosie) putem spune totusi ca aceasta tendinta apare vizibil mai voalat la clivajul pe
forma F suprarasucita. Deoarece lungimea ADN-ului intersemnal IS afecteaza eficienta clivajului prin RAG
rezultatele topoizomerilor F ale constructilor IS80 si IS101 trebuie interpretate ca maxime locale in grupul
lor. Puse impreuna aceste observatii experimentale favorizeaza ipoteza H3 demonstrand mai ales ca sub
constrangerile impuse de suprarasucire activitatea de cataliza a RAG este orientata de o topologie
preferentiala a ADN-ului, configuratie ce nu poate fi simplu reprodusa doar de orientarea unui nod de
intretaiere prin writhe.
In continuare am facut experimentele de clivaj
dublu mediat prin RAG intreprinse pe substrate ADN
similare cu cel IS264 de 431bp (prezentate in
propunerea de proiect D2A, Fig. 4A pg24) dar de data
aceasta avand o pereche 12/23-RSSuri orientate
inversional. Rezultatele unui gel reprezentativ sunt
ilustrate in Fig. 9B si a cuantificarilor rezultate in Fig.
9C. Aceste rezultate vin sa confirme faptul ca
suprarasucirea negativa prezenta la Unwc1,UnwC2 si
Unwc3 la o orientare inversionala a RSS-urilor
substratului inhiba in mare parte clivajul RAG fata de
cel liniar de referinta, un rezultat diametral opus celui
obtinut pe IS264 del 431bp prezentat in propunerea de
proiect(D2A, Fig. 4A pg24). Acest experiment
confirma puternic ipoteza noastra caci suprarasucirea
negativa ar facilita o aranjare a configuratiei optime a
RSS-urilor in cis numai in orientarea negativa si ar avea efect invers pe orientarea inversionala.
Ne-am decis sa investigam cu amanuntime detaliile si proprietatile structurale ale celor doua RSS-uri in
complexul sinaptic aspecte ce vor fi descrise la sectiunea D3B1.
D2B. Efectul suprarasucirii ADN-ului asupra synapsei RAG deficitare in trans
(obiectivul D2B este adresat in sectiunile experimentale D3B1, D3B2 a propunerii noastre de proiect, D3B2
adresand experimental si prima parte a obiectivului D2C.

9
Obiectiv 2 D3B1. Configuratia 23 si 12-RSS in complexul sinaptic. Pentru a investiga sinapsa in cis
versus trans precum si pentru a orienta design-ul modului cum metalohelicatii se leaga de ADN-ul
suprarasucit trebuie sa intelegem pe deplin detaliile topologice si structurale ale dispunerii celor doua RSS-
uri in complexul sinaptic. Avand in vedere faptul ca un nod cu
impletire writhe simplu nu poate reproduce configuratia celor
doua RSS-uri in PC ne-am decis sa folosim metodologia FRET
pentru adezvalui in detaliu aspectul elementelor ADN 12 si 23-
RSS in acest complex. Experimentele de FRET ce urmeaza in
descriere au fost desfasurate in colaborare cu grupurile Dr.
David G. Schatz si Dr. Elizabeth Rhoades's de la Yale
University iar asamblarea modelelor structurale 3D a fost
desavarsita in colaborare cu grupul domnului Dr. Andrei J.
Petrescu de la Institutul de Biochimie al Academiei Romane
din Bucuresti. Aceasta metodologie avea sa inlocuiasca cu
succes strategia propusa in proiectul initial cu topoizomeri
indusi prin IHF (D3A2 pg. 27) neproductiva si pe baza ei am
reusit sa descifram configuratia celor 2 RSSuri in PC. Pentru
a determina gradul de indoire al RSS-urilor am folosit
oligonucleotide dublu catenare fiecare individual marcate cu
o pereche de fluorofori Acceptor/Donor (A/D) localizati in
cis (marcare duala), necesari evidentierii fenomenului de
FRET (Fluorescence resonance energy transfer)(Fig. 10A si
B). Fiecare fluorofor a fost cuplat in cursul sintezei de ADN
prin intermediul unei punti amino modificate continand sase grupari de metilen C6 pentru a reduce la minim
posibilele interactiuni proteine-fluorofori din complexul cu RAG. Design-ul oligonucleotidelor 12 si 23-RSS
foloseste ca principiu orientarea fluoroforilor in raport cu directia indoiturilor induse de RAG (Fig. 10).
Studiul nostru compara eficienta fenomenului FRET a aceleiasi perechi. Acceptor/Donor in absenta fata de
eficienta detectata in prezenta proteinelor necesare formarii sinapsei (amestec RAG complet MBP-
cRAG1wt, GST-cRAG2 HMGB1 si un partener RSS nemarcat). Schimbarile de eficienta a fenomenului
FRET induse de fiecare tip de complex in parte detecteaza o schimbare a pozitionarii perechii A/D in
contextul creat de legarea RSS-ului. Principiul experimental este schematizat in desenele din Fig. 10C. In
cazul unor indoituri ascutite a caror convexitate este pozitionata intre fluoroforii A/D distanta dintre
fluoroforii perechi localizati pe aceeasi fata a elicei creste (scazand eficienta fenomenului FRET). Cand
indoiturile din RSS-uri sunt pozitionate cu concavitatea lor intre perechile de A/D, aceasta duce la o scadere

10
drastica a distantei dintre fluoroforii localizati pe aceeiasi fata a elicei (corespunde unei cresteri a eficientei
FRET).
Desi am evidentiat indoirea ADN-ului RSS folosind multe din substratele descrise in Fig 10A, substratele 12
si 23RSSdR2a (Fig.11-Spectre de Emisie Fluorescenta pt 23RSSdR2a) au fost cele mai informative si de
aceea au fost studiate in amanuntime. Experimentele descrise mai jos pentru 23RSSdR2a au fost intreprinse
si pentru 12RSSdR2a si fiindca rezultatele obtinute sunt similare la ambele elemente ADN prezentam in
acest raport doar varianta celor de la 23-RSS. In aceste substrate donorul este pozitionat 3bp 3' fata de
nonamer(cercul galben) si acceptorul
la 9bp 5' fata de heptamer(cutia
albastra) fiind sunt separate in cazul
23-RSS de 51bp(distanta 171-180Å)
iar in 12-RSS de 40bp(distanta 131-
140Å ). Cum transferul de energie
nu este detectabil la distante mai mari
90Å, nu ne-am asteptat sa obtinem
FRET in absenta proteinelor cand ADN-ul este liber. Acest lucru a fost confirmat experimental(in raport
ilustrat doar pentru 23-RSS in Fig. 11A, spectrele de emisie pentru 23RSSdR2a (linia rosie; indicata
"(d+a)") este similara cu reactia control in care donorul si acceptorul sunt pozitionate in trans pe substrate
23RSS distincte (un amestec echimolar de 23RSSdR si 23RSS2a; curbele negre continue; "(d) + (a)"). Aceste
spectre se caracterizeaza prin maxima de emisie la ~ 520 nm, caracteristic donorului FAM, fara a evidentia
emisia acceptorului sau diminuarea emisiei donorului in situatia in care fluoroforii sunt pozitionati in cis sau
in trans pe ADN. Spectrele din Fig11 au fost corectate pentru fluorescenta reziduala a
acceptorului.Adaugarea la 23RSSdR (contine numai donor-) a amestecului complet RAG proteic si a 12-RSS
duce la o reducere substantiala a
fluorescentei donorului (linia
neagra punctata; "(d) + proteins")
(Fig. 11A), datorata interactiunilor
locale dintre donorul FAM si
proteine. Cand sustratul dublu
marcat 23RSSdR2a este incubat cu
amestec complet RAG (curba
albastra; "(d+a) + Proteins"), observam o scadere suplimentara a fluorescentei donorului precum si
sensitizarea acceptorului (max. emisie intre 570-595 nm) (Fig. 11A, compara spectrul trasat in albastru cu
cel in negru punctat). Aceste modificari spectrale ce trebuiesc atribuite sensitizarii acceptorului, indica

11
transferul de energie intre fluorofori indus de complexul protein-ADN. Acest fenomen a fost consistent
reprodus in 17 experimente independente din a caror cuantificare s-a obtinut o eficienta FRET medie (E-
FRET) de 18.0% ± 1.2%. Acest lucru indica faptul ca in complexul PC RAG-DNA donorul si acceptorul
sunt separate de o distanta mai mica de 90Å. Folosind teoria FRET radiativa Forster am calculate distanta
dintre fluorofori in sinapsa pe 23RSSdR2a de 71Å ± 10Å. Acest lucru este posibil numai daca ADN-ul este
supus unei distorsiuni/indoiri majore in regiunea cuprinsa intre cei doi fluorofori.Pentru a descrie o harta cit
mai exacta a tuturor indoiturilor aparute la nivelul secventelor 12 sau 23-RSS, am studiat un nr de 14 perechi
de fluorofori in cis pentru 23-RSS si 16 perechi cis pentru 12-RSS (Fig.10A si B). Distantele interfluorofori
masurate in complexul synaptic si in signal complex SC (doar pentru 12-RSS) au fost folosite pentru
construirea modelelor structural moleculare a 23 si respectiv 12-RSS in PC si SC. Experimentele descriind
masuratorile FRET efectuate pentru 23-RSS in PC iar 12-RSS in SC si PC precum si modelele rezultate au
fost descrise in amanunt si publicate in ianuarie anul curent Ciubotaru et. al. 2013 si 2015 in revista Nucleic
Acids Research Oxford University Press (3,8).
Masuratorile si modelele structurale privind 12-RSS au fost submise spre publicatie in luna noiembrie anul
in curs, la aceeiasi revista stiintifica
fiind in prezent in stadiul de revizie
stiintifica. Fig. 12 prezinta
comparativ structurile moleculare
rezultate din masuratorile FRET
pentru 23-RSS si 12RSS in PC si
cu sageti situs-urile de maxima
hipersensibilitate la DNAazaI. Doua
concluzii majore se desprind din
aceste modele structurale. Atat 12
cat si 23-RSS sunt indoite de RAG
si HMGB1 in mod considerabil,
ADN-ul lor adoptand forma literei
U(23) sau V(12) intoarse cu
heptamerul si nonamerul fiecareia
(colorate pe ambele structuri)
localizate pe bratele literelor model
la jonctiunea cu regiunea
intermediara ce le separa si care
preia arcuirea maxima. La ambele
elemente ADN regiunile de contact
cu componentele proteice sunt

12
localizate la nivelul concavitatii curburii. Acest aspect este de maxima importanta pentru noi caci putem
orienta acum modul cum RSS- urile suprarasucite vin in contact cu complexul RAG-HMGB in PC. Singura
structura de inalta rezolutie disponibila a unui domeniu al RAG1 in complex cu elementul consens nonamer
(descris de F. F. Yin, din grupul Dr. Schatz's in colaborare cu cel al Dr. Tom Steitz's de la Yale
University)(2) este cea a peptidului de 75 aminoacizi Nonamer Binding Domain NBD of RAG1 (389-456)(F.
F. Yin et. al. 2009). Pentru a completa aceasta structura cu detaliile relationale dintre cele doua structuri
consens ale 12/23-RSS si RAG in interiorul PC am ancorat "in silico" cei doi nonameri proveniti de la doua
23 RSS-uri indoite din PC rezultate din FRET la sirul de interactiuni Aminoacid-baze descrise la motivul de
legare GGRPR din dimerul NBD. Deoarece structura 12-RSS este mult prea compacta pentru a permite
acomodarea in trans a NBD, modelarea s-a facut cu doua structuri 23RSS. Structura rezultata din “targeted
docking” a fost ulterior supusa modelarii molecular dinamice, cu minimizarea eventualelor conflicte sterice
si de potential electrostatic. Rezultatul este ilustrat in Fig.13 fiecare 23-RSS colorat distinct(unul rosu si
unul verde), literele N desemnand fiecare din nonameri iar cu H heptamerii celor 2 RSS-uri. Acest model a
fost descris de noi in articolul review teoretic publicat in acest an in revista Discoveries Ciubotaru et. al
2014(7). Desi structura model rezultata in Fig.13 provine din ancorarea unui domeniu restrains (NBD) al
RAG1 si cu siguranta prezinta o viziune mult simplificata a complexului RAG synaptic (PC), totusi ea ne
ofera cateva repere structurale importante ce ne ajuta sa interpretam rezultatele cinetice obtinute la ligarea
IS264 precum si cele de clivaj pe ADN-urile circulare topoizomerice prezentate la paginile 3-4 si 7 ale
acestui raport. Deoarece miezul complexului proteic RAG-HMGB pare sa fie chiar si partial inchis inlauntrul
plectonemelor (arcelor impletite) celor 2 RSS-uri acest lucru exclude contactul direct dintre regiunile lor de
ADN “spacer”, asa cum s-ar fi intamplat daca RAG ar fi legat un nod pozitiv sau negativ preformat ADN
impletit cu “writhe”. Asadar RAG ar lega preferential cu
afinitate sporita fiecare din cele doua RSS-uri in stare
suprarasucita formand 12 sau 23SC mult mai degraba decat
RSSuri in stare de ADN relaxat deoarece suprarasucirea
prefigureaza o stare de indoire plectonemica(mult mai
apropiata de starea indusa RSS-ului de RAG in PC). Totusi
formarea PC nu este in mod pasiv facilitat de simpla
juxtapunere a celor doua RSS-uri intr-un singur nod
suprarasucit preformat( asa cum propuneam in mod gresit in
ipoteza noastra de lucru prezentata in propunerea de proiect
la pg. 22 si 25). Dimpotriva, sinapsa este mediata de
interactiuni proteina-proteina care ancoreaza doua RSS-uri
suprarasucite complementare in sinaptozom. Tocmai
aceasta orientare optima impreuna cu adiacenta de orientare a interactiunilor protein-proteina (dupa
formarea SC) ceea ce faciliteaza sinapsarea preferentiala si optimizeaza cataliza constructelor IS80 si
IS101. Deoarece cele doua RSS-uri indoite in PC adopta structuri considerabil distincte (Fig.12) modelul
din Fig.13 propune o intrebare noua, si anume cea referitoare la modul cum sinaptozomul se formeaza

13
izotropic sau anisotropic(necesita o orientare unica a celor doua elemente 12 deasupra RAG la mijloc 23
subiacent sau vice versa)? In continuare am decis sa abordam acest tip de intrebari folosind fenomenul de
transcriere ca un factor regulator al semnului suprarasucirii (vezi sectiunea D2C-D3C).
Obiectiv3-D2C, D3C. Suprarasucirea ADN-ului indusa prin transcriere factor selectiv in favorizarea formarii PC in cis.
(objectivul D2C este addresat in sectiunea experimentala a propunerii de proiect subsectiunea D3C la pg
29). Ipoteza de lucru a proiectului propunea ca transcrierea initiata in preajma de la promotorilor din
proximitatea segmentelor V si D incorporeaza RSSurile lor in suprarasuciri ce le activeaza recombinarea.
Ca sa testam aceasta idee am propus un set de experimente paralele in vitro/in vivo folosind un vector ADN
substrat cu doua RSS-uri
identice (unul reporter si
celalalt constitutiv) localizate
langa un promoter heterolog al
fagului T7 drept activator si un
RSS distal complementar cu
cele 2 pentru imperechere. Am
modificat un pic design-ul
initial din propunerea de
proiect (pg. 29, Fig.7), in asa
fel incat noul substrat sa
raspunda mai bine efectelor
structurale dovedite in
experimentele anterioare cu
topoizomeri si FRET. Avand in vedere ca 23-
RSS are afinitate mai mica pentru legarea
RAG si necesita pentru aceasta prezenta
proteinei HMGB in noii constructi atat RSS –
ul constitutiv cat si cel reporter au fost alesi
12-RSS. Drept vector de baza am folosit
secventele pJH290 unde alaturi de 12-RSS-ul
sau am inserat unul identic cu acesta si un
promoter al RNA polimerazei fagului T7 intre
ele. Constructii au fost asamblati cu cei 2x12-RSS in orientare deletionala si inversionala fata de 23-RSS-ul
distal cum este ilustrat in Fig. 14. Pentru a testa efectele orientarii sinaptozomului ambele 12-RSSuri au fost

14
pozitionate in imediata vecinatate a promoterului T7 astfel incat transcrierea induce in 12-RSSul din aval
suprarasucire pozitiva(12-2 pt. T7Inv sau 12-1pt. T7del.) si negativa in cel pozitionat in amonte fata de
sensul sau(12-1 pentru T7 Inv si 12-2 pt T7 del.). Pentru oprirea transcrierii am folosit cate un terminator
pozitionat la 200 de baze in josul promotorului pt. fiecare orientare. In gelul din Fig.14 sunt migrate produsii
de amplificare PCR cu primerii 1M si4M care amplifica regiunea din construct dintre 12/23 RSS distal in
lipsa recombinarii. Atat pentru experimentele in vitro cat si pentru cele in vivo am obtinut date
preliminarii doar pentru substratul 2x12RSS T7Inv. Experimentele de recombinare in vivo au fost efectuate
cotransfectand fibroblaste umane 293T HEK cu substrat 2x12RSS T7Inv, vectori pEBB exprimand RAG1 si
RAG2 proteine intregi si vectorul pAR3132T7RNP_NLS(6) exprimand RNA polimeraza fagului T7. Fig.
15A ilustreaza schematic procesul de recombinare pe vectorul inversional mediat prin RAG fie la nivelul
12-1 sau al 12RSS-2, si pozitionarea primerilor folositi in amplificari PCR individualizate pentru detectia
fiecarui tip de produs de recombinare; P1(12-1) and P2(12-2). In Fig. 16B este ilustrat gelul de separare al
unui set tipic de amplificari PCR incluzand reactiile control si folosind drept matrite de copiere vectori
extrasi din celule transfectate si mentinute in cultura 48h. posttransfectii. PCR1 detecteaza referinta ADN-ul
intersemnal (1M si 4M) de pe vectorul nerecombinat PCR2 evidentiaza produsul P1 iar PCR3 produsul P2.
Cinci seturi de amplificari PCR au fost executate pentru fiecare tip de transfectie si produsii de reactie dupa
separare pe 5% PAGE si colorare cu SybrGreen au fost cuantificati folosind un scanner Typhoon
Fluorescent. Rezultatele provenind de la 5 seturi de amplificari corespunzatoare a trei experimente cu
transfectii individuale sunt ilustrate cumulativ ca medii in histograma din Fig. 16C. Rezultatele sunt
exprimate fie ca raport relativ ai produsilor detectati P1 (rosu) sau P2(albastru) in prezenta(+T7) sau
absenta(-T7) T7RNA polimerazei, sau a raportului P1/P2(verde) in conditii cu transcriere T7 divizat la
raportul dintre produsi obtinuti in absenta transcrierii. Fig. 15 D ilustreaza analiza de tip imunoblot aratand
prezenta sau absenta expresiei proteice dupa 48 h posttransfectii complet dirijate de acest process. Fig. 16A
ilustreaza reactii de RT-PCR facute cu diferite dilutii de ARN din extracte celulare tratate cu DNAase I,
urmate de +/-Reverse Transcriere si amplificare PCR cu un set de primeri flancand transcriptul de la
promoterul T7. Se vede clar prezenta marcata a transcrierii initiate de acest promoter in celulele transfectate
cu vector exprimand T7RNA polimeraza (+T7) spre deosebire de cele crescute in absenta acestui vector(-
T7). In Fig. 16 B sunt descrise dilutii ale reactiilor specifice de PCR folosite pentru cuantificarile obtinute in
Fig.15C.
Transcrierea initiata specific de la promoterul T7 activeaza doar moderat recombinarea RAG in
special la nivelul lui 12RSS-1 (Transcrierea aproape dubleaza eficienta recombinarii la acest sit, vezi P1
+T7/-T7 din Fig15C) care in acest regim devine suprarasucit negativ. Mult mai putin dramatica este
cresterea recombinarii P2 la 12RSS2 ceea ce ar corespunde in timpul unei initieri sustinute a transcrierii unei
stari de suprarasucire pozitiva. Faptul ca transcrierea la acest construct activeaza recombinarea la ambele
12RSS-uri este pus in evidenta indirect si de raporturile P1/P2 ce par putin afectate de transcriere desi asa
15
cum am evidentiat fiecare produs in parte este activat. Aceste rezultate sugereaza ca sinaptozomul s-ar forma
izotropic acceptand RSS-uri suprarasucite atat negativ cat si pozitiv ambele semne facilitand indoirea
extrema a ADN-ului.
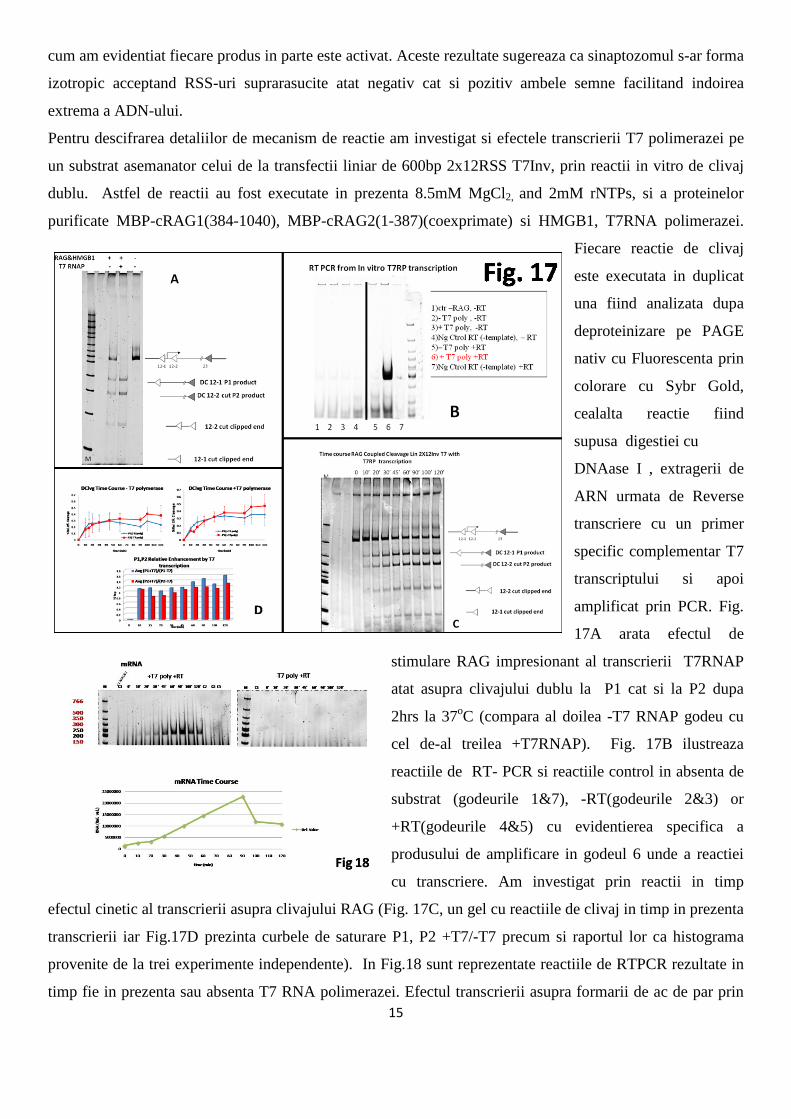
Pentru descifrarea detaliilor de mecanism de reactie am investigat si efectele transcrierii T7 polimerazei pe
un substrat asemanator celui de la transfectii liniar de 600bp 2x12RSS T7Inv, prin reactii in vitro de clivaj
dublu. Astfel de reactii au fost executate in prezenta 8.5mM MgCl2, and 2mM rNTPs, si a proteinelor
purificate MBP-cRAG1(384-1040), MBP-cRAG2(1-387)(coexprimate) si HMGB1, T7RNA polimerazei.
Fiecare reactie de clivaj
este executata in duplicat
una fiind analizata dupa
deproteinizare pe PAGE
nativ cu Fluorescenta prin
colorare cu Sybr Gold,
cealalta reactie fiind
supusa digestiei cu
DNAase I , extragerii de
ARN urmata de Reverse
transcriere cu un primer
specific complementar T7
transcriptului si apoi
amplificat prin PCR. Fig.
17A arata efectul de
stimulare RAG impresionant al transcrierii T7RNAP
atat asupra clivajului dublu la P1 cat si la P2 dupa
2hrs la 37oC (compara al doilea -T7 RNAP godeu cu
cel de-al treilea +T7RNAP). Fig. 17B ilustreaza
reactiile de RT- PCR si reactiile control in absenta de
substrat (godeurile 1&7), -RT(godeurile 2&3) or
+RT(godeurile 4&5) cu evidentierea specifica a
produsului de amplificare in godeul 6 unde a reactiei
cu transcriere. Am investigat prin reactii in timp
efectul cinetic al transcrierii asupra clivajului RAG (Fig. 17C, un gel cu reactiile de clivaj in timp in prezenta
transcrierii iar Fig.17D prezinta curbele de saturare P1, P2 +T7/-T7 precum si raportul lor ca histograma
provenite de la trei experimente independente). In Fig.18 sunt reprezentate reactiile de RTPCR rezultate in
timp fie in prezenta sau absenta T7 RNA polimerazei. Efectul transcrierii asupra formarii de ac de par prin
16
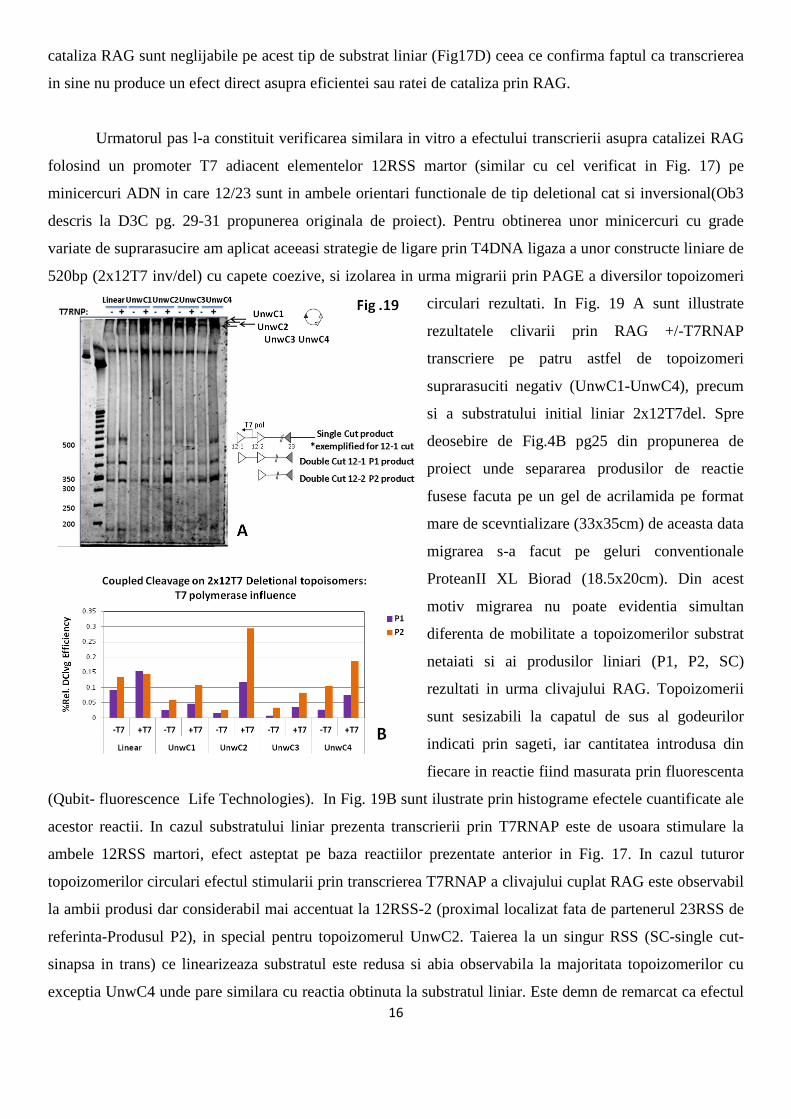
cataliza RAG sunt neglijabile pe acest tip de substrat liniar (Fig17D) ceea ce confirma faptul ca transcrierea
in sine nu produce un efect direct asupra eficientei sau ratei de cataliza prin RAG.
Urmatorul pas l-a constituit verificarea similara in vitro a efectului transcrierii asupra catalizei RAG
folosind un promoter T7 adiacent elementelor 12RSS martor (similar cu cel verificat in Fig. 17) pe
minicercuri ADN in care 12/23 sunt in ambele orientari functionale de tip deletional cat si inversional(Ob3
descris la D3C pg. 29-31 propunerea originala de proiect). Pentru obtinerea unor minicercuri cu grade
variate de suprarasucire am aplicat aceeasi strategie de ligare prin T4DNA ligaza a unor constructe liniare de
520bp (2x12T7 inv/del) cu capete coezive, si izolarea in urma migrarii prin PAGE a diversilor topoizomeri
circulari rezultati. In Fig. 19 A sunt illustrate
rezultatele clivarii prin RAG +/-T7RNAP
transcriere pe patru astfel de topoizomeri
suprarasuciti negativ (UnwC1-UnwC4), precum
si a substratului initial liniar 2x12T7del. Spre
deosebire de Fig.4B pg25 din propunerea de
proiect unde separarea produsilor de reactie
fusese facuta pe un gel de acrilamida pe format
mare de scevntializare (33x35cm) de aceasta data
migrarea s-a facut pe geluri conventionale
ProteanII XL Biorad (18.5x20cm). Din acest
motiv migrarea nu poate evidentia simultan
diferenta de mobilitate a topoizomerilor substrat
netaiati si ai produsilor liniari (P1, P2, SC)
rezultati in urma clivajului RAG. Topoizomerii
sunt sesizabili la capatul de sus al godeurilor
indicati prin sageti, iar cantitatea introdusa din
fiecare in reactie fiind masurata prin fluorescenta
(Qubit- fluorescence Life Technologies). In Fig. 19B sunt ilustrate prin histograme efectele cuantificate ale
acestor reactii. In cazul substratului liniar prezenta transcrierii prin T7RNAP este de usoara stimulare la
ambele 12RSS martori, efect asteptat pe baza reactiilor prezentate anterior in Fig. 17. In cazul tuturor
topoizomerilor circulari efectul stimularii prin transcrierea T7RNAP a clivajului cuplat RAG este observabil
la ambii produsi dar considerabil mai accentuat la 12RSS-2 (proximal localizat fata de partenerul 23RSS de
referinta-Produsul P2), in special pentru topoizomerul UnwC2. Taierea la un singur RSS (SC-single cut-
sinapsa in trans) ce linearizeaza substratul este redusa si abia observabila la majoritata topoizomerilor cu
exceptia UnwC4 unde pare similara cu reactia obtinuta la substratul liniar. Este demn de remarcat ca efectul
17
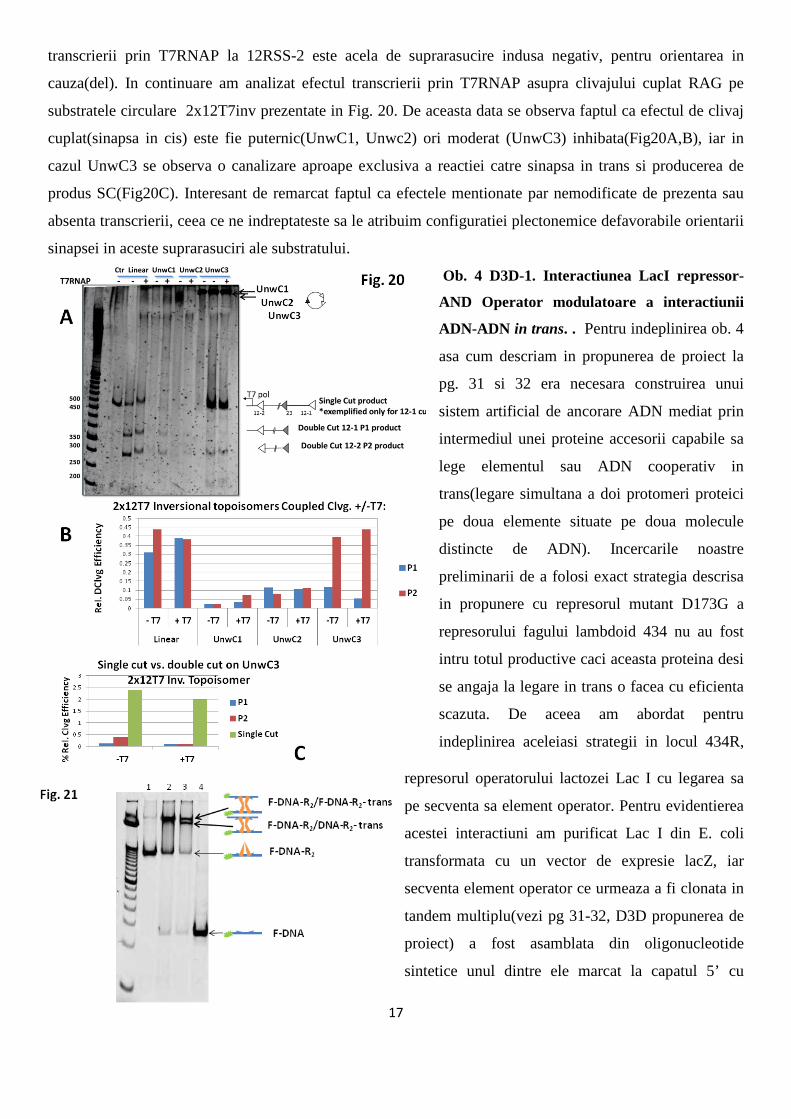
transcrierii prin T7RNAP la 12RSS-2 este acela de suprarasucire indusa negativ, pentru orientarea in
cauza(del). In continuare am analizat efectul transcrierii prin T7RNAP asupra clivajului cuplat RAG pe
substratele circulare 2x12T7inv prezentate in Fig. 20. De aceasta data se observa faptul ca efectul de clivaj
cuplat(sinapsa in cis) este fie puternic(UnwC1, Unwc2) ori moderat (UnwC3) inhibata(Fig20A,B), iar in
cazul UnwC3 se observa o canalizare aproape exclusiva a reactiei catre sinapsa in trans si producerea de
produs SC(Fig20C). Interesant de remarcat faptul ca efectele mentionate par nemodificate de prezenta sau
absenta transcrierii, ceea ce ne indreptateste sa le atribuim configuratiei plectonemice defavorabile orientarii
sinapsei in aceste suprarasuciri ale substratului.
Ob. 4 D3D-1. Interactiunea LacI repressor-
AND Operator modulatoare a interactiunii
ADN-ADN in trans. . Pentru indeplinirea ob. 4
asa cum descriam in propunerea de proiect la
pg. 31 si 32 era necesara construirea unui
sistem artificial de ancorare ADN mediat prin
intermediul unei proteine accesorii capabile sa
lege elementul sau ADN cooperativ in
trans(legare simultana a doi protomeri proteici
pe doua elemente situate pe doua molecule
distincte de ADN). Incercarile noastre
preliminarii de a folosi exact strategia descrisa
in propunere cu represorul mutant D173G a
represorului fagului lambdoid 434 nu au fost
intru totul productive caci aceasta proteina desi
se angaja la legare in trans o facea cu eficienta
scazuta. De aceea am abordat pentru
indeplinirea aceleiasi strategii in locul 434R,
represorul operatorului lactozei Lac I cu legarea sa
pe secventa sa element operator. Pentru evidentierea
acestei interactiuni am purificat Lac I din E. coli
transformata cu un vector de expresie lacZ, iar
secventa element operator ce urmeaza a fi clonata in
tandem multiplu(vezi pg 31-32, D3D propunerea de
proiect) a fost asamblata din oligonucleotide
sintetice unul dintre ele marcat la capatul 5’ cu

18
fluoroforul Alexa484. Experimentul descris in Fig. 21 este un experiment de EMSA(electrophoretic mobility
shift assay) ce evidentiaza legarea specifica a Lac I de operator intai in cis(DNA-R2-dimer Lac I legat de o
secventa operator-godeul 1, la un raport molar 1,25:1, LacI:opADN). Cand incubarea represorului cu ADN-
ul marcat se face prin adaugarea unui exces de ADN operator fie in totalitate marcat(godeul 2), fie
aproximativ jumatate marcat-godeul 3(1:1,6, lacI:opADN) complexul proteina ADN se formeaza prevalent
in trans astfel mediind reactia noastra de interes. Aceste rezultate preliminarii sunt incurajatoare, si pe baza
lor am ansamblat un polioperatorADN cu secvente multiple in tandem, construct folosit drept model de
ancorare in experimentele descrise mai jos.
D3D-2. Interactiunea LacI repressor-ADN Operator dirijeaza recombinarea VDJ in trans. Avand in
vedere faptul ca interactiunea de legare Lac I repressor
tetramer de cate un singur operator pe doua molecule ADN
poate media cu eficienta apropierea acestora in trans am
incercat sa amplificam acest efect generand doua constructe
de ADN plasmidial ce contin cate 8 secvente operator insiruite in tandem, adiacente, fiecare octamer 8xOp
flancand fie un 12 sau un 23 RSSr-reporteri (Fig. 22, sau Ob4 D3D Fig.8 pg 31 din propunerea de proiect)
de fiecare parte. Pentru a evidentia daca legarea de LacI represor poate in mod cooperativ reduce eficienta
clivajului RAG in cis, pe constructul cu 12RSS flancat de 8xOp am adaugat la distanta si un 23RSSc
constitutiv. Experimentele prin care testam efectul produs de tetramerii de Lac I repressor au fost descrise la
Ob4 D3D Fig.8 pg 31-32 din propunerea de proiect si aici vom aminti doar pe scurt rationamentul
desfasurarii lor. Primul set de experimente testeaza daca medierea prin complexele LacI represor tetramer
formate intre constructele cu 12 si 23RSSr reporter prin legarea de cele 4 multisituri repetate in tandem la
8xOp duce la o ghidare moleculara a complexului recombinazic RAG spre o cataliza preferentiala in
trans(versus cea naturala 12RSS reporter cu 23RSSc). Acest lucru presupune testarea efectului catalitic
RAG pe cele doua constructe din Fig. 22 in prezenta sau absenta unei concentratii corespunzatoare de LacI

19
Represor. In Fig. 23 avem imaginea marita (vezi gelul in ansamblu in coltul din dreapta jos al imaginii) a
separarii produsilor de clivaj ADN generati de RAG in urma a 13 reactii intreprinse distinct in conditiile
specificate de legenda de deasupra gelului. In diagrama din stanga imaginii gelului sunt reprezentati
schematic cei patru produsi rezultati in urma clivarii de catre RAG la nivelul 12-RSSr (cel mai redus in
mobilitate electroforetica), 23-RSSc (imediat sub cel descris anterior), 23-RSSr (cu mobilitate intermediara),
si cel de dublu clivaj cu sinapsa in cis 12-RSSr & 23-RSSc (banda mica de mobilitate maxima
electroforetica). In mod remarcabil clivarea de catre RAG a ADN-ului in trans necesita atat prezenta
ambelor tipuri de constructe plasmidiale unul cu 12-RSSr si celalalt cu 23-RSSr cat si cea a proteinei de
docare represorul LacI (compara rezultatele din godeurile 1,3, 4, 5, 7 cu cele din reactiile 2 si setul 8-13).
Pentru a pune in evidenta in ce masura concentratia de represor LacI influenteaza gradul de proximitate
mediata a 12-RSSr si 23-RSSr setul de reactii din reactiile 3-7 trebuie in mod corespunzator comparate cu
cele din setul 8-13. Deoarece efectul detectabil la nivelul clivajului unic 12-RSSr se datoreaza sumativ atat
sinapsei in trans (cu 23-RSSr) cat si a celei cu 23-RSSc de pe acelasi ADN, el poate fi urmarit numai
corelativ in sensul sincronizarii celor doua efecte. Produsul de clivaj unic mediat prin RAG la 23-RSSr este
cel ce indica electiv eficienta imperecherii sinaptice in trans si de aceea este indicatorul de maxima acurateta
a fenomenului urmarit si indus de noi si modulat prin concentratia de LacI represor. Pentru a urmari astfel de
efecte corelative produsii de reactie izolati electroforetic precum cei din reactiile din Fig. 23 au fost
cuantificati fluorescent pe imaginile gelurilor si intensitatea lor relativa reprezentata in functie de
concentratia de represor Lac I inclusa in reactie. Aceste curbe de eficienta a clivajului unic (in cazul sit-
urilor reporter)
sau dublu(12-
RSSr & 23-
RSSc) au fost
reprezentate
grafic in Fig. 24
Curbele
reprezentate in
albastru sunt
reactiile control
in care ADN-ul
partener reactiei
in trans este absent in timp ce cele in rosu contin ambele plasmide cu sit-urile in trans. Pe axa y este
reprezentata intensitatea relative fluorescent a produsului de reactie RAG urmarit in timp ce pe abscisa sunt
rprezentate concentratiile de represor Lac I din reactie. In Fig. 24C se vede efectul remarcabil al represorului
Lac I care intr-un ordin de 0.1-1µM creste eficienta reactiei de clivaj unic la 23-RSSr de aproape 5 ori,

20
numai in prezenta partenerului 12RSSr. Acest effect este observat si la sit-ul partener 12-RSSr(Fig. 24A),
confirmand corelativ faptul ca imperecherea trans 12RSSr/23-RSSr este substantial facilitata de legarea
cooperativa a tetramerilor represorilor LacI la cele doua set-uri de octameri 8xOct ce le flancheaza.
Interesant insa este si efectul de inhibitie al reactiei de clivaj RAG dublu 12RSSr/23-RSSc prin sinapsa in
cis pe acelasi ADN, efect independent insa de prezenta partenerului 23RSS reporter in trans (Fig. 24B
curbele in rosu si albastru arata acelasi efect stimulativ). Acest efect incercam sa-l atribuim posibil modularii
de proximitate a celor 2 sit-uri in cis prin faptul ca represorul Lac I probabil indoind ADN-ul inter-sit-uri
scade concentratia efectiva a celor doua RSS-uri de pe acelasi ADN unul fata de celalalt, astfel reducand
sansele sinapsarii in cis. Scopul primar al acestor experimente a fost atins si anume acela de a reproduce in
vitro(reactia chimica RAG+AND+LacI repressor) fenomenul ce se produce patologic in cazul translocatiilor
cromozomiale din limfoamele primare B sau T. Mentionam ca acest rezultat reprezinta prima tentativa de a
dirija artificial in vitro sinapsa RAG pe sit-uri RSS localizate in trans. Pasul urmator il reprezinta
caracterizarea cinetica a acestui fenomen si influenta transcrierii asupra modularii configuratiei RSS-urilor
in trans(vezi mai jos).
D3D-3. Efectul suprarasucirii trasncriptionale a unuia din situ-urile RSS implicate in recombinarea
VDJ in trans. Rezultatele pozitive ale experimentelor
prezentate la punctele D3D-1 si 2 ne permit reinvestigarea
reactiei de clivaj cu doua RSS-uri flancate de 8xOp localizate
in trans dar de aceasta data influentand configuratia 12RSSc
constitutiv prin transcrierea concomitenta cu clivajul cu T7RNA polimeraza (Fig. 25A). Acest experiment
este doar unul preliminar de tip pilot prin care am inceput sa investigam efectul transcrierii in afara
contextului de mediere a docarii ADN-ADN prin legarea represorului LacI. Asa cum este ilustrat in
diagrama din Fig. 25A unul din plasmide contine atat RSS-ul constitutiv 12RSSc, sub influenta promoterului
T7 in aval cat si pe cel reporter 23RSSr flancat in ambele parti de 8xOp. Celalalt plasmid contine doar
12RSS reporter flancat de octameri 8xOp. In Fig. 25B este ilustrat scan-ul obtinut prin fluorescenta din
gelul nedenaturat(nativ) folosit la separarea electroforetica a produsilor de reactie de clivaj indus prin RAG
in prezenta represorului LacI(godeurile 2,4,6 si 8) sau absenta sa (godeurile 1,3,5 si 7). In Fig. 25C sunt
prezentate rezultatele reactiilor de RT-PCR ce pun in evidenta un produs de aprox. 200bp. corespunzator
lungimii transcriptului nostru, in prezenta T7 polimerazei (3, 7 si8). In Fig 25C primele 9 godeuri din stanga
reprezinta reactiile in prezenta revers transcriptazei iar ultimele din dreapta reactiile control in absenta
acestei enzime. Numerele reactiilor corespund cu cele din Din pacate reactia pilot prezentata nu a fost
suficient de bine tratata cu RNAaza A si urmele de ARN prezente in fiecare reactie au influentat dramatic
mobilitatea electroforetica a produsilor de clivaj enzimatic detectati. In special trebuie sa mentionam
imposibilitatea detectiei clivajului unic la 12RSSc desenat in diagram intre paranteze. In ciuda acestor
inconveniente tehnice regretabile reactia de clivaj evidentiaza remarcabil cresterea substantiala a clivarii

21
duble 12RSSc/23RSSr (banda cu mobilitate maxima de jos) ori de cate ori reactia faciliteaza sinapsa prin
transcriere in cis (compara godeurile 1 cu 3, 3 cu 4, si 7 cu 8, atunci cand reactia nu contine represor LacI
/12RSSr LacI dar are transcriere. Aceasta observatie intareste si mai convingator concluziile privind efectul
stimulator al transcrierii pe RSS-uri orientate in cis deletional descrise la pg 16 si 17 ale acestui raport.
Urmeaza sa caracterizam efectul dinamic al transcrierii exercitat asupra unui RSS reporter pe reactia in trans
si ulterior sa construim plasmidele de transfectie pentru repetarea acestor experimente si evidentierea
efectului de docare al represorului LacI pe cataliza RAG in culture celulare. Experimentele de evidentiere a
recombinarii in trans la nivel celular definitiveaza cel de-al IVlea obiectiv propus in proiect.
CONCLUZII
In aceste 42 luni de activitate am reusit:
a) Sa exprimam si sa purificam in regim constant si de rutina proteine active RAG1, RAG2 and HMGB1/2
din diverse surse reactivi extrem de pretiosi pentru toate studiile recombinarii RAG in vitro.
b) Folosind un assay de ligare facilitata pe substratul IS264 am determinat ca RAG induce activ prin sinapsa
(<5min) acumularea substantiala de produs circular suprarasucit. Generarea acestor produsi de reactie initial
postula dar apoi am putut si demonstra prin studii de FRET ca RAG in timpul sinapsei impleteste
plectonemic activ 12si 23RSS-urile indoindu-le si suprarasucindu-le in sinaptozom. Acest efect faciliteaza
dramatic sinapsa in cis experimentele de ligare conducand la o acumulare rapida si eficienta de cercuri
monomerice suprarasucite in timp ce producerea prin sinapsa RAG in trans de multimeri este greu
detectabila.
c) Am investigat si mecanismul prin care clivajul RAG este activat de gradul de suprarasucire la care este
supus substratul sau ADN. Am dezvoltat o metoda noua de clivaj dublu RAG pe topoizomeri. Prin ea am

22
analizat cataliza RAG pe 10 topoizomeri cu lungime restransa ce constrange orientarea de faza elicoidala a
celor doua 12/23RSS-uri. Clivajul este in mod substantial crescut de catre topoizomerii suprarasuciti si
respecta orientarea optima de faza determinata si la sinapsa evidentiata prin ligari.Aceste rezultate
demonstreaza fara echivoc ca suprarasucirea RSS-urilor induce o reducere a barierei de energie de activare
pe care RAG ar trebui sa o invinga in timpul sinapsei prin indoirea RSS-urilor ADN relaxat.De aceea
aceasta stare induce activarea catalizei.
d) Pentru a investiga configuratia RSS-urilor in sinapsa am folosit metodologia FRET si din nevoia de a
depasi un design experimental neproductiv propus innitial. Prin aceasta am investigat configuratiile
12&23RSS in complexul sinaptic RAG si am construit pe baza datelor FRET un model detaliat atat al 12 cat
si al 23-RSS in PC si SC. Configuratia 23-RSS in sinaptozom a fost publicata in luna ianuarie 2013
(Ciubotaru et. al. in Nucleic Acids Research 2013, vol. 41, 2437-54) (3) intr-o revista prestigioasa in
domeniul biologiei moleculare cu factor de impact 8.4. Configuratia 12-RSS atat in sinaptozom cat si in
complexul unic RAG/HMGB1 a fost elucidata prin aceeiasi metodologie de FRET si rezultatele acestei
investigatii au fost publicate in luna ianuarie 2015 la aceeasi revista Nucleic Acids Research (Ciubotaru et.
al. in Nucleic Acids Research 2015, vol43, 917-931)(8). Rezultatele docarii si modelarii molecular-
structurale a configuratiilor 12/23RSS in interiorul sinaptozomului au fost publicate in review in 2014 in
revista Discoveries (7).
e) Am demonstrat atat prin experimente in vivo cat si in vitro faptul ca suprarasucirea indusa de transcriere
poate modula dramatic activitatea RAG atunci cand are loc la promoterii adiacenti RSS-urilor. Rezultatele
noastre subliniaza consistent stimularea clivajului prin RAG la orientarea deletionala in contextul
suprarasucirii negative, si presupune inhibitia severa a sinapsei in cis atunci cand situ-rile 12/23RSS sunt in
configuratia inversionala a aceleiasi suprarasuciri. Aceste rezultate au fost prezentate oral la a 9a Conferinta
de Biomolecular Dynamics Albany 2015 in SUA, iar abstractul acestei prezentari a fost publicat ca o
comunicare scurta in Journal of Biomolecular Structure and Dynamics 2015, vol33, Sup1, p71(9).
f) Cu ajutorul interactiunii represor LacI-operator ADN, evidentiata in trans, am edificat un sistem de
modulare a ancorarii intermoleculare ADN-ADN pentru indeplinirea obiectivului 4.
g) Am testat modul cum sistemul proiectat de noi LacI represor –operator ADN, dirijeaza cu prevalenta
cataliza de clivare RAG in trans, astfel simuland fenomenologia ce produce in mod patologic translocatiile
cromozomiale in cursul recombinarii V(D)J.
h) Am optimizat conditiile de producere a reactiei de clivaj prin RAG in trans, si aceste conditii au fost
testate ulterior prin reactii de echilibru. Din cuantificarea produsilor de reactie obtinuti estimam o crestere
de cel putin 5 ordine de marime a reactiei in trans atunci cand ea este dirijata prin Lac I versus reactiile
control in absenta sa. Vom reface reactiile in conditii optime(concentratia fiecarui plasmid ADN continand
un RSSr reporter, si a represorului Lac I) in conditii cinetice pentru a caracteriza sistemul complex prin
constantele enzimatice.

23
g) In reactii pilot am testat efectul transcrierii de a competitiona activ la un RSS constitutiv(neflancat de
operatori) cu sistemul de docare in trans, si rezultatele preliminare sustin convingator faptul ca transcrierea
sustine orientarea recombinarii in cis.
References 1)Ciubotaru, M., et al. Fluorescence resonance energy transfer analysis of recombination signal sequence
configuration in the RAG1/2 synaptic complex. Mol Cell Biol 27, 4745-4758, (2007).
2) Yin, F. F. et al. Structure of the RAG1 nonamer binding domain with DNA reveals a dimer that mediates DNA
synapsis. Nat Struct Mol Biol 16, 499-508, (2009).
3) Ciubotaru, M. Trexler A. J., Spiridon L., Surleac M. D., Rhoades E, ., Petrescu A. J., Schatz D.G. “RAG and
HMGB1 create a large bend in the 23RSS in the V(D)J recombination synaptic complexes. Nucleic Acids Research,
2013,vol 41, 2437-2454
4) Swanson, P. C Fine structure and activity of discrete RAG-HMG complexes on V(D)J recombination signalsMol
Cell Biol 22, 1340-1351, (2002). 5)Zwilling, S. et al. High mobility group protein 2 functionally interacts with the POU domains of octamer transcription factors, EMBO J. 14, 1198-1208, (1995). 6) Dunn, J. J. et al. Targeting bacteriophage T7 RNA polymerase to the mammalian cell nucleus,Gene, vol68,259-266,(1988). 7)Ciubotaru M., Surleac M.D., Musat G. M., Rusu A. M., Ionita E., Albu C. C. Paul " DNA bending in the synaptic complex in V(D)J recombination: turning an ancestral transpososome upside down”, Discoveries, 2(1):e13:1-15, 2359–7232(2014).
8) Ciubotaru M. , Surleac M. D., Metskas L. A., Koo P., Rhoades E., Petrescu A. J., Schatz D.G.” The architecture of the 12RSS in V(D)J recombination signal and synaptic complexes “, Nucleic Acids Research, 2015, vol43, 917-931 9) Ciubotaru M.”V(D)J recombination targeted in cis by transcription induced DNA supercoiling”, Journal of Biomolecular Structure and Dynamics, 2015, vol 33, Sup1, p71. DOI: 10.1080/07391102.2015.1032746.