raport ŞtiinŢific Şi tehnic etapa 3 / 2013 obiectivul...
TRANSCRIPT

1
RAPORT ŞTIINŢIFIC ŞI TEHNIC – ETAPA 3 / 2013
Obiectivul etapei
Desfăşurarea experimentului de electroterapie în vederea eradicării virusurilor la viţa-de-vie şi
cartof
Rezultate aşteptate
- evaluarea acţiunii curentului electric asupra evoluţiei subculturilor prin măsurători biometrice şi
cuantificarea unor compuşi biochimici;
- model al interacţiei câmp electromagnetic – material biologic;
- monitorizarea proceselor fiziologice la plantele de viţă-de-vie şi cartof selecţionate sănătoase prin
chimioterapie,
- elaborarea tehnologiilor de eliminare a virusurilor specifice la viţa-de-vie şi cartof (2),
- diseminarea rezultatelor prin publicaţii.
Rezumat
Stimularea electrică în cuva de electroforeză orizontală constituie una din metodele de eliminare
a virusurilor la viţa-de-vie şi cartof. Electroterapia se bazează pe aplicarea unui câmp electric intens în
apropierea materialului vegetal care duce prin efect corona, la apariţia în apropierea membranei
celulare a unor distribuţii de sarcină electrică spaţială, ce face ca tensiunea transmembranară şi curentul
transmembranar să fie diferite de cele corespunzătoare stării staţionare şi să inducă modificări
intracelulare soldate cu inactivarea proteinei virale. Tratamentul lăstarilor erbacei cu diferite intensităţi
ale curentului electric (40, 50, 100 mA) timp de 5, 10, 15 minute şi inocularea lor pe medii de cultură
specifice nu a influenţat în sens negativ viabilitatea explantelor. Eventuala influenţă a tratamentului
electric a fost apreciată prin cuantificarea ratelor de multiplicare şi a unor compuşi biochimici, pe
parcursul a trei subculturi. La viţa-de-vie, la prima subcultură, apexurile tratate electric au multiplicat
semnificativ diferit pe variantele experimentale, la ambele genotipuri studiate, fără o variaţie liniară cu
creşterea intensităţii curentului sau a timpului, în timp ce la fragmentele nodale nu s-au înregistrat
diferenţe semnificative comparativ cu martorul. La următoarele două subculturi, variaţia ratelor de
multiplicare pe variante comparativ cu matorul s-a menţinut, valorile acestui indice a crescut conform
tehnicii de micropropagare in vitro, asigurând regenerarea unui număr mare de plante. La cartof, rata de
multiplicare a crescut în toate variantele experimentale faţă de martorul netratat, ceea ce denotă faptul
că, efectul benefic al electroterapiei este mai intens decât al altor tehnici convenţionale de devirozare
(cultura de meristem, termoterapia). Rata de multiplicare a explantelor in vitro a fost influenţată
semnificativ atât de tipul virusului, cât şi de intensitatea curentului şi durata tratamentului. După prima
subcultură post tratament electric cuantificarea unor indicatori biochimici (conţinutul în zaharuri
solubile, polifenoli totali, pigmenţi clorofilieni şi carotenoizi) la plantulele de viţă-de-vie, nu a pus în
evidenţă diferenţe semnificative la variantele experimentale comparativ cu martorul. Diferite intensităţi
ale curentului precum şi timpi de tratament diferiţi nu şi-au manifestat acţiunea pe termen lung,
indicatorii evaluaţi nu au înregistrat diferenţe semnificative comparativ cu martorul pe parcursul celor
trei subculturi. La cartof, dozarea conţinutului în amidon a înregistrat o dependenţă direct proporţională
cu intensitatea curentului electric şi implicit cu durata de expunere. După trei subculturi consecutive, la
explantele infectate cu ambele virusuri s-a înregistrat o scădere a conţinutului de proteină brută, în
special în cazul variantelor de tratament mai severe şi mai ales la materialul infectat cu virusul Y.
Analiza unor indicatori fiziologici (intensitatea fotosintezei, a respiraţiei, pigmenţi clorofilieni etc.) a
pus în evidenţă diferenţe semnificative la plantele de viţă-de-vie şi cartof regenerate libere de virusuri,
care nu pot fi puse numai pe seama prezenţei chimioterapicelor, ci şi a influenţelor induse de condiţiile

2
de mediu şi genotip. Rezultatele cercetărilor obţinute până în această fază de derulare a proiectului au
fost valorificate prin publicarea de lucrări ştiinţifice.
Descrierea ştiinţifică şi tehnică
1. Parcurgerea etapelor de regenerare in vitro a plantulelor de viţă-de-vie libere de virusuri, după
aplicarea electroterapiei - P1
Viţa-de-vie este una din speciile horticole cele mai susceptibile a fi infectate de diverşi agenţi
infecţioşi printre care şi virusurile. Selecţia riguroasă din punct de vedere al prezenţei infecţiei virale, a
plantelor utilizate la producerea materialului de înmulţire horticol constituie obiectivul principal atât al
amelioratorilor cât şi al cultivatorilor agricoli. Într-adevăr, începând cu 1960, state sau uniuni de state,
ca fosta Comunitate Economică Europeană, s-au implicat în definirea cadrului legal privind statutul
fitosanitar al viţei-de-vie (68/93 EEC), plante ornamentale (77/93 EEC) şi pomi fructiferi (92/34 EEC).
O abordare similară a fost întreprinsă de Organizaţia Nord Americană pentru Protecţia Plantelor
(NAPPO) (Panattoni şi colab., 2013).
Eliminarea virusurilor şi obţinerea materialului de înmulţire viticol sănătos a devenit necesară în
cazul genotipurilor de interes economic, în curs de omologare şi menţinerea unei surse de material
necesar amelioratorilor, liber de virusuri.
În cadrul proiectului a fost experimentată o modalitate de utilizare a curentului electric în
blocarea multiplicării virale. Fragmente erbacee provenite de la plante de viţă-de-vie infectate au fost
tratate electric în cuva de electroforeză orizontală la intensităţi crescătoare ale curentului (40, 50, 100
mA) şi timpi de stimulare diferiţi (5, 10, 15 minute). Materialul biologic luat în studiu a fost reprezentat
de genotipurile Tămâioasă românească 3-2-2 în curs de omologare aparţinând INCDBH Ştefăneşti-
Argeş, infectat cu fleck (GFkV) şi Frâncuşă 15 Od infectat cu virusul asociat răsucirii frunzei serotip 1
(GLRaV 1) aparţinând SCDVV Odobeşti.
Lăstarii trataţi au fost fragmentaţi în apexuri şi fragmente nodale, sterilizaţi şi cultivaţi pe mediu
specific viţei-de-vie în condiţii aseptice. Culturile au fost incubate în camere de creştere cu parametri
controlaţi (25±10C, 16/8 fotoperioada). După iniţiere, explantele au fost transvazate pe mediu proaspăt
la fiecare 30 de zile, timp de 3 subculturi.
După fiecare subcultură a fost cuantificată rata de multiplicare pe variantele experimentale, ca
indicator al evoluţiei proceselor de organogeneză ale explantelor tratate electric. Fiecare explant a fost
evaluat individual astfel încât, ratele de multiplicare constituie medii pe variantele experimentale.
Evaluarea ratelor de multiplicare după prima subcultură a pus în evidenţă atât la genotipul
Tămâioasă românească 3-2-2 cât şi la Frâncuşă 15 Od, diferenţe semnificative pe variante
experimentale comparativ cu martorul, în cazul apexurilor, ca tip de explant cu care au fost iniţiate
culturile.
Astfel, la genotipul Tămâioasă românească 3-2-2 infectat cu GFkV, stimularea electrică cu 40
mA conduce la scăderea semnificativă a ratei de multiplicare la 10 minute timp de tratament (V2). O
creştere a intensităţii curentului de numai 10 mA conduce la scăderea semnificativă a ratei la 5 minute
timp de stimulare, după care, creşterea timpului determină o mărire semnificativă a numărului de
fragmente de multiplicare, după 15 minute de stimulare a explantului iniţial. Dublarea intensităţii
curentului determină scăderea ratei de multiplicare la 5 minute, după care, indicatorul creşte
semnificativ odată cu creşterea timpului de tratament. Aşadar, o stimulare cu 50 mA timp de 15 minute
a unui apex şi cultivarea acestuia pe mediu de cultură steril conduce la o creştere semnificativă a
numărului de fragmente de multiplicare (Fig. 1.1). Stimularea electrică în cuva de electroforeză
orizontală a fragmentelor nodale nu s-a soldat cu diferenţe semnificative ale ratelor de multiplicare,
indiferent de varianta experimentală.
Comportarea diferită la stimuli electrici a diferitelor tipuri de explante demonstrează încă o dată
că, regenerarea plantelor prin micropropagare depinde de numeroşi factori printre care: genotipul,
stadiul fiziologic al explantului cultivat pe mediul de cultură, condiţiile de cultură, mediul de cultură,

3
precum şi interacţiile dintre aceşti factori (Svetleva şi colab., 2003). În cazul de faţă, la toate acestea se
adaugă prezenţa infecţiei virale, care are un rol determinant în declanşarea proceselor regenerative ale
explantelor cultivate in vitro (Guţă şi colab., 2009). Mai mult decât atât, curentul electric sub formă de
pulsuri determină stimularea diferenţierii in vitro a plantulelor (Goldsworthy, 1997). A fost raportată
îmbunătăţirea regenerării plantelor de cartof prin expunerea explantelor la curent electric (Lozoya-
Saldana şi colab., 1996).
Fig. 1.1 Influenţa curentului electric asupra ratei de multiplicare după prima subcultură,
la genotipul Tămâioasă românească 3-2-2 infectată cu GFkV. Valorile sunt medii pe 3 repetiţii, barele
indică deviaţiile standard ale mediilor, literele reprezintă semnificaţiile mediilor comparativ cu
martorul la P<0,05
Genotipul Frâncuşă 15 Od infectat cu GLRaV 1 se comportă similar, în sensul că ratele de
multiplicare ale explantelor provenite din apexuri stimulate electric, prezintă diferenţe semnificative
comparativ cu martorul, pe variantele experimentale, spre deosebire de cele determinate de evoluţia
fragmentelor nodale. Tratamentul apexurilor cu curent electric de 40, 50 mA conduce la scăderea
semnificativă a ratelor de multiplicare, în timp ce 100 mA determină creşterea semnificativă a
proceselor de regenerare după numai 5 minute de tratament. Mărirea timpului de stimulare electrică
influenţează în sens negativ declanşarea proceselor de morfogeneză (Fig. 1.2)
Fig. 1.2 Influenţa curentului electric asupra ratei de multiplicare după prima subcultură,
la genotipul Frâncuşă 15 Od infectată cu GLRaV 1. Valorile sunt medii pe 3 repetiţii, barele indică
deviaţiie standard ale mediilor, literele reprezintă semnificaţiile mediilor
comparativ cu martorul la P<0,05
a
a
b
a
a
a
b
a
a a
d a
b
a c a c
a a
a
0
2
4
6
8
1 2
Rat
a d
e m
ult
iplic
are
(x)
Tip de explant: 1-apex; 2-fragment nodal
V1
V2
V3
V4
V5
V6
V7
b
a
b
a
c a
b
a
b a b
a
d a
a a
b
a
a
a
0
1
2
3
4
5
1 2Rat
a d
e m
ult
iplic
are
(x)
Tip de explant: 1-apex;2-fragment nodal
V1
V2
V3
V4
V5
V6
V7

4
Este normal ca influenţa curentului electric să fie resimţită de către explante, la prima
subcultură. Apexurile ambelor genotipuri subcultivate pe mediu proaspăt (S2, S3), continuă să prezinte
diferenţe semnificative ale ratelor de multiplicare pe variantele experimentale comparativ cu martorul.
Valorile sunt crescătoare asigurând micropropagarea şi regenerarea unui număr mare de plante de viţă-
de-vie. Diferenţele de mărime între ratele celor două genotipuri studiate derivă din diferenţa dintre
virusuri şi chiar datorită potenţialului regenerativ diferit, specific soiului de viţă-de-vie (Fig. 1.3 şi 1.4).
Fragmentele nodale stimulate electric parcurg încă o subcultură fără modificări semnificative
ale ratelor de multiplicare. La subcultura a treia, genotipul Tămâioasă românească 3-2-2 înregistrează
cea mai mare valoare a indicatorului la varianta trei (40 mA, 15 min) şi anume, 19,33 ± 1,5275
comparativ cu 11 ± 8,7178, martorul. Genotipul Frâncuşă 15 Od se comportă similar, cea mai mare rată
de multiplicare diferenţiindu-se la prima variantă (40 mA, 5 min), (21,33 ± 4,1633) comparativ cu
martorul (7 ± 1,00).
Fig. 1.3 Influenţa curentului electric asupra ratei de multiplicare a apexurilor, în dinamică,
pe subculturi, la genotipul Tămâioasă românească 3-2-2 infectată cu GFkV. Valorile sunt medii pe 3
repetiţii, barele indică deviaţiile standard ale mediilor, literele reprezintă semnificaţiile mediilor
comparativ cu martorul la P<0,05
Fig. 1.4 Influenţa curentului electric asupra ratei de multiplicare a apexurilor, în dinamică, pe
subculturi, la genotipul Frâncuşă 15 Od infectată cu GLRaV 1. Valorile sunt medii pe 3 repetiţii, barele
indică deviaţiie standard ale mediilor, literele reprezintă semnificaţiile mediilor
a b
d
a a
b c
d c
a
d
i
a
c
h
a b
g
d
f
g
e
e
f
b
c
e
a a
a
0
5
10
15
20
25
S1 S2 S3
Rat
a m
ult
iplic
are
(x)
Subculturi
V1
V2
V3
V4
V5
V6
V7
V8
a a
a
b
a
d
a a
a
b a
b
c
c
e
e
d a
b
b
a
d
f
c
d
e
e
a a
a
0
5
10
15
20
25
S1 S2 S3
Rat
a m
ult
iplic
are
(x)
Subculturi
V1
V2
V3
V4
V5
V6
V7
V8

5
comparativ cu martorul la P<0,05
Bibliografie
Goldsworthy A., 1997 - Electrical stimulation of tissue culture growth and morphogenesis. Agriculture
cell report 8, 14.
Guţă I.C., Buciumeanu E.-C., Vişoiu E., 2009 – Comparative study of in vitro behavior of grapevine
(V.Vinifera L., Fetească neagră cv.) under the influence of various infections. Extended abstacts
16th
Meeting of ICVG, Dijon, France, 31 Aug.- 4 Sept., 247.
Lozoya-Saldaña H.F., Abelló J., García G., 1996 - Electrotherapy and shoot-tip culture eliminate potato
virus X in potatoes. Am. J. Potato Res. 73, 149-154.
Panattoni A., Luvisi A., Triolo E., 2013 – Review. Elimination of viruses in plants: twenty years of
progress. Span. J. Agric. Res.11 (1), 173-188.
Svetleva D., Velcheva M., Bhowmik G., 2003 - Biotechnology as a useful tool in common bean
(Phaseolus vulgaris L.) improvement. Euphytica 13, 189-200.
2. Evaluarea acţiunii curentului electric asupra dezvoltării plantulelor de viţă-de-vie pe parcursul
subculturilor prin cuantificarea unor compuşi biochimici - P1
Micropropagarea este o metodă de înmulţire vegetativă care se bazează pe totipotenţa celulei
vegetale, conform căreia, fiecare celulă conţine întreaga informaţie genetică necesară regenerării unui
alt individ. Mediile de cultură au o compoziţie complexă, combinaţia chimică şi proporţia substanţelor
fiind stabilită pe baza analizelor calitative şi cantitative făcute asupra cenuşii diferitelor specii de plante
şi a gazelor emanate la incinerarea acestora, precum şi din experienţe de nutriţie.
Macroelementele sunt elemente chimice de bază care intră în structura materiei vii: C, O, H, N,
S, P, K, şi Ca. Prezenţa lor este indispensabilă în mediile de cultură. Proporţia între diferiţi
macronutrienți variază de la o plantă la alta, dar, în general se apreciază că oxigenul alcătuieşte 70%
din materia vie, carbonul 18%, hidrogenul 10,5%, cele trei elemente însumate deţinând 98,5% din
compoziţia plantelor. P, K, S şi N sunt participante cu zecimi de procente. Ca şi K joacă un rol
important în funcţionarea celulei, în gradul de fluiditate a citoplasmei, în schimbul ionic, în
permeabilitatea membranei celulare etc. O parte mai redusă o au macroelementele: Mg, Na, Cl, Al, sub
formă de ioni, în mediul de dispersie sau în vacuole. Mg intră în molecula de clorofilă, Na joacă un rol
important în schimburile de ioni, dar excesul de Na poate cauza substituirea K din celule şi
alcalinizarea mediilor. Ionii de Ca în exces pot provoca acidifierea acestora, iar Al serveşte drept
catalizator într-o serie de reacţii metabolice. Aceste 12 elemente realizează cca. 99,99% din substanţa
vie. Microelementele, deși sunt necesare în cantități reduse, devin indispensabile în compoziția
mediilor de cultură, exercitând un rol deosebit asupra ţesuturilor crescute in vitro, deoarece intră în
compoziţia a numeroase combinaţii organometalice, complexe, cu valoare biologică ridicată, alcătuiesc
sisteme enzimatice-metaloenzimale (citocromoxidaza şi peroxidaza) fără de care procesele vitale nu se
pot desfăşura. Suprimarea lor totală produce reducerea creşterilor cu 40% începând cu prima subcultură
şi provoacă moartea explantelor la cea de-a treia subcultură. Fierul, molibdenul şi manganul împreună
măresc ritmul creşterii celulare. Fe intră în compoziţia unor metalo-enzime, este implicat în sinteza
proteică, iar împreună cu Mo sporeşte conţinutul de proteine în celule şi acumularea în ţesuturi de N
neproteic. Cuprul este necesar pentru creşterea rădăcinilor. Manganul este şi el implicat în sinteza
proteinelor, carenţa represând ARN din celule. Mn este necesar pentru creşterea rădăcinilor. Trebuie
avută în vedere proporţia între Mn/Mg, deoarece Mn într-o serie de reacţii metabolice provocând
perturbaţii celulare grave. Se va avea în vedere că unele substanţe chimice pot imobiliza chimic unii
ioni sau pot provoca precipitarea unor săruri. Cu toate că inoculii sunt verzi, celulele lor deşi posedă
cloroplaste, nu au capacitatea de a fi total autotrofe, menţinerea lor în viaţă depinzând de prezenţa unei
surse de carbon glucidic. Ţesuturile devin heterotrofe faţă de carbon. Cu timpul ţesuturile se
depigmentează, semnal că în mediul de cultură sunt absenţi compuşi implicaţi în sinteza clorofilei, ori a

6
slabei funcţionări a cloroplastelor. Acest fenomen de anemie clorofiliană, se poate datora şi lipsei de
fier. Ca sursă de carbon, în mediile de cultură se folosesc glucidele, în special zaharoza. Concentraţia
optimă este de 2%. Sunt situaţii când se cer concentraţii de 8% la care zaharoza fără a devenii toxică
este mai puţin eficientă. Vitaminele sunt prezente în toate mediile de cultură şi sunt indispensabile unei
multiplicări şi creşteri optime. Regulatorii de creștere (auxine, citochinine, gibereline) sunt compuși
similari fitohormonilor, având un rol primordial în multiplicarea, creşterea şi diferenţierea explantelor
cultivate „in vitro”, dar sunt de natură exogenă și nu produși de către organismul plantei ca și
fitohormonii (Petruş, 2009).
De-a lungul timpului, numeroşi cercetători au formulat reţete de mediu pentru diferite specii de
plante, în funcţie de necesităţile lor fiziologice. Momentul de răscruce în acest domeniu, l-a reprezentat
reţeta publicată de Murashige şi Skoog în 1962, care le poartă numele şi care a fost concepută iniţial
pentru cultura ţesuturilor de tutun. Ulterior a fost preluată şi pentru alte specii, fiind în prezent cea mai
utilizată reţetă de mediu. Regenerarea viţei-de-vie prin culturi de ţesuturi utilizează mediul de bază
Murashige-Skoog suplimentat cu concentraţii diferite de hormoni: 1 mg/L benzilaminopurină (BAP) +
0,5 mg/L acid indolilacetic în fazele de multiplicare şi 1,8 mg/L AIA + 0,022 mg/L kinetină la
înrădăcinarea in vitro. Aşadar, ţesuturile de viţă-de-vie cultivate pe mediu de cultură specific dispun de
nutrienţii necesari declanşării proceselor de organogeneză care conduc la regenerarea de noi indivizi.
Electroterapia a constat în cazul de faţă, în stimularea electrică în cuva de electroforeză
orizontală a porţiunilor de lăstari erbacei provenite de la plante infectate cu virusuri şi cultivarea lor
ulterioară pe medii de cultură sterile. Curentul electric a acţionat deci un timp relativ scurt, 5-15 minute
după care, explantele au parcurs etapele specifice micropropagării. Cuantificarea unor compuşi
biochimici în vitroplante pe parcursul subcultivării lor a avut ca scop aprecierea unei eventuale
influenţe a curentului asupra dezvoltării propagulilor.
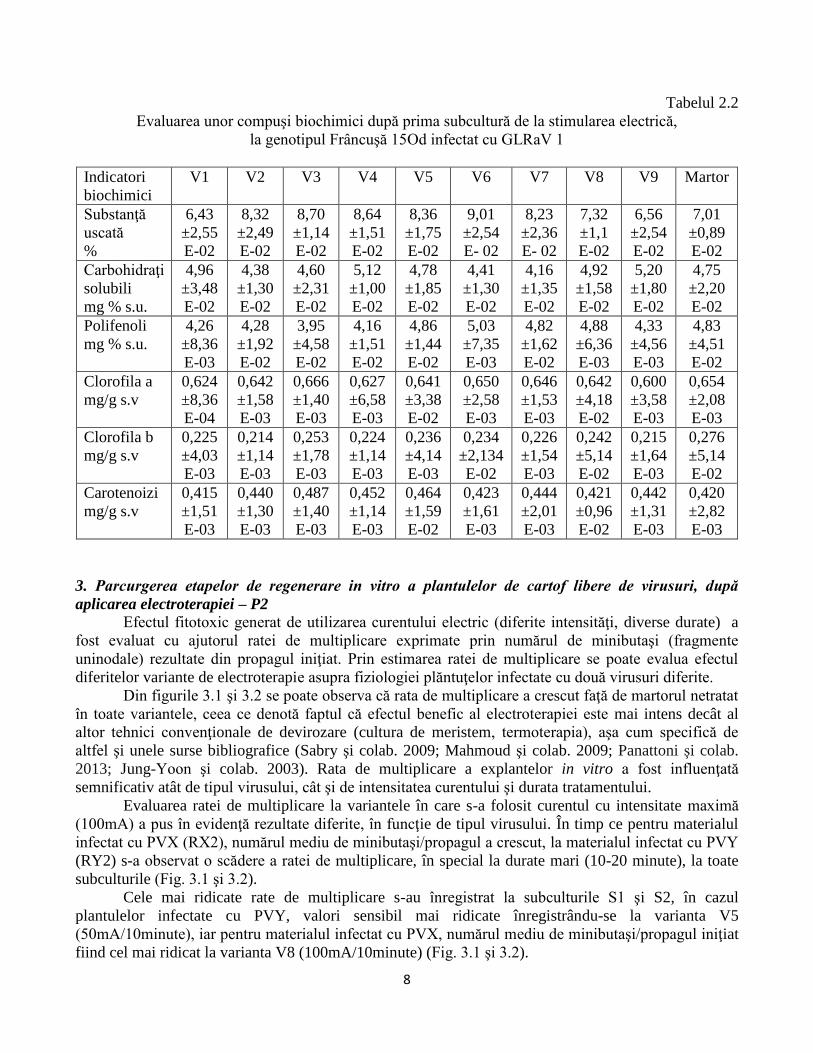
După prima subcultură post tratament electric cuantificarea unor indicatori (conţinutul în
zaharuri solubile, polifenoli totali, pigmenţi clorofilieni şi carotenoizi) nu a pus în evidenţă diferenţe
semnificative la variantele experimentale comparativ cu martorul (Tabelul 2.1 şi 2.2, Fig. 2.1 şi 2.2).
Fig. 2.1 Evoluţia subculturilor de Tămâioasă românească 3-2-2, după 60 de zile de la tratamentul
electric. De la stânga la dreapta: Martor-V3 (40mA, 15 min), Martor – V6 (50 mA, 15 min),
Martor - V9 (100 mA, 15 min)

7
Tabelul 2.1
Evaluarea unor compuşi biochimici după prima subcultură de la stimularea electrică,
la genotipul Tămâioasă românească 3-2-2 infectat cu GFkV
Indicatori
biochimici
V1 V2 V3 V4 V5 V6 V7 V8 V9 Martor
Substanţă
uscată
%
8,56
±2,55
E-02
9,42
±2,49
E-02
9,70
±1,14
E-02
9,94
±1,51
E-02
8,96
±1,75
E-02
9,05
±2,54
E- 02
8,68
±2,36
E- 02
9,12
±1,1
E-02
8,76
±2,54
E-02
9,01
±0,89
E-02
Carbohidraţi
solubili
mg % s.u.
4,75
±3,48
E-02
5,16
±1,30
E-02
4,64
±2,31
E-02
5,01
±1,00
E-02
4,58
±1,85
E-02
4,83
±1,30
E-02
4,76
±1,35
E-02
5,12
±1,58
E-02
4,95
±1,80
E-02
5,10
±2,20
E-02
Polifenoli
mg % s.u.
4,35
±8,36
E-03
3,68
±1,92
E-02
3,40
±4,58
E-02
4,26
±1,51
E-02
4,56
±1,44
E-02
4,53
±7,35
E-03
5,02
±1,62
E-02
4,85
±6,36
E-03
4,63
±4,56
E-03
4,13
±4,51
E-02
Clorofila a
mg/g s.v
0,614
±8,36
E-04
0,654
±1,58
E-03
0,646
±1,40
E-03
0,670
±6,58
E-03
0,624
±3,38
E-02
0,612
±2,58
E-03
0,646
±1,53
E-03
0,640
±4,18
E-02
0,624
±3,58
E-03
0,634
±2,08
E-03
Clorofila b
mg/g s.v
0,230
±4,03
E-03
0,254
±1,14
E-03
0,221
±1,78
E-03
0,216
±1,14
E-03
0,252
±4,14
E-03
0,231
±2,134
E-02
0,246
±1,54
E-03
0,242
±5,14
E-02
0,213
±1,64
E-03
0,249
±5,14
E-02
Carotenoizi
mg/g s.v
0,410
±1,51
E-03
0,443
±1,30
E-03
0,457
±1,40
E-03
0,480
±1,14
E-03
0,452
±1,59
E-02
0,461
±1,61
E-03
0,450
±2,01
E-03
0,418
±0,96
E-02
0,435
±1,31
E-03
0,427
±2,82
E-03
Fig. 2.2 Evoluţia subculturilor de Frâncuşă 15 Od, după 60 de zile de la tratamentul electric.
De la stânga la dreapta: Martor-V1 (40mA, 5 min), Martor - V5 (50 mA, 10 min),
Martor - V9 (100 mA, 15 min)
Diferite intensităţi ale curentului precum şi timpi de tratament diferiţi nu şi-au manifestat
acţiunea pe termen lung, indicatorii evaluaţi nu au înregistrat diferenţe semnificative comparativ cu
martorul pe parcursul celor trei subculturi (aprox. 100 zile) de multiplicare
Bibliografie
Petruş A., 2009 - Lucrări practice de biotehnologie generală. Universitatea din Oradea. [online].
Disponibil pe internet:
bioresearch.ro/.../Petrus%20A%20-%20Biotehnologie%20generala%20L.[accesat 12.11.2013]
8
Tabelul 2.2
Evaluarea unor compuşi biochimici după prima subcultură de la stimularea electrică,
la genotipul Frâncuşă 15Od infectat cu GLRaV 1
Indicatori
biochimici
V1 V2 V3 V4 V5 V6 V7 V8 V9 Martor
Substanţă
uscată
%
6,43
±2,55
E-02
8,32
±2,49
E-02
8,70
±1,14
E-02
8,64
±1,51
E-02
8,36
±1,75
E-02
9,01
±2,54
E- 02
8,23
±2,36
E- 02
7,32
±1,1
E-02
6,56
±2,54
E-02
7,01
±0,89
E-02
Carbohidraţi
solubili
mg % s.u.
4,96
±3,48
E-02
4,38
±1,30
E-02
4,60
±2,31
E-02
5,12
±1,00
E-02
4,78
±1,85
E-02
4,41
±1,30
E-02
4,16
±1,35
E-02
4,92
±1,58
E-02
5,20
±1,80
E-02
4,75
±2,20
E-02
Polifenoli
mg % s.u.
4,26
±8,36
E-03
4,28
±1,92
E-02
3,95
±4,58
E-02
4,16
±1,51
E-02
4,86
±1,44
E-02
5,03
±7,35
E-03
4,82
±1,62
E-02
4,88
±6,36
E-03
4,33
±4,56
E-03
4,83
±4,51
E-02
Clorofila a
mg/g s.v
0,624
±8,36
E-04
0,642
±1,58
E-03
0,666
±1,40
E-03
0,627
±6,58
E-03
0,641
±3,38
E-02
0,650
±2,58
E-03
0,646
±1,53
E-03
0,642
±4,18
E-02
0,600
±3,58
E-03
0,654
±2,08
E-03
Clorofila b
mg/g s.v
0,225
±4,03
E-03
0,214
±1,14
E-03
0,253
±1,78
E-03
0,224
±1,14
E-03
0,236
±4,14
E-03
0,234
±2,134
E-02
0,226
±1,54
E-03
0,242
±5,14
E-02
0,215
±1,64
E-03
0,276
±5,14
E-02
Carotenoizi
mg/g s.v
0,415
±1,51
E-03
0,440
±1,30
E-03
0,487
±1,40
E-03
0,452
±1,14
E-03
0,464
±1,59
E-02
0,423
±1,61
E-03
0,444
±2,01
E-03
0,421
±0,96
E-02
0,442
±1,31
E-03
0,420
±2,82
E-03
3. Parcurgerea etapelor de regenerare in vitro a plantulelor de cartof libere de virusuri, după
aplicarea electroterapiei – P2 Efectul fitotoxic generat de utilizarea curentului electric (diferite intensităţi, diverse durate) a
fost evaluat cu ajutorul ratei de multiplicare exprimate prin numărul de minibutaşi (fragmente
uninodale) rezultate din propagul iniţiat. Prin estimarea ratei de multiplicare se poate evalua efectul
diferitelor variante de electroterapie asupra fiziologiei plăntuţelor infectate cu două virusuri diferite.
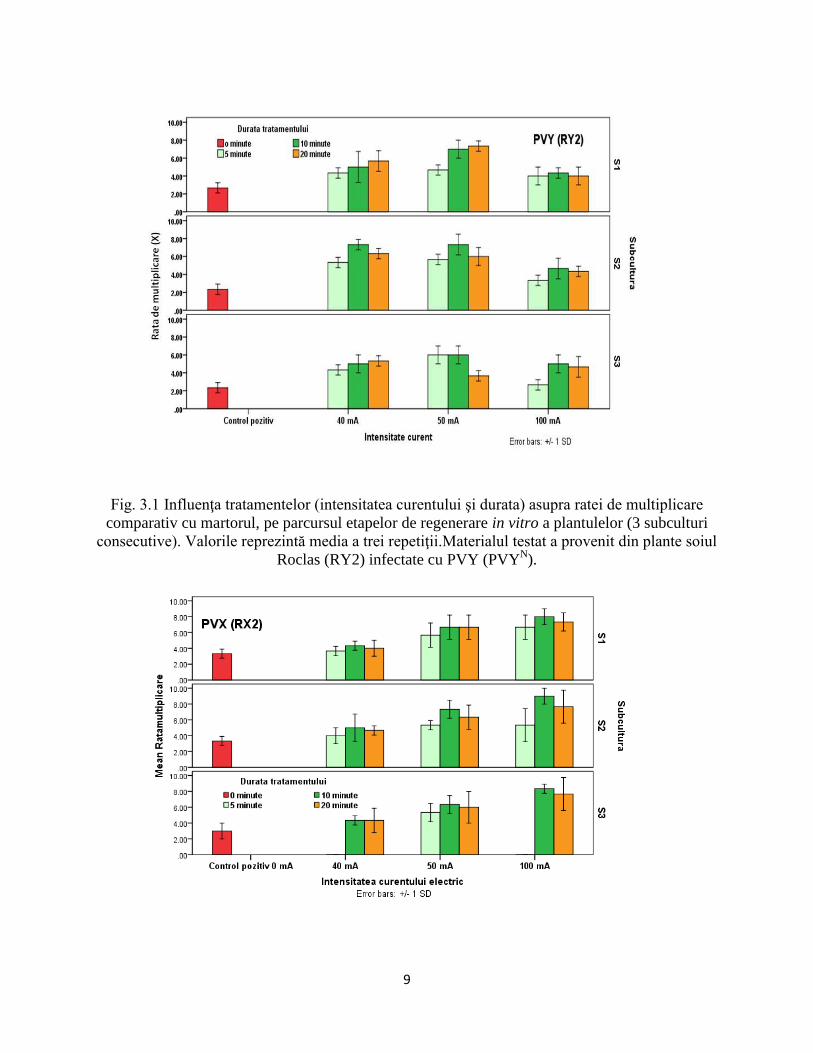
Din figurile 3.1 şi 3.2 se poate observa că rata de multiplicare a crescut faţă de martorul netratat
în toate variantele, ceea ce denotă faptul că efectul benefic al electroterapiei este mai intens decât al
altor tehnici convenţionale de devirozare (cultura de meristem, termoterapia), aşa cum specifică de
altfel şi unele surse bibliografice (Sabry şi colab. 2009; Mahmoud şi colab. 2009; Panattoni şi colab.
2013; Jung-Yoon şi colab. 2003). Rata de multiplicare a explantelor in vitro a fost influenţată
semnificativ atât de tipul virusului, cât şi de intensitatea curentului şi durata tratamentului.
Evaluarea ratei de multiplicare la variantele în care s-a folosit curentul cu intensitate maximă
(100mA) a pus în evidenţă rezultate diferite, în funcţie de tipul virusului. În timp ce pentru materialul
infectat cu PVX (RX2), numărul mediu de minibutaşi/propagul a crescut, la materialul infectat cu PVY
(RY2) s-a observat o scădere a ratei de multiplicare, în special la durate mari (10-20 minute), la toate
subculturile (Fig. 3.1 şi 3.2).
Cele mai ridicate rate de multiplicare s-au înregistrat la subculturile S1 şi S2, în cazul
plantulelor infectate cu PVY, valori sensibil mai ridicate înregistrându-se la varianta V5
(50mA/10minute), iar pentru materialul infectat cu PVX, numărul mediu de minibutaşi/propagul iniţiat
fiind cel mai ridicat la varianta V8 (100mA/10minute) (Fig. 3.1 şi 3.2).
9
Fig. 3.1 Influenţa tratamentelor (intensitatea curentului şi durata) asupra ratei de multiplicare
comparativ cu martorul, pe parcursul etapelor de regenerare in vitro a plantulelor (3 subculturi
consecutive). Valorile reprezintă media a trei repetiţii.Materialul testat a provenit din plante soiul
Roclas (RY2) infectate cu PVY (PVYN).

10
Fig 3.2 Influenţa tratamentelor (intensitatea curentului şi durata) asupra ratei de multiplicare
comparativ cu martorul, pe parcursul etapelor de regenerare in vitro a plantulelor (3 subculturi
consecutive, cu excepţia variantelelor V1 şi V7. Valorile reprezintă media a trei repetiţii. Materialul
testat a provenit din plante soiul Roclas (RX2) infectate cu PVX.
Bibliografie
Clark M.F. and Adams A.N., 1977 - Characterization of the microplate method of the enzyme-linked
immunosorbent assay for the detection of plant virus. J. Gen.Virol. 34, 475-483.
Lozoya-Saldana H., Abello F.J., Garcia G.R.,1996 - Electrotherapy and shoot tip culture eliminate
potato virus X in potatoes. Am. J. Potato Res. 73, 149-154
Jung-Yoon Y., Hyo-won S., Young-Mee C., Young-Eun P., 2003 - Ribavirin, electric current and
shoot tip culture to eliminate several potato viruses. Journal of Plant Biotechnology 5, 101-105.
Panattoni A., Luvisi A., Triolo E., 2013 - Elimination of viruses in plants: twenty years of progress.
Span. J. Agric. Res. 11(1), 173–178.
Sabry Y.M. Mahmoud, Maher H. Hosseny and Mamdouh H. Abdel-Ghaffar, 2009 - Evaluation of
some therapies to eliminate Potato Y Potyvirus from potato plants, Int. J. Virol. 5, 64-76
Svetleva D., Velcheva M., Bhowmik G., 2003 - Biotechnology as a useful tool in common bean
(Phaseolus vulgaris L.) improvement. Euphytica 131, 189-200.
4. Evaluarea acţiunii curentului electric asupra dezvoltării plantulelor de cartof pe parcursul
subculturilor prin cuantificarea unor compuşi biochimici – P2
Cuantificarea unor compuşi biochimici în plantulele de cartof supuse electroterapiei a urmărit
evaluarea unor potenţiale efecte fitotoxice ale curentului electric (în special în cazul variantelor celor
mai severe 100mA/10-20minute) asupra dezvoltării explantelor, în vederea alegerii variantei optime,
care să permită nu numai supravieţuirea plantelor, dar şi eliminarea infecţiei virale.
Dozarea amidonului din plantulele de cartof supuse electroterapiei a fost efectuată conform
metodei spectrofotometrice cu antrona (Pena Valdivia, 1991). S-a folosit 1 gram material vegetal
prelevat de la trei frunze ale aceleiaşi plante, materialul a fost mărunţit şi omogenizat în 10ml etanol
80% (v/v). Rezultatele testelor au evidenţiat o dependenţă direct proporţională a concentraţiilor acestor
compuşi biochimici, cu intesitatea curentului electric şi implicit durata de expunere la curent, pentru
ambele tipuri de materiale virozate (RX2, RY2). Comparativ cu martorul netratat, valorile medii ale
concentraţiilor acestor compuşi au crescut în toate variantele experimentale, în special în cazul
plantelor infectate cu virusul Y al cartofului. După trei subculturi consecutive, la explantele infectate cu ambele virusuri s-a înregistrat o
scădere a conţinutului de proteină brută, în special în cazul variantelor mai severe (V7,V8,V9) şi mai
ales la materialul infectat cu virusul Y. Probabil, intensitatea ridicată a curentului a denaturat o parte
din proteinele plantei şi de aceea, în aceste variante şi rata de multiplicare a fost mai scăzută. La toate
plantulele testate, provenite din subcultura a doua (S2) s-au înregistrat valori descrescătoare ale
conţinutului de proteină brută (% SU), odată cu intensificarea condiţiilor tratamentelor aplicate, în timp
ce conţinutul de amidon (% SU) a crescut (Tabel 4.1 şi 4.2).
Bibliografie
Peña Valdivia C. B., Ortega-Delgado M. L., 1991- Non structural carbohydrate partitioning in
Phaseolus vulgaris after vegetative growth. J. Sci. Food Agric. 55, 563-577.

11
Tabelul 4.1.
Efectul tratamentelor prin electroterapie asupra conţinutului de amidon şi proteină brută la plantulele de
cartof infectate artificial cu PVX (RX2), pe parcursul subculturilor S1, S2, S3.
Varianta Tratament
mA/min
Conţinut amidon % SU Conţinut proteină brută % SU
S1 S2 S3 S1 S2 S3
V0 0/0 5,6 6,8 6,6 55,9 57,4 58,1
V1 40/5 6 7 - 51,2 52 -
V2 40/10 5,8 7,1 6,8 49,2 48,2 47,1
V3 40/20 6,2 6,7 6,4 45,2 44,4 45,1
V4 50/5 7,5 8 7,8 50,5 49,7 50,1
V5 50/10 7,1 7,4 7 47,1 46 46,8
V6 50/20 7,8 8,6 8,2 44,9 43,8 44,7
V7 100/5 8,4 9 - 47,2 42,6 -
V8 100/10 9 9,4 8,8 48,1 43,8 45,7
V9 100/20 9,5 9,7 9,1 43,1 41,8 44,4
Tabelul 4.2
Efectul tratamentelor prin electroterapie asupra conţinutului de amidon şi proteină brută la plantulele de cartof
infectate artificial cu PVY (RY2), pe parcursul subculturilor S1, S2, S3.
Varianta Tratament
mA/min
Conţinut amidon % SU Conţinut proteină brută %SU
S1 S2 S3 S1 S2 S3
V0 0/0 5,2 5,7 6,4 70,9 72 70,4
V1 40/5 5 5,7 5,4 68,2 67,1 68
V2 40/10 5,8 6,3 6 64 63,2 64,2
V3 40/20 5,9 6,7 6,4 63,7 64,7 62,4
V4 50/5 6,7 5,7 5,8 65 63 64,1
V5 50/10 6,4 6,3 7 62 64 65,2
V6 50/20 6,9 7,4 7,5 60,4 58,7 58
V7 100/5 7,7 8 7,9 58,7 56,7 56,1
V8 100/10 8,1 8,4 8,5 56,7 55,5 56,7
V9 100/20 8,7 9 9,4 52 50,7 49,7
5. Modelarea efectului câmpului electric de înaltă tensiune asupra proceselor fiziologice din plante –
CO

12
Modelul de transport membranar prezentat mai jos stă la baza explicării efectului câmpului
elecric de înaltă tensiune asupra celulelor vegetale. Aplicarea unui câmp electric intens în apropierea
materialului vegetal duce, prin efect corona, la apariţia în apropierea membranei celulare a unor
distribuţii de sarcină electrică spaţială.
Aceste distribuţii de sarcină electrică determină câmpuri electrice locale de mare intensitate.
Apariţia rapidă a acestor structuri modifică condiţiile pe frontiera membranei celulare. Aceasta face ca
tensiunea transmembranară şi curentul transmembranar să fie diferite de cele corespunzătoare stării
staţionare. Noile valori ale celor doi parametrii constituie starea inţială a S.D.-ului asociat celulei. În
funcţie de starea iniţială şi de parametrii specifici, celula evoluează pe o anumită traiectorie, în general,
spre starea staţionară. Există situaţii în care celula se îndepartează indefinit de starea de echilibru.
Aceasta înseamnă de fapt ruperea membranei celulare şi moartea acesteia.
Dacă privim membrana celulară doar din punct de vedere electric, ea poate fi descrisă ca un
sistem dinamic bidimensional în care variabilele independente sunt diferenţa de potenţial
transmembranară şi intensitatea curentului electric total ce străbate membrana. Din considerente
experimentale de dată relative recentă, propunem pentru circuitul electric echivalent schema din figura
1.
Deoarece pompele asigură un anumit curent şi la potenţialul de echilibru, curentul injectat de
acestea se poate reprezenta printr-o funcţie de forma:
Deoarece f este o funcţie continuă şi derivabilă, această funcţie poate fi dezvoltată în serie de puteri în
jurul lui V0. Oprindu-ne la aproximaţia de ordinul trei, obţinem:
Deoarece pentru tensiuni de membrană mai mari decât V0 sistemul tinde să refacă potenţialul de
echilibru, rezultă că a<0. Din condiţia:
rezultă b=0. Deci aproximaţia de ordinal trei a funcţiei f este:
Pe baza consideraţiilor biofizice expuse mai sus, prima ecuaţie ce descrie sistemul dinamic
poate fi derivată direct din circuitul electric echivalent al membranei celulare (fig. 1).
Pentru aflarea celei de a doua ecuaţii diferenţiale, pornim de la forma canonică pe care trebuie
să o îndeplinească aceasta:
Din păcate, nu se cunoaşte forma exactă a funcţiei g(V,I). Însă, se pot face unele consideraţii de natură
generală asupra proprietăţilor acesteia. Să introducem, în locul variabilelor V şi I, două mărimi
adimensionale. Pentru a realiza acest lucru, se pot împărţi cele două variabile la două constante
intrinseci sistemului. Fie acestea, potenţialul de echilibru al membranei celulare V0, respectiv I0
curentul injectat de pompele ionice la echilibru:
Fig. 1 Schema electrică echivalentă a transportului transmembranar
care ia în consideraţie şi transportul activ

13
Variabilele adimensionale au avantajul obţinerii unor informaţii cu un grad mai mare de
generalitate deoarece parametrii V0 şi I0 sunt dependenţi de tipul de celulă pe când variabilele
adimensionale nu. Ca urmare a celor spuse anterior, scriem sistemul de ecuaţii folosind variabilele
adimensionale: ( )
[ √ ]
√
1
unde constantele A, B, C, D, E, F, G, H, I şi J sunt parametrii celulari.
Rezolvarea S.D.-ului se reducela determinarea portretului de fază al E.D.O..Prima etapă în
rezolvarea problemei este determinarera punctelor de echilibru ale S.D.-ului neliniar. Pentru aceasta
trebuie rezolvat sistemul de ecuaţii: ( )
[ √ ]
√
Pornind de la natura fizică a problemei, ştim că acest sistem trebuie să admită soluţia (0, 0). Cea de a
doua ecuaţie acceptă în mod evident această soluţie. Prima ecuaţie, conţinând termeni liberi, nu admite
în general, această soluţie. Pentru a se realiza această soluţie, trebuie îndeplinită următoarea relaţie între
parametrii de sistem:
Folosind această restricţie, E.D.O. se rescrie astfel:
[ √ ]
√
2
Se pune problema dacă E.D.O. mai prezintă şi un alt punct de echilibru. Cea de a doua ecuaţie se
anulează dacă:
√
Dacă este îndeplinită condiţia: ,
atunci soluţiile:
√
√
sunt reale. Pentru ca aceste soluţii să aibă sens fizic, trebuie ca valoarea lor să fie pozitivă.
Din prima ecuaţie a sistemului 2, rezultă:
[ ]
Introducând acest rezultat în soluţia de mai sus, rezultă:
{ [
[ ]]
}
√
Pentru fiecare valoare a lui v există o valoare a lui i calculată cu ecuaţia precedentă astfel încât numărul
punctelor de echilibru este maxim de patru plus (0, 0).
Odată determinate punctele de echilibru, următoarea etapă în analiza S.D. –ului este
determinarea naturii punctelor de echilibru.Pentru analiza punctului de echilibru (0, 0), calculăm
expresia Jacobianului în acest punct şi rezolvăm ecuaţia caracteristică:
Aceasta are douăvalorile proprii. Partea reală a celei de a doua soluţii este întotdeauna negativă. Pentru
ca partea reală a primei soluţii să fie negativă, trebuie ca . În aceste condiţii,partea reală a
soluţiilor ecuaţiei caracteristice este întotdeauna negativă, şi punctul de echilibru (0,0) este stabil.În

14
privinţa celorlalte puncte de echilibru, deoarece acestea se obţin dintr-o ecuaţie transcendentă, valoarea
lor exactă nu poate fi calculată. Singura metodă este cea numerică. Aceste puncte de echilibru
corespund la două valori pozitive ale lui v şi două valori negative ale lui v. Să notăm aceste valori cu
, . Se constată imediat că valorile curenţilor corespunzătoare primelor două
tensiuni sunt întotdeauna pozitive ( ). În mod analog, pentru ultimele două valori ale tensiunii,
valorile curenţilor sunt întotdeauna negative ( ). Rezolvând ecuaţia caracteristică pentru aceste
puncte, constatăm că: . Adică, punctele de echilibru altele decât originea sunt
puncte de şa.
Simularea numerică se face folosind softuri specializate. Noi folosim mediul de lucru
MATLAB 2010b.Pentru început vom simula comportamentul celulei pentru cazul în care toţi
parametrii celulari sunt mai mari ca zero. Aceasta înseamnă că doar originea este punct de echilibru.
a) Pentru ca originea să fie nod atractiv nedegenerattrebuie ca H să îndeplineasă condiţia:
[ ( )
].
Alegând parametrii A=1, B=1, C=1, D=1, E=1, G=1, I=1, J=1, putem alege pentru H valoarea
0,1.Modelarea numerică în acest caz este prezentată în fig.4.
b) Pentru ca originea să fie nod atractiv degenerat, alegând parametrii A=1, B=1, C=1, D=1,
G=1, I=1, J=1,pentru a realiza condiţiile de degenerare, parametrul E trebuie să aibă valoarea:
Modelarea numerică în acest caz este prezentată în fig.5. Se observă proprietatea de tangenţă a tuturor
orbitelor în apropierea originii.
c) Pentru ca originea să să fie punct de şa, trebuie ca H>0. Alegând valorile celorlalţi parametrii
egale cu 1, putem alege, ca exemplu, pe H=1. Reprezentarea grafică este dată în figura 6.
d) Pentru ca Originea să fiefocar atractiv,alegem parametrii celulari astfe:A=1, B=1, C=1, D=1,
G=1, I=1, J=1. H trebuie să îndeplinească restricţia:
( )
.
Fig. 4 Portretul de fază în cazul
nodului atractiv
Fig. 7 Portretul de fază pentru focar atractiv Fig. 8 Portretul de fază pentru centru
Fig. 6 Portretul de fază pentru punctul
de şa
Fig. 5 Portretul de fază pentru
nodul atractiv degenerat

15
Rezultăcăîlputemalegepe H=-3.
e) Pentru ca Originea să fiecentru,alegem parametrii celulari astfel A=1, B=0, C=1, D=1, E=0,
G=1, H=-3, I=1, J=1. Constatăm apariţia unor orbite eliptice în spaţiul fazelor. Aceasta înseamnă o
evoluţie periodică a sistemului dinamic.
Vom selecta coeficienţii sistemului astfel încât să îndeplinească cerinţele necesare apariţiei şi a
altor puncte de echilibru decât originea. Din condiţia: ,
rezultă restricţia:
| | √| |
Dacă J este pozitiv, criteriul de apariţie al punctelor de echilibru se rescrie astfel:
√
Această restricţie este îndeplinită numai dacă I>0. Din cele două inegalităţi rezultă condiţia de apariţie
a punctelor de echilibru altele decât originea:
>2√ .
Alegem parametrii celulari astfel: A=1, B=1, C=1, D=1, E=1, G=1, H=-1, I=2, J=1. Se constată
comportamentul fazelor în apropierea punctelor de echilibru instabil tipică punctelor de tip şa: iniţial
faza se apropie de punctul de echilibru, apoi se îndepărtează. Se constată că există şi puncte
reprezentative care, după apropierea de punctul de echilibru instabil, se îndepărtează nedefinit de
origine. Astfel de orbite sunt reprezentate în figura10.
6. Monitorizarea plantelor de viţă-de-vie şi cartof regenerate sănătoase prin chimioterapie, privind
desfăşurarea proceselor fiziologice – CO
Plantele de viţă-de-vie regenerate sănătoase prin chimioterapie in vitro cu ribavirină şi
oseltamivir au fost urmărite în perioada de vegetaţie în ceea ce priveşte desfăşurarea proceselor
fiziologice. Determinările au fost efectuate în seră, la o valoare a intensităţii luminii de 20 000 lucşi,
temperatura de 28oC şi umiditatea relativă a aerului de 52%. Intensitatea fotosintezei şi respiraţiei
(µmol CO2/m2/s) s-a realizat cu ajutorul analizorului de dioxid de carbon S151. Determinarea
suprafeţei foliare s-a realizat cu ajutorul metodei mulajelor. Determinarea conţinutului în apă (%) la
nivelul frunzelor s-a realizat prin uscare la temperatura camerei (până la masă constantă), apoi
materialul vegetal a fost uscat, cu ajutorul termobalanţei, la 100-105oC.
În ceea ce priveşte influenţa viricidelor asupra dimensiunii aparatului foliar, s-a constatat o
valoare apropiată de martor în cazul variantei V2. Rezultate semnificativ mai mari s-au determinat
pentru variantele V1 şi V3 (Fig. 6.1a ). Suprafaţa foliară medie/plantă a înregistrat valori semnificativ
mai mari faţă de martor în cazul tuturor variantelor influenţate (Fig. 6.1b). Numărul de frunze/plantă nu
a fost influenţat în mod semnificativ de concentraţiile de viricid utilizate (Fig. 6.1c).
Determinarea intensităţii respiraţiei (la întuneric) la nivelul frunzelor viţelor libere de virus, a
evidenţiat valori scăzute în cazul variantei V3, în comparaţie cu varianta V2. Între martor şi variantele
V1 şi V2, diferenţele nu sunt semnificative (Fig.6.1d).
Fig 10 Portret de faze în jurul punctelor de echilibru
instabil cu orbite ce se îndepărtează
de origine
Fig 9 Portret de faze în jurul punctelor de echilibru
instabil cu orbite ce se îndreaptă spre origine

16
Determinarea intensităţii fotosintezei evidenţiază influenţa negativă a concentraţiei de 40mg/l
ribavirină, în combinaţie cu 40mg/l oseltamivir, faţă de martor. Între variantele influenţate (V1, V2 şi
V3), diferenţele înregistrate nu sunt semnificative (Fig. 6.1e).
Fig. 6.1 Determinări fiziologice la plantele de viţă-de-vie regenerate sănătoase prin chimioterapie in
vitro. Reprezentarea grafică şi interpretarea statistică a rezultatelor s-a realizat cu ajutorul programului
SPSS 16,0 for Windows. S-a aplicat testul Duncan pentru comparaţii multiple (literele diferite indică
diferenţe semnificative, pentru pragul de semnificaţie p<0,05).
Conţinutul în apă liberă determinată la nivelul frunzelor viţelor din varianta V1 a fost
semnificativ mai mic faţă de valorile înregistrate la martor, V2 şi V3 (Fig. 6.1f).
Conţinutul în apă legată pentru variantele V1, V2 şi V3 a fost semnificativ mai scăzut în
comparaţie cu martorul (Fig. 6.1g). Conţinutul în apă totală la V1 şi V3 nu a prezentat diferenţe
semnificative faţă de martor, o scădere semnificativă înregistrându-se doar la V1 (Fig 6.1h).

17
Calcularea coeficientului de corelaţie simplă Pearson şi a semnificaţiei acestuia a evidenţiat o
scădere semnificativă a intensităţii fotosintezei şi a procentului de apă totală odată cu creşterea
concentraţiei de ribavirină (Tabelul 6.1).
Tabelul 6.1
Corelaţia dintre indicii fiziologici şi cantitatea de viricid
Pentru monitorizarea plantelor de cartof provenite prin electroterapie au fost alese plantule
obţinute din explante provenite de la materialul biologic RX1, RX2 şi RY1 (plante aclimatizate în
condiţii de seră după subcultura 3, după 32 zile de vegetaţie pentru RX2 şi după 28 zile de vegetaţie
pentru restul plantelor).
Monitorizarea stării de vegetaţie a plantelor regenerate sănătoase s-a realizat prin determinarea
conţinutului de clorofilă la nivel de frunză cu ajutorul dispozitivului portabil SPAD 502 (Chlorophyll
Meter). Valorile determinate de acest instrument indică suma relativă a cantităţii de clorofilă prezentă
Corelatia dintre indicatorii fiziologici si concentratia de ribavirina/oseltamivir
-.672* -.427
.017 .167
12 12
-.083 -.536
.798 .072
12 12
.560 .734**
.058 .007
12 12
.938** .551
.000 .063
12 12
-.263 .263
.410 .410
12 12
-.847** -.304
.001 .337
12 12
-.456 -.975**
.137 .000
12 12
-.877** -.556
.000 .061
12 12
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Coef icientul de corelatie Pearson
Pragul de semnif icat ie
N
Intensitatea
f otosintezei
Intensitatea
respiratiei
Supraf ata
f oliara
totala/planta
Supraf ata
f oliara medie
Numar de
f runze/planta
Apa libera
Apa legata
Apa totala
Concentratia
ribav irina
Concentratia
oseltamiv ir
Corelatia este semnif icativa pentru p<0,05*.
Corelatia este semnif icativa pentru p<0,01**.
18
în frunzele plantei, măsurată prin transmitanţa frunzei la două lungimi de undă, 650 nm (roşu) şi 940
(infraroşu apropiat - NIR).
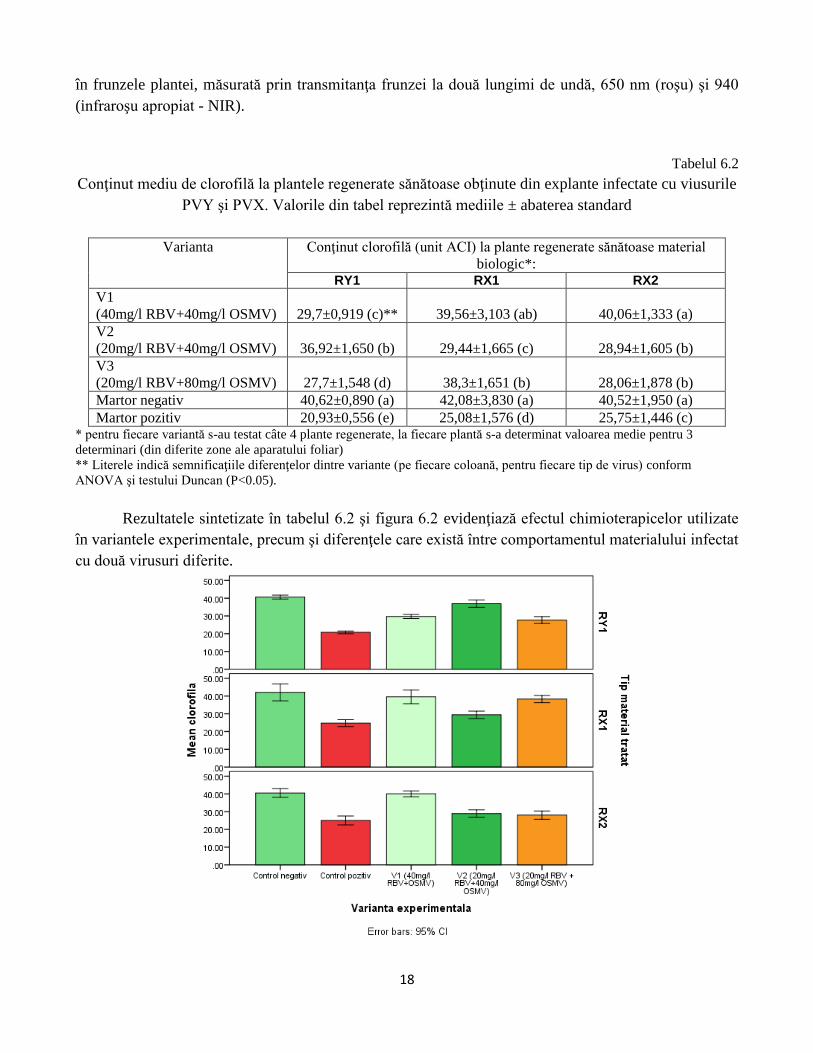
Tabelul 6.2
Conţinut mediu de clorofilă la plantele regenerate sănătoase obţinute din explante infectate cu viusurile
PVY şi PVX. Valorile din tabel reprezintă mediile ± abaterea standard
Varianta Conţinut clorofilă (unit ACI) la plante regenerate sănătoase material
biologic*: RY1 RX1 RX2
V1
(40mg/l RBV+40mg/l OSMV) 29,7±0,919 (c)** 39,56±3,103 (ab) 40,06±1,333 (a)
V2
(20mg/l RBV+40mg/l OSMV) 36,92±1,650 (b) 29,44±1,665 (c) 28,94±1,605 (b)
V3
(20mg/l RBV+80mg/l OSMV) 27,7±1,548 (d) 38,3±1,651 (b) 28,06±1,878 (b)
Martor negativ 40,62±0,890 (a) 42,08±3,830 (a) 40,52±1,950 (a)
Martor pozitiv 20,93±0,556 (e) 25,08±1,576 (d) 25,75±1,446 (c) * pentru fiecare variantă s-au testat câte 4 plante regenerate, la fiecare plantă s-a determinat valoarea medie pentru 3
determinari (din diferite zone ale aparatului foliar)
** Literele indică semnificaţiile diferenţelor dintre variante (pe fiecare coloană, pentru fiecare tip de virus) conform
ANOVA şi testului Duncan (P<0.05).
Rezultatele sintetizate în tabelul 6.2 şi figura 6.2 evidenţiază efectul chimioterapicelor utilizate
în variantele experimentale, precum şi diferenţele care există între comportamentul materialului infectat
cu două virusuri diferite.

19
Fig. 6.2 Influenţa tratamentelor chimioterapice asupra dezvoltării plantelor regenerate sănătoase.
Conţinutul de clorofilă (unităţi ACI) determinat cu dispozitivului portabil SPAD 502 (Chlorophyll
Meter) la plante provenite din material biologic infectat cu virusurile PVY şi PVX şi regenerate
după 3 subculturi consecutive (două cu viricide şi unul MS) şi aclimatizat în condiţii de seră.
Barele de eroare reprezintă intervalul de confidenţă al erorilor (CI).
Diferenţele mai mult sau mai puţin semnificative privind anumiţi indicatori fiziologici la
plantele de viţă-de-vie şi cartof, regenerate libere de virusuri în urma tratamentelor cu chimioterapice,
nu pot fi puse strict pe seama concentraţiilor de viricide sau a timpului de tratament, deoarece, fiecare
plantă constituie un individ singular şi complex, care se dezvoltă independent în funcţie de condiţiile de
mediu sau de genotip. Este cunoscut faptul că, procesele de fotosinteză şi respiraţie sunt influenţate
direct de temperatură, umiditate, intensitate luminoasă la nivelul fiecărui individ. Pe de altă parte,
genotipul este răspunzător de dezvoltarea aparatului foliar la plantele regenerate de viţă-de-vie şi cartof.
Important este faptul că, tratamentele cu viricide au condus la obţinerea de plante libere de
virusuri. Ca urmare, a fost atins unul din obiectivele proiectului şi anume, au fost elaborate tehnologii
de eliminare a virusurilor la viţa-de-vie şi cartof prin chimioterapie in vitro cu ribavirină şi oseltamivir.
Proiectul îşi propune în continuare experimentarea aplicării curentului electric pentru
inactivarea proteinei virale, precum şi experimentarea terapiilor combinate, electroterapie urmată de
chimioterapie, în vederea îmbunîtîţirii ratei de eliminare virală. Analiza economică a tehnologiilor va
decide care dintre metodele propuse este cea mai eficientă şi va conduce la brevetarea acesteia.
I. Tehnologie de eliminare a virusului fleck la viţa-de-vie prin chimioterapie in vitro cu ribavirină şi
oseltamivir
1. Identificarea infecţiei virale;
2. Cultivarea in vitro a fragmentelor de lăstari erbacei proveniţi de la plantele infectate, pe mediu de
cultură specific viţei-de-vie (Murashige şi Skoog, 1962; Vişoiu şi Teodorescu, 2001), suplimentat cu
două chimioterapice, ribavirină şi oseltamivir:
- recoltarea lăstarilor, fragmentarea în minibutaşi uninodali, dezinfecţia cu hipoclorit de calciu 6-10%
timp de 4 min;
- inocularea pe mediu agarizat suplimentat cu 20 mg/L ribavirină şi 40 mg/L oseltamivir, în condiţii
sterile la hota cu flux de aer laminar;
- incubarea timp de 30 de zile pe mediu cu viricide, urmată de o subcultură pe mediu fără
chimioterapice, în camere de creştere la temperatura de 24 ± 1C, fotoperioada 16 ore lumină şi
iluminarea 3000-3500 lx;
- înrădăcinarea in vitro pe mediu specific viţei-de-vie;
3. Aclimatizarea şi fortificarea plantelor regenerate (90 de zile).
4. Analiza virologică a plantelor regenerate în vederea selecţiei plantelor libere de virus.
II. Tehnologie de eliminare a virusurilor X, izolatele 1 şi 2 (PVX1, PVX2), Y , izolatele 1 şi 2 (PVY1,
PVY2) ale cartofului prin chimioterapie in vitro cu ribavirină şi oseltamivir
1. Identificarea infecţiei virale;
2. Iniţierea culturilor prin detaşarea colţilor de cartof de 2-3 cm lungime, sterilizarea cu clorură
mercurică 0,5% timp de 3 minute, inocularea pe mediul de cultură specific cartofului (Murashige şi
Skoog, 1962; Nistor şi colab., 2010) şi incubarea în camere de creştere la temperatura de 24 ± 1C,
fotoperioada 16 ore lumină şi iluminarea 3000-3500 lx;

20
3. Butăşirea plantulelor şi cultivarea acestora pe mediu de cultură specific suplimentat cu viricide:
ribavirină 40mg/L + oseltamivir 40 mg/L timp de 2 subculturi (60 zile);
4. Butăşirea plantulelor tratate cu chimioterapice şi cultivarea lor pe mediu fără viricide;
5. Aclimatizarea şi fortificarea plantelor regenerate (90 de zile).
6. Analiza virologică a plantelor regenerate în vederea selecţiei plantelor libere de virus.
Bibliografie
Murashige T. şi Skoog F., 1962 - A revised medium for rapid grow and bioassays with Tabacco tissue
culture. Physiol. Plant. 15, 473-49
Nistor A., Câmpeanu G., Atanasiu N., Chiru N., Caracsoniy D., 2010- Influence of potatto genotypes
on in vitro production of microtubers. Romanian Biotechnological Letters 15 (3), 5317-5324
Vişoiu E. şi Teodorescu Al., 2001 – Biotehnologii de producere a materialului săditor viticol. Ed.
Ceres, Bucureşti.
7. Diseminare rezultate – CO, P1, P2
Rezultatele cercetărilor obţinute până în această fază de derulare a proiectului au fost
valorificate prin publicarea următoarelor lucrări ştiinţifice:
Topală CM (2013). Temperature Effects On The FTIR Spectra Of Ribavirin. Rev Chim 64(2): 132-135.
Topală C., Oprescu B., Tătaru L., 2013 - FT/IR study on changes induced by chemotherapy and
electrotherapy application in grapevine. Analele Universităţii din Craiova, seria Biologie,
Horticultură, tehnologia prelucrării produselor agricole, Ingineria mediului. ISSN 1453-1275,
VIII (LIV), 333-338
Bădărău C.L., Chiru N., Guţă I.C., 2013 - Effect of some chemotherapies on potato virus Y and X
infected Solanum tuberosum L. plantlets (cv. Roclas. Proceedings of the Fourth International
Conference, Research People and actual tasks on multidisciplinary sciences, Lozenec, Bulgaria,
12-16 June, 2013, ISSN 1313-7735, 1, 69-74.
Bădărău C.L., Chiru N., Damşa F., Guţă I.C., 2013 - Effect of some chemotherapies on potato virus Y
and X infected Solanum tuberosum L. microplants (preliminary studies). Volum de rezumate Al
V-lea Simpozion de Etnofarmacologie cu participare internaţională “Etnofarmacologia în
sprijinul sănătăţii omului şi mediului”, 21-23 iunie Braşov ISSN 1844-6604, 65.
Bădărău C.L., Chiru N., Guţă I.C., 2013 - Effect of some chemo- and electrotherapies on potato virus Y
and X infected Solanum tuberosum L. plantlets (cv. Roclas). Proceedings of the EAPR
Conference Pathology Section Meeting Climate Change/Global Warming: Effects on Potato
Diseases/Pests.16-22 November, Ierusalem, Israel, 46.
Guţă I.C., Buciumeanu E-C., Vişoiu E. - Elimination of grapevine fleck virus by in vitro chemotherapy.
Lucrare în curs de evaluare la revista Notulae Botanicae Horti Agrobotanici Cluj-Napoca.
Director proiect
Conf. univ.dr. Carmen Mihaela TOPALĂ