ing. tiberia ioana pop - usamv cluj · universitatea de ȘtiinȚe agricole Și medicinĂ...
TRANSCRIPT

UNIVERSITATEA DE ȘTIINȚE AGRICOLE ȘI MEDICINĂ
VETERINARĂ
CLUJ-NAPOCA
ȘCOALA DOCTORALĂ
FACULTATEA DE HORTICULTURĂ
Ing. TIBERIA IOANA POP
REZUMATUL TEZEI DE DOCTORAT
CONTROLUL GENETIC AL FORMĂRII RĂDĂCINILOR
ADVENTIVE- UN FACTOR LIMITATIV ÎN PROPAGAREA
VEGETATIVĂ
CONDUCĂTOR ȘTIINȚIFIC
Prof. Univ. Dr. DORU PAMFIL
CLUJ-NAPOCA
2011

2
CUPRINS
INTRODUCERE........................................................................................................... 3
I. IMPORTANȚA STUDIULUI RĂDĂCINILOR ADVENTIVE ............................. 5
1.1. IMPORTANŢA COMERCIALĂ A RĂDĂCINILOR ADVENTIVE PENTRU
AGRICULTURĂ, HORTICULTURĂ ȘI SILVICULTURĂ................................................... 5
1.2. IMPORTANȚA RĂDĂCINILOR ADVENTIVE PENTRU PROGRAMELE DE
CONSERVARE A BIODOVERSITĂȚII ............................................................................... 6
II. BIOLOGIA RĂDĂCINILOR ADVENTIVE .......................................................... 7
2.1. DESCRIEREA ȘI CARACTERIZAREA RĂDĂCINILOR ADVENTIVE ........... 7
2.2. FACTORI CARE INFLUENȚEAZĂ FORMAREA RĂDĂCINILOR
ADVENTIVE ........................................................................................................................ 8
III.STUDIUL FORMĂRII RĂDĂCINILOR ADVENTIVE ....................................... 9
3.1. ARABIDOPSIS THALIANA - PLANTĂ MODEL ÎN STUDIUL FORMĂRII
RĂDĂCINILOR ADVENTIVE ............................................................................................. 9
3.1.1. The Arabidopsis Information Resource (TAIR) .................................................... 11
3.2. SUPRESIA GENEI SUPPERROOT- STRATEGIE ÎN IDENTIFICAREA DE
GENE IMPLICATE ÎN FORMAREA RĂDĂCINILOR ADVENTIVE ................................ 11
3.3. MUTAGENEZA CU EMS ................................................................................ 12
3.4. CARTAREA MUTAȚIILOR ............................................................................. 13
3.4.1. Tipuri de markeri utilizați în cartare....................................................................... 13
3.4.2. Procedeul de cartare fină .......................................................................................... 14
IV.SCOPUL ȘI OBIECTIVELE TEZEI DE DOCTORAT ...................................... 15
V. MATERIALE ȘI METODE .................................................................................. 16
5.1. MATERIALE ȘI METODE UTILIZATE PENTRU ÎNRĂDĂCINAREA IN
VITRO A SPECIILOR DE DIANTHUS ................................................................................. 16
5.2. MATERIALE ȘI METODE PENTRU CARACTERIZAREA MUTANȚILOR
SUPRESORI SUPERROOT2 LA ARABIDOPSIS THALIANA .............................................. 17
VI.REZULTATE ȘI DISCUȚII ................................................................................. 20
6.1. REZULTATE ȘI DISCUȚII PRIVIND REACTIVITATEA ȘI
ÎNRĂDĂCINAREA IN VITRO A SPECIILOR DE DIANTHUS ......................................... 20
6.2. REZULTATE ȘI DISCUȚII PRIVIND CARACTERIZAREA MUTANȚILOR
SUPRESORI SUPERROOT2 LA ARABIDOPSIS THALIANA .............................................. 24
6.3. REZULTATE ȘI DISCUȚII PRIVIND CARTAREA MUTAȚIILOR SUPRESOR
.......................................................................................................................... 31
6.4. REZULTATE PRIVIND IDENTIFICAREA DE GENE CANDIDATE
CORESPUNZĂTOARE MUTANȚILOR ȘI SECVENȚIEREA ACESTORA ...................... 36
VII.CONCLUZII ........................................................................................................ 38
BIBLIOGRAFIE SELECTIVĂ.................................................................................. 39

3
INTRODUCERE
Înmulţirea clonală este folosită pe scară largă în horticultură, agricultură,
silvicultură, în programele de conservare a biodoversității, pentru a înmulţi plantele elită
obţinute în ameliorare sau selectate din populaţiile naturale. Formarea rădăcinilor
adventive constituie un pas cheie în înmulţirea vegetativă, prezentând un interes
economic major deoarece multe specii manifestă dificultăţi la procesul de înrădăcinare.
Înțelegerea mecanismelor moleculare care controlează apariția rădăcinilor adventive
reprezintă o prioritate a studiului biologiei rădăcinilor adventive.
Tema prezentei teze de doctorat este axată pe studiul controlului genetic al
formării rădăcinilor adventive, prin identificarea de noi gene implicate în formarea
acestora. Partea experimentală s-a desfășurat în două locații: la USAMV Cluj-Napoca și
UPSC Umeå, Suedia. La USAMV, s-a testat înrădăcinarea adventivă și reactivitatea in
vitro la trei specii rare de Dianthus din România, în condițiile în care biodiversitatea
vegetală este amenințată și conservarea acesteia reprezintă o prioritate la nivel
internațional. Deoarece unele specii rare din Lista Roșie manifestă dificultăți în formarea
de rădăcini adventive in vitro, cercetările au fost extinse pe specia model Arabidopsis
thaliana, a cărei studiu prezintă o serie de avantaje.
Originalitatea prezentei lucrări constă în îmbinarea studiului a două genuri diferite,
prin abordări genetice diferite, în scopul edificării unor aspecte privind formarea
rădăcinilor adventive. Astfel, experimentele au fost continuate la UPSC Umeå, prin
identificarea și caracterizarea de mutanți supresori superroot2 la Arabidopsis thaliana și
studii de genetică moleculară privind cartarea mutațiilor supresor și secvențierea genelor
candidate.
Chiar dacă de-a lungul timpului au fost identificate gene implicate în formarea
rădăcinilor adventive la specii ca Arabidopsis, orezul, porumbul, există în continuare
dileme în ceea ce privește mecanismele moleculare care stau la baza acestui proces
biologic. Datele existente într-un număr limitat de studii sugerează faptul că această
capacitate de a forma rădăcini adventive in vitro este controlată cantitativ de gene majore
la specii ca stejarul, plopul sau eucaliptul. La specia model Arabidopsis thaliana au fost
identificaţi o serie de mutanţi afectaţi în răspunsul la auxine, transportul sau conjugarea

4
acestora. Între acești mutanți superroot2 (sur2) (DELARUE et al., 1998) produce o
cantitate excesivă de auxine şi, ca urmare, plantele dezvoltă în mod spontan rădăcini
adventive la nivelul hipocotilului (DELARUE et al., 1998; SORIN et al., 2005).
Mutantul sur2 a fost identificat şi caracterizat în grupul BELLINI şi s-a demonstrat că
gena mutantă codifică citocromul P450, CYP83B1, care este implicat în oxidarea indole-
3-acetaldoximei în procesul de biosinteză a glucozinolaţilor (BARLIER et al., 2000).
Fenotipul particular al acestui mutant în relaţie directă cu nivelul endogen de auxine
indică faptul că aspecte importante ale homeostazei auxinelor şi ale capacităţii de formare
a rădăcinilor adventive pot fi dezvăluite prin studiul în detaliu a acestui mutant şi a
supresorilor lui. Adresarea funcţiei unei gene se poate face prin intermediul a două
strategii bazate pe genetica directă sau pe genetica inversă (ALONSO-BLANCO et al.,
2009; ALONSO and ECKER, 2006). Deşi clonarea pozițională este un instrument al
geneticii directe, utilizat pe scară largă pentru a izola gene de la diferite organisme (CHI
et al., 2008), utilitatea acesteia poate fi exploatată pe deplin doar în cadrul sistemelor
model, cum este specia Arabidopsis thaliana. Principiul din spatele clonării poziționale
constă în restrângerea sistematică a intervalului genetic care conţine o mutaţie, prin
excluderea secvenţiată a tututor celorlalte regiuni din genom (LUKOWITZ et al., 2000).
Acest lucru poate fi realizat prin utilizarea de markeri deja disponibili şi/sau a unora nou
generați, care să fie polimorfici între accesiunile utilizate pentru generarea populaţiilor de
cartare. Diferite strategii bazate pe clonare au fost deja descrise (JANDER et al., 2002;
LUKOWITZ et al., 2000; PETERS et al., 2003) şi toate se bazează pe disponibilitatea
unei colecţii de markeri genetici extrem de dense, pentru a oferi o rezoluţie de cartare
adecvată. Această disponibilitate de markeri reprezintă un factor major, limitativ, al
progresului procesului de cartare.
Pentru a depăși acest inconvenient provocat de lipsa formării rădăcinilor adventive
şi pentru o mai bună înţelegere a procesului de formare a acestora, au fost efectuate
numeroase studii fiziologice, biochimice şi moleculare în vederea identificării punctelor
cheie care controlează formarea rădăcinilor adventive. În pofida tuturor acestor eforturi
colective, sunt încă multe aspecte care trebuie elucidate şi multe întrebări legate de acest
subiect își așteaptă în continuare răspunsul.

5
I. IMPORTANȚA STUDIULUI RĂDĂCINILOR ADVENTIVE
În ceea ce priveşte studiul formării rădăcinilor adventive, secolul XXI abia începe
să asimileze observaţii, ipoteze şi teorii lansate în cursul secolului XX, iar cercetătorii,
profitând de noile tehnologii disponibile în biologia moleculară au şansa să avanseze cu
investigaţiile la un nivel neimaginat într-o perioadă relativ scurtă (HAISSIG and DAVIS,
1994).
Cercetarea în domeniul biologiei rădăcinilor adventive deţine un loc special în
ştiinţă deoarece oferă o platformă excelentă pentru cercetarea fundamentală în direcţia
elucidării proceselor de creştere şi dezvoltare a plantelor. Rezultatele obţinute în urma
cercetării sunt foarte importante, deoarece ele pot fi aplicate în agronomie, horticultură,
silvicultură, în programele de conservare a biodiversității prin culturi in vitro. În urma
cercetărilor efectuate asupra procesului de formare a rădăcinilor adventive, propagarea
plantelor a înregistrat progrese prin dezvoltarea sistemelor necesare menţinerii unor
condiţii favorabile înrădăcinării butaşilor.
Descoperirea auxinei şi al rolului acesteia în stimularea înrădăcinării a avut un
impact major atât asupra cercetării procesului de formare a rădăcinilor cât şi în industria
înmulţirii plantelor. Ulterior, au fost sintetizaţi şi alţi compuşi care au fost chiar mai
eficienţi decât AIA; acidul indolilbutiric (IBA) şi acidul naftlilacetic (ANA) devenind
substanţe standard folosite. Dezvoltarea biologiei moleculare oferă abilitatea de a adresa
întrebări şi de a găsi răspunsuri care nici nu puteau fi abordate cu doar câţiva ani în urmă.
Este necesar să se ajungă la o înţelegere completă a elementelor regulatoare ale
procesului de formare a rădăcinilor adventive.
1.1. IMPORTANŢA COMERCIALĂ A RĂDĂCINILOR ADVENTIVE PENTRU
AGRICULTURĂ, HORTICULTURĂ ȘI SILVICULTURĂ
Culturile agricole au o mare arie de răspândire, fiind utilizate pentru consum
uman, ca hrană pentru animale sau pentru producera de materii prime. Aceste culturi se
pot împărţi în două categorii: cele care se înmulţesc vegetativ şi cele la care produsul
final (fructele sau materia uscată) este influenţat de prezenţa rădăcinilor adventive
(DAVIES et al., 1994).

6
Înmulţirea vegetativă este cea mai importantă metodă de înmulţire în scopuri
comerciale pentru foarte multe specii horticole, horticultura fiind ramura agronomiei în
care înrădăcinarea adventivă are un rol şi un impact major. Înrădăcinarea slabă reprezintă
o limitare comercială serioasă a propagării asexuate la multe specii horticole, sistemele
de înmulțire utilizate fiind costisitoare și necesitând timp îndelungat. Înmulțirea clonală
rămâne una dintre cele mai importante metode de propagare, în scopul producerii
comerciale, a celor mai multe specii horticole. Din păcate, înrădăcinarea proastă continuă
să reprezinte un factor limitativ al propagării asexuate a multor specii lemnoase horticole
și silvice. Abilitatea de a înrădăcina specii recalcitrante va duce la dezvoltarea de noi
produse și a unei noi piețe de desfacere. Procesul de înrădăcinare și aclimatizare a
plantelor din culturile de țesuturi trebuie îmbunătățit pentru ca biotehnologia să fie
integrată comercial în propagarea și producția de culturi (DAVIES et al., 1994).
1.2. IMPORTANȚA RĂDĂCINILOR ADVENTIVE PENTRU PROGRAMELE DE
CONSERVARE A BIODOVERSITĂȚII
În condițiile în care biodiversitatea vegetală este amenințată, conservarea acesteia
reprezintă o prioritate la nivel internațional. În general, strategiile de conservare se
bazează pe managementul și gestionarea populațiilor sălbatice din natură, dar acestea
necesită metode complementare de conservare ex situ, care ajută conservarea plantelor și,
în unele cazuri, reprezintă singura alternativă. Conservarea in vitro presupune
prezervarea de ţesuturi de plante incluzând protoplaşti, suspensii de celule, culturi de
calus, embrioni etc. şi se practică în special la speciile înmulţite vegetativ şi cele cu
seminţe recalcitrante (GHIDRA et al., 2004). Printre obstacolele care se pot întâlni în
cadrul procesului de conservare a biodiversității prin culturi in vitro pot fi amintiți
contaminanții exogeni și endogeni ai culturilor in vitro, cât și lipsa procesului de
înrădăcinare. Formarea rădăcinilor adventive poate constitui, altfel, un factor limitativ în
ceea ce privește reușita micropropagării și, mai departe, a aclimatizării materialului
vegetal, în vederea reintroducerii lui în zonele de habitat natural (POP and PAMFIL,
2011).

7
II. BIOLOGIA RĂDĂCINILOR ADVENTIVE
2.1. DESCRIEREA ȘI CARACTERIZAREA RĂDĂCINILOR ADVENTIVE
În funcție de originea lor, rădăcinile se pot clasifica în: rădăcini primare
(principale) care provin din radicula embionului și cresc după geminație, secundare
(laterale) care se formează postembrionar, pe rădăcinile principale sau pe alte rădăcini
secundare și rădăcini adventive, care apar postembrionar pe alte organe decât pe
rădăcină, luând naștere din celule diferențiate.
Sistemul radicular al monocotiledonatelor (cerealelor) este, în general, compus din
rădăcini adventive, care se produc în mod natural, făcând parte din dezvoltarea normală a
unei plante (HOCHHOLDINGER et al., 2004). În literatura de specialitate, există autori
care consideră că numele de rădăcini adventive este impropriu pentru acest tip de rădăcini
la cereale, deoarece la monocotiledonate acestea sunt determinate genetic, în timp ce la
dicotiledonate acestea se formează în condiții speciale (TAKAHASHI et al., 2003).
Rădăcinile adventive pot apărea, în mod natural, din ţesutul tulpinii sub condiţii de
stres. Acestea pot fi, de asemenea, induse de deteriorarea mecanică sau în urma
regenerării microbutașilor prin culturi de țesuturi in vitro (LI et al., 2009). Rădăcinile
adventive sunt rădacini postembrionare care iau naștere din tulpină şi frunze şi din
ţesuturile dinafara periciclului rădăcinilor bătrâne (GEISS et al., 2009). Există cel puţin
două căi prin care rădăcinile adventive se formează: prin organogeneză directă din celule
specializate, cum ar fi cambiul, sau din calus, în urma unei deteriorări mecanice (Figura
1). Rădăcinile adventive reprezintă una dintre cele mai importante căi de propagare
vegetativă şi una dintre cele mai importante metode de producţie comercială a culturilor
agricole, horticole, silvice din întreaga lume (POP et. al., 2011).

8
Figura 1. Rădăcini adventive naturale (a, b, c) și induse (d) (după GEISS et al., 2009)
2.2. FACTORI CARE INFLUENȚEAZĂ FORMAREA RĂDĂCINILOR
ADVENTIVE
Procesul de dezvoltarea a plantelor este reglat de factori genetici şi de mediu, care
au efecte asupra biosintezei, metabolismului, transportului şi căii de semnalizare a
auxinelor (HAN et al.,2009). Mulţi factori de mediu şi endogeni, cum ar fi temperatura,
lumina, hormonii (în special auxinele), zaharurile, sărurile minerale și alte molecule
funcţionează ca semnale şi induc grupuri de celule care își schimbă structura, influențând
formarea și reglarea rădăcinilor adventive (POP et al., 2011).
Fitohormonii joacă un rol complex, exercitând efecte directe, care acţionează
asupra diviziunii celulare sau creșterii celulelor sau indirecte, interacționează cu alte
molecule sau fitohormoni. Auxinele s-au dovedit a fi intim implicate în procesul de
formare a rădăcinilor adeventive (WIESMAN et al., 1988) şi etapele fiziologice
interdependente ale procesului de înrădăcinare sunt asociate cu modificări ale
concentraţiilor de auxine endogene (HELOIR et al., 1996). Mai multe studii au subliniat
faptul că poliaminele joacă un rol în formarea rădăcinilor adventive (BIONDI et al.,
1990; HAUSMAN et al., 1994; HELOIR et al., 1996). Unii cercetători au sugerat o
posibilă interdependenţă între auxine şi poliamine în ceea ce privește inducţia formării
rădăcinilor adventive (HAUSMAN et al., 1995). Alte studii au raportat că etilena

9
reprezintă o moleculă semnal în rădăcinile adventive (BIONDI et al., 1990). Producția de
etilenă indusă de AIA poate fi un factor implicat în stimularea rădăcinilor adventive
(PAN et al., 2002). Unele dintre lucrările recente au raportat alte molecule implicate în
semnalizarea rădăcinilor adeventive, cum ar fi oxidul de azot (NO) (PAGNUSSAT et al.,
2003; PAGNUSSAT et al., 2004; PAGNUSSAT et al., 2002), pe bază de peroxid de
hidrogen (H2O2) (LI et al., 2007) şi monoxid de carbon (CO) (XU et al., 2006). Se știe,
de asemenea, că activitatea peroxidazei reglementează catabolism AIA şi acţionează ca
un marker pentru faze succesive, atingând, de obicei, un minim în faza de inducţie şi un
maxim în faza de iniţiere. În ultimii ani s-a realizat un mare progres în ceea ce privește
înțelegerea semnalizării auxinelor și a genelor cu răspuns la auxine. (PARRY and
ESTELLE, 2006; QUINT and GRAY, 2006). Abilitatea de a identifica caracterele care
controlează capacitatea de înmulţire vegetativă este extrem de importantă în perspectiva
direcţiilor de urmat, în vederea încorporării lor în programele de ameliorare. În acest
sens, identificarea de markeri moleculari asociaţi acestor caractere devine crucială în
selecţia genotipurilor valoroase.
III. STUDIUL FORMĂRII RĂDĂCINILOR ADVENTIVE
3.1. ARABIDOPSIS THALIANA - PLANTĂ MODEL ÎN STUDIUL FORMĂRII
RĂDĂCINILOR ADVENTIVE
O plantă model, aleasă pentru studiul rădăcinilor adventive, trebuie să prezinte
următoarele caracteristici genetice (SOMERVILLE ȘI KOORNNEEF, 2002):
să aibă un genom bine caracterizat.
să dispună de hărți fizice și genetice.
să aibă un genom de mici dimensiuni cu grupuri de linkage puține (cromozomi
puțini).
să aibă colecții vaste de mutanți caracterizați și cartați pentru trăsături de interes și
noi mutanți ușor de produs și examinat.
să existe o bibliotecă genomică vastă disponibilă pentru analiză (ex. în YAC-
uri=Yeast Artificial Chromosome).

10
planta trebuie să fie capabilă de autoîncrucișare și să fie fertilă, să permită
realizarea ușoară de încucișări, să fie ușor de emasculat, de dimensiune mică.
să producă un număr mare de semințe pe încrucișare și pe plantă.
să aibă intervale scurte ale generației (de la câteva săptămâni la câteva luni).
să manifeste o variabilitate genetică pentru trăsăturile de interes.
să existe un sistem disponibil de marcare a genelor (transpozomi, T-DNA).
să existe sisteme de recombinare homoloagă disponibile pentru mutageneza
direcționată.
complementare genetică disponibilă și ușor de realizat cu ajutorul ameliorării
tradiționale sau folosind bibliotecile genomice.
să beneficieze de sisteme de transformare celulară și tisulară bine dezvoltate.
O altă caracteristică de bază a unei specii model constă în faptul că rezultatele
cercetărilor trebuie să fie capabile să fie transferate la alte specii vegetale sau alte
sisteme. Dintre toate speciile de plante, Arabidopsis thaliana (Figura 2) iese în evidență,
fiind cea mai studiată. Alte plante ca porumbul, soia, petunia, tomatele, mazărea, odată
considerate candidatele ideale în cercetarea vegetală, sunt acum depășite de numărul
impresionant al publicațiilor științifice axate pe această specie model.
Figura 2. Planta model Arabidopsis thaliana (original)
Arabidopsis are o serie de avantaje față de alte plante model, printre care se
numără un genom mic (125 Mb) care a fost secvențiat în anul 2000 (SequenceViewer,
AGI). Există hărţi genetice şi fizice extinse pentru toți cei cinci cromozomi

11
(MapViewer). Planta are un ciclu de viaţă scurt (circa 6 saptamani de la germinare la
obținerea de seminţe mature), având o producție prolifică de seminţe şi cultivare, fiind
accesibilă în spaţiu limitat. Există o serie de metode eficiente de transformare utilizând
Agrobacterium tumefaciens și un număr mare de linii mutante şi resurse genomice, dintre
care multe sunt disponibile de la centrele de resurse (stock centres). Există, de asemenea
o comunitate de cercetare multinaţională în laboratoare academice, guvernamentale şi
industriale. Astfel de avantaje au făcut ca Arabidopsis să fie considerat un organism
model pentru studiile de biologie celulară și moleculară ale plantelor cu flori. TAIR (The
Arabidopsis Information Resource) colectează şi pune la dispoziţie informaţii provenite
în urma eforturilor menționate mai sus.
3.1.1. The Arabidopsis Information Resource (TAIR)
TAIR (The Arabidopsis Information Resource, http://arabidopsis.org) reprezintă
baza de date pentru organismul model în întregime secvențiat și intensiv studiat
Arabidopsis thaliana. Datele de pe TAIR provin în mare parte din literatura de
specialitate având ca temă Arabidopsis și observațiile efectuate de comuniunea de
cercetare.
3.2. SUPRESIA GENEI SUPPERROOT- STRATEGIE ÎN IDENTIFICAREA DE
GENE IMPLICATE ÎN FORMAREA RĂDĂCINILOR ADVENTIVE
În urma unor analize genetice caracteristice geneticii directe (forward genetics), în
care se dorea identificarea de mutanți care să producă auxine în exces, au fost izolați la
Arabidopsis thaliana doi mutanţi denumiţi superroot 1 (sur1) şi superroot2 (sur2).
Aceștia au fost izolaţi în două screening-uri independente de către BOEJERAN și
colaboratorii (1995) și respectiv de DELARUE și colaboratorii (1998). S-a demonstrat că
ambii mutanţi au produs auxine în exces şi au dezvoltat în mod spontan rădăcini
adventive la nivelul hipocotilului, manifestând, de asemenea, un număr ridicat de
rădăcini laterale ca și caracteristică fenotipică a nivelului ridicat de auxine. S-a descoperit
că gena SUR1 (AT2G20610.1) codifică o C-S-liază care are rol în sinteza
glucozinolaţilor, determinând conversia S-alkiltiohidroximat-ului în thiohidroximat
(WOODWARD and BARTEL, 2005).

12
În stare homozigotă, mutantul sur1 poate fi menținut doar in vitro, în schimb
plantele mutante homozigote sur2 pot crește în sol, fiind fertile, iar la maturitate prezintă
un fenotip asemănător tipului sălbatic (DELARUE et al., 1998). Mai târziu, gena SUR2
(AT4G31500) a fost cartată de către BARLIER și colaboratorii (2000) la baza
cromozomului patru și codifică pentru citocromul P450 enzima CYP83B1, fiind implicat
în oxidarea indol-3-acetaldoxinei în procesul de sinteză a glucozinaților. Identificarea
locusului SUR2 având rol în codificarea unei enzime implicate în biosinteza metabolitului
secundar a respins ipoteza că SUR2 este un modulator al homeostaziei auxinelor și a
demonstrat o cuplare a acestora cu sinteza glucozinaților (BAK et al., 2001).
Fenotipul specific al mutantului sur2 corelat cu concentrația ridicată de auxine
sugerează că studiul detaliat al acestui mutant împreună cu studiul supresorilor acestuia
vor putea duce la elucidarea unor importante aspecte legate de formarea rădăcinilor
adventive și homeostaza auxinelor.
Studiul supresorilor este o strategie folosită deseori pentru a identifica relații
funcționale între gene care nu au fost încă descoperite prin alte mijloace genetice sau
biologice. Multe mecanisme care explică fenomenul supresiei genetice au fost descrise,
dar marea diversitate de mecanisme posibile reprezintă o provocare în definirea relației
dintre gena supresor și gena inițială (PRELICH, 1999). Studiul supresorilor este foarte
important, deoarece numeroase gene sunt dificil de identificat prin metode directe de
selecţie. O mutaţie preexistentă adesea sensibilizează procesul pe care îl afectează,
permiţând astfel identificarea de componente adiţionale prin selecţia de supresori.
3.3. MUTAGENEZA CU EMS
Progresele realizate în analiza genetică a dezvoltării plantelor se datorează, de
asemenea și tehnicilor de mutageneză. Mutageneza prin intermediul agenţilor mutageni
chimici, fizici sau genetici realizează legătura între genotip şi fenotip, prin introducerea
de mutaţii la Arabidopsis thaliana (MEINKE et al., 1998). Agenții mutageni prezintă
particularităţi în ce priveşte eficienţa mutagenezei prin numărul de leziuni/genom
transmise în descendenţă şi prin modificările introduse la nivel molecular. Modificările
pot varia de la substituţia unei singure nucleotide până la inserţii sau rearanjări ale unor
regiuni mari de cromozomi. Agenții alchilanții, cum ar fi etil-metan-sulfonat-ul (EMS)

13
induc modificarea chimică a nucleotidelor, ceea ce duce la împerecheri greșite şi
schimbări ale bazelor.
3.4. CARTAREA MUTAȚIILOR
Genetica directă a fost folosită extensiv la speciile model precum Arabidopsis
thaliana. Astfel, funcţia unei gene specifice este în mod normal stabilită în urma
analizării efectelor fenotipice ale alterării secvenţei genei respective. Cu alte cuvinte,
funcţia unei anumite gene poate fi determinată când gena respectivă dobândeşte o
mutaţie, iar mutantul rezultat este analizat şi comparat cu tipul sălbatic pentru a înregistra
diferenţele apărute. Clonarea bazată pe cartare, de asemenea cunoscută sub numele de
clonare de poziție sau cartare, se bazează pe analiza legăturii dintre un fenotip mutant și
markeri genetici sau moleculari ale căror poziţii pe cromozom sunt cunoscute (JANDER,
2006; JANDER et al., 2002; PETERS et al., 2003). Cartarea bazată pe clonare este
eficientă pentru caracterizarea tuturor tipurilor de mutanţi, inclusiv mutanţi T-ADN.
Aceasta rămâne în continuare cea mai bună metodă pentru mutaţii punctiforme sau
inserţii/deleții mici generate de EMS sau de bombardarea cu neutroni, până în momentul
dezvoltării și utilizării unei tehnologii de secvenţiere ieftină şi eficientă (JANDER, 2006;
JANDER et al., 2002).
3.4.1. Tipuri de markeri utilizați în cartare
Pe parcursul dezvoltării cartării bazate pe clonare, markeri folosiți s-au schimbat
de la markeri genetici la markeri moleculari, de exemplu, markeri RFLP (Restriction
Fragment Length Polymorphism) și markeri bazați pe PCR incluzând RAPD (Random
Amplified Polymorphic DNA) și AFLP (Amplified Fragment Length Polymorphism)
(ALONSO-BLANCO et al., 1998; CHANG et al., 1988; CHANG and MEYEROWITZ,
1991; WILLIAMS et al., 1990). În ultimii ani, markerii INDEL (inserție/deleție) şi CAPS
(Cleaved Amplified Polymorphic Sequence) au devenit cei mai fiabili markeri pentru
cartare (BELL and ECKER, 1994; KONIECZNY and AUSUBEL, 1993). Atât markerii
CAPS, cât și cei INDEL sunt markeri frecvent utilizați bazați pe PCR. Cu sau fără
digestia cu enzime de restricție, produșii PCR sunt supuși analizei prin electroforeză, în
urma căreia datele sunt obţinute într-o manieră directă și ușoară (BELL and ECKER,

14
1994; KONIECZNY and AUSUBEL, 1993). Numeroși markeri INDEL şi CAPS au fost
creați și raportați, şi sunt, de asemenea, prezenți în colecția TAIR
(http://www.arabidopsis.org/marker). Cu toate acestea, acești markeri disponibili nu sunt
distribuiți în mod uniform pe cromozomi, existând diferite motive care ar explica
numărul limitat de markeri disponibili. De exemplu, numai acei markeri strâns linkați de
fenotipul mutanților pot fi raportați şi transmiși la TAIR, iar, în multe cazuri, markerii nu
sunt trimiși după clonarea genei.
3.4.2. Procedeul de cartare fină
Cartarea fină (fine mapping) reprezintă procesul prin care se determină poziţia
exactă a unei gene pe cromozom și trebuie parcurse o serie de etape succesive (Figura 3).
Figura 3. Procedura de cartare fină (original)
În final, intervalul genetic conţinând mutaţia este redus treptat prin identificarea de
ambele părţi de noi markeri situaţi tot mai aproape de mutaţie, astfel ca la final să
flancheze o regiune mai mică de 100-150 kb, regiune care la Arabidopsis poate conţine în

15
medie 10-30 de gene. Odată această regiune definită se poate trece la selectarea
posibilelor gene candidate pentru a fi secvenţiate în vederea identificării mutaţiei
(JANDER et al., 2002).
IV. SCOPUL ȘI OBIECTIVELE TEZEI DE DOCTORAT
Formarea rădăcinilor adventive constituie un factor limitativ în propagarea
vegetativă, iar în lipsa acestora se înregistrează pierderi economice semnificative în
agricultură, silvicultură, horticultură și în conservarea ex situ a resurselor vegetale prin
culturi in vitro. Scopul prezentei teze de doctorat și al proiectului de cercetare la care am
colaborat constă în elucidarea mecanismelor moleculare care stau la baza formării
rădăcinilor adventive, prin identificarea de gene implicate în controlul acestora.
Rezultatele acestei teze de doctorat vor contribui la o mai bună înțelegere a controlului
genetic al formării rădăcinilor adventive și se vor alătura eforturilor comunității științifice
internaționale de a elucida noi aspecte ale biologiei vegetale.
Obiectivele cercetării au constat în:
Testarea reactivității și înrădăcinării in vitro a trei specii endemice din
România (din genul Dianthus).
Generarea de mutanți supresori ai mutantului superroot2-1 de la planta
model Arabidopsis thaliana, care produce o cantitate excesivă de auxine și un număr
foarte mare de rădăcini adventive.
Caracterizarea genetică și fenotipică a mutanților supresori identificați.
Generarea populațiilor de cartare pentru mutanții supresori identificați
Generarea de markeri moleculari care să faciliteze procesul de cartare
Cartarea a opt mutații supresor pe cromozomi
Alegerea de gene candidate și secvențierea acestora

16
V. MATERIALE ȘI METODE
5.1. MATERIALE ȘI METODE UTILIZATE PENTRU ÎNRĂDĂCINAREA IN
VITRO A SPECIILOR DE DIANTHUS
Pentru realizarea primelor experimente desfășurate la USAMV Cluj-Napoca, care
au vizat testarea reactivițății in vitro a speciilor de Dianthus, s-a utilizat urmatoarele
materiale și metode: s-a recoltat materialului biologic reprezentat de plante întregi și
semințe apartinând la trei specii rare de Dianthus (Dianthus spiculifolius, Dianthus
henteri și Dianthus giganteus banaticus) din Lista Roşie a Florei României (OPREA,
2005). S-a realizat sterilizarea explantelor și a semințelor cu ajutorul diferiților agenți de
sterilizare și inițierea de culturi in vitro din semințe și fragmente nodale. S-au testat cinci
variante de medii de cultură având la bază mediul MS simplu, suplimentate cu
fitoregulatori de creștere și s-a urmărit reactivitatea celor trei specii de Dianthus pe
acestea. S-a determinat rata de germinare a semințelor, rata de contaminare a culturilor in
vitro inițiate din semințe și fragmente nodale, randamentul microlăstăririi și lungimea
medie a lăstarilor. S-au elaborat observații cu privire la înrădăcinarea adventivă in vitro.
a b c
Figura 4. Flori și semințe a) D. spiculifolius b) D. giganteus banaticus c) D. henteri
(original)

17
5.2. MATERIALE ȘI METODE PENTRU CARACTERIZAREA MUTANȚILOR
SUPRESORI SUPERROOT2 LA ARABIDOPSIS THALIANA
În continuare, cercetările au fost continuate în centrul de cercetare Umeå Plant
Science Center, Suedia. S-au identificat mutanți supresori ai superroot2 (DELARUE et
al., 1998) prin realizarea mutagenezei cu agentul chimic EMS a semințelor mutantului
sur2 și s-au obținut noi mutanți la specia model Arabidopsis thaliana (Figura 5), s-au
analizat liniile mutante obținute și s-a confirmat fenotipul supresor prin obținerea unui
număr de rădăcini adventive mai redus decât în cazul mutantului sur2. S-au analizat în
continuare raporturile de segregare a opt mutanți supresori identificați în prezenta teză de
doctorat.
a b
Figura 5 a) Rozetă de Arabidopsis thaliana b) Plantă
întreagă de Arabidopsis thaliana (original)
Caracterizarea fenotipică a mutanților în condiții de lumină s-a realizat prin
determinarea lungimii rădăcinilor principale, a hipocotilelor, a numărului de rădăcini
laterale și a densității acestora. Caracterizarea fenotipică a mutanților în condiții de
întuneric s-a determinat prin evaluarea numărului de rădăcini adventive și laterale. S-a
determinat cantitatea de AIA (acid indolil-acetic) în stare liberă din partea radiculară și
apicală a plantulelor mutante la doi dintre mutanții supresori identificați (2146.4 și 2344)
prin faza solidă de extracție și analiza GC-MS (cromatografie în gaz/spectrofotometrie de
masă). Plantulele mutante au fost crescute conform protocolului descris de SANTONI et
al. (1994) și SORIN et al. (2005), pe un mediu de cultură Milieu Arabidopsis (INRA)
(Figura 6).
a b

18
a
b
c
d
Figura 6. a) semințe germinate in vitro, semănate în linii b) placă Petri pătrată cu
plantule de Arabidopsis pe mediu de cultură c) plantarea în sol a plantulelor d)
etichetarea și păstrarea în seră (original)
Generarea populațiilor de cartare corespunzătoare mutanților identificați s-a
realizat prin obținerea plantelor mutante F2, selecția plantelor cu fenotip mutant
reprezentând 25% din totalul de plante, urmată apoi de izolarea ADN-ului din plantele
selectate în plăci de stocare a ADN-ului de 96 de godeuri.
Pentru realizarea cartării mutațiilor pe cromozomi s-au testat markeri clasici SSLP
și INDEL din baze de date existente (TAIR, Cereon), care să fie polimorfici între
ecotipurile Ws-4 și Col-0. S-au proiectat markeri noi INDEL și s-a testat polimorfismul
între ecotipurile Ws-4 și Col-0. S-a amplificat ADN-ului mutant prin PCR (Figura 7 a) și
produșii PCR au fost analizați prin electroforeză în gel de agaroză de 4% sau 2%, în
funcție de mărimea alelelor și vizualizați (Figura 7 b). S-a încercat găsirea de intervale
genetice restrânse care să conțină mutația de interes.
c
d
b

19
a b c
Figura 7. a) Termociclu BioRad b) Molecular Imager GelDoc XR de la BioRad
c) Spectrofotometrul Nanodrop ND-1000
Prin consultarea bazei de date www.arabidopsis.org (TAIR) s-au aleges gene
candidate din regiunile respective, având în vedere date din literatura de specialitate. S-au
proiectat perechi de primeri care să flancheze genele candidate, s-au amplificat prin PCR
și analizat elecroforetic în gel de agaroză produșii rezultați, s-au verificat
spectofotometric concentrațiile de ADN (Figura 7 c). Probele au fost pregătite pentru
secvențiere, iar după primirea rezultatelor secvențierii, la final, s-au aliniat secvențele
mutației cu secvența referință Col-0 (cu ajutorul programului ClustalW) și s-a încercat
identificarea mutației.
20
VI. REZULTATE ȘI DISCUȚII
6.1. REZULTATE ȘI DISCUȚII PRIVIND REACTIVITATEA ȘI
ÎNRĂDĂCINAREA IN VITRO A SPECIILOR DE DIANTHUS
Au fost identificate și recoltate din siturile Natura2000 trei specii endemice din
genul Dianthus (Dianthus spiculifolius, Dianthus henteri și Dianthus giganteus
banaticus), care se găsesc în Lista Roșie a plantelor pe cale de dispariție din România. În
condițiile în care biodiversitatea vegetală este amenințată, s-a urmărit conservarea acestor
specii prin păstrarea lor în culturi in vitro repetate, în vederea aclimatizării și repopulării,
în final, a zonelor de unde speciile provin. Experimentele s-au desfășurat în
Departamentul de genetică și ameliorarea plantelor, din USAMV Cluj-Napoca.
În cazul speciilor de Dianthus luate în studiu, sterilizarea explantelor desprinse de
pe plantele mature a fost dificil de efectuat din cauza gradului ridicat de contaminare cu
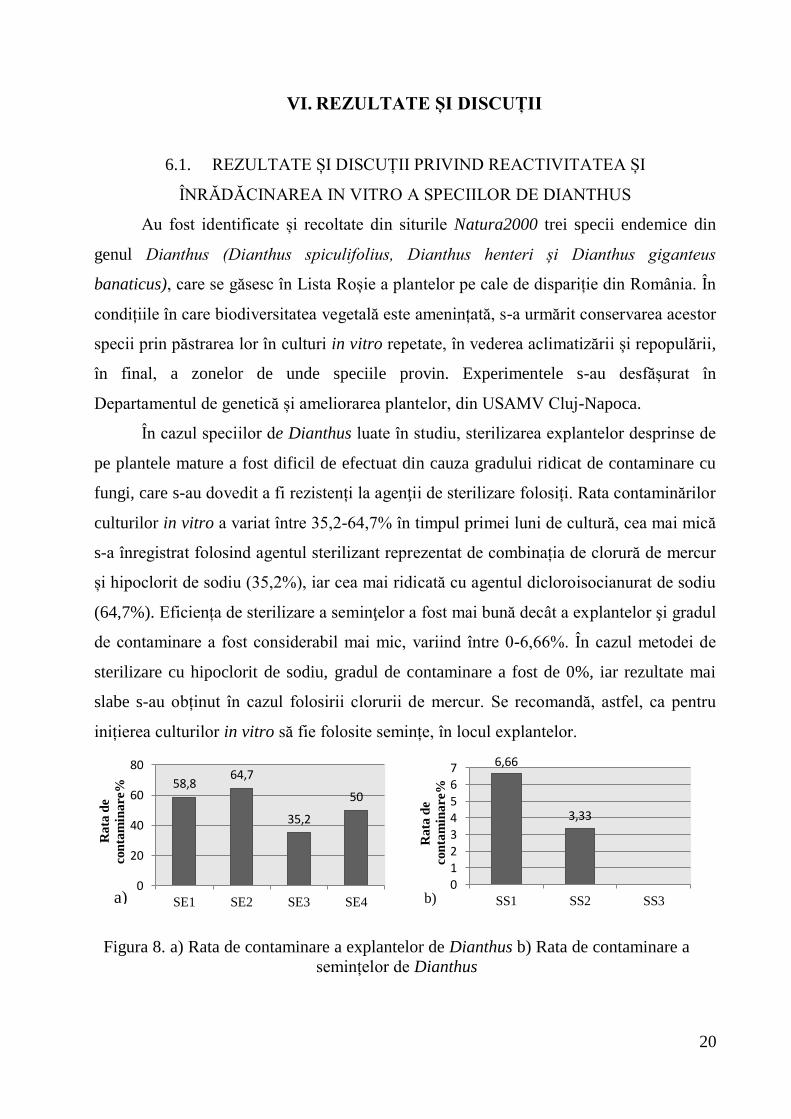
fungi, care s-au dovedit a fi rezistenți la agenţii de sterilizare folosiți. Rata contaminărilor
culturilor in vitro a variat între 35,2-64,7% în timpul primei luni de cultură, cea mai mică
s-a înregistrat folosind agentul sterilizant reprezentat de combinația de clorură de mercur
și hipoclorit de sodiu (35,2%), iar cea mai ridicată cu agentul dicloroisocianurat de sodiu
(64,7%). Eficiența de sterilizare a seminţelor a fost mai bună decât a explantelor şi gradul
de contaminare a fost considerabil mai mic, variind între 0-6,66%. În cazul metodei de
sterilizare cu hipoclorit de sodiu, gradul de contaminare a fost de 0%, iar rezultate mai
slabe s-au obținut în cazul folosirii clorurii de mercur. Se recomandă, astfel, ca pentru
inițierea culturilor in vitro să fie folosite semințe, în locul explantelor.
Figura 8. a) Rata de contaminare a explantelor de Dianthus b) Rata de contaminare a
semințelor de Dianthus
58,8 64,7
35,2
50
0
20
40
60
80
SE1 SE2 SE3 SE4
Rata
de
con
tam
inare%
6,66
3,33
0 1 2 3 4 5 6 7
SS1 SS2 SS3
Rata
de
con
tam
inare%
b)
a
a)

21
Semințele au germinat în următoarea ordine: D. giganteus banaticus, D.
spiculifolius, D. henteri, timpul de germinare fiind cuprins între 3-30 de zile de la
inoculare, pe mediul MS suplimentat cu AIA și BAP. Rata de germinare a fost de 83,33%
(D. giganteus banaticus), 70,83% (D. spiculifolius) respectiv 52,5% (D. henteri),
rezultatele sunt satisfăcătoare și similare celor obținute în cazul altor specii de Dianthus
(CRISTEA et al., 2009; PAPAFOTIOU and STRAGAS, 2009). Viabilitatea semințelor a
fost influențată de condițiile în care acestea au fost păstrate până în momentul semănării
lor pe mediul de cultură.
Au fost testate cinci variante de mediu de cultură pentru observarea
comportamentului speciilor in vitro, din punctul de vedere al înrădăcinării adventive.
Speciile au avut o reactivitate in vitro medie, calea de regenerare a explantelor a fost
morfogeneza directă. S-au înregistrat diferențe în ceea ce privește comportamentul in
vitro al culturilor, gradul de regenerare, de creștere a lăstarilor, de vitrificare și
înrădăcinare, în funcție de mediul de cultură folosit.
Cele mai bune rezultate ale ratei de regenerare s-au obținut pe mediul MSV1
(raport de auxine/citochinine de 1/10) pentru toate cele trei specii de Dianthus. După 30
de zile, D. giganteus banaticus a înregistrat cea mai ridicată rată de regenerare pe MSV1,
cu o medie de 12 lăstari/explant, D. spiculifolius a avut o valoare medie a numărului de
lăstari mai scăzută, de 7,8, iar D. henteri de 6,7. Comparând rezultatele obținute pe
MSV1 și MSV4, s-a observat că diferenţele dintre specii s-au menţinut în cazul ambelor
formule fitohormonale, valorile medii pe MSV4 fiind ușor mai scăzute (Figura 9).
Înrădăcinarea adventivă a fost slabă, s-a format calus la baza explantelor, dominanța
citochininei favorizând formarea de lăstari. Randamentele neoformării lăstarilor pe
mediile MSV4 și MSV3 au fost comparabile la D. spiculifolius și la D. henteri, în timp ce
la D. giganteus banaticus, reducerea raportului auxină/citochinină la 1/20 a avut efect
negativ, cu o medie de 8,3 lăstari/explant. Scăderea concentraţiei de auxină a avut un
efect pozitiv asupra formării de rădăcini adventive şi nu s-a format calus. Modificarea
balanţei hormonale prin creșterea raportului auxine/citochinine la ½ (MSV2) nu a afectat
randamentul lăstăririi decât slab la D. gigantheus banaticus. Efectul pozitiv asupra
rizogenezei s-a manifestat doar la D. spiculifolius, iar la D. henteri nu s-a observat nici o
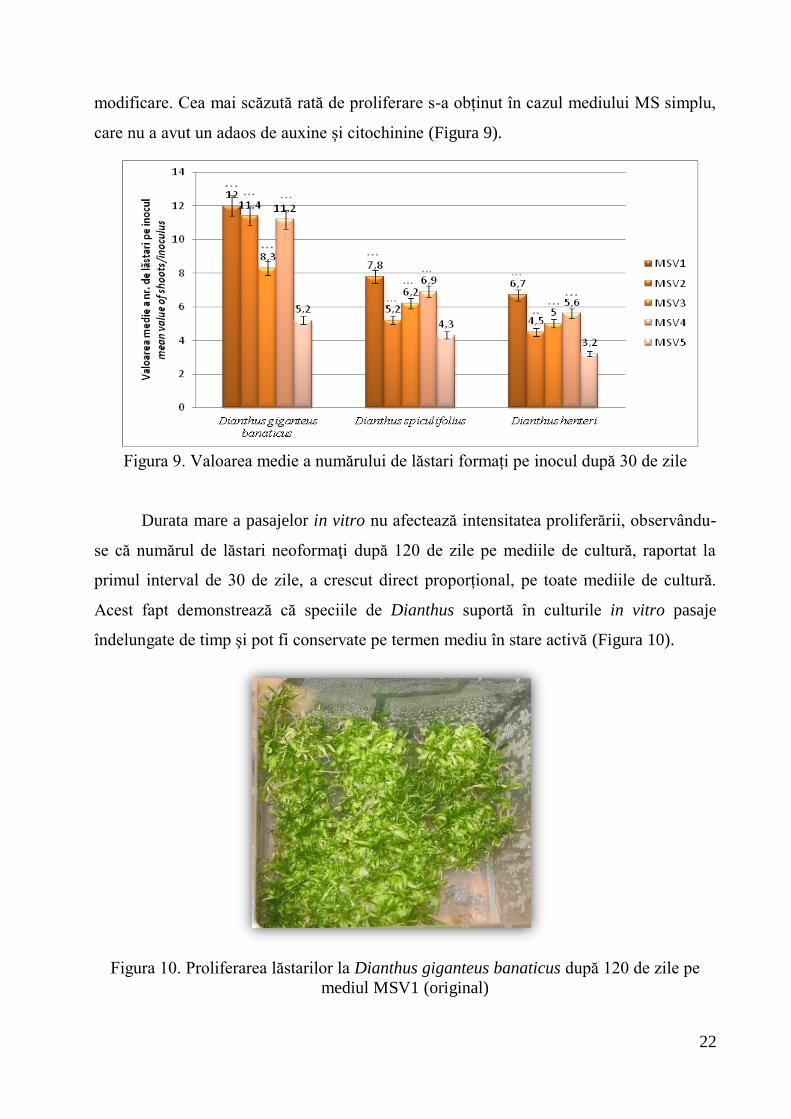
22
modificare. Cea mai scăzută rată de proliferare s-a obținut în cazul mediului MS simplu,
care nu a avut un adaos de auxine și citochinine (Figura 9).
Figura 9. Valoarea medie a numărului de lăstari formați pe inocul după 30 de zile
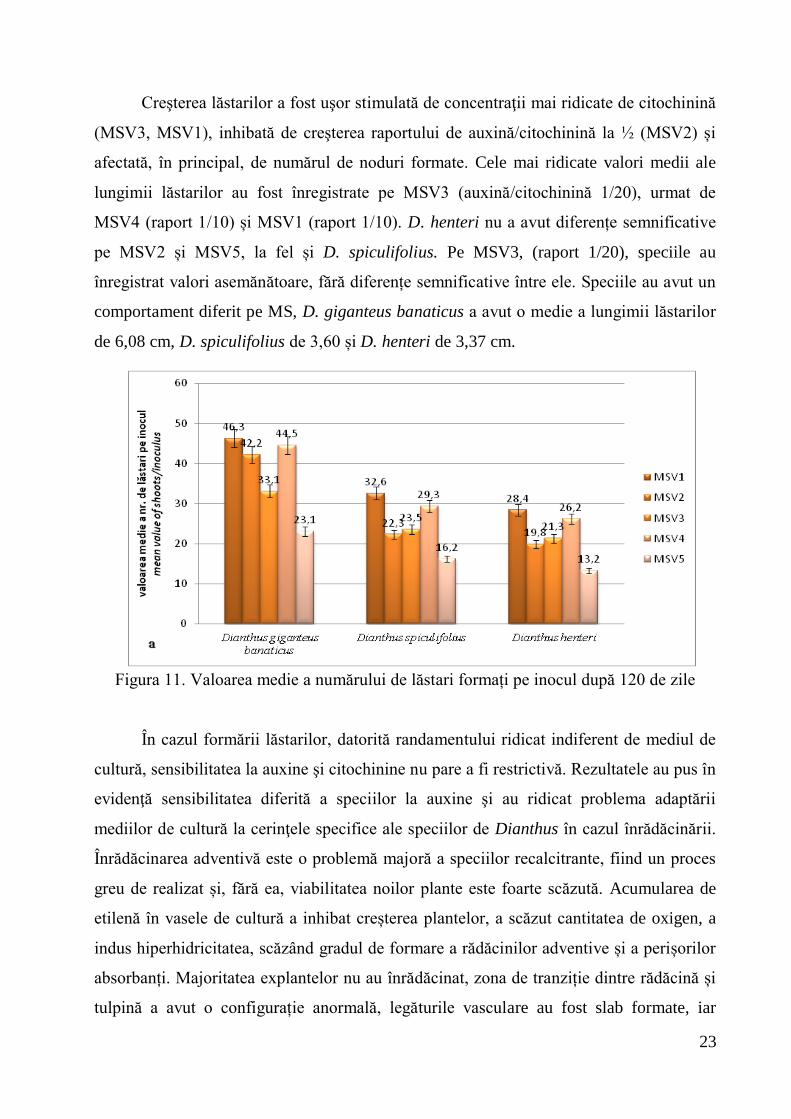
Durata mare a pasajelor in vitro nu afectează intensitatea proliferării, observându-
se că numărul de lăstari neoformaţi după 120 de zile pe mediile de cultură, raportat la
primul interval de 30 de zile, a crescut direct proporțional, pe toate mediile de cultură.
Acest fapt demonstrează că speciile de Dianthus suportă în culturile in vitro pasaje
îndelungate de timp şi pot fi conservate pe termen mediu în stare activă (Figura 10).
Figura 10. Proliferarea lăstarilor la Dianthus giganteus banaticus după 120 de zile pe
mediul MSV1 (original)
23
Creşterea lăstarilor a fost uşor stimulată de concentraţii mai ridicate de citochinină
(MSV3, MSV1), inhibată de creşterea raportului de auxină/citochinină la ½ (MSV2) și
afectată, în principal, de numărul de noduri formate. Cele mai ridicate valori medii ale
lungimii lăstarilor au fost înregistrate pe MSV3 (auxină/citochinină 1/20), urmat de
MSV4 (raport 1/10) și MSV1 (raport 1/10). D. henteri nu a avut diferențe semnificative
pe MSV2 și MSV5, la fel și D. spiculifolius. Pe MSV3, (raport 1/20), speciile au
înregistrat valori asemănătoare, fără diferențe semnificative între ele. Speciile au avut un
comportament diferit pe MS, D. giganteus banaticus a avut o medie a lungimii lăstarilor
de 6,08 cm, D. spiculifolius de 3,60 și D. henteri de 3,37 cm.
Figura 11. Valoarea medie a numărului de lăstari formați pe inocul după 120 de zile
În cazul formării lăstarilor, datorită randamentului ridicat indiferent de mediul de
cultură, sensibilitatea la auxine şi citochinine nu pare a fi restrictivă. Rezultatele au pus în
evidenţă sensibilitatea diferită a speciilor la auxine şi au ridicat problema adaptării
mediilor de cultură la cerinţele specifice ale speciilor de Dianthus în cazul înrădăcinării.
Înrădăcinarea adventivă este o problemă majoră a speciilor recalcitrante, fiind un proces
greu de realizat și, fără ea, viabilitatea noilor plante este foarte scăzută. Acumularea de
etilenă în vasele de cultură a inhibat creșterea plantelor, a scăzut cantitatea de oxigen, a
indus hiperhidricitatea, scăzând gradul de formare a rădăcinilor adventive și a perișorilor
absorbanți. Majoritatea explantelor nu au înrădăcinat, zona de tranziție dintre rădăcină și
tulpină a avut o configurație anormală, legăturile vasculare au fost slab formate, iar

24
rădăcinile nu au prezentat perișori absorbanți și au murit după scurt timp de la
transplantare, ceea ce a dus la formarea unui sistem radicular slab dezvoltat.
Deoarece formarea rădăcinilor adventive reprezintă un factor limitativ în
înrădăcinarea in vitro a speciilor endemice și biodiversitatea fiind în pericol, este
importantă înțelegerea mecanismelor moleculare care stau la baza formării rădăcinilor
adventive.
6.2. REZULTATE ȘI DISCUȚII PRIVIND CARACTERIZAREA MUTANȚILOR
SUPRESORI SUPERROOT2 LA ARABIDOPSIS THALIANA
Un studiu mai detaliat al proceselor genetice care stau la baza formării rădăcinilor
adventive s-a realizat prin experimentele efectuate în centrul de cercetare UPSC (Umeå
Plant Science Center) din Umeå, Suedia, în grupul de cercetare condus de prof. dr.
Catherine Bellini, sub îndrumarea dr. Daniel Păcurar.
Pentru a permite izolarea de gene implicate în procesele de formare a rădăcinilor
adventive, s-au proiectat și efectuat teste de analiză a mutanților supresori ai genei
SUPERROOT 2-1 de la specia model Arabidopsis thaliana, prin analiza a 2345 familii
independente M2, obţinute din seminţe homozigote pentru sur2-1, mutagenizate cu EMS.
Familiile au fost evaluate pentru identificarea indivizilor cu un număr redus de rădăcini
adventive sau care nu prezintă deloc rădăcini adventive în comparație cu sur2-1.
S-au selectat 300 de indivizi considerați potenţiali mutanți supresori, s-a analizat
descendența acestora în M3, pentru confirmarea fenotipului de supresor, s-au ales 48 de
mutanți supresori confirmaţi în generaţia M3, care au prezentat o reducere a numărului de
rădăcini adventive în comparaţie cu sur2. Mutanții au fost încrucișați în vederea testului
de complementare, ce a permis identificarea potenţialelor alele răspunzătoare de mutații.
În urma testului de complementare, supresorii confirmaţi în generaţia M3 au fost
identificaţi ca aparţinând la 37 de grupuri de complementaritate.
Mutanții supresori au fost retroîncrucișați cu sur2-1 (cu fond genetic în Col-0), a
fost analizată generația BC1F2 și analiza segregării a evidenţiat că cele opt mutații
studiate au fost recesive și au avut un raport de segregare de 3:1 (fenotip sur2-1:fenotip
supresor), fapt reconfirmat și în generația BC2F2.
25
Srs.
Srs.
Semințe
semănate
Sawn
seeds
Semințe
germinate
Germinated
seeds
Rata
germinației
%
Germination
rate
Plante cu
fenotip
sur2 (O)
Plants with
sur2
phenotype
(O)
Plante cu
fenotip
supresor (O)
Plants with
srs phenotype
(O)
Plante cu
fenotip sur2
(A)
Plants with
sur2
phenotype
(E)
Plante cu
fenotip
supresor (A)
Plants with
srs
phenotype
(E)
Raportul
de
segregare
Segregation
ratio
chi2
p-value
Semnifi-
cație
Significance
1788 158 158 100 116 42 118,5 39,5 3:1 0,21 0,7 ns
1844 131 131 100 116 15 98,25 32,75 3:1 12,83 < 0.001 ***
1848 128 128 100 92 36 96 32 3:1 0,67 0,5 ns
1932 150 150 100 120 30 112,5 37,5 3:1 2 0,157 ns
2059 183 183 100 138 45 137,25 45,75 3:1 0,016 0,898 ns
2146.1 146 146 100 131 15 109,5 36,5 3:1 16,88 0,001 ***
2146.4 144 144 100 114 30 108 36 3:1 1,33 0,3 ns
2344 212 212 100 171 41 159 53 3:1 3,62 0,057 ns
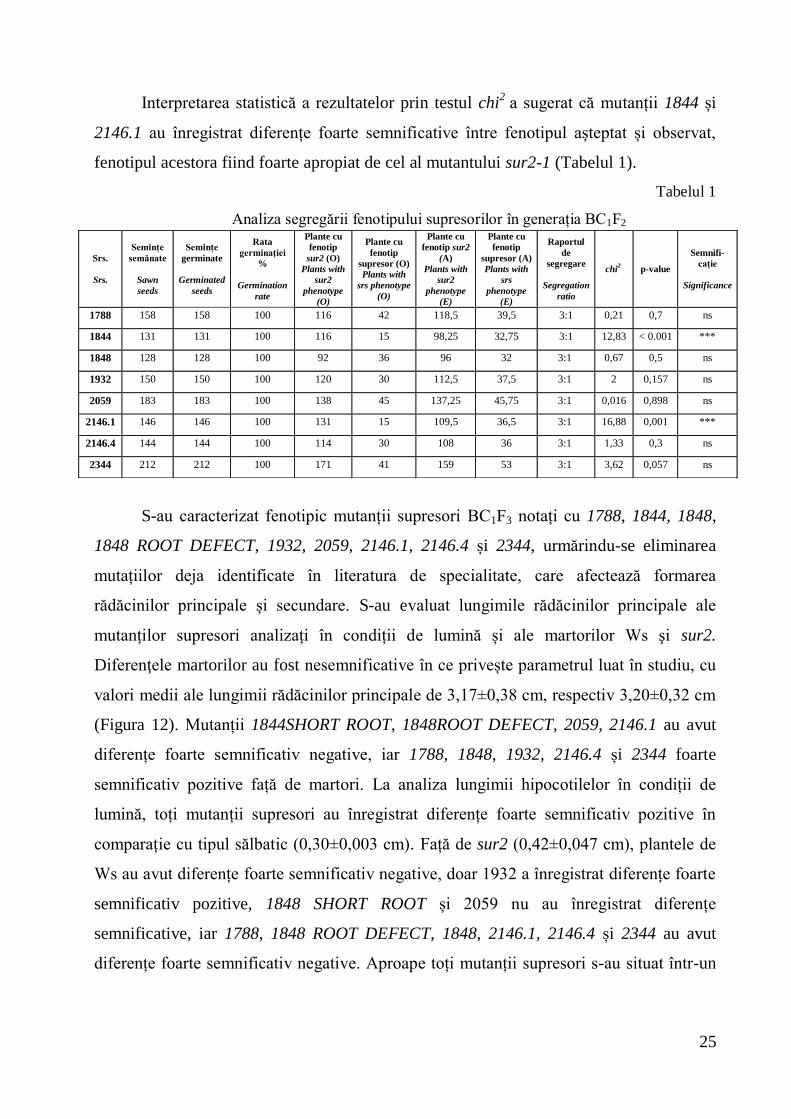
Interpretarea statistică a rezultatelor prin testul chi2
a sugerat că mutanții 1844 și
2146.1 au înregistrat diferențe foarte semnificative între fenotipul așteptat și observat,
fenotipul acestora fiind foarte apropiat de cel al mutantului sur2-1 (Tabelul 1).
Tabelul 1
Analiza segregării fenotipului supresorilor în generația BC1F2
S-au caracterizat fenotipic mutanții supresori BC1F3 notați cu 1788, 1844, 1848,
1848 ROOT DEFECT, 1932, 2059, 2146.1, 2146.4 și 2344, urmărindu-se eliminarea
mutațiilor deja identificate în literatura de specialitate, care afectează formarea
rădăcinilor principale și secundare. S-au evaluat lungimile rădăcinilor principale ale
mutanților supresori analizați în condiții de lumină și ale martorilor Ws şi sur2.
Diferenţele martorilor au fost nesemnificative în ce privește parametrul luat în studiu, cu
valori medii ale lungimii rădăcinilor principale de 3,17±0,38 cm, respectiv 3,20±0,32 cm
(Figura 12). Mutanții 1844SHORT ROOT, 1848ROOT DEFECT, 2059, 2146.1 au avut
diferențe foarte semnificativ negative, iar 1788, 1848, 1932, 2146.4 și 2344 foarte
semnificativ pozitive față de martori. La analiza lungimii hipocotilelor în condiții de
lumină, toți mutanții supresori au înregistrat diferențe foarte semnificativ pozitive în
comparație cu tipul sălbatic (0,30±0,003 cm). Față de sur2 (0,42±0,047 cm), plantele de
Ws au avut diferențe foarte semnificativ negative, doar 1932 a înregistrat diferențe foarte
semnificativ pozitive, 1848 SHORT ROOT și 2059 nu au înregistrat diferențe
semnificative, iar 1788, 1848 ROOT DEFECT, 1848, 2146.1, 2146.4 și 2344 au avut
diferențe foarte semnificativ negative. Aproape toți mutanții supresori s-au situat într-un
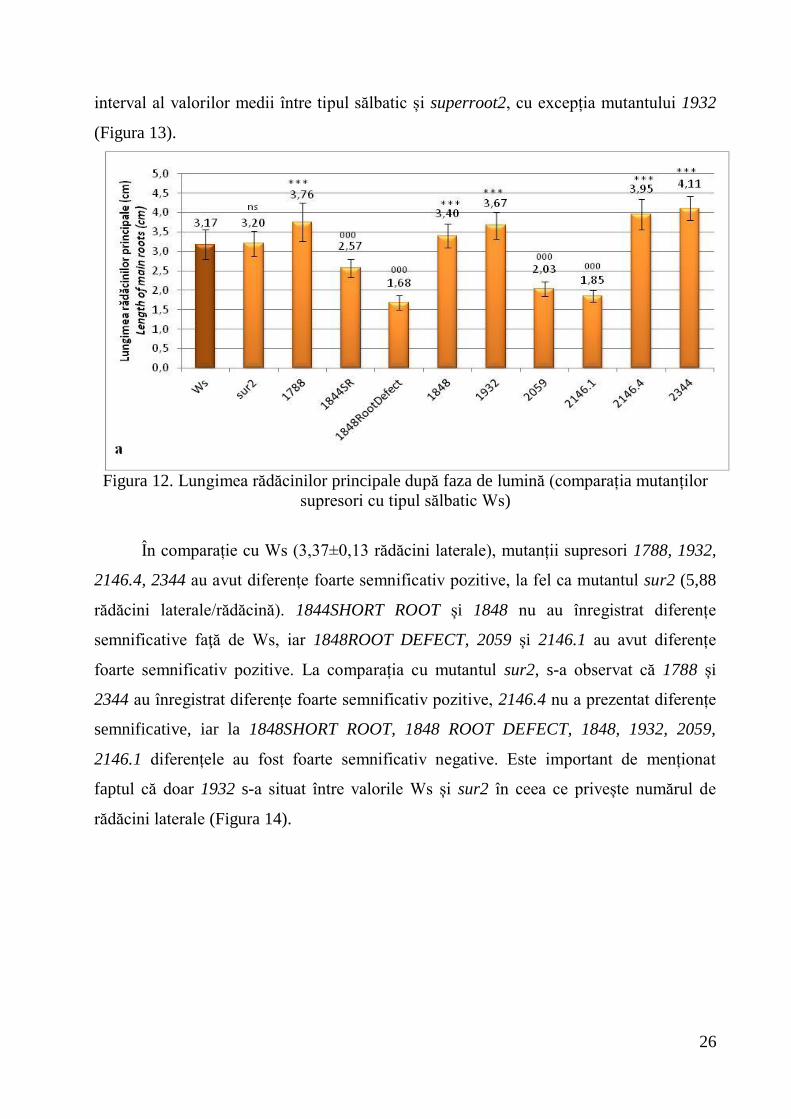
26
interval al valorilor medii între tipul sălbatic și superroot2, cu excepția mutantului 1932
(Figura 13).
Figura 12. Lungimea rădăcinilor principale după faza de lumină (comparația mutanților
supresori cu tipul sălbatic Ws)
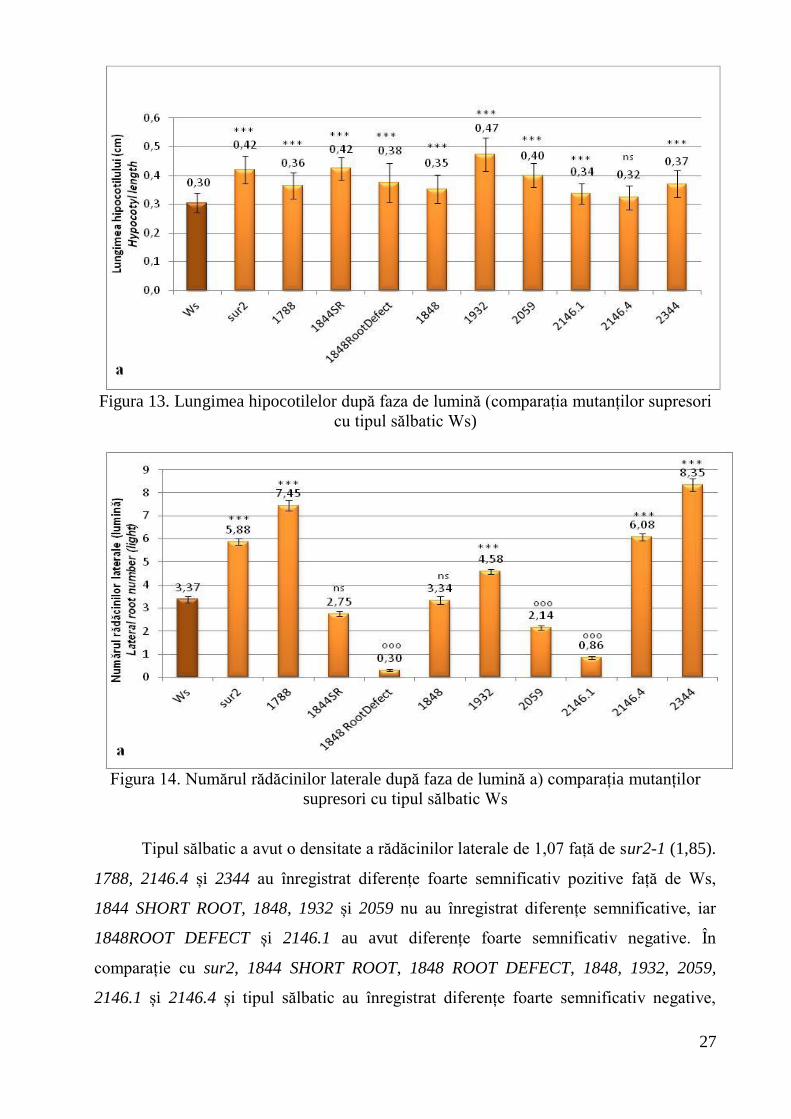
În comparație cu Ws (3,37±0,13 rădăcini laterale), mutanții supresori 1788, 1932,
2146.4, 2344 au avut diferențe foarte semnificativ pozitive, la fel ca mutantul sur2 (5,88
rădăcini laterale/rădăcină). 1844SHORT ROOT și 1848 nu au înregistrat diferențe
semnificative faţă de Ws, iar 1848ROOT DEFECT, 2059 și 2146.1 au avut diferențe
foarte semnificativ pozitive. La comparația cu mutantul sur2, s-a observat că 1788 și
2344 au înregistrat diferențe foarte semnificativ pozitive, 2146.4 nu a prezentat diferențe
semnificative, iar la 1848SHORT ROOT, 1848 ROOT DEFECT, 1848, 1932, 2059,
2146.1 diferențele au fost foarte semnificativ negative. Este important de menționat
faptul că doar 1932 s-a situat între valorile Ws și sur2 în ceea ce privește numărul de
rădăcini laterale (Figura 14).
27
Figura 13. Lungimea hipocotilelor după faza de lumină (comparația mutanților supresori
cu tipul sălbatic Ws)
Figura 14. Numărul rădăcinilor laterale după faza de lumină a) comparația mutanților
supresori cu tipul sălbatic Ws
Tipul sălbatic a avut o densitate a rădăcinilor laterale de 1,07 față de sur2-1 (1,85).
1788, 2146.4 și 2344 au înregistrat diferențe foarte semnificativ pozitive față de Ws,
1844 SHORT ROOT, 1848, 1932 și 2059 nu au înregistrat diferențe semnificative, iar
1848ROOT DEFECT și 2146.1 au avut diferențe foarte semnificativ negative. În
comparație cu sur2, 1844 SHORT ROOT, 1848 ROOT DEFECT, 1848, 1932, 2059,
2146.1 și 2146.4 și tipul sălbatic au înregistrat diferențe foarte semnificativ negative,
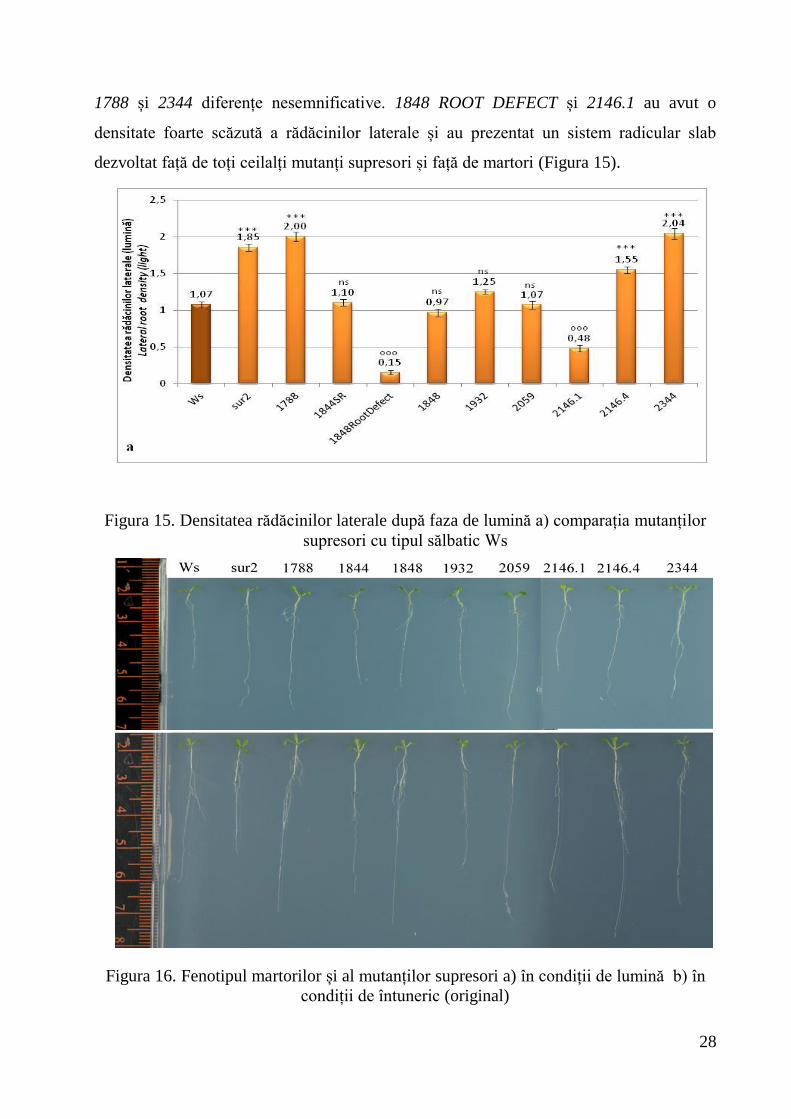
28
1788 și 2344 diferențe nesemnificative. 1848 ROOT DEFECT și 2146.1 au avut o
densitate foarte scăzută a rădăcinilor laterale și au prezentat un sistem radicular slab
dezvoltat față de toți ceilalți mutanți supresori și față de martori (Figura 15).
Figura 15. Densitatea rădăcinilor laterale după faza de lumină a) comparația mutanților
supresori cu tipul sălbatic Ws
Figura 16. Fenotipul martorilor și al mutanților supresori a) în condiții de lumină b) în
condiții de întuneric (original)
29
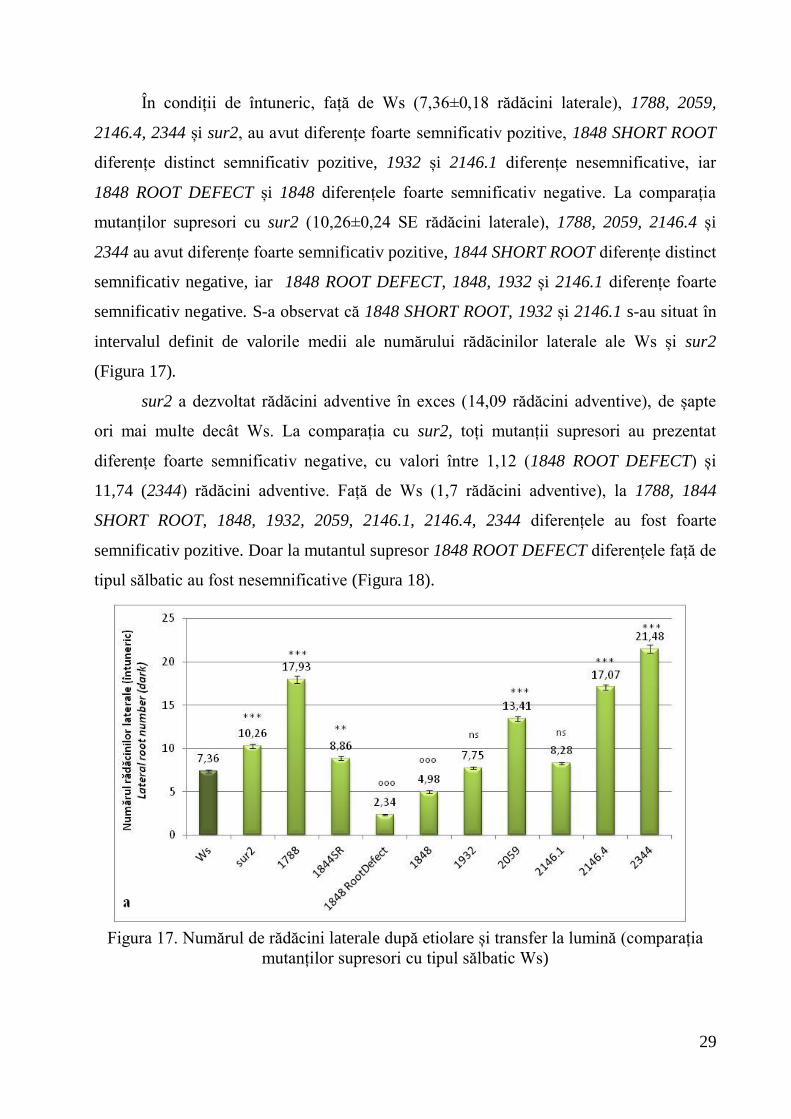
În condiții de întuneric, față de Ws (7,36±0,18 rădăcini laterale), 1788, 2059,
2146.4, 2344 și sur2, au avut diferențe foarte semnificativ pozitive, 1848 SHORT ROOT
diferențe distinct semnificativ pozitive, 1932 și 2146.1 diferențe nesemnificative, iar
1848 ROOT DEFECT și 1848 diferențele foarte semnificativ negative. La comparația
mutanților supresori cu sur2 (10,26±0,24 SE rădăcini laterale), 1788, 2059, 2146.4 și
2344 au avut diferențe foarte semnificativ pozitive, 1844 SHORT ROOT diferențe distinct
semnificativ negative, iar 1848 ROOT DEFECT, 1848, 1932 și 2146.1 diferențe foarte
semnificativ negative. S-a observat că 1848 SHORT ROOT, 1932 și 2146.1 s-au situat în
intervalul definit de valorile medii ale numărului rădăcinilor laterale ale Ws și sur2
(Figura 17).
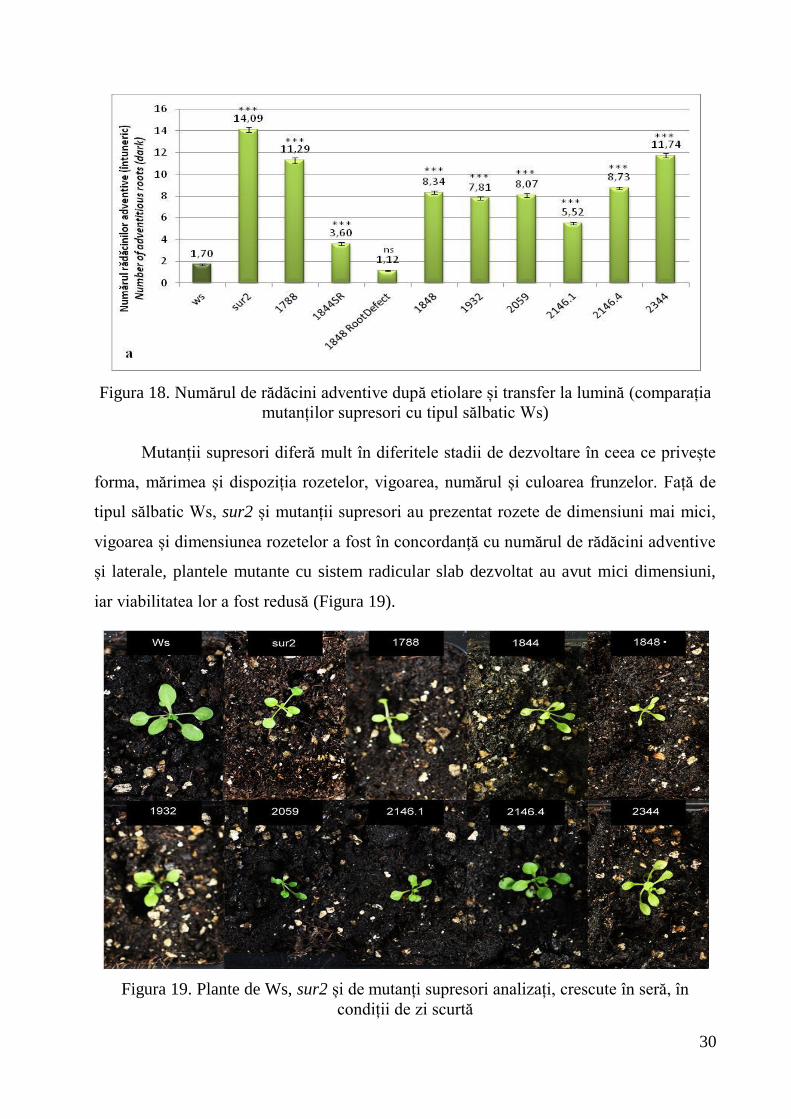
sur2 a dezvoltat rădăcini adventive în exces (14,09 rădăcini adventive), de șapte
ori mai multe decât Ws. La comparația cu sur2, toți mutanții supresori au prezentat
diferențe foarte semnificativ negative, cu valori între 1,12 (1848 ROOT DEFECT) și
11,74 (2344) rădăcini adventive. Față de Ws (1,7 rădăcini adventive), la 1788, 1844
SHORT ROOT, 1848, 1932, 2059, 2146.1, 2146.4, 2344 diferențele au fost foarte
semnificativ pozitive. Doar la mutantul supresor 1848 ROOT DEFECT diferențele față de
tipul sălbatic au fost nesemnificative (Figura 18).
Figura 17. Numărul de rădăcini laterale după etiolare și transfer la lumină (comparația
mutanților supresori cu tipul sălbatic Ws)
30
Figura 18. Numărul de rădăcini adventive după etiolare și transfer la lumină (comparația
mutanților supresori cu tipul sălbatic Ws)
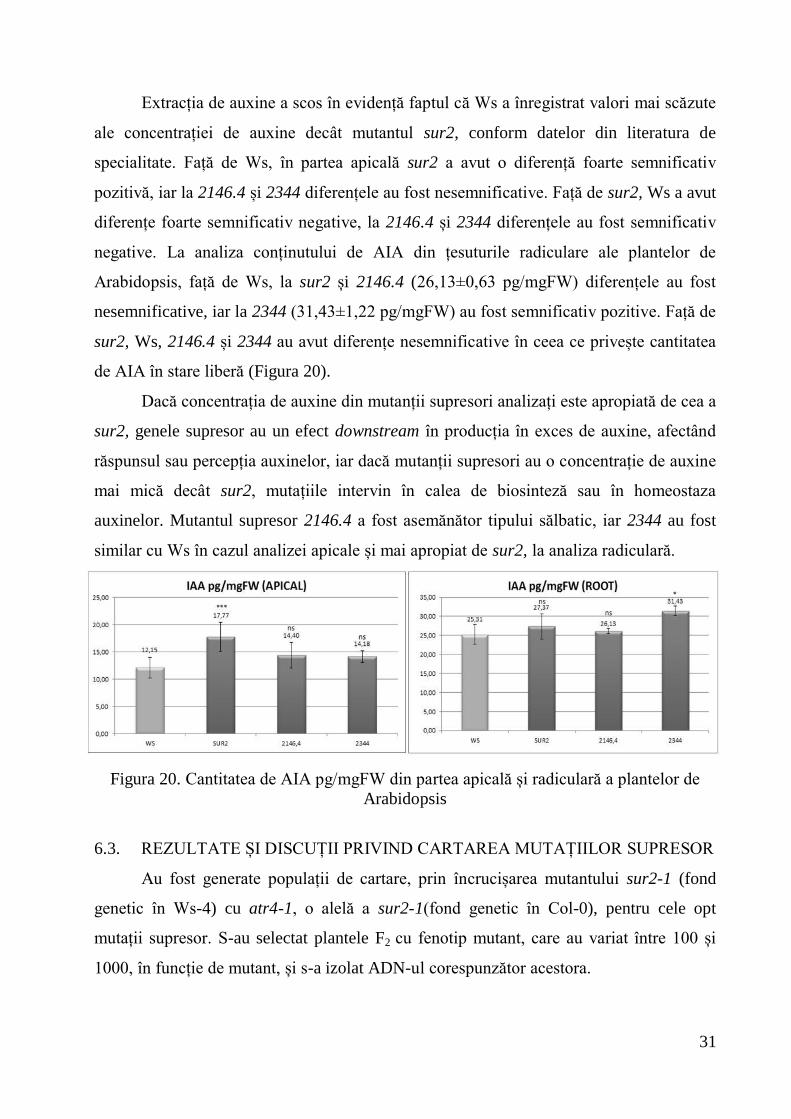
Mutanții supresori diferă mult în diferitele stadii de dezvoltare în ceea ce privește
forma, mărimea și dispoziția rozetelor, vigoarea, numărul și culoarea frunzelor. Față de
tipul sălbatic Ws, sur2 și mutanții supresori au prezentat rozete de dimensiuni mai mici,
vigoarea și dimensiunea rozetelor a fost în concordanță cu numărul de rădăcini adventive
și laterale, plantele mutante cu sistem radicular slab dezvoltat au avut mici dimensiuni,
iar viabilitatea lor a fost redusă (Figura 19).
Figura 19. Plante de Ws, sur2 și de mutanți supresori analizați, crescute în seră, în
condiții de zi scurtă
31
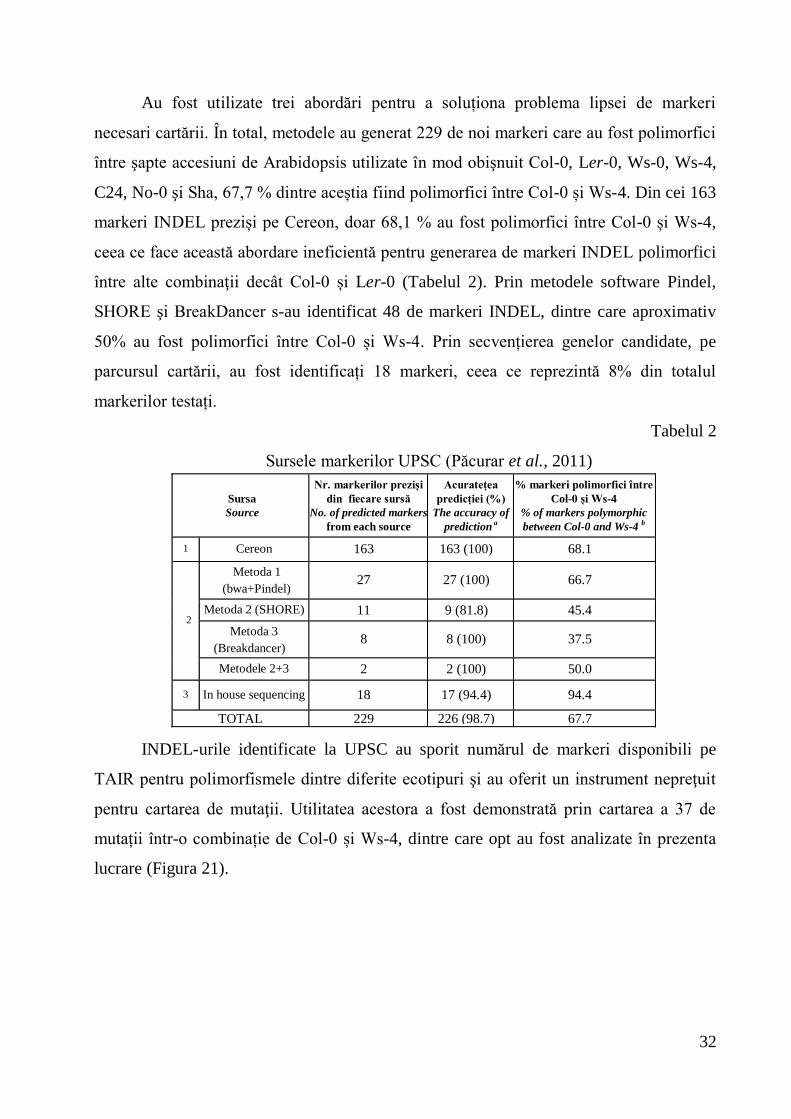
Extracția de auxine a scos în evidență faptul că Ws a înregistrat valori mai scăzute
ale concentrației de auxine decât mutantul sur2, conform datelor din literatura de
specialitate. Față de Ws, în partea apicală sur2 a avut o diferență foarte semnificativ
pozitivă, iar la 2146.4 și 2344 diferențele au fost nesemnificative. Față de sur2, Ws a avut
diferențe foarte semnificativ negative, la 2146.4 și 2344 diferențele au fost semnificativ
negative. La analiza conținutului de AIA din țesuturile radiculare ale plantelor de
Arabidopsis, față de Ws, la sur2 și 2146.4 (26,13±0,63 pg/mgFW) diferențele au fost
nesemnificative, iar la 2344 (31,43±1,22 pg/mgFW) au fost semnificativ pozitive. Față de
sur2, Ws, 2146.4 și 2344 au avut diferențe nesemnificative în ceea ce privește cantitatea
de AIA în stare liberă (Figura 20).
Dacă concentrația de auxine din mutanții supresori analizați este apropiată de cea a
sur2, genele supresor au un efect downstream în producția în exces de auxine, afectând
răspunsul sau percepția auxinelor, iar dacă mutanții supresori au o concentrație de auxine
mai mică decât sur2, mutațiile intervin în calea de biosinteză sau în homeostaza
auxinelor. Mutantul supresor 2146.4 a fost asemănător tipului sălbatic, iar 2344 au fost
similar cu Ws în cazul analizei apicale și mai apropiat de sur2, la analiza radiculară.
Figura 20. Cantitatea de AIA pg/mgFW din partea apicală și radiculară a plantelor de
Arabidopsis
6.3. REZULTATE ȘI DISCUȚII PRIVIND CARTAREA MUTAȚIILOR SUPRESOR
Au fost generate populații de cartare, prin încrucișarea mutantului sur2-1 (fond
genetic în Ws-4) cu atr4-1, o alelă a sur2-1(fond genetic în Col-0), pentru cele opt
mutații supresor. S-au selectat plantele F2 cu fenotip mutant, care au variat între 100 și
1000, în funcție de mutant, și s-a izolat ADN-ul corespunzător acestora.
32
Sursa
Source
Nr. markerilor preziși
din fiecare sursă
No. of predicted markers
from each source
Acuratețea
predicției (%)
The accuracy of
prediction a
% markeri polimorfici între
Col-0 și Ws-4
% of markers polymorphic
between Col-0 and Ws-4 b
1 Cereon 163 163 (100) 68.1
2
Metoda 1
(bwa+Pindel) 27 27 (100) 66.7
Metoda 2 (SHORE) 11 9 (81.8) 45.4
Metoda 3
(Breakdancer) 8 8 (100) 37.5
Metodele 2+3 2 2 (100) 50.0
3 In house sequencing 18 17 (94.4) 94.4
TOTAL 229 226 (98.7) 67.7
Au fost utilizate trei abordări pentru a soluționa problema lipsei de markeri
necesari cartării. În total, metodele au generat 229 de noi markeri care au fost polimorfici
între şapte accesiuni de Arabidopsis utilizate în mod obişnuit Col-0, Ler-0, Ws-0, Ws-4,
C24, No-0 şi Sha, 67,7 % dintre aceștia fiind polimorfici între Col-0 și Ws-4. Din cei 163
markeri INDEL preziși pe Cereon, doar 68,1 % au fost polimorfici între Col-0 și Ws-4,
ceea ce face această abordare ineficientă pentru generarea de markeri INDEL polimorfici
între alte combinaţii decât Col-0 și Ler-0 (Tabelul 2). Prin metodele software Pindel,
SHORE şi BreakDancer s-au identificat 48 de markeri INDEL, dintre care aproximativ
50% au fost polimorfici între Col-0 și Ws-4. Prin secvențierea genelor candidate, pe
parcursul cartării, au fost identificați 18 markeri, ceea ce reprezintă 8% din totalul
markerilor testați.
Tabelul 2
Sursele markerilor UPSC (Păcurar et al., 2011)
INDEL-urile identificate la UPSC au sporit numărul de markeri disponibili pe
TAIR pentru polimorfismele dintre diferite ecotipuri şi au oferit un instrument nepreţuit
pentru cartarea de mutaţii. Utilitatea acestora a fost demonstrată prin cartarea a 37 de
mutații într-o combinație de Col-0 și Ws-4, dintre care opt au fost analizate în prezenta
lucrare (Figura 21).

33
Figura 21. Poziția pe harta cromozomală a markerilor UPSC în genomul Col-0
(PĂCURAR et al., 2011)
Genele mutante au fost distribuite pe cuprinsul lungimilor cromozomilor, excepție
făcând regiunile din vecinătatea centromerilor, șase dintre acestea au fost localizate pe
brațul inferior al cromozomilor (1844, 1932, 2059, 2146.1, 2146.4 și 2344), iar 1844 a
fost localizată pe brațul superior al cromozomului. Trei dintre mutații au fost cartate pe
cromozomul II (1844, 1932 și 2344), două pe cromozomul V (2146.1 și 2146.4), una pe
cromozmomul I (1848), una pe cromozomul IV (2059), iar la mutația 1788 nu s-a putut
identifica cromozomul pe care aceasta este localizată (Figura 22).

34
Figura 22. Distribuția mutațiilor pe cromozomi (original)
Cei 22 markeri testați pentru cartarea mutației 1788, răspândiți pe cei cinci
cromozomi, nu au reușit să localizeze mutația. Datorită plantelor clasificate greșit ca
având fenotip mutant, existența contaminanților din populațiile de cartare a împiedicat
identificarea unor markeri linkați cu mutația.

35
Pentru cartarea mutației 1844, s-a utilizat o populație de cartare de 400 de indivizi
și 14 markeri INDEL, dintre care patru nou creați, răspândiți pe patru din cei cinci
cromozomi. Mutația a fost cartată pe cromozomul II, fiind flancată de markerii MSAT
2.7 și 2-UPSC_12898_M1, care definesc un interval genetic de 286 kb cu 105 gene
repartizate în 85 loci adnotați.
Mutația 1848 a fost cartată cu ajutorul unei populații de cartare de 300 indivizi F2
și a 13 markeri, din care șapte nou creați. Rezultatele au reușit să plaseze mutația pe
cromozomul I, între markerii 1-UPSC_13122 și MSAT 1.4, care definesc un interval de 1
044 kb, situat la nord de centromer, și care conține 206 gene aflate pe 200 loci.
Mutația 1932, cartată cu ajutorul a 200 de indivizi mutanți F2 și 15 markeri
INDEL, din care nouă nou creați, a fost localizată pe cromozomul II, în apropierea
mutației 1844. Aceasta a fost flancată de markerii 2-UPSC_11088 și MSAT 2.41, care
definesc un interval de 148 kb ce conține 52 gene distincte în 45 loci adnotați.
Cartarea mutației 2059 s-a realizat cu ajutorul unei populații de cartare de 100
plantule mutante și 12 markeri, din care opt nou creați. Mutația a fost localizată pe
cromozomul IV, între markerii 4-UPSC_13880 și 4-UPSC_14985, ce definesc un intrval
de 1 104 kb ce conține 329 loci adnotați cu 396 gene distincte.
Pentru cartarea mutației 2146.1 s-au utilizat 100 indivizi mutanți și 12 markeri, din
care 2 nou creați. Cartarea fină a localizat mutația pe cromozomul V, în partea terminală
a brațului lung, între markerii SSLP JV75/76 și JV58/59, de pe TAIR, într-un interval de
547 kb care include 178 loci adnotați și 225 gene distincte.
Cartarea mutației 2146.4 a necesitat o lungă perioadă de timp, iar populația de
cartare a fost crescută de la 100 la 800 indivizi mutanți din cauza pierderii
recombinanților. Au fost utilizați 27 markeri INDEL, din care 15 nou creați, care au
localizat mutația pe cromozomul V, între 5-UPSC_18641 și 5-UPSC_18642. Intervalul
genetic definit de aceștia conține două gene.
Mutația 2344 a fost cartată cu o populație de cartare de 200 de indivizi mutanți și
11 markeri INDEL, din care trei nou creați. În final, mutația a fost localizată în treimea
inferioară a cromozomului II, într-un intervalul de 1 189 kb delimitat de markerii 2-
UPSC_15109 și NGA347, cu 348 loci adnotați și 440 gene.
36
6.4. REZULTATE PRIVIND IDENTIFICAREA DE GENE CANDIDATE
CORESPUNZĂTOARE MUTANȚILOR ȘI SECVENȚIEREA ACESTORA
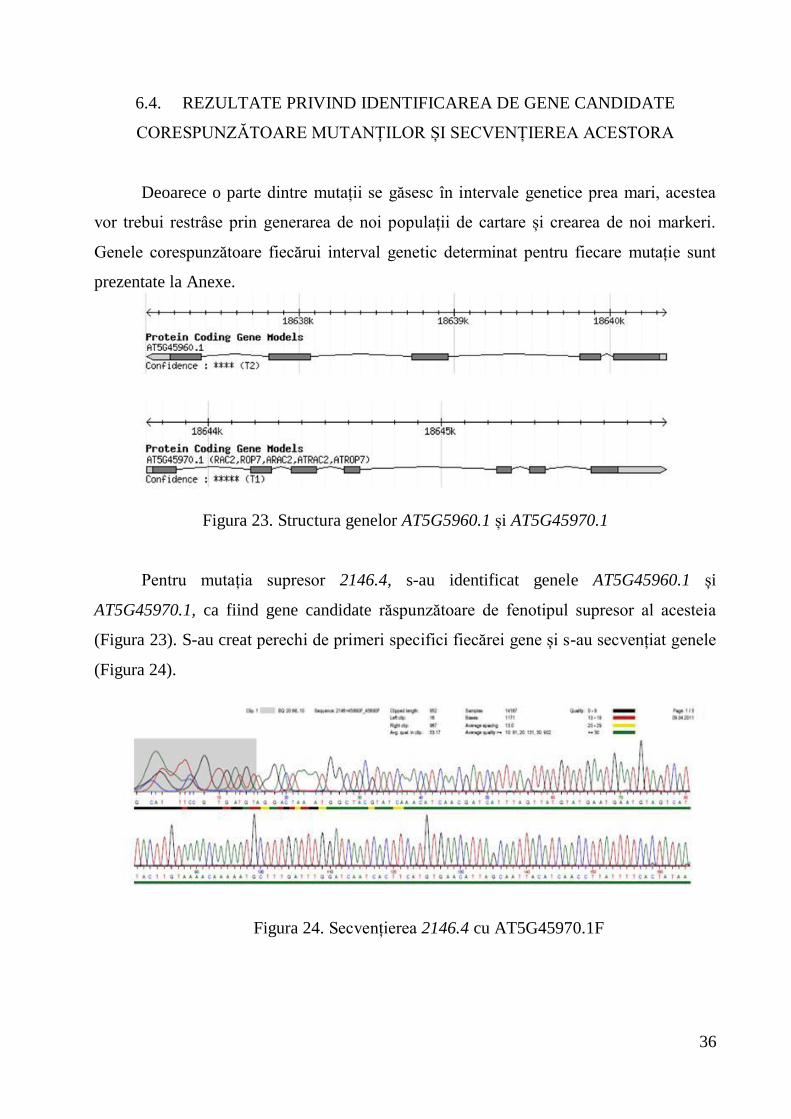
Deoarece o parte dintre mutații se găsesc în intervale genetice prea mari, acestea
vor trebui restrâse prin generarea de noi populații de cartare și crearea de noi markeri.
Genele corespunzătoare fiecărui interval genetic determinat pentru fiecare mutație sunt
prezentate la Anexe.
Figura 23. Structura genelor AT5G5960.1 și AT5G45970.1
Pentru mutația supresor 2146.4, s-au identificat genele AT5G45960.1 și
AT5G45970.1, ca fiind gene candidate răspunzătoare de fenotipul supresor al acesteia
(Figura 23). S-au creat perechi de primeri specifici fiecărei gene și s-au secvențiat genele
(Figura 24).
Figura 24. Secvențierea 2146.4 cu AT5G45970.1F

37
Secvențierea genei AT5G45970.1 a identificat o mutație punctiformă în mutantul
2146.4, care transformă C în T și determină schimbarea prolinei în leucină (Figura 25).
Cercetările viitoare vor confirma dacă gena secvențiată este responsabilă sau nu de
emiterea rădăcinilor adventive.
Figura 25. Alinierea secvențelor cu ClustalW și identificarea mutației

38
VII. CONCLUZII
Rezultatele obținute în urma cercetărilor sugerează faptul că prin secvenţierea
tuturor mutanţilor supresori se pot identifica gene noi supresoare ale SUR2, implicate în
formarea rădăcinilor adventive, crescând, astfel, gradul de importanță și noutate a temei
de cercetare.
Prin studiul speciei model Arabidopsis thaliana se va realiza transferul cu succes
al cunoştinţelor dobândite la speciile valoroase din punct de vedere economic.
Identificarea, clonarea şi caracterizarea genelor de interes la Arabidopsis va permite
elucidarea mecanismelor moleculare care guvernează procesul de formare a rădăcinilor
adventive.
Astfel, va fi posibilă identificarea de gene ortoloage la alte specii cu interes
economic ridicat, la care formarea rădăcinilor adventive reprezintă un factor limitativ.
Datele obţinute în urma secvenţierii şi clonării genelor supresor pot fi utilizate
pentru identificarea de markeri afați în linkage cu acestea, care să fie folosiți în
ameliorare. Astfel, găsirea unor markeri pentru genele care controlează înrădăcinarea
adventivă poate constitui o unealtă extrem de utilă în mâna amelioratorilor.
Apelând la rezerva genetică a speciilor, prin identificarea de gene implicate în
controlul formării rădăcinilor adventive, se poate realiza o selecție a combinaţiior
genetice favorabile unei bune înrădăcinări, menţinând în acelaşi timp celelalte
caracteristici de interes.

39
BIBLIOGRAFIE SELECTIVĂ
1.ALONSO, J.M. and J.R. ECKER (2006). Moving forward in reverse: genetic
technologies to enable genome-wide phenomic screens in Arabidopsis. Nat Rev
Genet 7(7): 524-536.
2.ALONSO-BLANCO, C., A.J.M. PEETERS, M. KOORNNEEF, C. LISTER, C. DEAN,
N. VAN DEN BOSCH, J. POT and M.T.R. KUIPER (1998). Development of an
AFLP based linkage map of Ler, Col and Cvi Arabidopsis thaliana ecotypes and
construction of a Ler/Cvi recombinant inbred line population. Plant Journal 14(2):
259-271.
3.ALONSO-BLANCO, C., M.G. AARTS, L. BENTSINK, J.J. KEURENTJES, M.
REYMOND, D. VREUGDENHIL and M. KOORNNEEF (2009). What has
natural variation taught us about plant development, physiology, and adaptation?
Plant Cell 21(7): 1877-1896.
4.BAK, S. and R. FEYEREISEN (2001). The involvement of two P450 enzymes,
CYP83B1 and CYP83A1, in auxin homeostasis and glucosinolate biosynthesis.
Plant Physiology 127(1): 108-118.
5.BARLIER, I., M. KOWALCZYK, A. MARCHANT, K. LJUNG, R. BHALERAO, M.
BENNETT, G. SANDBERG and C. BELLINI (2000). The SUR2 gene of
Arabidopsis thaliana encodes the cytochrome P450 CYP83B1, a modulator of
auxin homeostasis. Proc Natl Acad Sci U S A 97(26): 14819-14824.
6.BELL, C.J. and J.R. ECKER (1994). Assignment of 30 microsatellite loci to the linkage
map of Arabidopsis. Genomics 19(1): 137-144.
7.BIONDI, S., T. DIAZ, I. IGLESIAS, G. GAMBERINI and N. BAGNI (1990).
Polyamines and ethylene in relation to adventitious root-formation in Prunus
avium shoot cultures. Physiologia Plantarum 78(3): 474-483.
8.BOERJAN, W., M.T. CERVERA, M. DELARUE, T. BEECKMAN, W. DEWITTE, C.
BELLINI, M. CABOCHE, H.V. ONCKELEN, M.V. MONTAGU and D. INZE
(1995). superroot, a Recessive Mutation in Arabidopsis, Confers Auxin
Overproduction. The Plant Cell Online 7(9): 1405-1419.
9.CHANG, C. and E.M. MEYEROWITZ (1986). Molecular cloning and DNA-sequence of
the Arabidopsis thaliana alcohol-dehydrogenase gene. Proceedings of the
National Academy of Sciences of the United States of America 83(5): 1408-1412.
10.CHANG, C., J.L. BOWMAN, A.W. DEJOHN, E.S. LANDER and E.M.
MEYEROWITZ (1988). Restriction Fragment Length Polymorphism Linkage
map for Arabidopsis thaliana.
11.CHI, X.F., X.Y. LOU and Q.Y. SHU (2008). Progressive fine mapping in experimental
populations: an improved strategy toward positional cloning. J Theor Biol 253(4):
817-823.

40
12.CRISTEA, V., C. DELIU, B. OLTEAN, A. BUTIUC-KEUL, A. BRUMMER, C. ALBU
and G.L. RADU (2009). Soilless cultures for pharmaceutical use and biodiversity
conservation. Acta Hort. 843: 157-164
13.DAVIES, F.T., T.D. DAVIS and D.E. KESTER (1994). Commercial importance of
adventitious rooting to horticulture. Biology of adventitious root formation.
DAVIS, T.D. and HAISSIG B.E.
14.DELARUE, M., E. PRINSEN, H.V. ONCKELEN, M. CABOCHE and C. BELLINI
(1998). Sur2 mutations of Arabidopsis thaliana define a new locus involved in the
control of auxin homeostasis. Plant J 14(5): 603-611.
15.GEISS, G., L. GUTIERREZ and C. BELLINI (2009). Adventitious Root formation: new
insights and perspectives, Wiley-Blackwell.
16.GHIDRA, V., M. BOTU, R. SESTRAȘ and I. BOTU, Eds. (2004). Biodiversitate şi
Bioconservare. Cluj Napoca, Editura AcademicPres.
17.HAISSIG and DAVIS (1994). An historical evaluation of adventitious rooting research to
1993 In: Davis T.D., Haissig B.E., eds. Biology of adventitious root formation.
New York, London: Plenum Publishing Corporation: 275-331.
18.HAN, H., S. ZHANG and X. SUN (2009). A review on the molecular mechanism of
plants rooting modulated by auxin. African Journal of Biotechnology 8(3): 348-
353.
19.HAUSMAN, J.F., C. KEVERS and T. GASPAR (1994). Involvement of putrescine in the
inductive rooting phase of poplar shoots raised in vitro. Physiologia Plantarum
92(2): 201-206.
20.HAUSMAN, J.F., C. KEVERS and T. GASPAR (1995). Auxin-Polyamine interaction in
the control of the rooting inductive phase of poplar shoots in vitro. Plant Science
110(1): 63-71.
21.HELOIR, M.C., C. KEVERS, J.F. HAUSMAN and T. GASPAR (1996). Changes in the
concentrations of auxins and polyamines during rooting of in vitro-propagated
walnut shoots. Tree Physiol 16(5): 515-519.
22.HOCHHOLDINGER, F. and R. ZIMMERMANN (2008). Conserved and diverse
mechanisms in root development. Current Opinion in Plant Biology 11(1): 70-74.
23.HOU, X.H., L.C. LI, Z.Y. PENG, B.Y. WEI, S.J. TANG, M.Y. DING, J.J. LIU, F.X.
ZHANG, Y.D. ZHAO, H.Y. GU and L.J. QU (2010). A platform of high-density
INDEL/CAPS markers for map-based cloning in Arabidopsis. Plant Journal
63(5): 880-888.
24.JANDER, G., Ed. (2006). Gene Identification and cloning by molecular marker mapping.
Gene identification and cloning by molecular marker mapping.
25.JANDER, G., S.R. NORRIS, S.D. ROUNSLEY, D.F. BUSH, I.M. LEVIN and R.L.
LAST (2002). Arabidopsis map-based cloning in the post-genome era. Plant
Physiology 129(2): 440-450.

41
26.KONIECZNY, A. and F.M. AUSUBEL (1993). A procedure for mapping arabidopsis
mutations using codominant ecotype-specific pcr-based markers. Plant Journal
4(2): 403-410.
27.LI, S.-W., L. XUE, S. XU, H. FENG and L. AN (2009). Mediators, genes and signaling
in adventitious rooting. The Botanical Review 75(2): 230-247.
28.LI, S.W., L.G. XUE, S.J. XU, H.Y. FENG and L.Z. AN (2007). Hydrogen peroxide
involvement in formation and development of adventitious roots in cucumber.
Plant Growth Regulation 52(2): 173-180.
29.LUKOWITZ, W., C.S. GILLMOR and W.R. SCHEIBLE (2000). Positional cloning in
Arabidopsis. Why it feels good to have a genome initiative working for you. Plant
Physiology 123(3): 795-805.
30.MEINKE, D.W., J.M. CHERRY, C. DEAN, S.D. ROUNSLEY and M. KOORNNEEF
(1998). Arabidopsis thaliana: A model plant for genome analysis. Science
282(5389).
31.OPREA, A., Ed. (2005). Lista critică a plantelor vasculare din România. 2009 3rd
International Conference on Bioinformatics and Biomedical Engineering, Vols 1-
11. Iaşi., Editura Universitaţii „Al.I.Cuza”.
32.PAGNUSSAT, G.C., M. SIMONTACCHI, S. PUNTARULO and L. LAMATTINA
(2002). Nitric oxide is required for root organogenesis. Plant Physiology 129(3):
954-956.
33.PAGNUSSAT, G.C., M.L. LANTERI and L. LAMATTINA (2003). Nitric oxide and
cyclic GMP are messengers in the indole acetic acid-induced adventitious rooting
process. Plant Physiology 132(3): 1241-1248.
34.PAGNUSSAT, G.C., M.L. LANTERI, M.C. LOMBARDO and L. LAMATTINA
(2004). Nitric oxide mediates the indole acetic acid induction activation of a
mitogen-activated protein kinase cascade involved in adventitious root
development. Plant Physiology 135(1): 279-286.
35.PAN, R.C., J.X. WANG and X.S. TIAN (2002). Influence of ethylene on adventitious
root formation in mung bean hypocotyl cuttings. Plant Growth Regulation 36(2):
135-139.
36.PAPAFOTIOU, M. and J.L. STRAGAS (2009). Seed germination and in vitro
propagation of Dianthus fructicossus. Acta Hort. (ISHS) 813: 481-484.
37.PARRY, G. and M. ESTELLE (2006). Auxin receptors: a new role for F-box proteins.
Current Opinion in Cell Biology 18(2): 152-156.
38.PĂCURAR, D., M. PĂCURAR, N. STREET, J. BUSSELL, T.I. POP, L. GUTIEREZ
and C. BELLINI (2011). A collection of INDEL markers for map-based cloning in
seven Arabidopsis accessions.
39.PETERS, J.L., F. CNUDDE and T. GERATS (2003). Forward genetics and map-based
cloning approaches. Trends in Plant Science 8(10): 484-491.

42
40.POP, T.I. (2011). The Importance of biodiversity preservation in Romania. Bulletin
UASVM Horticulture 68(1).
41.POP, T.I. and D. PAMFIL (2011). In vitro Preservation of Three Species of Dianthus
from Romania. Bulletin UASVM Horticulture 68(1).
42.POP, T.I., D. PAMFIL and C. BELLINI (2011). Auxin control in the formation of a
adventitious roots. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 39(1):307-
316.
43.PRELICH, G. (1999). Suppression mechanisms: themes from variations. Trends in
Genetics 15(7): 261-266.
44.QUINT, M. and W.M. GRAY (2006). Auxin signaling. Current Opinion in Plant Biology
9(5): 448-453.
45.SOMERVILLE, C. and M. KOORNNEEF (2002). Timeline - A fortunate choice: the
history of Arabidopsis as a model plant. Nature Reviews Genetics 3(11): 883-889.
46.SORIN, C., J.D. BUSSELL, I. CAMUS, K. LJUNG, M. KOWALCZYK, G. GEISS, H.
MCKHANN, C. GARCION, H. VAUCHERET, G. SANDBERG and C.
BELLINI (2005). Auxin and light control of adventitious rooting in Arabidopsis
require ARGONAUTE1. Plant Cell 17(5): 1343-1359.
47.TAKAHASHI, F., K. SATO-NARA, K. KOBAYASHI, M. SUZUKI and H. SUZUKI
(2003). Sugar-induced adventitious roots in Arabidopsis seedlings. Journal of
Plant Research 116(2): 83-91.
48.WIESMAN, Z., J. RIOV and E. EPSTEIN (1988). Comparison of movement and
metabolism of indole-3-acetic-acid and indole-3-butyric acid in mung bean
cuttings. Physiologia Plantarum 74(3): 556-560.
49.WILLIAMS, J.G.K., A.R. KUBELIK, K.J. LIVAK, J.A. RAFALSKI and S.V. TINGEY
(1990). DNA polymorphisms amplified by arbitrary primers are useful as genetic-
markers. Nucleic Acids Research 18(22): 6531-6535.
50.WOODWARD, A.W. and B. BARTEL (2005). Auxin: regulation, action, and
interaction. Ann Bot 95(5): 707-735.
51.XU, J., W. XUAN, B.K. HUANG, Y.H. ZHOU, T.F. LING, S. XU and W.B. SHEN
(2006). Carbon monoxide-induced adventitious rooting of hypocotyl cuttings from
mung bean seedling. Chinese Science Bulletin 51(6): 668-674.