cromozomi
DESCRIPTION
geneticaTRANSCRIPT

285
ORGANIZAREA GENOMULUI UMAN
L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA Institutul de Genetica al Universitatii din Bucuresti Aleea Portocalilor Nr.1-3 Sector 6 Bucuresti
Introducere
Termenul “genom” a fost introdus in anul 1920 de catre botanistul german Hans Winkler, pentru a desemna setul haploid de cromozomi al unui organism eucariot. Termenul a fost extins pentru a se putea folosi si la organismele non-eucariote, genomul desemnand in acest caz cantitatea de material genetic ce defineste zestrea ereditara a unui organism. Genomul nu este insa doar o suma a partilor sale componente, datorita faptului ca diferite tipuri de interactiuni, functionale, evolutive, apar intre diferitele parti ale genomului, mai ales intre secventele sale codificatoare si cele necodificatoare.
Genomul uman poate fi impartit in doua componente: genomul nuclear si genomul mitocondrial. Cele doua componente au putine lucruri in comun, atat in ceea ce priveste organizarea cat si in ceea ce priveste expresia genelor. In fapt, cele doua componente pot fi privite ca fiind doua sisteme genetice separate si independente. In cele ce urmeaza ne vom referi doar la componenta nucleara a genomului uman.
Dimensiune
Dimensiunea totala a genomului nuclear uman este estimata, in urma ultimelor descoperiri realizate in cadrul Proiectului Genomului Uman, la aproximativ 3000Mb, dimensiune conforma, de altfel, cu estimarile anterioare. Genomul nuclear este impartit intr-un numar de molecule ADN individuale, fiecare dintre acestea fiind continuta si formand un cromozom diferit. Aceste molecule de ADN sunt liniare. In majoritatea celulelor din organismul uman exista doua copii ale fiecarui cromozom, deci doua copii ale fiecarei gene. Acest ansamblu formeaza ceea ce este denumit complementul diploid; termenul haploid se refera la situatia intalnita de obicei in celulele germinale, unde nucleul contine o singura copie a fiecarui cromozom.
Cromozomi
Genomul uman este impartit in 23 cromozomi, fiecare continand deci o singura molecula de ADN liniara, dublu-catenara, cu dimensiunea cuprinsa intre 51Mb si 279Mb (vezi Tabelul 1).

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
286
Tabelul 1. Dimensiunile cromozomilor umani
Cromozomul Dimensiune (Mb) 1 279
2 251
3 221
4 197
5 198
6 176
7 163
8 148
9 140
10 143
11 148
12 142
13 118
14 107
15 100
16 104
17 88
18 86
19 72
20 66
21 45
22 48
X 163
Y 51
Daca genomul uman ar fi fost reprezentat de o singura molecula de ADN, lungimea totala a acesteia ar fi fost de aproximativ 1m. Cum cantitatea de ADN din celula umana este impartita intre 23 de cromozomi, lungimea medie a acestora ar trebui sa fie de aproximativ 5cm. Totusi lungimea cromozomilor umani este de ordinul micrometrilor. Trebuie sa existe deci un sistem inalt organizat de impachetare a unei molecule de o asemenea lungime intr-o structura atat de mica.
La inceputul secolului, citologii au propus termenul cromatina pentru a desemna componenta cromozomului care se coloreaza mai puternic cu coloranti specifici pentru cromozom. Astazi, prin cromatina se intelege complexul format prin asocierea intre molecula de ADN continuta in cromozom si proteinele legate de aceasta, proteine responsabile de impachetarea ADN intr-o structura regulata in interiorul cromozomului.

ORGANIZAREA GENOMULUI UMAN
287
Cromozomii sunt formati aproximativ 50% din ADN si 50% din proteine. Din componenta proteica, o portiune este reprezentata de o mixtura heterogena de molecule, incluzand ADN si ARN polimeraze si proteine reglatoare. Restul de proteine, asociate mai intim cu componenta ADN a cromozomilor, este reprezentat de un grup de proteine numite histone, denumite respectiv, H1, H2A, H2B, H3 si H4. Histonele sunt proteine cu un pronuntat caracter bazic, fiind inalt conservate intre diferite specii.
In anul 1973 s-a demonstrat ca in urma digestiei cromatinei purificate cu o endonucleaza (enzima care taie molecula de ADN la nivelul legaturilor fosfodiesterice interne) si extractiei ADN-ului, fragmentele obtinute nu sunt de dimensiuni randomice, asa cum ar fi fost previzibil, ci sunt multimeri de aproximativ 200bp lungime. Acest experiment arata ca proteinele din cromatina sunt asociate cu ADN-ul intr-un mod regulat si ca aceste complexe proteice protejeaza molecula ADN de atacul endonucleazelor. Aceste rezultate au fost completate anul urmator de prima observare in microscopie electronica a fibrei de cromatina, supusa unei noi metode preparative. La microscopul electronic s-au observat aranjamente liniare de structuri sferice car au fost numite nucleosomi. Acestia sunt formati dintr-un miez histonic (octamer) format din cate 2 molecule din histonele H2A, H2B, H3 si H4, miez in jurul caruia se afla infasurata molecula de ADN. Exact 146 nucleotide formeaza portiunea de ADN implicata in edificarea nucleosomului, portiune protejata deci de atacul endonucleazelor. Singura portiune din molecula de ADN accesibila endonucleazelor este reprezentata de ADN linker (50-70bp) care uneste nucleosomii intre ei. Taierea moleculei de ADN la nivelul acestui ADN linker va genera fragmente de 200bp.
Figura 1. Structura cromatinei

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
288
Impachetarea ADN in nucleosomi, reduce lungimea moleculei de acid nucleic de aproximativ 6 ori. Deci exista un alt nivel de organizare a cromatinei pentru a se putea ajunge la dimensiunea reala a cromozomilor.
Nucleosomii se asociaza pentru a forma fibra de cromatina cu diametrul de 30nm. Al doilea nivel de organizare a fibrei de cromatina a fost caracterizat intre anii 1977-1980 de catre Aaron Klug prin observatii de microscopie electronica, observatii care au revelat o fibra de cromatina, mai densa decat simpla insiruire a nucleosomilor, fibra cu un diametru de aproximativ 30nm. Nucleosomii sunt aranjati intr-o structura numita solenoid, pentru a da nastere fibrei de cromatina. Aceasta maniera de impachetare reduce dimensiunea cromozomilor de aproximativ 7 ori, ceea ce ar face ca dimensiunea cromozomilor sa fie de aproximativ 1.4mm. In celulele care se divid, intervine un al treilea nivel de impachetare a fibrei de cromatina, prin formarea cromozomilor metafazici. Aceasta conformatie reprezinta o reminiscenta a nucleoidului bacterian. Pentru formarea acestei structuri, fibra de cromatina formeaza bucle superrasucite care emerg dintr-un miez proteic de natura non-histonica. Aproximativ 85kb ADN sunt continute in fiecare bucla. Dimensiunea acestei structurii este de aproximativ 0.75�m, in acord cu dimensiunea cromozomului metafazic, sugerand ca acesta este cel mai inalt nivel structural de organizare prezentat de cromozomul eucariot, deci si uman.
Aspectul tipic al unui cromozom metafazic este prezentat in figura 1. In realitate este vorba aici de doi cromozomi uniti la nivelul unei structuri numite centromer. Pozitia centromerului este caracteristica pentru un cromozom dat si este unul din criteriile utilizate pentru a distinge membrii cariotipului (intregul complement cromozomal din nucleu). In cromozomul metafazic, fiecare din cei doi cromozomi uniti la nivelul centromerului se numeste cromatida, aspectul acestora fiind un alt caracter utilizat in identificarea unui cromozom dat. Acest lucru se datoreaza faptului ca in urma unor coloratii specifice, cromatidele diferitilor cromozomi nu se coloreaza identic, aparand un model de benzi pozitive si negative (pentru coloratia respectiva) caracteristic fiecarei cromatide, deci fiecarui cromozom metafazic.
Cromozomii umani au fost repartizati in 7 grupe morfologice la Conferinta de la Denver (Colorado) din anul 1960. Caracterizarea morfologica a cromozomilor mitotici a fost completata ulterior la Conferintele de la Londra (1963), Chicago (1971) si Paris (1972) consacrate cariotipului uman.
Pe baza lungimii relative a cromozomilor, indicelui centromeric, raportului bratelor, prezentei satelitilor si a constrictiilor secundare, a sincroniei replicarii si modelului de bandare, cei 46 de cromozomi au fost repartizati in urmatoarele grupe morfologice ale cariotipului uman:
1. Grupa A (1-3). Cuprinde cromozomi mari, dintre care 1 si 3 au centromerul situat median, iar cei din perechea 2 submedian. Cromozomii din perechea 1 prezinta o constrictie secundara in regiunea proximala a bratului q.
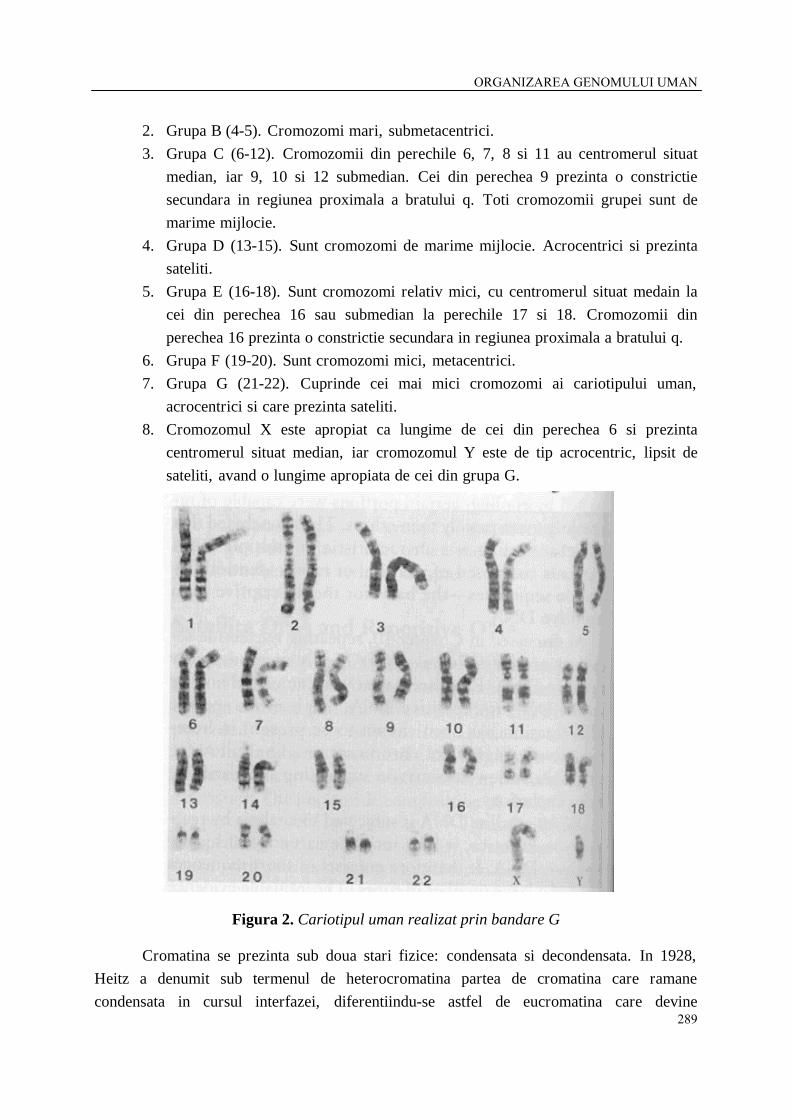
ORGANIZAREA GENOMULUI UMAN
289
2. Grupa B (4-5). Cromozomi mari, submetacentrici. 3. Grupa C (6-12). Cromozomii din perechile 6, 7, 8 si 11 au centromerul situat
median, iar 9, 10 si 12 submedian. Cei din perechea 9 prezinta o constrictie secundara in regiunea proximala a bratului q. Toti cromozomii grupei sunt de marime mijlocie.
4. Grupa D (13-15). Sunt cromozomi de marime mijlocie. Acrocentrici si prezinta sateliti.
5. Grupa E (16-18). Sunt cromozomi relativ mici, cu centromerul situat medain la cei din perechea 16 sau submedian la perechile 17 si 18. Cromozomii din perechea 16 prezinta o constrictie secundara in regiunea proximala a bratului q.
6. Grupa F (19-20). Sunt cromozomi mici, metacentrici. 7. Grupa G (21-22). Cuprinde cei mai mici cromozomi ai cariotipului uman,
acrocentrici si care prezinta sateliti. 8. Cromozomul X este apropiat ca lungime de cei din perechea 6 si prezinta
centromerul situat median, iar cromozomul Y este de tip acrocentric, lipsit de sateliti, avand o lungime apropiata de cei din grupa G.
Figura 2. Cariotipul uman realizat prin bandare G
Cromatina se prezinta sub doua stari fizice: condensata si decondensata. In 1928, Heitz a denumit sub termenul de heterocromatina partea de cromatina care ramane condensata in cursul interfazei, diferentiindu-se astfel de eucromatina care devine

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
290
condensata. Ulterior s-a propus ca termenul de heterocromatina sa fie utilizat doar pentru desemnarea heterocromatinei constitutive sau cariotipice, caracterizata printr-un continut inalt de ADN repetitiv. Termenul de heterocromatina facultativa trebuie utilizat numai pentru a desemna cromozomul X hetropicnotic, inactivat, al femelelor la mamifere. In cazul genomului uman, heterocromatina formeaza aproximativ 6.5% din totalul genomului.
Tipuri de secvente
Genomul uman poate fi impartit in ADN genic si ADN extragenic. Prin ADN extragenic intelegem orice secventa ADN care nu este continuta intr-o gena (sub forma de exoni sau introni), nu este asociata cu o gena (secvente leader, trailer, promotor, situsuri de reglare) si care nu este recunoscuta ca provenind dintr-o gena (pseudogene, fragmente genice). Orice ADN care nu se incadreaza in aceste categorii este extragenic. ADN-ul extragenic reprezinta aproximativ 70-80% din intregul genom uman.
Majoritatea ADN extragenic consta din secvente unice sau aflate in numar mic de repetitii. Restul, aproximativ 0.7Mb, este format din ADN moderat si inalt repetitiv, fie dispersat in genom, fie aglomerat in clustere.
ADN repetitiv dispersat Secvente SINE (Short Interspersed Nuclear Elements). Cel mai cunoscut exemplu
de SINE este reprezentat de familia Alu. Membrii acestei familii prezinta o dimensiune de aproximativ 250bp, aceste secvente fiind repetate de aproximativ 700000 in intregul genom, inclusiv in introni. Se pare ca aceste secvente poseda activitate de transpozoni, permitandu-le sa se replice si sa se deplaseze in noi locuri in genom.
Figura 3. Secvente SINE
Secvente LINE (Long Interspersed Nuclear Elements). Si aceste secvente se pare ca s-au propagat prin transpozitii. Secventele de tip LINE-1 (aproximativ 60000 copii) reprezinta un retroelement de tip non-viral, un transpozon care se poate replica si deplasa in genom printr-un proces care implica revers-transcrierea.
Figura 4. Secvente LINE

ORGANIZAREA GENOMULUI UMAN
291
ADN repetitiv grupat in clusteri Genomul uman contine regiuni intinse formate din secvente repetate aranjate in
tandem. Acest tip de ADN este numit ADN satelit si este impartit in trei categorii, in functie de dimensiunea clusterilor:
1. ADN satelit clasic Acesta a fost primul tip de ADN satelit identificat in genomul uman si consta in
regiuni de 100-5000kb lungime. Clusterle de lanivelul centromerului formate din repetiiile Alphoid reprezinta exemplul clasic de ADN satelit.
2. ADN minisatelit ADN minisatelit formeaza clustere de 100bp-20kb lungime. Telomerele (10-15kb)
sunt clasificate ca ADN minisatelit. 3. ADN microsatelit Dimensiunea clusterilor este aceeasi ca in cazul minisatelitilor, dar in cazul
microsatelitilor unitatile repetitive sunt rareori mai mari de 4bp. Secvente dinucleotidice, de tipul 5’-CACACACACACACACACACA-3’, sunt foarte frecvente in genomul uman, reprezentand aproximativ 0.5% din total. Monorepetitii de tipul 5’-AAAAAAAAAAAAAAAA-3’ formeaza inca 0.3% din totalul genomului.
Familii de gene - caracterizare
O familie multigenica este un cluster de gene inrudite, gene care prezinta secventa identica sau similara de nucleotide si deci care contin informatie identica sau inrudita. Genele cu secventa de nucleotide similara sunt numite gene omoloage.
Exista doua tipuri de familii multigenice: 1. Familii multigenice simple, in care toate genele sunt identice. De exemplu,
familia de gene pentru ARN ribozomal 5S, reprezentata la om de aproximativ 2000 de copii; acest numar mare este probabil datorat necesitatii de cantitati mari de produs genic in acelasi moment de timp, cantitate pe care o singura copie sau doar cateva copii ale genei respective nu ar fi putut sa o satisfaca.
2. Familii multigenice complexe, formate din gene similare, dar nu identice. Exemplul cel mai bun este reprezentat de genele pentru globine la toate vertebratele, inclusiv la om.
Genomul uman ilustreaza toate cele trei modalitati posibile in care familiile multigenice pot fi organizate:
1. In unele cazuri genele individuale sunt localizate impreuna, formand un cluster genic, intr-un singur loc in genom. Exemple sunt familia de gene pentru hormonul de crestere, ale carui 5 gene sunt localizate pe cromozomul 17 si familia de gene pentru ARN ribozomal 5S, care cuprinde 2000 de gene repetate in tandem pe bratul lung al cromozomului 1.

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
292
2. In cazul altor familii genice, genele sunt raspandite in genom. De exemplu cei 5 membri ai familiei genei aldolazei sunt localizati pe cromozomii 3, 9, 10, 16 si 17.
3. Unele familii genice mari sunt atat raspandite in genom cat si organizate in clusteri genici, in acelasi timp. Astfel, exista aproximativ 280 de copii pentru unitatea transcriptionala a ARN ribozomal, grupate in 5 clusteri de cate 50-70 unitati fiecare, clusteri localizati pe bratele scurte ale cromozomilor 13, 14, 15, 21 si 22.
Sa analizam acum cateva astfel de familii genice pentru o mai buna intelegere a organizarii lor.
Familia de gene pentru globine umane La om, patru catene polipeptidice, 2� si 2� sunt combinate cu un cofactor hem pentru
a forma o singura molecula de hemoglobina, proteina raspunzatoare de transportul oxigenului in torentul sanguin. Fiecare tip de polipeptid care intra in componenta hemoglobinei, catenele � si �, exista ca o familie de molecule inrudite, care difera una de cealalta prin cativa aminoacizi. Astfel, la om exista de fapt doua molecule de tip �, � si �, si cinci molecule de tip �, respectiv �, �, A�, G� si �. Tipul de globina din sange depinde de stadiul de dezvoltare a organismului, cu tipurile � si �, de exemplu, gasite doar in stadiile embrionare. Genele pentru globinele umane au fost printre primele studiate cu ajutorul tehnologiei ADN recombinant. Tom Maniatis si colaboratorii sai au descoperit la sfarsitul anilor 1970, ca genele pentru globinele umane sunt organizate intr-o familie multigenica pentru globina � pe cromozomul 16 si o familie multigenica pentru globina � pe cromozomul 11. Analize ulterioare au permis stabilirea cu precizie a pozitiei fiecarei gene in cadrul familiei.

ORGANIZAREA GENOMULUI UMAN
293
Figura 5. Familiile de gene pentru globinele umane
Familiile genelor pentru globine ilustreaza un alt aspect al organizarii genelor: pseudogenele. Astfel, clusterii pentru globinele de tip � si � includ, in afara genelor deja prezentate, genele ��1, ��1, ��2 si ��1. Aceste “gene” sunt foarte asemanatoare celorlalti membri ai familiei, dar informatia codificata nu mai este activa.
Izolarea, secventierea, caracterizarea unor gene umane: gena pentru maladia Huntington, gena pentru mucoviscidoza.
Maladia Huntington Maladia Huntington este o boala neurodegenerativa progresiva, autosomala, cu
transmitere dominanta, caracterizata fenotipic prin miscari involuntare (chorea), distonie, tulburari emotionale si scaderea capacitatii intelectuale. Gena HD, care a suferit o mutatie la bolnavi, a fost localizata pe cromozomul 4p16.3 in anul 1983 si a fost clonata 10 ani mai tarziu.
Deoarece polimorfismele la nivelul ADN sunt frecvente, Gusella si colaboratorii au adoptat o strategie sistematica de identificare a unui astfel de marker ADN asociat cu maladia Huntington. Ca subiect pentru acest studiu au fost alese doua familii, una americana si una venezueleana. Analiza incidentei bolii a aratat ca aproximativ jumatate din descendenti, de ambele sexe, contracteaza boala daca unul din parinti este afectat. Acest lucru a condus la concluzia ca boala este de natura autosomala, dominanta. In continuare s-a incercat localizarea genei raspunzatoare pe cromozomi.

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
294
Pentru aceasta, limfocite colectate de la ambele familii au fost permanentizate in cultura, pentru a se constitui surse permanente de ADN pentru analiza. In screeningul initial s-au folosit un numar de sonde pentru a analiza ADN-ul provenit de la familia americana. Una din aceste sonde, denumita G8, a aratat o puternica asociere cu gena maladiei Huntington. Clona G8 era un bacteriofag � recombinant continand o secventa unica de ADN uman de 17.6kb.
Pentru a stabili ca sonda G8 continea secvente din apropierea genei pentru maladia Huntington au fost necesare doua etape:
1. Mai intai s-a stabilit ca un numar de polimorfisme ale unor situsuri de restrictie poate fi detectat cu aceasta sonda.
2. In al doilea rand s-a stabilit ca un model caracteristic de situsuri de restrictie era transmis ereditar impreuna cu maladia
Aceasta analiza a fost destul de complexa deoarece s-a aratat ca asociat cu boala nu era un singur polimorfism al unui sit de restrictie, ci doua astfel de polimorfisme, ambele detectabile cu ajutorul sondei G8, si denumite situsul 1, respectiv 2. Acestea au fost detectate prin metoda Southern, utilizand HindIII ca enzima de restrictie. Cele patru combinatii de alele, sau haplotipuri, rezultate din aceste doua polimorfisme la nivelul situsurilor 1, respectiv 2, au fost desemnate ca fiind haplotipurile A, B, C si D. Gena pentru maladia Huntington segrega intotdeauna cu halpotipul A, in cazul familiei din America si cu haplotipul C, in cazul familiei din Venezuela.
Pentru a se localiza pe cromozom gena bolii Huntington, s-a constatat ca celule hibride om-soarece continand cromozomul 4 uman hibridizau cu sonda G8, in timp ce toti hibrizii lipsiti de cromozomul 4 nu hibridizau. Cum gena pentru maladia Huntington este linkata cu secventa G8, rezulta ca de asemenea ea este linkata cu cromozomul 4.
Mutatia responsabila de generarea maladiei consta in extinderea repetitiei unei trinucleotide, (CAG)n, care codifica pentru glutamina, in interiorul exonului 1 al genei, cu 17 codoni in aval de codonul de initiere ATG. La indivizii normali, numarul de repetitii al acestui codon este cuprins intre 6-35, in timp ce la indivizii afectati, numarul de repetitii este cuprins intre 40-121. Numarul de repetitii este invers proportional cu varsta la care apare boala.
Mucoviscidoza Mucoviscidoza este maladia recesiva autosomala cea mai frecventa in populatiile de
origine europeana, avand o incidenta de 1:2500 nou-nascuti. Aplicand legea Hardy-Weinberg, aceasta frecventa implica faptul ca 1:25 indivizi este heterozigot I populatie si ca frecventa genei mutate este de 2%. In forma sa cea mai obisnuita, maladia antreneaza o patologie a secretiilor exocrine, care afecteaza mai ales epiteliul cailor respiratorii. Nu exista un tratament specific si in majoritatea cazurilor maladia este extrem de invalidanta, evoluand inexorabil spre o insuficienta respiratorie mortala, care survine la diferite varste, de multe ori peste 20 de ani, datorita dezvoltarii tratamentelor paliative. Subiectii masculi

ORGANIZAREA GENOMULUI UMAN
295
ajunsi la maturitate sunt sterili datorita unei azoospermii generat de impermeabilitatea canalelor deferente.
Cautarea genei CF (Cystic Fibrosis) a fost demarata inca de la inceputurile strategiei geneticii inverse (1980). Aplicarea strategiei geneticii inverse la genele autosomale responsabile de o maladie recesiva se loveste de o dificultate intrinseca: imposibilitatea de a distinge heterozigotii, sanatosi dar purtatori ai genei anormale, de homozigotii normali.
Localizarea primara a genei CF pe cromozomul 7. Aceasta prima etapa a fost una laborioasa, deoarece absenta unui indice in favoarea
unui cromozom a impus o abordare pur aleatoare a cartografiei de excluziune. Au durat aproximativ 4 ani pentru a exclude in jur de 50% din genom. O problema foarte dezbatuta la inceput s-a referit la heterogenitatea genetica posibila a maladiei, data fiind heterogenitatea clinica a acesteia. Totusi, descoperirea unei prime legaturi semnificative intre un marker proteic polimorf, cu localizare cromozomala necunoscuta insa, paroxonaza (locusul PON), pleda in favoarea unui locus unic implicat in maladie. Acest locus a fost identificat gratie legaturii dintre boala si un situs polimorf explorat cu sonda anonima DOCRI 917, localizata pe bratul lung al cromozomului 7, descoperire facuta de Tsui in 1985. Acest prim marker era situat la aproximativ 15cM de locusul CF. Rapid acesta din urma a fost incadrat de doi markeri mai apropiati, proto-oncogena met si sonda anonima pJ3.11, situati respectiv la 0.4 cM si 0.3cM de locusul CF.
Cautarea genei CF Gena se afla deocamdata circumscrisa intr-un teritoriu care nu depasea 1 milion de
perechi de baze (Williamson, 1987). In subclonele derivate din aceasta regiune s-au pus in evidenta sonde (sondele XV-2c, CS7 si KM-19 la locusul anonim D7S23) care au permis detectarea unor RFLP in puternic dezechilibru de linkage cu gena CF mutanta (Estivil et al, 1987). Cu ajutorul acestor markeri s-au definit 4 haplotipuri, desemnate respectiv A, B, C, si D (vezi Tabelul 2)
Tabelul 2. Haplotipuri asociate cu gena CF
Alela Haplotip Xv-2c KM-19 (sau CS7)
A 1 1
B 1 2
C 2 1
D 2 2
O asociere preferentiala intre haplotipul B si gena mutanta CF a fost descoperita
pentru prima data in populatiile din Europa de Nord, pentru ca ulterior sa fie descoperite in populatiile de origine europeana din Statele Unite ale Americii si in Europa. Acest fenomen implica pe de o parte ca gena este foarte aproape de domeniul de cateva zeci de kilobaze

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
296
explorate de markerii respectivi, iar pe de alta parte, ca mutatia a avut loc o singura data, pe un haplotip comun, si anume pe haplotipul B care insoteste alela mutanta in peste 80% din cazuri
Descoperirea genei Gena CF a fost in cele din urma identificata in anul 1989 de catre echipele lui L. C.
Tsui (Toronto) si F. S. Collins (Ann Arbor). Acest succes a fost obtinut printr-o strategie de deplasare si salt pe un teritoriu de mai multe sute de kilobaze in aval de situsul KM-19, in cautarea unor secvente in acelasi timp conservate si exprimate, intr-o banca de cADN din celule de glande sudoripare. Una din aceste secvente, corespunzatoare unei insule HTF, a permis izolarea unei clone corespunzatoare inceputului genei. Aceasta prima secventa ( de doar 113 bp) a permis reconstituirea intregului puzzle de cADN. Validarea acestei gene s-a facut prin descoperirea unei deletii de 3 nucleotide care antrena pierderea unui codon Phe in pozitia 508 (mutatia numita �F508), regasita in 70% dintre cromozomii 7 purtatori ai anomaliei CF si in nici un caz la cromozomii nepurtatori. Acesta a fost argumentul decisiv pentru a se accepta ca gena pentru mucoviscidoza a fost desoperita. Un alt argument a fost furnizat de modelarea structurii proteinei putative codificate de aceasta gena, structura care este aceea a unei proteine transmembranare, apartinand superfamiliei transportorilor activi ATP-dependenti.
Datorita rolului presupus indirect al acestei proteine in transferul ionilor, in particular al ionilor de Cl-, ea a fost denumita CFTR (Cystic Fibrosis Transmembrane Regulator.
Gena este formata din 27 de exoni repartizati pe un teritoriu de 250Kb, ceea ce explica dificultatile intalnite in identificarea ei, deoarece aceasta reprezinta cea mai mare gena din genomul uman.
Patologia genei CFTR este dominata de un factor major in populatiile caucaziene: existenta unei alele comune, mutatia �F508, reprezentand 70% dintre mutatii, restul de 30% fiind constituite de o constelatie de alele rare si variate. Astfel, mai mult de 230 de mutatii non-�F508 sunt actualmente repertoriate.
Descifrarea genomului uman: Proiectul Genom Uman
Decifrarea secventei ADN care codifica genomul uman a fost anticipata de
sperantele pe care un astfel de proiect le genera in ceea ce priveste intelegerea evolutiei speciei umane, cauzele bolilor si relatia dintre mediu si organismul uman. Un proiect cu scopul de determina secventa completa de nucleotide a genomului uman a fost pentru prima oara adus in discutie in 1985. In anul 1990, Proiectul Genom Uman (HGP, Human Genome Project) a fost in mod oficial initiat in USA, sub conducerea Institutului National de Sanatate (NIH, National Institutes of Helth) si Departamentului Energiei al USA, proiect

ORGANIZAREA GENOMULUI UMAN
297
care urma sa se intinda pe durata a 15 ani, cu un buget de 3 miliarde USD, pentru descifrarea completa a genomului uman.
EST (Expressed Sequence Tags) Genomul organismelor superioare este reprezentat doar intr-o mica masura de ADN
codificator de proteine (1-3% in genomul uman). Totusi, ADN-ul complementar (ADNc), fiind revers-transcris din ARNm, reprezinta o sursa directa de secvente codificatoare ale genelor. Din acest motiv, secventierea ADNc a devenit o tehnica larg acceptata care completeaza intr-o masura importanta eforturile de secventiere ale genomului.
In anul 1991 a fost descrisa prima aplicatie a acestei metode de secventiere a ADNc, clone din creier uman fiind alese randomic si secventiate. Aceste secvente partiale reprezentau gene exprimate in tesutul nervos la un anumit moment de timp si ele au fost denumite EST-uri. Abordari similare pentru alte tesuturi, ca si pentru alte organisme au urmat rapid. EST-urile reprezinta cea mai semnificativa forma de informatie per baza secventiata, iar posibilitatea de a identifica rapid toate genele umane a condus la dezvoltarea unor baze de date comerciale care contineau secventele EST. Pentru a capta atentia publicului si a oferi o alternativa in acest domeniu, Merck Inc. a initiat in anul 1994 sponzorizarea efortului public de secventiere a EST-urilor la Univeritatea Washington, prin utilizarea de clone ADNc provenite de la consortiul I.M.A.G.E. (Lennon et al., 1996).
Secventele EST sunt extrem de utile pentru identificarea de noi gene, dar sunt de asemenea importante si din alte puncte de vedere. Secventele obtinute prin selectia randomica a clonelor ADNc furnizeaza o imagine statistica a nivelului si complexitatii expresiei genice in tesutul de provenienta. Influenta factorilor de mediu, precum si expresia genica specifica tesutului respectiv pot fi astfel studiate (Okubo et al., 1992). Secventele genice obtinute pot fi utilizate eficient pentru cartare fizica prin determinarea pozitiei lor pe cromozomi. Mai mult, secventele EST contribuie la intelegerea granitelor exoni-introni, ceea ce va permite prezicerea regiunilor transcrise ale genomului. Aceste fapte confirma necesitatea secventierii ADNc ca o completare a secventierii genomului. Seventele EST pot fi de asemenea utilizate pentru a evidentia variatii la nivelul unei singure baze in cadrul genelor. Detectarea acestor polimorfisme SNP (Single Nucleotide Polymorphism) poate fi un insrtument important de caracterizarea genelor pentru maladii umane. In sfarsit, clonarea pozitionala a genelor pentru diferite maladii va fi mult facilitata de combinarea intre secventierea EST-urilor si cartarea fizica de mare densitate, iar secventierea EST-urilor provenite de la diferite organisme va permite efectuarea de studii evolutioniste si clonarea unor gene interesante trecand peste barierele taxonomice.
Tehnologii de secventiere a ADN In ciuda resurselor implicate in eforturile de secventiere a genomului, tehnicile de
baza de secventiere a ADN dezvolate in urma cu 20 de ani sunt inca in uz. Cele mai importante tehnici de secventiere a ADN sunt metoda enzimatica de blocare a lantului

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
298
sintetizat (Sanger et al., 1977) si metoda de degradare chimica (Maxam si Gilbert, 1977). In comparatie cu metoda Maxam-Gilbert, metoda Sanger genereaza siruri de date mai usor de interpretat, ceea ce a facut ca aceasta tehnica sa devina cea mai larg utilizata.
Secventierea genomului In ciuda diferentei semnificative de talie intre diferitele genomuri, pentru a fi supus
secventierii orice genom trebuie sa fie subclonat in fragmente suficient de mici pentru a putea fi utilizate tehnicile de secventiere, iar secventierea trebuie sa se realizeze pe ambele catene la nivelul intregului genom, pentru a sigura acuratete procesului de secventiere.
Cartarea fizica. Consta in ordonarea clonelor continand inserturi mari de-a lungul cromozomilor. Clonele ordonate vor constitui baza pentru subclonarea ulterioara in vectori corespunzatori, care sa contina inserturi de dimensiuni accesibile tehnicilor de secventiere. Scopul ultim il constituie generarea unei harti fizice care sa acopere intregul genom.
Un numar de vectori diferiti este disponibil pentru construirea hartilor fizice, fiecare dintre acestia putand contine inserturi de diferite dimensiuni.
Astfel, YAC-urile (Yeast Artificial Chromosome) tolereaza inserturi cu dimensiunea intre 250kb-1Mb, si au fost utilizate pentru a produce contiguri (contig= set de clone care se suprapun) care acopereau majoritatea genomului uman (Cohen et al., 1993). Dezavantajul YAC-urilor este rata mare de himerism si rearanjari pe care le sufera. Pe de alta parte, YAC-urile pot contine regiuni dificil de clonat in constructe mici (de ex. cosmide).
Cosmidele sunt vectori plasmidiali care contin situsul cos de la fagul �, ceea ce permite impachetarea in vitro de particule fagice. Astfel, ADN-ul clonat poate fi cu usurinta introdus in celule de E.coli. Cosmidele pot contine inserturi de aproximativ 40kb si au fost frecvent utilizate ca legatura intre YAC-uri si vectori de secventiere.
O data cu dezvoltarea HGP a devenit tot mai imperioasa nevoia existentei unor vectori care sa mentina in stare stabila fragmente de ADN cu dimensiuni cuprinse intre cele continute de YAC-uri si cele continute de cosmide. Prin utilizarea factorului F de la E.coli, a fost construit un cromozom artificial bacterian numit BAC (Bacterial Artificial Chromosome), in anul 1992 (Shizuya et al.). acest vector poate contine inserturi de 15-300kb, uzual 100-200kb. Replicarea vectorilor ce contin factorul F se afla sub strict control in E.coli, ceea ce face ca BAC-urile sa fie mentinute intr-un numar mic per celula, in mod obisnuit una sau doua copii, ceea ce reduce riscul de recombinare. Eficienta mare in clonare prin electroporare, manipularea facila si stabilitatea au facut din acesti vectori instrumente mult mai potrivite pentru cartarea fizica decat Cosmidele.
Un al doilea sistem a fost dezvoltat prin modificarea unui vector bazat pe bacteriofagul P1. Acest sistem are proprietati similare cu BAC, si a fost denumit PAC (P1-derived Artificial Chromosome) (Ioannou et al., 1994). In ultimii ani atat BAC cat si PAC au fost larg utilizati in secventierea genomurilor mari.

ORGANIZAREA GENOMULUI UMAN
299
Pentru a crea harti fizice de mare rezolutie trebuiesc aplicate diferite tehnici de ordonare a clonelor. O imbunatatire in acest sens s-a produs odata cu utilizarea PCR pentru a amplifica regiuni genomice unice, de aproximativ 100-1000bp. Aceste secvente, denumite STS (Sequence Tagged Site) (Olson et al., 1989), apar o singura data in genom, si pot fi utilizate deci pentru a ordona YAC-urile sau BAC-urile prin metode de screening PCR.
Secventierea randomica Exista doua strategii majore de sceventiere a fragmentelor mari de ADN, randomica
si directa. Cele doua metode difera in ceea ce priveste procedura de clonare, dimensiunea inserturilor si strategia de secventiere.
In cazul metodei randomice, uzual denumita secventiere “shotgun”, este generata o librarie de subclone plasmidiale sau M13, cu dimensiunea inserturilor de 1-2kb. Un mare numar din aceste clone sunt apoi izolate randomic si secventiate utilizand primeri specifici pentru vectorul utilizat, generand astfel secvente raspandite randomic in fragmentul original (insert dintr-un cosmid sau BAC). Secventele sunt apoi combinate pentru a genera contiguri de pe ambele catene. Secventierea randomica este realizata pana cand se acopera 75-95% din fragmentul initial, dupa care se aplica strategii directe de secventiere pentru a umple golurile ramase.
Fractionarea inserturilor mari initiale in subclone se poate obtine prin cateva tehnici: digestia cu enzime de restrictie, tratamentul cu Dnaza I, ruperea ADN prin sonicare, presiune scazuta, HPLC, sau “nebulizare”.
Caracteristica tehnicii “shotgun” este numarul mare de clone care trebuiesc procesate pentru a se acoperi regiunea de interes. Numarul de reactii de secventiere este deci mai mare decat in cazul tehnicii directe; pe de lata parte insa, toate aceste reactii de secventiere se pot realiza cu aceeasi primeri, ceea ce permite standardizarea metodei. Secventierea “shotgun” necesita utilizarea unor algoritmi computerizati de calitate pentru a asambla datele in contiguri. Umplerea golurilor ramase in urma acestei metode se realizeaza prin metode directe.
Metode directe de secventiere. Se refera la tehnici in care reactia de secventiere este realizata la pozitii cunoscute in molecula de analizat. Strategia conduce la o minima redundanta si un numar semnificativ mai mic de reactii de secventiere, comparativ cu metoda “shotgun”.
Cea mai comuna metoda de secventiere directa este cea denumita “primer walking”. In acest caz, secventa obtinuta in urma unei reactii este utilizata pentru a construi urmatorul primer. Astfel, o plimbare de-a lungul secventei se poate realiza pe ambele catene. Asamblarea datelor este mai simpla decat in cazul metodelor “shotgun”, deoarece pozitia exacta a fiecarei reactii este cunoscuta. Dezavantajul metodei consta in continua nevoie de construire de primeri, sinteza care este scumpa si lenta.

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
300
Rezultatele HGP Incheierea HGP a condus la o serie de concluzii care acopera diferite aspecte ale
genomului uman. Numarul mic de gene umane. In urma secventierii si asamblarii a aproximativ 95% din portiunea eucromatinica a
genomului uman s-a estimat existenta a 26000-38000 gene, mult mai putine decat in estimarile anterioare (50000-140000).
Numarul mic de gene umane inseamna ca mecanismele care genereaza complexitatea inerenta in dezvoltarea umana si semnalele complicate implicate in mentinerea homeostaziei trebuiesc cautate in alta parte, nu in numarul mare de gene.
Variatia secventei ADN uman si distributia acesteia de-a lungul genomului. Desi s-au identificat peste 3 milioane de SNP-uri, aceasta nu inseamna ca sarcina
identificarii de alte polimorfisme SNP este incheiata. Acestea reprezinta doar o parte din numarul de SNP prezente in intregul populatiei umane. Totusi, s-a evidentiat o puternica heterogenitate in distributia acestor polimorfisme in genom.
Complexitatea genomului Simpla examinare a numarului de neuroni, tipuri genice, gene sau dimensiune a
genomului nu poate fi raspunzatoare de diferentele de complexitate constatate intre diferite organisme, tesuturi, etc. complexitatea este datorata mai degraba a interactiei dintre toti acesti factori.
Aplicatii ale rezultatelor HGP In cele mai recente lucrari, autorii pot doar specula asupra viitoarelor aplicatii ale
acestui proiect. Totusi pot fi deja citate cateva aplicatii directe, in principal din domeniul medical.
Gene implicate in fenomene patologice O aplicatie importanta a cercetarilor in domeniul genomului uman este descoperirea
de gene cu functie biochimica necunoscuta prin cartare pozitionala. Cel putin 30 de gene implicate in procese patologice au fost pozitionate in acest fel (Tabelul 3). In aditie, sunt multe cazuri in care secventa genica joaca un rol de suport, furnizand candidati pentru markeri microsatelitici asociati cu diferite maladii.

ORGANIZAREA GENOMULUI UMAN
301
Tabelul 3. Gene implicate in procese patologice Locus Maladie BRCA2 Breast cancer susceptibility
AIRE Autoimmune polyglandular syndrome type 1 (APS1 or APECED)
PEX1 Peroxisome biogenesis disorder
PDS Pendred syndrome
XLP X-linked lymphoproliferative disease
DFNA5 Nonsyndromic deafness
ATP2A2 Darier’s disease
SEDL X-linked spondyloepiphyseal dysplasia tarda
WISP3 Progressive pseudorheumatoid dysplasia
CCM1 Cerebral cavernous malformations
COL11A2/DFNA13 Nonsyndromic deafness
LGMD 2G Limb-girdle muscular dystrophy
EVC Ellis-Van Creveld syndrome
ACTN4 Familial focal segmental glomerulosclerosis
SCN1A Generalized epilepsy with febrile seizures plus type 2
AASS Familial hyperlysinaemia
NDRG1 Hereditary motor and sensory neuropathy –Lom
CNGB3 Total colour-blindness
MUL Mulibrey nanism
USH1C Usher type 1C
MYH9 May_Hegglin anomaly
PRKAR1A Carney’s complex
SCA10 Spinocerebellar ataxia type 10
OPA1 Optic atrophy
XLCSNB X-linked congenital stationary night blindness
FGF23 Hypophosphataemic rickets
GAN Giant axonal neuropathy
AAAS Triple-A syndrome
HSPG2 Schwartz-Jampel syndrome

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
302
Tinte pentru medicamente In ultimul secol industria farmaceutica a depins de numarul limitat de tinte cunoscute
pentru medicamente, in incercarea de a dezvolta noi produse. O lista recenta inventariaza 483 tinte pentru virtual toate medicamentele existente pe piata. Cunoasterea completa a secventei genelor, precum si a proteinelor codificate de acestea permite extinderea efortului de identificare de noi tinte. Desi doar o minoritate dintre genele umane pot fi tinte pentru medicamente, se estimeaza ca numarul acestora va depasi cateva mii si aceasta prezumtie a condus la o extindere masiva a cercetarii genomice in asociatie cu cercetari farmacologice.
Biologie fundamentala Desi exemplele discutate pana acum se refera doar la domeniul medical, exista
aplicatii similare in domeniul fiziologiei sau al biologiei celulare. Astfel, secventierea genomului uman a permis elucidarea unei probleme care ramanea nerezolvata de cateva decenii: baza moleculara a gustului amar. Oamenii si alte animale prezinta polimorfism in ceea ce priveste perceperea gustului amar. Recent cercetatorii au cartat aceasta trasatura atat in genomul uman cat si in cel murin, iar apoi au cautat regiuni relevante in secventa genomica uman pentru receptori cuplati cu proteina G. Aceste studii au condus rapid la descoperirea unei noi familii de astfel de proteine, la demonstrarea ca acestea sunt exprimate aproape exclusiv in papilele gustative si la confirmarea experimentala a faptului ca receptorii din celulele in cultura raspund la substante specifice amare.
Perspective Desi s-a realizat un progres semnificativ in secventierea genomului uman, raman
inca pasi care trebuiesc parcursi pentru a produce o secventa completa. Finalizarea secventei genomului uman Secventa genomului uman va servi ca baza pentru cercetarea biomedicala in anii
urmatori si, de aceea, este crucial ca golurile ramase sa fie umplute iar ambiguitatile sa fie eliminate cat mai repede. Se estimeaza ca pana in anul 2003 toti cromozomii vor fi complet secventiati.
Identificarea de regiuni reglatoare Genomul uman, regasit in toate celulele, in toate tesuturile, contine suficienta
informatie pentru diferentierea a sute de celule diferite si pentru a raspunde la o gama vasta de stimuli interni si externi. O mare parte din aceasta plasticitate rezulta din controlul fin la nivel transcriptional. Desi s-au invatat multe de spre motivele reglatoare cis ale diferitelor gene, semnalele genice ale majoritatii genelor raman in ca necunoscute. Analiza comparativa a genomului de la multiple vertebrate ofera cea mai buna speranta de identificare pe scara larga de astfel de motive. De asemenea, va fi interesant de studiat modificarile epigenetice, cum ar fi metilarea resturilor citozina la scara intregului genom, si determinarea consecintelor biologice ale metilarii. In acest scop, un proiect pilot al epigenomului uman a fost lansat.

ORGANIZAREA GENOMULUI UMAN
303
Secventierea altor genomuri mari Pentru realizarea cat mai completa si eficienta a analizei comparative in ceea ce
priveste genomul, determinarea secventei genomului a cat mai multe organisme este un deziderat important.
Completarea catalogului variabilitatii umane Secventierea genomului uman realizata pana in prezent a permis identificarea a peste
1.4 milioane SNP-uri, care sunt raspunzatoare intr-o proportie semnificativa de variabilitatea umana. Acest program trebuie extins pentru a se identifica virtual toate variantele genomice si pentru a se identifica haplotipurile ancestrale prezente in populatii.
De la secventa la functie HGP s-a concentrat pe identificare secventei genomului uman ca sursa de informatie
biologica. In completare, secventa genomului va furniza instrumente pentru o abordare functionala, intr-o maniera sistematica. Acest deziderat implica perfectionarea tehnicilor si bazelor de date pentru analiza globala a: expresiei ARN si a proteinelor; localizarii proteinelor; interactiei proteina-proteina; modalitatii chimice de inhibare a diferite cai metabolice.
Concluzii Proiectul Genom Uman va avea consecinte profunde pe termen lung asupra societatii
umane. Nu este vorba doar de aspectele medicale implicate, ci si de cele filosofice, etice. Rezultatele obtinute vor permite o intelegere mai adanca a ceea ce suntem, a directiei in care mergem. Pentru aceasta insa mai sunt de strabatut pasi, caci se poate spune ca pe masura ce invatam mai multe despre genomul uman, se ivesc mai multe aspecte de explorat.

L. O. POPA, I. MOANŢĂ, F. RAICU, D. CIMPONERIU, G. BORDEIANU, P. APOSTOL, L. DAN, O. POPA
304
Bibliografie 1. ***. A Physical Map of 30,000 Human Genes. Science 1998, 282: 744-746 2. ***. The sequence of the Human Genome. Science 2001, 291: 1304-1351 3. Bernardi G. The Human Genome: Organization and Evolutionary History.
Annu. Rev. Genetics. 1995. 29: 445-476 4. Brookes AJ. The essence of SNPs. Gene 1999, 234: 177-186 5. Brown TA. Genetics a molecular aproach. Chapman & Hall, 1992 6. Collins FS, Patrinos A, Jordan E, Chakravarti A, Gesteland R, Walters LR, and
the members of the DOE and NIH planning groups. New Goals for the U.S. Human Genome Project: 1998–2003. Science 1998, 282: 682-689
7. Gardiner K. Human genome organization. Current Opinion in Genetics & Development 1995, 5: 315-322
8. Gavrila L, Lungeanu A, Rogoz I. Citogenetica moleculara si evolutionista. Editura Stiintifica si Enciclopedica, Bucuresti, 1989
9. Gusella JF, MacDonald ME. Huntington’s disease: CAG genetics expands neurobiology. Current Opinion in Neurobiology 1995, 5: 656-662
10. International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome.Nature 2001, 409: 860-921
11. Kaplan J-C, Delpech M. Biologie moleculaire et medecine. Flammarion Medecine – Science, 1994
12. Kozian DH, Kirschbaum BJ. Comparative gene-expression analysis. Trends in Biotechnology 1999, 17: 73-77
13. Lewin B. Genes V. oxford University Press, 1994 14. Reddy PH, Williams M, Tagle DA. Recent advances in understanding the
pathogenesis of Huntington’s disease. Trends Neurosci. 1999, 22, 248-255 15. Sterky F, Lundeberg J. Sequence analysis of genes and genomes. Journal of
Biotechnilogy 2000, 76: 1-31 16. Zubay G. Genetics. The Benjamin / Cummings Company, Inc.,1987 17. Zweiger G, Scott RW. From expressed sequence tags to ‘epigenomics’: an
understanding of disease processes. Current Opinion in Biotechnology 1997, 8: 684-687