anatomical properties of plantago arenariabiologie.uvt.ro/annals/vol_14/vol_xiv_23-34.pdf ·...
TRANSCRIPT

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
23
ANATOMICAL PROPERTIES OF PLANTAGO
ARENARIA
Nicoleta IANOVICI, Adrian SINITEAN, Aurel FAUR
West University of Timisoara, Faculty of Chemistry-Biology-Geography, Department of Biology, Pestalozzi 16, Romania
Corresponding author e-mail: [email protected]
ABSTRACT Psammophytes are marked by a number of adaptations that enable them to exist in
the hard environmental conditions of the sand habitats. In this study, the
anatomical characteristics of Plantago arenaria were examined. Studies were
conducted to assess the diversity of anatomical adaptations of vegetative organs in
this taxa. Results are presented with original photographs. The analysis of leaf
anatomy in P. arenaria showed that the leaves contained a contained xeromorphic
traits. Arbuscular mycorrhizal symbiosis seems to be critical for their survival.
KEY WORDS: Plantago arenaria, xeromorphic traits, psammophytes
INTRODUCERE
Vegetaţia psamofită este supusă la o serie de factori ecologici care afectează în mod direct supravieţuirea şi distribuţia plantelor: nivelul scăzut al nutrienţilor şi apei, temperaturi ridicate, instabilitatea substratului (Rozema et al 1985; Hesp 1991; Maun 1997; Huang et al, 1997; Ciccarelli et al, 2009). Sistemul defensiv la psamofite poate fi activat continuu, atenuând daunele şi îmbunătăţind toleranţa lor la secetă (Luo et al, 2011).
Plantago arenaria caracterizează psamosolul (Filipov & Slonovschi, 2007). Aparţine familiei Plantaginaceae şi este cunoscută drept plantă medicinală în ultimele decenii (Hendawy, 2008; Ianovici et al, 2010; Nikolova et al, 2011). Pe teritoriul României, dintre cele 47 habitate care prezintă în structura şi compoziţia lor floristică specii ale genului Plantago (Doniţă et al, 2005; Andrei et al, 2009; Ianovici, 2009), 6 sunt caracterizate de existenţa speciei P. arenaria şi au o valoare conservativă mare şi foarte mare.
Plantago arenaria este geoelement eurasiatic continental, indiferentă faţă de reacţia solului, termofilă, xerofilă, oligotrofă, terofită (Ianovici et al, 2010), garnitura diploidă cu 12 cromozomi (Samantaray et al, 2010), descrisă morfologic în Flora României cu rădăcina pivotantă, tulpina ramificată (10 - 40 cm) erectă şi frunzele opuse, liniare (0.5–2 mm lăţime), întregi. Spicele capituliforme au 1–1.5 cm. Sepalele sunt inegale, cele două posterioare libere, ciliate iar cele anterioare concrescute, spatulate. Corola lungă de 4 mm are gâtul glabru şi lacinii lanceolate, albe brunii. Capsulă prezintă două seminţe lungi de 2–2.5 mm, în formă de luntriţă, pe o parte excavate. P. arenaria germinează foarte repede (24 - 48 ore) (Panayotova et al, 2008).
Lucrarea de faţă îşi propune evidenţierea la Plantago arenaria a principalelor caracteristici histoanatomice şi a unor parametri morfofiziologici.

IANOVICI et al: Anatomical properties of Plantago arenaria
24
MATERIALE SI METODE
Materialul biologic a fost recoltat în 2006 din preajma Staţiunii de Cercetare de la Agigea, pe coasta Mării Negre. Plantele au fost fixate în alcool etilic 70℅ şi s-au efectuat secţiuni transversale de mână şi preparate proaspete. Am folosit rectivul genevez şi albastru de toluidină pentru colorare. Pentru detaşarea epidermei am utilizat metoda jupuirii. S-au efectuat câte zece preparate de pe ambele suprafeţe ale frunzei. Epidermele au fost decolorate într-o soluţie de 5% hipoclorit de sodiu timp de 30-60 de minute. Densitatea stomatică (DS) şi a perilor epidermici (DP) reprezintă numărul de stomate per mm², respectiv numărul de peri per mm² (Ianovici, 2009). Pentru determinarea DS şi DP, dar şi a celorlalţi parametri foliari am folosit coeficientul micrometric pentru fiecare pereche obiectiv-ocular la microscopul Optika B500. Au fost realizate fotografii cu ajutorul aparatului Cannon PowerSchot A630. Parametri calculaţi (tab.1) au fost verificaţi cu softul QuickPHOTO MICRO 2.3. Prelucrarea statistică am realizat-o cu Microsoft Office Excel 2007 (Ianovici, 2009).
Probele radiculare au fost supuse colorării cu albastru de metilen şi acid acetic. Determinarea cantitativă a preparatelor radiculare a fost realizată potrivit metodei lui Trouvelot et al (1986) iar datele obţinute au fost introduse în programul Mycocalc pentru evaluare. În lucrarea de faţă sunt prezentaţi 2 parametri specifici (tab.2): abundenţa relativă a arbusculelor în sistemul radicular (A%) şi intensitatea colonizării micorizale în sistemul radicular (M%). Detalii privind protocolul de lucru au fost descrise în lucrări anterioare (Ianovici, 2009; Ianovici et al, 2009; Ianovici, 2010a; Ianovici, 2010b).
REZULTATE ŞI DISCUŢII
La nivelul frunzei, celulele epidermice văzute din faţă au contur regulat, sunt poligonale cu pereţii laterali drepţi dar îngroşaţi moniliform (în secţiune apicală). Aspectul este dat de prezenţa punctuaţiunilor simple. Stomatele sunt prezente în ambele epiderme, deci limbul este amfistomatic. Aparatele stomatice din epidermele foliare de la P.arenaria se încadrează în tipurile anisotricitic, anisocitic şi diacitic. Tipul predominant anisotricitic, cu două celule anexe mici şi una evident mai mare a fost sporadic pus în evidenţă la P. major ssp. intermedia f. minor (Ianovici, 2009). Densitatea stomatică în epiderma inferioară (223 stomate/mm2) este mai mare decât densitatea stomatică în epiderma superioară (164 stomate/mm2). Celulele stomatice acoperă un perimetru de 160 µm şi o arie de 2200µm2. Perii secretori cu glanda tetracelulară au celula bazală epidermică mică. Perii capitaţi sunt scurt pedicelaţi. Un păr glandular are în medie o lungime de 41,4µm. Perii tectori pluricelulari sunt de două tipuri: flagelaţi foarte lungi şi aculeiformi tetracelulari curbaţi. Pedicelul perilor tectori tetracelulari are o lungime de 392µm. Aceşti peri se remarcă printr-o cuticulă foarte groasă. Cuticula ce delimitează celulele trihomilor are o grosime de 9,2 µm, fiind asemănători din acest punct de vedere cu P. major ssp winteri şi P.maritima (Ianovici, 2009). Cei flagelaţi sunt vizibili cu ochiul liber, depăşind şi 2 mm. Celula bazală epidermică a acestora este mai mare (41.4 µm). Foarte rar pot fi evidenţiaţi şi peri tectori

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
25
unicelulari verucoşi. Densitatea perilor în epiderma superioară (22 peri/mm2) este foarte apropiată de densitatea perilor în epiderma inferioară (34 peri/mm2).
Fig. 1. Plantago arenaria – habitus, frunze şi peri tectori foliari, seminţe
Fig. 2. Plantago arenaria – epidermele foliare cu peri şi stomate anisotricitice, anisocitice şi diacitice, 400x şi 200x
Fig. 3. Celulele epidermice cu pereţi celulari moniliformi, 400x zoom 4 şi 900x

IANOVICI et al: Anatomical properties of Plantago arenaria
26
La Plantago arenaria limbul are structură omogenă, mezofilul nefiind diferenţiat. Celulele epidermice sunt mari, alungite uşor tangenţial şi cu pereţii îngroşaţi, mai ales cel tangenţial extern. (A=1156 µm2 şi P=129 µm). La extremităţile limbului, celulele epidermice se micşorează. Întreg clorenchimul este palisadic, doar puţine celule dispuse central fiind izodiametrice. O celulă palisadică are un perimetru de 83µm şi aria medie de 369 µm2.
În secţiune transversală, nervura mediană proemină la faţa abaxială, iar la faţa opusă se află un şanţ uşor adâncit. În acest şanţ sunt prezenţi cei mai mulţi peri secretori. Această nervură este complet deosebită de cele descrise la genul Plantago (Ianovici, 2009). Grosimea limbului este de 593 µm şi de 399 µm în dreptul nervurii. Unicul fascicul vascular este conectat la epiderme prin două extensii ale tecilor de legătură constituite din celule mari cu pereţi îngroşaţi. Fasciculul (P=1091µm; A=90162 µm2) este înconjurat de o endodermă a cărei celule sunt de aceeaşi mărime cu celulele tecii de legătură. Vasele lemnoase sunt dispuse radiar şi sunt separate prin parenchim lemnos biseriat. Liberul orientat spre faţa abaxială are o dispunere în semicerc. Fasciculul este prevăzut cu două calote colenchimatice, cea adaxială fiind mai slab reprezentată. Cuticula este mai groasă în dreptul nervurii (11.5µm). Cuticula în restul limbului prezintă o grosime de 9,2 µm. Cuticula la extremităţi poate fi chiar mai groasă, având în medie 13.8 µm. Părţile laterale ale frunzei sunt rotunjite şi niciodată nu se formează colenchim. Cel mai frecvent aparatele stomatice sunt foarte uşor adâncite în epidermă. Este vizibilă de asemenea şi camera substomatică. Creşterea suprafeţei interne libere este datorată creşterii dezvoltării ţesutului palisadic. Cu creşterea în volum a ţesutului palisadic, la care conductibilitatea în direcţia paralelă cu suprafaţa frunzei este mică, distanţa dintre nervuri devine mai mică. În frunză se pot observa numeroase fascicule cu organizare mai simplă, fără teci de susţinere şi mărginite de
Fig. 4. Peri secretori cu glanda tetracelulară şi celula bazală epidermică, 400x zoom 4
Fig. 5. Peri tectori flagelaţi şi aculeiformi curbaţi cu celula bazală epidermică, 400x zoom 4

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
27
endodermă (P= 309 µm; A= 7342 µm2). Celulele palisadice sunt aşezate radiar în jurul legăturilor centrului vascular şi astfel, în condiţii favorabile de aprovizionare cu apă, transportul apei de la legături la epidermă este amplificat. Apreciem în ansamblu această structură ca fiind xeromorfică cu celule palisadice. Iluminarea intensă şi împiedicarea cursului apei datorită deficienţei acesteia, rezultă în creşterea dezvoltării ţesutului palisadic (Shields, 1950). Dezvoltarea din punct de vedere structural a ţesutului palisadic este în mod clar corelată cu capacitatea de fotosinteză. Fotosinteza depinde de echilibrul existent dintre concentraţiile de lumină şi de dioxid de carbon, o grosime mai accentuată a frunzei şi dezvoltarea straturilor palisadice influenţează în mod direct acest echilibru şi optimizează fotosinteza la nivelul întregii frunze. Dezvoltarea crescută a ţesutului palisadic probabil că duce la o creştere a activităţii fotosintezei (Ianovici, 2009). Dezvoltarea accentuată a straturilor epidermice poate duce la creşterea capacitaţii de reflecţie a frunzei şi să protejeze celulele fotosintetice inferioare subiacente de la o iradiere excesivă. În timpul acestui proces temperatura frunzei se menţine la un nivel optim pentru a asigura o bună desfăşurare a proceselor fiziologice (Ianovici, 2009). În jurul legăturilor centrului vascular, celulele palisadice sunt aşezate radiar şi astfel, în condiţii favorabile de aprovizionare cu apă, transportul apei de la legături la epidermă este amplificat (Thoday, 1931). Părţile laterale ale frunzei sunt rotunjite şi nu se formează colenchim. Apa din frunze este condusă nu numai de nervuri dar şi de celulele mezofilului şi epidermei (Shull, 1934; Wylie, 1943). Transportul apei către epidermă este mult mai mare prin ţesutul palisadic decât prin parenchimul lacunar. Prezenţa spaţiilor intercelulare, în special la celulele palisadice, totuşi, limitează transportul apei în planul paralel cu suprafaţa frunzei (Wylie, 1943). Frunza conţine ţesuturi cu multe conexiuni laterale (epiderma şi parenchimul lacunar) şi altele cu puţine conexiuni laterale (parenchimul palisadic).
Fig. 6. Fascicul vascular din nervura principala a limbului foliar şi extensiile tecii de legătură, 400x
Fig. 7. Aparatele stomatice secţionate transversal, 400x zoom 4

IANOVICI et al: Anatomical properties of Plantago arenaria
28
a
b
Wylie a ajuns la concluzia că există o corelaţie între densitatea nervurilor şi volumul ţesuturilor din mezofil. Pe de o parte, cu creşterea în volum a ţesutului palisadic, la care conductibilitatea în direcţia paralelă cu suprafaţa frunzei este mică, distanţa dintre nervuri devine mai mică şi, pe cealaltă parte, cu creşterea în volum a ţesutului lacunar, la care conductibilitatea este eficientă în direcţia menţionată de mai sus, distanţa dintre nervuri devine mai mare (Fahn, 1967). Nervurile mari sunt înconjurate de mult parenchim care nu prezintă cloroplaste sau acestea sunt puţine. Nervurile mai mici, de asemenea, sunt de obicei înconjurate de un strat de celule parenchimatice împachetate strâns. Un asemenea strat este definit
Fig. 9. Plantago arenaria - sectiune prin frunza colorată cu albastru de toluidină, 40x şi 100x (a); coloraţie cu reactiv genevez, 100x şi 100x zoom4 (b)
Fig. 8. Cuticula şi ornamentaţiile cuticulare, 400x zoom 4 şi 900x

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
29
teacă de legătură (Fahn, 1964) sau endodermă, termen preluat din Toma & Rugină (1998). Prezenţa endodermei este semnalată şi de Metcalfe & Chalk, 1950. La P.
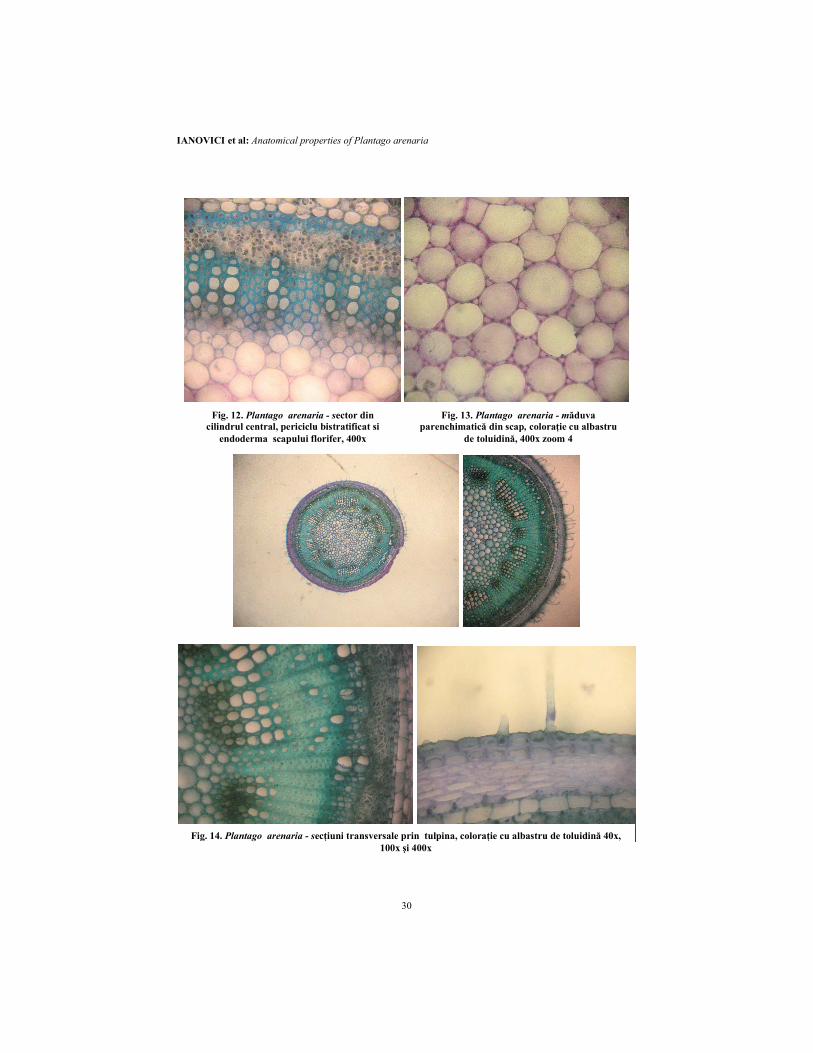
arenaria am întâlnit singura situaţie în care parenchimul tecii de legătură (endoderma fasciculului vascular unic corespunzător nervurii frunzei) se extinde către epidermele frunzei. Aceste plăci de celule parenchimatice care ajung la epidermă sunt definite extensiuni ale tecii de legătură. Există dovezi că extensiunile tecii de legătură au un rol de dirijare a apei către celulele epidermale. Cele din urmă sunt strâns conectate lateral şi funcţionează în dirijarea într-un plan paralel cu suprafaţa frunzei. Apa se poate mişca de asemenea de la endodermă şi extensiunile ei, prin mezofil, la celulele ce delimitează camerele substomatice (Wylie, 1943, 1951, 1952; Ianovici, 2009). Conturul secţiunii transversale prin scapul florifer (fig. 10-13) este circular. Unele secţiuni prezintă patru coaste abia schiţate. Epiderma prezintă celule izodiametrice, cu pereţii intern şi extern mai îngroşaţi decât ceilalţi. Cuticula este groasă. Din loc în loc se observă stomate. Camerele substomatice sunt foarte evidente. Perii prezenţi pe scapul florifer sunt aculeiformi cu picior tetracelular şi vârf curbat. Scoarţa este parenchimatic-asimilatoare, de tip meatic, din 6-7 straturi celulare. Stratul hipodermic este colenchimatizat. Stratul cortical cel mai intern reprezintă o endodermă de tip primar cu celulele bine individualizate, alungite tangenţial (P=81µm; A=469 µm2). Cilindrul central începe cu un periciclu sclerenchimatic de regulă bistratificat. Ţesuturile conducătoare au o dispunere aproape continuă, în puţine sectoare fiind întrerupte de raze medulare. Liberul formează un inel continuu, nefiind prezente fibre liberiene. Vasele lemnoase sunt dispuse în şiruri radiare uni şi biseriate. Între ele este dispus parenchim bi şi triseriat lignificat. Nu se observă liber intern. Sectoarele curbate mai accentuat prezintă celule parenchimate uşor lignificate la faţa internă a lemnului. Măduva este groasă, parenchimatic-celulozică, de tip meatic, cu celule foarte mari. Apreciem că pubescenţa crescută are legătură cu adaptarea la uscăciune pentru xerofite şi mezoxerofite, aşa cum este cazul la P. arenaria. Suprafeţele foarte pubescente pot să reflecte lumina soarelui şi să contribuie la descreşterea ratelor de transpiraţie. P. arenaria prezintă trei tipuri de peri tectori: aculeiformi curbaţi, flagelaţi multicelulari şi unicelulari verucoşi (Ianovici, 2009).
Fig. 10. Plantago arenaria - secţiune
transversală prin scapul florifer, 40x Fig. 11. Plantago arenaria - sector extern din scapul florifer
cu peri aculeiformi, 200x
IANOVICI et al: Anatomical properties of Plantago arenaria
30
Fig. 12. Plantago arenaria - sector din cilindrul central, periciclu bistratificat si
endoderma scapului florifer, 400x
Fig. 13. Plantago arenaria - măduva parenchimatică din scap, coloraţie cu albastru
de toluidină, 400x zoom 4
Fig. 14. Plantago arenaria - secţiuni transversale prin tulpina, coloraţie cu albastru de toluidină 40x,
100x şi 400x

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
31
Conturul secţiunii transversale prin tulpina aeriană (fig. 14 şi 15) de la P.
arenaria este circular. Epiderma prezintă celule izodiametrice, cu pereţii îngroşaţi. Cuticula este groasă. Perii tectori unicelulari verucoşi sunt foarte deşi. Scoarţa este parenchimatică, de tip meatic, din 6-7 straturi celulare. Stratul hipodermic este colenchimatizat. Celulele parenchimatice sunt puternic aplatizate (P=114 µm; A=683 µm2) cu peretii celulari mult ingrosati comparativ cu celulele corticale din tulpina avand structura primară. Stratul cortical cel mai intern reprezintă o endodermă de tip primar cu celulele bine individualizate, dreptunghiulare (P=133 µm; A=994 µm2). Cilindrul central începe cu un periciclu unistratificat, cu celule parenchimatice, de mici dimensiuni, aşezate în alternanţă cu celulele endodermice. Ţesuturile conducătoare au o dispunere inelară caracteristică structurii secundare. Liberul formează un inel continuu. Straturile din preajma periciclului sunt colenchimatizate. În rest sunt prezente doar tuburi ciuruite şi celule anexe. Vasele lemnoase sunt dispuse în şiruri radiare uni şi biseriate. Între ele este dispus parenchim lignificat. Structura secundară prezentă aici este caracterizată de prezenţa masivă a fibrelor lignificate, şi ele aşezate radiar. Cambiul funcţionează mai mult unifacial, prin urmare lemnul secundar este în cantitate mai mare. Măduva este groasă, de tip meatic, cu celule foarte mari ce manifestă tendinţă de îngroşare prin depunerea de lignină.
La P. arenaria rădăcina (fig. 16) are la maturitate o structură tipic secundară, edificată pe seama ambelor meristeme laterale. La nivelul scoarţei externe se diferenţiază o peridermă cu tendinţă de exfoliere. Suberul are celule mari, alungite tangenţial, cu pereţii suberificaţi. Felodermul rezultat din diviziunea felogenului se diferenţiază greu de scoarţa primară. Cilindrul central nu este delimitat cu precizie. Felodermul, scoarţa primară şi liberul secundar sunt de natură celulozică şi limita între ele este imprecisă. Ţesuturile conducătoare secundare, rezultate din activitatea cambiului, formează un inel extern subţire de liber şi un corp lemnos central gros. Liberul este format din tuburi ciuruite, celule anexe şi celule de parenchim, cu dispoziţie în şiruri radiare. Elementele liberului extern sunt vizibil colenchimatizate şi aplatizate. Lemnul secundar este format din numeroase vase dispersate în masa fundamentală de libriform. Vasele sunt solitare sau grupate în şiruri radiare discontinue.
Fig. 15. Plantago arenaria – perii unicelulari verucoşi la nivelul tulpinii aeriene, 200x şi 400x

IANOVICI et al: Anatomical properties of Plantago arenaria
32
Intensitatea colonizării micorizale în sistemul radicular (M=72.51% ) şi
abundenţa relativă a arbusculelor în sistemul radicular (A=61.32%) sunt mari. Aceste valori sunt peste cele raportate pentru alte specii ale genului (Ianovici, 2010b). Cercetările arată că VAM ameliorează şi cresc toleranţa la condiţiile de sol nefavorabile, pot spori productivitatea plantelor, protejează împotriva efectelor dăunătoare cauzate de stresul salin şi prin secetă (Sanchez-Diaz & Honrubia, 1994; Ruiz-Lozano et al., 1996; Brundrett et al., 1996; Ianovici, 2010a).
BIBLIOGRAFIE
• Andrei M., Ardelean A., Ianovici N. 2009. Habitatele din România cu Plantago sp. şi valoarea lor conservativă, NATURA – Biologie, Seria III, 51 (1): 41-67
• Brundrett M, 1991. Mycorrhizas in natural ecosystem. In: Macfayden, Begon, M and Fitter AH (eds). Advances in Ecological Research vol 21, Academi Press London. pp.171-173.
• Choong MF, Lucas PW, Ong JSY, Pereira B, Tan HTW, Turner IM. 1992. Leaf fracture toughness and sclerophylly: their correlations and ecological implications. New Phytologist 121: 597-610.
Tabel 1. Parametri histoanatomici la Plantago arenaria
Densitatea stomatică în epiderma superioară (DS) 164 stomate/mm2
Densitatea stomatică în epiderma inferioară (DS) 223 stomate/mm2 Densitatea perilor în epiderma superioară (DP) 22 peri/mm2 Densitatea perilor în epiderma inferioară (DP) 34 peri/mm2 Grosimea limbului între nervuri 399 µm Cuticula în dreptul nervurii 11.5-13.8 µm Cuticula între nervuri 9,2 µm Intensitatea colonizării micorizale în sistemul radicular (M) 72.51% Abundenţa relativă a arbusculelor în sistemul radicular (A) 61.32%)
Fig. 16. Plantago arenaria - secţiune transversală prin rădăcina cu structura secundară, coloraţie cu albastru de toluidină 40x si 100x

Annals of West University of Timişoara, ser. Biology, 2011, vol XIV, pp. 23-34
33
• Ciccarelli D., Forino L. M. C., Balestri M., Pagni A.M. 2009. Leaf anatomical adaptations of Calystegia soldanella, Euphorbia paralias and Otanthus maritimus to the ecological conditions of coastal sand dune systems, Caryologia, 62 (2): 142-151
• Cunningham S.A., Summerhayes B., Westoby M. 1999. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecological Monographs 69, 569-588.
• Eamus D., Myers B., Duff G., Williams R. 1999. A cost-benefit analysis of leaves of eight Australian savanna tree species of differing leaf life-span. Photosynthetica 36: 575-586.
• Fahn A., 1967, Plant Anatomy, Oxford, Pergaman Press Ltd. Headington Hill Hall Oxford • Filipov F., Slonovschi V. 2007. Plants indicating soil characteristics, Cercetări
Agronomice în Moldova, Anul XXXX , Vol. 4 (132) , 15-20 • Hendawy S.F. 2008. Comparative Study of Organic and Mineral Fertilization on
Plantago arenaria Plant, Journal of Applied Sciences Research, 4(5): 500-506 • Hesp P.A. 1991. Ecological processes and plant adaptations on coastal dunes. Journal
of Arid Environments, 21: 165-191 • Huang Zhenying, Wu Hong, Hu Zhenghai. 1997. The Structures of 30 Species of
Psammophytes and Their Adaptation to The Sandy Desert Environment In Xinjiang, Chinese Journal of Plant Ecology, 21 (6): 521-530
• Ianovici N. 2010b. Some preliminary data about vesicular – arbuscular mycorrhizas at different species of Plantago, Annals of West University of Timişoara, ser. Biology, 13: 129-134
• Ianovici N. 2009. Morphoanatomical researches on Plantago species from România, PhD Thesis, University of Bucharest, 306 p.
• Ianovici N. 2010a. Preliminary investigations on the arbuscular mycorrhizas in
Plantago lanceolata. In: Şesan T. (Eds.), Romanian approaches on mycorrhizas in the frame of European Researches, Ed. Universităţii din Bucureşti, p. 61-72
• Ianovici N., Ţărău G., Mujescu M., 2009. Quantification of Vesicular-Arbuscular Mycorrhizal Colonization on Plantago lanceolata in Zlatna, Valea Rizei and Timişoara,
Fungi & Mycotoxins, 3: 291-296 • Ianovici N., Ţărău G., Todosi A.L., Iriza E., Danciu A., Ţolea L., Tudosie D.,
Munteanu F., Bogdan D., Ciobănică V. 2010. Contributions to the characterization of Plantago species from Romania. Review, Annals of West University of Timişoara, ser. Biology, 13: 37-76
• King D.A. 1999. Juvenile foliage and the scaling of tree proportions, with emphasis on Eucalyptus. Ecology 80: 1944-1954.
• Lambers H, Poorter H. 1992. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Advances in Ecological Research 23: 187-261.
• Landsberg J. 1990. Dieback of rural eucalypts: Does insect herbivory relate to dietary quality of tree foliage? Australian Journal of Ecology 15: 73-87.
• Li C.Y., Berninger F., Koskela J., Sonninen E. 2000. Drought responses of Eucalyptus microtheca provenances depend on seasonality of rainfall in their place of origin. Australian Journal of Plant Physiology 27: 231-238.
• Maun M.A. 1997. Adaptations of plants to burial in coastal sand dunes. Canadian Journal of Botany, 76: 713-738
• Metcalfe, C.R. & Chalk, L., 1950. Anatomy of the Dicotyledons, vol.2, Oxford University Press, Oxford

IANOVICI et al: Anatomical properties of Plantago arenaria
34
• Nikolova M., Evstatieva L., Nguyen T.D. 2011. Screening of plant extracts for antioxidant properties, Botanica Serbica, 35 (1): 43-48
• Panayotova L.G., Ivanova T.A., Bogdanova Y.Y., Gussev C. V., Stanilova M.I., Bosseva Y. Zh., Stoeva T.D. 2008. In vitro cultivation of plant species from sandy dunes along the Bulgarian Black Sea Coast, PHYTOLOGIA BALCANICA 14 (1): 119 –123
• Patel A., Saravanan R. 2010. Screening of Plantago species for physiological parameters in relation to seed Yield, Electronic Journal of Plant Breeding, 1(6):1454-1460
• Pinkard E.A., Beadle C.L. 1998. Aboveground biomass partitioning and crown architecture of Eucalyptus nitens following green pruning. Canadian Journal of Forest Research 28: 1419-1428.
• Poorter H, Garnier E. 1999. Ecological signi®cance of inherent variation in relative growth rate. In: Pugnaire F, Valladares X,eds. Handbook of functional plant ecology. New York, USA: Marcel Dekker, 81-120.
• Rozema J., Bijwaard P., Prast G., Broekman R. 1985. Ecophysiological
adaptations of coastal halophytes from foredunes and salt marshes. Vegetatio, 62: 499-521
• Ruiz-Lozano JM, Azcon R, Gomez M, 1996. Alleviation of salt stress by arbuscular mycorrhizal Glomus species in Lactuca sativa plants. Physiol. Plant. 98:767-772.
• Ryser P. 1996. The importance of tissue density for growth and life span of leaves and roots: a comparison of five ecologically contrasting grasses. Functional Ecology 10: 717-723.
• Samantaray S., Dhagat U.M., Maiti S. 2010. Evaluation of genetic relationships in Plantago species using Random Amplified Polymorphic DNA (RAPD) markers, Plant Biotechnology 27, 297–303
• Sanchez-Diaz M, Honrubia M. 1994. Water relations and alleviation of drought stress in mycorrhizal plants. In Impact of Arbuscular Mycorrhizas on Sustainable Agriculture and Natural Ecosystems (S.Gianninazi and H. Schuepp (eds). Birkhauser Verlag, Basel, Switzerland. pp167-178.
• Shull C. A. 1934 Lateral water transfer in leaves of Ginkgo biloba. Plant Physiology 9: 387-389 • Steinbauer M.J. 2000. Specific leaf weight as an indicator of juvenile leaf toughness
in Tasmanian bluegum (Eucalyptus globulus ssp. globulus): implications for insect defoliation, Australian Forestry, 64 (1), 32-37
• Thoday, D. 1931. The significance of reduction in the size of leaves. Jour. Ecol. 19: 297. 40. • Toma Const., Rugină Rodica, 1998, Anatomia plantelor medicinale. Atlas. Ed. Academiei
Române, Bucureşti • Wylie R. B.. 1943. The role of the epidermis in foliar organization and its relations to the minor
venation. American Journal of Botany 30: 273-280 • Wylie R. B.. 1952. The bundle sheath extension in leaves of dicotyledons. American Journal of
Botany 39: 645-651 • Wylie R. B.1951. Principles of foliar organization shown by sun–shade leaves from ten species
of deciduous dicotyledonous trees. American Journal of Botany 38: 355-361