ssttuuddiiaa - ibn.idsi.md · emoţional. de aceea, modul în care părinţii îngrijesc copilul...
TRANSCRIPT

UNIVERSITATEA DE STAT DIN MOLDOVA
ISSN 1814-3237
ISSN online 1857-498X
Revistă Ştiinţifică Categoria C
SS TT UU DD II AA UUNNIIVVEERRSSIITTAATTIISS MMOOLLDDAAVVIIAAEE
SE
RIA
Ştiinţe reale
şi
ale naturii
• Biologie
• Chimie
Fondată în anul 2007
Chişinău Nr.6(66)
CEP USM 2013

Redactor-şef
Mihail REVENCO, profesor universitar, doctor habilitat, membru corespondent al AŞM (USM)
Redactori-şefi adjuncţi
Aurelia CRIVOI, profesor universitar, doctor habilitat (USM)
Viorica GLADCHI, conferenţiar universitar, doctor (USM)
Mihai LEŞANU, conferenţiar universitar, doctor (USM)
Consiliul academic Gheorghe DUCA, profesor universitar, doctor habilitat, academician (AŞM)
Aurelian GULEA, profesor universitar, doctor habilitat, academician (USM)
Maria GONŢA, profesor universitar, doctor habilitat (USM)
Vasile GUŢANU, profesor universitar, doctor habilitat (USM)
Petru CHETRUŞ, profesor universitar, doctor (USM)
Valentin BOBEICA, conf. universitar, doctor habilitat (USM)
Galina DRAGALINA, conf. universitar, doctor (USM)
Valerii LUNIN, profesor universitar, doctor, academician (Universitatea „M.V. Lomonosov”
din Moscova, Federaţia Rusă)
Rodolphe CLERAC, profesor universitar, doctor (Universitatea Bordeaux, Franţa)
Vucadin LEOVAC, profesor universitar, doctor (Universitatea Novi Sad, Serbia)
Vladimir ARION, profesor universitar, doctor habilitat (Universitatea din Viena, Austria)
Alexandru CECAL, profesor universitar, doctor (Universitatea „Al.I. Cuza” din Iaşi, România)
Constantin TOMA, profesor universitar, academician (Universitatea „Al.I. Cuza” din Iaşi, România)
Alexandru UNGUREANU, profesor universitar, membru corespondent al Academiei Române
(Universitatea „Al.I. Cuza” din Iaşi, România)
Vasile ŞALARU, profesor universitar, doctor habilitat, membru correspondent al AŞM (USM)
Victor ŞALARU, profesor universitar, doctor habilitat (USM)
Mihai COŞCODAN, conf. universitar, doctor (USM)
Ana BÂRSAN, conf. universitar, doctor (USM)
Ion TODERAŞ, profesor universitar, doctor habilitat, academician (AŞM)
Valeriu RUDIC, profesor universitar, doctor habilitat, academician (AŞM)
Leonid VOLOŞCIUC, profesor universitar, doctor habilitat (AŞM)
Coordonatori
Maria ŞLEAHTIŢCHI, conferenţiar universitar, doctor
Leonid GORCEAC, conferenţiar universitar, doctor
Raisa CREŢU
Lilia CEBAN
Redactori literari
Ariadna STRUNGARU (limba română)
Valentina MLADINA (limba rusă)
Dumitru MELENCIUC, conferenţiar universitar, doctor (limba engleză)
Asistenţă computerizată Ludmila REŞETNIC Tatiana VASILICĂ
Articolele incluse în prezentul volum au fost recomandate de catedre, consiliile profesorale
ale facultăţilor, consiliile ştiinţifice ale instituţiilor în cadrul cărora activează autorii,
recenzate de specialişti în domeniu şi aprobate spre publicare de către Senatul USM
© Universitatea de Stat din Moldova, 2013

190
CUPRINS SUMARRY
Biologie
Lidia COJOCARI, Aurelia CRIVOI
PERCEPŢIA INDIVIDUALĂ A TIMPULUI ÎN
RAPORT CU NIVELUL FUNCŢIONAL AL
EMISFERELOR CEREBRALE
3
TIME PERCEPTION IN RELATION TO
INDIVIDUAL FUNCTIONAL LEVEL
OF THE CEREBRAL HEMISPHERES
Inga DELEU
APRECIEREA NIVELULUI ELEMENTELOR
MINERALE DE CALCIU, MAGNEZIU ŞI FIER
LA SPORTIVII–ÎNOTĂTORI DE PERFORMANŢĂ
PÂNĂ LA ŞI DUPĂ EFORT FIZIC EFECTUAT PE
VELOERGOMETRU
9
ESTIMATION OF THE MINERAL ELEMENTS OF
CALCIUM, MAGNESIUM
AND IRON AT ELITE SWIMMERS BEFORE AND
AFTER PHYSICAL EFFORT ON THE BICYCLE
Анна ЛЕОРДА, Светлана ГАРАЕВА,
Влада ФУРДУЙ, Галина РЕДКОЗУБОВА,
Галина ПОСТОЛАТИ, Нина КОВАРСКАЯ
ОСОБЕННОСТИ АЗОТИСТОГО ОБМЕНА У
ЛИЦ С ПСИХИЧЕСКОЙ ДИЗОНТОГЕНИЕЙ
17
FEATURES OF NITROGEN METABOLISM IN
INDIVIDUALS WITH MENTAL DIZONTOGENY
Valentina BULIMAGA, Liliana ZOSIM,
Maria PISOV, Olga OLAN
INFLUENŢA CITRATULUI DE FIER ŞI A
INTENSITĂŢII LUMINII ASUPRA PRODUCTI-
VITĂŢII SPIRULINEI ŞI CONŢINUTULUI DE
CAROTENOIZI ŞI FIER ÎN BIOMASĂ
24
THE INFLUENCE OF IRON CITRATE AND
LIGHT REGIM ON SPIRULINA PRODUCTIVITY
AND CAROTENOIDS AND IRON CONTENT IN
BIOMASS
Natalia DONŢU
CULTIVAREA ALGELOR CHLORELLA
VULGARIS BEIJER, SYNECHOCYSTIS SALINA
WISL., PHORMIDIUM FOVEOLARUM (MONT.)
GOM. ŞI TRIBONEMA VIRIDE PASCH. PE MEDII
CU ADAOS DE APE REZIDUALE ORĂŞENEŞTI
28
CULTIVATION OF THE ALGAE CHLORELLA
VULGARIS BEIJER, SYNECHOCYSTIS SALINA WISL.,
PHORMIDIUM FOVEOLARUM (MONT.) GOM.
AND TRIBONEMA VIRIDE PASCH IN MEDIA
WITH ADDED MUNICIPAL WASTEWATERS
Vasile ŞALARU, Valentina BULIMAGA, Victor
ŞALARU, Alina TROFIM, Liliana ZOSIM,
Maria PISOV
ROLUL UNOR ALGE CIANOFITE
AZOTFIXATOARE ÎN REZOLVAREA
PROBLEMEI ALIMENTARE
33
THE ROLE OF NITROGEN FIXING
CYANOPHYTE ALGAE IN THE SOLVING
OF NUTRITION PROBLEM
Liliana ZOSIM, Cezara BIVOL, Ludmila BATÎR,
Daniela ELENCIUC, Svetlana DJUR, Olga OLAN
INFLUENŢA UNOR COMPUŞI COORDINATIVI
AI Cu(II) ASUPRA PRODUCTIVITĂŢII ŞI
CONŢINUTULUI DE PIGMENŢI FOTOSINTETICI
LA MICROALGA DUNALIELLA SALINA CNM-AV-02
42
THE INFLUENCE OF SOME COORDINATION
COMPOUNDS OF Cu (II) ON PRODUCTIVITY
AND PHOTOSYNTHETIC PIGMENT CONTENT
IN MICROALGA DUNALIELLA SALINA CNM-AV-02

191
Natalia CHISELIŢA, Agafia USATÎI,
Elena MOLODOI, Nadejda EFREMOVA,
Ludmila FULGA, Tamara BORISOVA
OPTIMIZAREA MATEMATICĂ A MEDIULUI
DE CULTURĂ PENTRU PRODUCEREA
ß-GLUCANILOR LA TULPINA
SACCHAROMYCES CEREVISIAE CNMN-Y-20
49
THE MATHEMATICAL OPTIMIZATION OF
CULTURE MEDIUM FOR
Β- GLUCANES PRODUCTION BY
SACCHAROMYCES CEREVISIAE CNMN-Y-20
Aliona GLIJIN, Maria JOIŢA-PĂCUREANU,
Adriana ACCIU, Maria GORCEAG, Maria DUCA
EFFECT OF BIOSTIMULATOR
“FERTILEADER GOLD” ON DEVELOPMENT
OF SUNFLOWER PLANTS
54
EFECTUL BIOSTIMULATORULUI
„FERTILEADER GOLD” ASUPRA DEZVOLTĂRII
PLANTELOR DE FLOAREA-SOARELUI
Дмитрий КУРШУНЖИ, Анатолий ГАНЯ
ИЗМЕНЧИВОСТЬ И НАСЛЕДОВАНИЕ
НЕКОТОРЫХ ПРИЗНАКОВ В F3 И F4- ПОКО-
ЛЕНИЯХ НУТА (CICER ARIETINUM L.)
60
VARIABILITY AND INHERITANCE OF SOME
TRAITS IN GENERATIONS F3 AND F4 OF
CHICKPEA (CICER ARIETINUM L.)
Rodica MARTEA, Irina DOROŞ, Alexandru MANOLE
VARIABILITATEA GENETICĂ INTRASPECIFICĂ
LA SALVIA SCLAREA L.
66
INTRASPECIFIC GENETIC VARIABILITY OF
SALVIA SCLAREA L.
Victoria NECHIFOR
CONTROLUL GENETIC AL MEIOZEI LA PLANTE
72
GENETIC CONTROL OF MEIOSIS IN PLANTS
Elena SAŞCO
EVALUAREA REZISTENŢEI UNOR HIBRIZI
AVANSAŢI DE TRITICUM AESTIVUM L. LA
AGENŢII PUTREGAIULUI DE RĂDĂCINĂ
ÎN CONDIŢII DE TEREN ŞI DE LABORATOR
80
EVALUATION OF RESISTANCE ADVANCEDS
HYBRIDS TRITICUM AESTIVUM TO THE
AGENTS ROOT ROT IN THE FIELD AND
LABORATORY CONDITIONS
Аркадий НИКОЛАЕВ, Светлана НИКОЛАЕВА,
Виктория ШУБИНА
ВЛИЯНИЕ БАКТЕРИАЛЬНЫХ МЕТАБОЛИТОВ
НА ХАРАКТЕР ПРОРАСТАНИЯ КОНИДИЙ
ГРИБА АЛЬТЕРНАРИЯ
86
EFFECT OF BACTERIAL METABOLITES ON
THE CHARACTER OF GERMINATION CONIDIA
OF FUNGUS ALTERNARIA
Аркадий НИКОЛАЕВ, Светлана НИКОЛАЕВА,
Виктория ШУБИНА
ДЕЙСТВИЕ ЭКСТРАКТОВ СЕМЯН
ВИНОГРАДА НА ФИТОПАТОГЕНЫ
В УСЛОВИЯХ IN VITRO
91
EFFECT OF EXSTRACTS OF GRAPE SEEDS ON
PHYTOPATHOGENS IN VITRO
Aurelia STÎNGACI
PROCEDEE BIOTEHNOLOGICE DE PRODUCERE
A PREPARATELOR VIRALE PENTRU
COMBATEREA DEFOLIATORULUI OMIDA
PĂROASĂ A DUDULUI (Hyphantria cunea Drury)
98
BIOTEHNOLOGICAL PRODUCTION PROCESSES
VIRAL PPREPARATION FOR CONTROL
OF THE FALL WEBWORM MOTH
(Hyphantria cunea Drury)
AIDAROSS NASSER NASR NAJI, Diana CRISTMAN
ЭФФЕКТИВНОСТЬ ФЕРОМОНА ФРУКТОВОЙ
ПОЛОСАТОЙ МОЛИ (Anarsia lineatella Z.)
105
EFFICIENCY OF PHEROMONES FRUIT MOTH
STIPED (Anarsia lineatella Z.)
AIDAROSS NASSER NASR NAJI
МИНИРУЮЩИЕ МОЛИ В ЯБЛОНЕВЫХ
САДАХ МОЛДОВЫ 109
MINED MOTH IN APPLE ORCHARDS OF
MOLDOVA

192
Виктория ШУБИНА
ПОЛУЧЕНИЕ ИНОКУЛЮМА ALTERNARIA
КУЛЬТИВИРОВАНИЕМ ПАТОГЕНА НА
БУМАЖНЫХ ФИЛЬТРАХ
114
PRODUCTION OF INOCULUM OF ALTERNARIA
WAS ACHIEVED BY CULTURING THE
PATHOGEN ON PAPER FILTERS
Андрей БАБИЦКИЙ
ИЗОЗИМЫ ЭСТЕРАЗЫ ИНБРЕДНЫХ ЛИНИЙ
И ГИБРИДОВ КУКУРУЗЫ
119
ESTERASE ISOZYMES OF MAIZE INBRED
LINES AND HYBRIDS
Андрей БАБИЦКИЙ
ОТ ЧЕГО ЗАВИСИТ СОДЕРЖАНИЕ ФОСФОРА
В СЕМЕНАХ ПШЕНИЦЫ
127
WHAT DETERMINES THE PHOSPHORUS
CONTENT IN WHEAT SEEDS
Vitalie SOCHIRCĂ, Corneliu IAŢU,
Alexandru-Iulian CIOBANU
CONSIDERAŢII CONCEPTUALE ŞI METODE
GEOGRAFICE DE ANALIZĂ A
FENOMENULUI TURISTIC
134
CONCEPTUAL CONSIDERATIONS AND
GEOGRAPHIC ANALYSIS METHODS
OF THE TOURISM PHENOMENON
Valeriu BRAŞOVEANU
ASPECTE PRIVIND REPARTIŢIA DEPUNERILOR
DE IONI MINERALI ÎN ECOSISTEMELE
FORESTIERE DIN REPUBLICA MOLDOVA ŞI
DIN EUROPA
141
ASPECTS OF REPARTITION THE
ION MINERAL DEPOSITION IN
THE FOREST ECOSYSTEMS IN
MOLDOVA AND EUROPE
Iulian MAMAI
PARTICULARITĂŢI ŞI TENDINŢE ALE UNOR
VALORI CLIMATICE DIN CADRUL
REZERVAŢIEI ŞTIINŢIFICE ,,PĂDUREA
DOMNEASCĂ”
148
PECULIARITIES AND TRENDS OF CLIMATE
VALUES OF THE NATURAL RESERVE
“PĂDUREA DOMNEASCĂ”
Mihai MÂRZA, Andrei NEGRU , Iulian MAMAI
FLORA SINANTROPĂ NECULTIVATĂ A
REPUBLICII MOLDOVA
154
FALLOW SYNANTHROPE FLORA OF REPUBLIC
MOLDOVA
Р.Ш. ИЗБАСАРОВА, А.Ш. ИЗБАСАРОВА
МОДЕРНИЗАЦИЯ БИОЛОГИЧЕСКОГО
ОБРАЗОВАНИЯ В 12-летней
ШКОЛЕ КАЗАХСТАНА
169
RENEWAL OF THE CONTENT OF
BIOLOGICAL EDUCATION IN THE
KAZAKHSTAN 12-YEAR SCHOOL
Chimie
Alexei MAFTULEAC, Nina ŢÎMBALIUC
PROCEDEELE DE AGITARE ŞI CENTRIFUGARE –
POSIBILE SURSE DE ERORI METODOLOGICE
ÎN STUDIUL ADSORBŢIEI
SUBSTANŢELOR DIN SOLUŢII
172
SHAKING AND CENTRIFUGATION PROCESSES
– POSSIBLE SOURCES OF METHODOLOGICAL
ERRORS IN THE STUDY OF ADSORPTION
FROM SOLUTIONS
Vasile RUSU, Alexei MAFTULEAC, Oleg PETUHOV
INTERCALAREA BENTONITULUI LĂRGUŢA
(R. MOLDOVA) CU OLIGOMERI DE ALUMINIU
176
ALUMINUM-PILLARING OF BENTONITE
LARGUTZA (R. MOLDOVA)
Eduard COROPCEANU
PROPRIETĂŢI UTILE ALE UNOR COMPUŞI
COORDINATIVI ÎN BAZA LIGANZILOR
DIOXIMICI
183
SOME USEFUL PROPERTIES OF
COORDINATION COMPOUNDS BASED ON
DIOXIME LIGANDS

193
Adresa redacţiei:
str. A.Mateevici, 60
MD2009, Chişinău, Republica Moldova
Tel. (37322) 577414; 577442; FAX (37322) 577440
e-mail: [email protected]
http://usm.md/studiau/
Formatul 60 84 1/8.
Coli de tipar 24,0. Coli editoriale 27,6. Comanda 479. Tirajul 100 ex.
Centrul Editorial-Poligrafic al USM
str. Al.Mateevici, 60. Chişinău, MD 2009

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
3
PERCEPŢIA INDIVIDUALĂ A TIMPULUI ÎN RAPORT CU NIVELUL FUNCŢIONAL
AL EMISFERELOR CEREBRALE
Lidia COJOCARI*, Aurelia CRIVOI
*Universitatea de Stat din Tiraspol
Universitatea de Stat din Moldova
A fost determinată dominanţa emisferică stângă la 48,19% din studenţi. Persoanele cu dominanţă emisferică dreaptă
şi ambidextrii posedă o capacitate mai bună de orientare în timp decât cele cu dominanţă stângă. La persoanele cu do-
minanţă emisferică stângă şi dreaptă mai frecventă este simpatotonia.
Cuvinte-cheie: dominanţă emisferică dreaptă şi stângă, ambidextri, perceperea individuală a timpului, indicele ve-
getativ Kerdo.
TIME PERCEPTION IN RELATION TO INDIVIDUAL FUNCTIONAL LEVEL
OF THE CEREBRAL HEMISPHERES
Left hemisphere dominance was determined at 48.19% of students. People with right hemispheric dominance and
ambidexters havea better ability of orientation in time than those with left dominance. The sympathicotonie is more
common for people with left and right hemisphere dominance
Keywords: left and right hemispheric dominance, ambidexters, individual perception of time, Kerdo vegetative index.
Introducere
Încă din perioada de dezvoltare embrionară există o asimetrie a sistemului nervos. Cele două emisfere ce-
rebrale au proprietăţi funcţionale diferite. Stânga controlează partea dreaptă a corpului, şi invers. Funcţiile
„superioare” ale creierului, cele legate de gândirea complexă şi abstractă, sunt direct influenţate de forţele
motivaţionale, emoţionale ale substraturilor subiacente ale aceleiaşi emisfere [15]. La adulţi emisferele cere-
brale funcţionează prin circuite neuronale care procesează în mod diferit informaţia [9].
Din punct de vedere anatomic, cele două emisfere sunt separate, legătura dintre ele realizându-se doar
prin fibrele de ţesut nervos din zona corpului calos şi prin comisura anterioară, care se dezvoltă în prima de-
cadă a vieţii individului [10].
La copil, creierul nu este încă specializat, maturizat şi, până în jurul vârstei de un an, copilul nu răspunde
la stimulările verbale cu întregul creier. Emisfera dreaptă domină dezvoltarea în perioada de început a vieţii,
până la declanşarea vorbirii.
A.Muntean (2006) menţionează că electroencefalogramele creierului la sugar, înregistrate la acţiunea sti-
mulilor de lumină discontinuă, vizează receptivitate doar în emisfera dreaptă a cortexului spaţio-vizual. Dacă
i se aplică unui adult acelaşi tip de stimulare, apare o dominanţă pe emisfera dreaptă, dar activitatea e reflec-
tată prin corpul calos şi în emisfera stângă. Aşadar, la sugar corpul calos este incapabil încă, din punct de ve-
dere funcţional, să asigure o bună comunicare între emisfere, în primele zile de viaţă.
Emisfera dreaptă este implicată mai mult în încercările sugarului de a se autocalma şi deţine o activitate
mai integră a sistemului somatosenzorial, reglează direct anumite procese ale organismului, e implicată în
percepţiile şi exprimările afective, este specializată în procesarea imaginilor perceptive, mediază datele auto-
biografice, memoria implicită, dar şi cea episodică şi procesează informaţia într-o manieră mai degrabă holistică.
Capacitatea empatică poate depinde, în mare măsură, de integrarea unor informaţii corporale, emoţionale şi
sociale, proces ce-şi are sediul predominant în emisfera dreaptă, precum şi de modul în care aceste informaţii
sunt interconectate cu emisfera stângă.
Emisfera stângă este implicată în acţiunile cu caracter explorator ale copilului, are o reprezentare şi o re-
glare a corpului mai scăzute, precum şi o capacitate diminuată de citire a expresiilor emoţionale nonverbale
ale celorlalţi. Emisfera stângă îndeplineşte predominant sarcini lingvistice, procesează informaţia într-un
mod silogistic (relaţia cauză-efect), liniar, logic.
Dominanţa emisferei drepte în primii ani ai copilăriei se manifestă atât printr-o creştere mai rapidă, cât şi
printr-o activitate mai intensă. Mai exact, în această perioadă apare o dezvoltare predominantă a ariei cortica-
le prefrontale a emisferei drepte, zonă ce reglează funcţionarea organismului şi comunicarea sincronizată

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
4
emoţional. De aceea, modul în care părinţii îngrijesc copilul în primii lui ani de viaţă, felul cum comunică cu
el influenţează formarea unor capacităţi care ţin de emisfera dreaptă: autoreglarea, relaţiile cu ceilalţi, sensul
autobiografic al sinelui, elementele de bază ale capacităţii empatice [15].
Toate aceste funcţii complexe se realizează fiziologic prin integrarea şi diferenţierea neuronală, într-un
proces de autoorganizare de o complexitate progresivă [2].
Deci, realizarea funcţiilor complexe ale creierului solicită integrarea unor aspecte care ţin de funcţionarea
emisferei drepte şi a celei stângi.
În urma observaţiilor clinice asupra bolnavilor cu leziuni cerebrale în focar, de tipul afecţiunilor neurochi-
rurgicale, s-a pus în evidenţă faptul (fie pe animalele de laborator sau în cadrul experimentelor clinice) că cele
două emisfere cerebrale nu sunt egale ca structură şi nu sunt echivalente funcţional [12].
S-a stabilit că leziunile corticocerebrale localizate într-o emisferă cerebrală şi cele corespunzătoare situate
în emisfera cerebrală contralaterală sunt diferite, atât ca formă de manifestare clinică, cât şi ca gravitate a
evoluţiei lor. Această observaţie pledează pentru ipoteza asimetriei funcţionale a celor două emisfere cere-
brale, dreaptă şi stângă [12]. Observaţiile clinice de ordin chirurgical au subliniat dominanţa emisferei stângi
asupra celei drepte, datorită integrării într-o mai mare măsură a unor funcţii neuropsihice în emisfera stângă
în raport cu cea dreaptă.
Asimеtria funcţională interemisferică apare deosebit de pregnant în situaţiile în care conexiunile interemi-
sferice sunt afectate lezional [8]. Separarea lezională a celor două emisfere cerebrale va avea consecinţe
funcţionale imediate. În cazul separării celor două emisfere prin secţionarea corpului calos se vor constata
următoarele: sarcinile verbale adresate emisferei cerebrale stângi şi sarcinile videoconstructive adresate emi-
sferei cerebrale drepte pot fi rezolvate corect; sarcinile verbale adresate emisferei cerebrale drepte şi sarcinile
videoconstructive adresate emisferei cerebrale stângi nu pot fi rezolvate corect.
Cele două emisfere au o specializare funcţională caracterizată prin următoarele [4,18]: emisfera cerebrală
stângă deţine funcţiile proprii limbajului, abstractizarea şi conceptualizarea; emisfera cerebrală dreaptă deţine
funcţiile nonverbale, în special recunoaşterea perceptivă şi construcţia spaţială.
Din acestea nu ar rezulta faptul că o emisferă se relevă superior faţă de cealaltă, ci, dimpotrivă, că fiecare
emisferă tratează informaţia într-un fel care-i este propriu: emisfera cerebrală stângă tratează informaţiile
care-i parvin într-un mod analitic şi secvenţial, iar emisfera dreaptă – după un model global sintetic.
Limbajul nu este proprietatea exclusivă a emisferei stângi. Emisfera cerebrală dreaptă nu apare ca privată
de funcţia limbajului, ci ca mai puţin specializată în limbaj prin modul său de tratare a informaţiei. Ea perce-
pe semnalele verbale, auditive sau vizuale, dar posibilităţile sale sunt foarte limitate [17].
Asimetria funcţională interemisferică trebuie considerată într-o manieră dinamică şi nu statică, ţinându-se
cont de condiţiile care o pot modifica: natura sarcinii de îndeplinit, tipul de atenţie pe care-l creează, experi-
enţa anterioară a subiectului, strategiile de realizare adoptate. În plus, diferit de procesele instrumental-sim-
bolice, fiecare emisferă cerebrală pare să controleze în mod diferit relaţiile emoţionale ale individului. H.Hecaen
(1963) a stabilit un model propriu al modurilor de funcţionare interemisferică, privind în mod special perfor-
manţele verbale. În acest sens, competiţia dintre cele două emisfere pare să presupună o cooperare interemis-
ferică, fiecare aducându-şi aportul caracteristicilor sale funcţionale, în raport cu natura sarcinilor care trebuie
îndeplinite. În sensul acesta, emisfera cerebrală dreaptă poate interveni în tratarea lor analitică, care este funcţia
emisferei cerebrale stângi [3].
C.Enăchescu (1996) caracterizează asimetria funcţională interemisferică, în cazul funcţiilor instrumental-
simbolice, prin două note specifice – relativitatea şi caracterul pendulant al acestora. Ele depind, în mare mă-
sură, de faptul că funcţiile instrumental-simbolice sunt reprezentate bilateral pe suprafaţa corticală. În condi-
ţii patologice, funcţia perturbată a unei arii corticocerebrale a uneia dintre emisfere poate fi preluată de aria
corespunzătoare din emisfera cerebrală contralaterală rămasă intactă, suplinind în felul acesta funcţia pierdu-
tă. În acelaşi timp, poate fi observat faptul că în condiţii patologice, cu leziuni ale ariei limbajului, această
funcţie poate fi suplinită, în unele privinţe, de zona corticocerebrală a emisferei drepte. În condiţiile lezării
ariei corticale a funcţiilor instrumental-simbolice din emisfera cerbrală stângă dominantă, funcţiile sale
instrumental-simbolice vor fi preluate de emisfera cerebrală dreaptă, care va prelua şi rolul dominant funcţio-
nal. Prin cunoscutul experiment „split-brain” devine mai explicită asimetria funcţională interemisferică.
Prin urmare, specializarea funcţională este următoarea: conştiinţa, funcţia verbală, ideaţia, conceptualiza-
rea, analiza timpului, analiza detaliilor, calculul aritmetic se realizează de emisfera stângă; funcţiile nonver-

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
5
bale, perceperea, reprezentarea şi orientarea spaţială, construcţia spaţială, imaginile globale, sinteza timpului,
geometria spaţiului se realizează de emisfera dreaptă.
Se consideră că encefalul în ultimii 100-150 mii de ani practic nu s-a schimbat. Însă, cercetările ştiinţifice
din ultimii ani atestă modificări esenţiale în asimetria funcţională a encefalului. Până în anii 70 ai sec. XX,
la copii numărul stângacilor era constant – 7-8% din populaţie [11], date mai recente atestă 5-10% din 800
de elevi ai claselor primare [16].
În anii 80 ai sec. XX savanţii au atras atenţia asupra creşterii numărului de copii stângaci şi al copiilor cu
,,stângăcie ascunsă”. Acest fenomen în SUA a fost observat încă cu 10 ani în urmă. În anii 90 ai sec. XX numărul
lor a crescut până la 15-17%. Apoi s-a declarat despre o categorie calitativ nouă de copii – ambidextri sau ,,bima-
nuali”, consideraţi, ca şi cei stângaci, ,,cu defect” sau ca variantă cu dereglări în dezvoltare. Zece ani mai târziu
atitudinea faţă de cei ,,bimanuali” s-a schimbat radical. La momentul actual este una din variantele normă [14].
Ambidextrul este o persoană cu o dezvoltare a emisferelor cerebrale la acelaşi nivel. Lipsa dominanţei de-
termină că ambidextrul poate să lucreze la fel de bine atât cu mâna stângă, cât şi cu dreapta. Ponderea ambi-
dextrilor nu depăşeşte 0,4% şi variază de la o specie la alta. Potrivit datelor recente ale lui V.Pugaci, astăzi
ponderea copiilor ambidextri este de 37,95%. Un număr mai mare de ambidextri se întâlneşte printre copiii
claselor primare şi ai clasei a V-a.
Oamenii de ştiinţă [14,16] susţin că atunci când ambele emisfere încep să proceseze informaţia paralel şi
concomitent, apar noi posibilităţi calitative în activitatea creierului. Însă, acest fenomen are şi particulari-
tăţi negative. În clasele primare ambidextrietatea este un obstacol serios în procesul intructiv-educativ. La
copiii ambidextri, mai ales de vârstă şcolară mică şi medie, au loc comutări spontane ale activităţii emisferei
drepte şi emisferei stângi ale creierului, din cauza imaturităţii legăturilor interemisferice. Din acest motiv, ei
demonstrează un comportament polar, un fel de personalitate multiplă. Elevii-ambidextri au nevoie de ceva
mai mult timp pentru însuşirea materialului nou, un număr mai mare de explicaţii. De asemenea, este im-
portant ca explicaţiile să se bazeze pe imagini, scheme şi desene.
Scopul cercetării noastre rezidă în evaluarea percepţiei timpului şi reacţiei sistemului nervos vegetativ în
dependenţă de nivelul funcţional al emisferelor cerebrale.
Material şi metode
În studiu au fost implicate 83 de studente – de la Universitatea de Stat din Tiraspol şi de la Universitatea
de Stat din Moldova. S-a determinat nivelul funcţional al emisferelor cerebrale, gradul asimetriei funcţionale
după metoda propusă de E.S. Juricov şi E.B. Zolotov (1990), conform căreia preferinţa pentru activităţile
care implică manifestarea funcţiilor psihice controlate în special de una din emisferele cerebrale indică pre-
dominanţă emisferică a individului.
Asimetria funcţională s-a calulat după formula:
AsF = ES(%) – ED(%) ,
unde: AsF – asimetria funcţională; ES – emisfera stângă; ED – emisfera dreaptă.
Evaluarea rezultatelor s-a efectuat în felul următor: AsF = 0 atestă funcţionarea simetrică a celor două
emisfere; AsF > 0 arată o dominanţa a emisferei stângi; AsF < 0 reprezintă dominanţa emisferei drepte.
Indicii de prevalenţă a membrelor s-a monitorizat prin probe repetate ale pozei lui Napoleon, deschiderea
flacoanelor, sărituri într-un picior etc.
Pentru determinarea nivelului funcţional al sistemului nervos vegetativ s-a determinat indicele Kerdo
după formula:
IK = 100 x (1 – TAD/P),
unde: TAD – tensiunea arterială diastolică (mmHg), P – frecvenţa contracţiilor cardiace (contracţii/ minut).
Evaluarea rezultatelor s-a făcut în felul următor: dacă IK > 0, atestă o predominare a sistemului nervos
vegetativ simpatic; dacă IK < 0 – o predominare parasimpatică, iar IK = 0 – un echilibru funcţional.
Pentru evaluarea perceperii timpului s-a determinat perceperea individuală a duratei minutului. Persoane-
le investigate trebuiau să aprecieze intervalul de timp – 60 secunde. Deviaţiile admise fiind de ±3 secunde.
Deoarece perceperea individuală a timpului se poate modifica sub influenţa diferiţilor factori, noi am deter-
minat minutul individual în decurs de 4 zile. Aprecierea subiectivă a minutului individual de la 57 secunde
până la 63 secunde corespunde unei perceperi adecvate a timpului, peste 63 secunde – unei perceperi lente,
mai puţin de 57 secunde – unei perceperi rapide [13].

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
6
Rezultate
Analizând rezultatele probei ce estimează nivelul funcţional al emisferelor cerebrale, am determinat la
48,19% din studenţi o dominanţă stângă. La ei ca indicator de preferinţă a membrelor s-a estimat dominanţa
mâinii drepte şi a piciorului drept legate de funcţiile emisferei cerebrale stângi. Gradul asimetriei funcţionale
la aceste persoane a variat între 6,45 şi 47,87%. Pentru aceşti studenţi activităţile în care pot obţine cele mai
bune rezultate sunt cele legate de prelucrarea logică a informaţiei – calcule, rezolvarea problemelor de mate-
matică, transmiterea verbală sau în scris a informaţiei sau cele care presupun capacitatea de a argumenta şi
susţine ideile şi propunerile proprii sau ale altora.
Predominarea emisferei drepte s-a estimat la 28,91% din studenţi, la care ca indicator de preferinţă a
membrelor s-a estimat dominanţa mâinii stângi şi a piciorului stâng legate de competenţa emisferei cerebrale
drepte, considerate dominantă din punct de vedere funcţional. Gradul asimetriei funcţionale la ei a variat între
valorile -2,47 şi -66,99%. Aceste persoane percep lumea în întregul ei, foarte rapid înţeleg esenţa evenimen-
telor pe cale intuitivă, fără o analiză logică consecventă a acestora, au o capacitate de imaginaţie şi proiecţie
videospaţială.
O simetrie funcţională a celor două emisfere cerebrale s-a estimat la 22,89% din studenţi, la ei gradul asi-
metriei funcţionale fiind 0. La aceşti studenţi s-a evidenţiat o relativă echivalenţă funcţională între membrele
drept şi stâng, fapt ce traduce o reprezentare a procesului de asimetrie funcţională interemisferică.
Cu problema timpului omul se confruntă zilnic, în fiecare minut. Timpul este o dimensiune în care se des-
făşoară existenţa umană. Nicio activitate nu se desfăşoară fără perceperea timpului. De aceea, este important
a cunoaşte ce este ,,timpul”, cum este perceput de către o persoană, de care factori depinde percepţia lui.
Acest lucru poate ajuta omul în organizarea activităţii, utilizarea raţională a timpului. Cercetările lui D.G. Elkin
arată că între percepţia timpului şi activitate este o relaţie directă: cu cât mai precisă este percepţia timpului,
cu atât mai prosperă este activitatea [6].
Timpul este forma de derulare a tuturor proceselor mecanice, organice şi psihice, condiţie a posibilităţii
de mişcare, schimbare şi dezvoltare; fie o deplasare spaţială, o schimbare calitativă, naşterea sau moartea,
toate au loc în timp.
Pentru orice individ sunt extrem de importante caracteristicile obiectelor din jur – forma lor, distanţa,
viteza de deplasare şi distanţa dintre ele. În legătură cu aceste cerinţe la animale s-a dezvoltat un aparat psi-
hic deosebit, a cărui funcţie constă în reconstrucţia spaţiului în interior, percepţia subiectivă a unei imagini
a lumii, care ar reproduce cel mai fidel caracteristicile spaţiale şi în timp ale obiectelor şi fenomenelor. În
această imagine trebuie să fie prezentate, într-o formă sau alta, şi alte calităţi ale obiectelor, iar imaginea
pentru subiect trebuie să fie veridică, astfel încât să nu existe nicio îndoială în ceea ce priveşte existenţa
obiectului, perceperea lui sau a unui fenomen.
Timpul există în mod obiectiv şi continuu: un minut se schimbă cu altul, o zi – cu alta, se scurg luni, ani,
secole. Dar, în pofida faptului că scurgerea timpului în toată lumea se supune aceloraşi legi, pentru fiecare
individ aceeaşi perioadă de timp trăită obiectiv poate părea mai scurtă sau mai lungă decât a fost de fapt.
Pentru persoane diferite aceeaşi perioadă de timp poate avea importanţă complet diferită. Pentru o persoană
săptămâna ,,durează”, pentru alta săptămâna ,,zboară” asemeni câtorva zile.
Percepţia este o reflectare subiectivă plus o imaginare conformă cu aceasta în conştiinţa omului a faptelor,
obiectelor şi fenomenelor din realitatea obiectivă care acţionează direct asupra organelor de simţ. În acest
context organele de simţ sunt categorisite ca analizatori. Percepţia este proprietatea psihicului de a reflecta
impresiile obiectelor, implicând gândirea, memoria, imaginaţia, formând imagini sintetice ale obiectelor re-
cepţionate; ea se fundamentează pe experienţa subiectivă, provoacă interese, aptitudini, stări afective. Per-
cepţia presupune prezenţa diferitelor senzaţii şi decurge împreună cu senzaţiile. Comparativ cu senzaţia,
percepţia constituie un nivel superior de prelucrare şi integrare a informaţiei despre lumea externă şi despre
propriul nostru ,,eu”. Superioritatea percepţiei constă în realizarea unei imagini sintetice, unitare, în care
obiectele şi fenomenele care acţionează direct asupra organelor analizatoare sunt reflectate ca totalităţi inte-
grale, în individualitatea lor specifică [1, 3].
Percepţia timpului este reflectarea duratei şi succesiunii fenomenelor sau evenimentelor. Intervalele tem-
porale sunt determinate de procesele ritmice ce au loc în organismul omului. Percepţia duratei timpului de-
pinde de conţinutul activităţii omului. Timpul, plin de ocupaţii interesante, semnificative, trece repede. Dacă,
însă, evenimentele nu sunt interesante, puţin importante, timpul se scurge încet [5].

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
7
Perceperea timpului depinde de un şir de factori: omul foloseşte trei sisteme de referinţă [5]:
sistemul fizico-cosmic, care oferă cunoaşterea reperelor naturale – zi, noapte, poziţie a aştrilor, anotim-
puri etc.;
sistemul biologic, ce depinde de succesiunea funcţiilor organismului, permiţând individului uman să
aprecieze ora mesei, timpul somnului, trezirii etc.;
sistemul sociocultural, constituit pe parcursul activităţii umane, care oferă repere cronologice – divi-
zarea timpului în secunde, minute, ore, luni, ani etc.
Pe baza experienţei ce o posedă, a cunoştinţelor omul în procesul percepţiei uneşte elementele separate
într-o imagine integrală.
La baza percepţiei stau reflexele condiţionate, conexiunile neuronale temporare, care se formează în cor-
texul cerebral ca răspuns la acţiunea exercitată asupra receptorilor de către obiectele sau fenomenele me-
diului înconjurător. Ultima acţionează ca un complex de stimuli. În nucleele centrilor corticali ai analizatori-
lor are loc o analiză şi sinteză complexă a acestor stimuli. La baza procesului complex de formare a percep-
ţiei imaginii sunt conexiunile dintre analizatori şi cele din cadrul analizatoruliui, care oferă cele mai bune
condiţii de evidenţiere a stimulilor şi de interacţiune a caracteristicilor unui obiect ca un întreg complex [1].
Studiind durata minutului individual, ce caracterizează capacitatetea de recepţionare subiectivă individua-
lă a timpului, am stabilit că persoanele cu dominanţă emisferică stângă (DS) supraapreciază secvenţa de timp
analizată (69,1±1,94) şi atestă o supraîncărcare informaţională şi o tensionare a mecanismelor de reglare [13],
fapt vizat şi de reactivitatea sistemului nervos vegetativ. Studenţii cu dominanţă dreaptă (DD) fie că apre-
ciază adecvat decurgerea timpului, fie că o subapreciază (57,3 ± 1,02), iar persoanele ambidextre (A) percep
adecvat derularea timpului (59,4 ± 2,03).
Fig.1. Perceperea subiectivă a timpului în dependenţă de dominanţa emisferică.
Astfel, datele obţinute atestă că persoanele cu dominanţă dreaptă şi ambidextrii posedă o capacitate mai
bună de orientare în timp decât persoanele cu dominanţă stângă. Datele obţinute nu contravin datelor din lite-
ratură [7], potrivit cărora subiecţii cu dominanţă dreaptă au o fidelitate mai mare în reproducerea etalonului
intervalului de timp pentru recunoaşterea stimulilor verbali.
Este cunoscut că persoanele cu percepere diferită a duratei minutului se caracterizează prin divers grad al
capacităţii de muncă, reacţii diverse ale sistemului cardiovascular, sistemului nervos vegetativ.
Sistemul nervos vegetativ nu poate fi controlat în mod conştient de individ. Deşi prezintă o anumită indi-
vidualitate, sistemul nervos vegetativ are relaţii strânse cu sistemul nervos al vieţii de relaţie, atât la nivel
central, cât şi la nivel periferic, ceea ce ne-a motivat să determinăm indicele vegetativ Kerdo în dependenţă
de nivelul funcţional al emisferelor cerebrale.
A fost stabilită o diferenţă după distribuirea indicelui vegetativ Kerdo la studenţii cu diferit tip de domi-
nanţă cerebrală (Fig.2).
La persoanele cu dominanţă stângă şi dreaptă mai frecvent se întâlneşte simpatotonia, iar la persoanele
ambidextre se întâlnesc diferite variante de reacţionare a sistemului nervos vegetativ.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
8
Fig.2. Distribuirea procentuală a studenţilor după indicele vegetativ.
Concluzii S-a determinat dominanţă emisferică stângă la 48,19% din studenţi, care supraapreciează secvenţa de
timp analizată şi o tensionare a mecanismelor de reglare. Persoanele cu dominanţă emisferică dreaptă şi am-bidextrii posedă o capacitate mai bună de orientare în timp decât cele cu dominanţă stângă. La persoanele cu domiinaţă emisferică stângă şi dreaptă mai frecventă este simpatotonia.
Bibliografie:
1. ENĂCHESCU, C. Neoropsihologie. Bucureşti: Victor, 1996. 159 p. ISBN 9739731848 2. FLOR-HENRY, P., KOLES, Z. EEG characterisics of normal subjects: a comparasion of men and women and dextrals
and sinistrals. In: Res. Commun. Psychol. And. Psychiat. Bechav., 1982, vol.7, no1, p.21-29. 3. OLĂRESCU, V. Psihoneurologia. Chişinău, 2001. 245 p. ISBN 9975-921-10-8 4. БРАГИНА, Н. Н., ДОБРОХОТОВА, Т.А. Функциональная асимметрия мозга. Москва: Медицина, 1998. 240 с.
ISBN 5-225-00102-5 5. БУРЛАЧУК, Л.Ф., МОРОЗОВ С.М. Словарь–справочник по психологической диагностике. Киев: Наукова
думка, 1989. 200 с. ISBN 5-12-000482-2 6. ГОДФРУА, Ж. Что такое психология: В 2-х т. Т.1 / Пер. с франц. Москва: Мир, 1992. 496 с. ISBN 5-03-001901-4 7. ГОРБАЧЕВ, М.Н. Психологические и психофизиологические особенности восприятия времени людьми с
различным профилем латеральной организации головного мозга: Автореф. дисс. канд. психол. наук. Ростов-на-Дону, 2006.
8. ГОРОШКО, Е.И. Функциональная асимметрия мозга, язык, пол:Аналитический обзор: Монография. Москва: ИД «ИНЖЭК», 2005. 280 с.
9. КИНЯГИНА, О. и др. Мозг на 100%. Интеллект. Память. Креатив. Интуиция. В: Интенсив-тренинг по развитию суперспособностей. Москва: Эксмо, 2011. 848 с. ISBN 978-5-699-49283-1
10. КУРЕПИНА, М.М., ОЖИГОВА, А.А. и др. Анатомия человека. Москва: Владос, 2003. 384 с. ISBN 5-691-00905-2 11. ЛОХОВ, М.И. Заикание: неврология или логопедия? СПб.: ЭЛБИ-СПб., 2005. 600 с. (Пограничные психи-
ческие расстройства у детей). ISBN 5-93979-150-6 12. ЛУРИЯ, А.Р. О функциональном взаимодействии полушарий головного мозга. В: Физиология человека,
1975, №3, c.411-417. 13. МОИСЕЕВА, Н.И., СЫСУЕВА, В.М. Временная среда и биологические ритмы. Ленинград: Наука, 1981. 128 с. 14. ПУГАЧ, В.Н. Функциональная асимметрия мозга: амбидекстрия и амбицеребральность, новые тенденции.
В: Актуальные вопросы функциональной межполушарной асимметрии и нейропластичности: Материалы Всероссийской конференции с международным участием. Москва: Научный мир, 2008, с.79-83.
15. СЕМЕНОВИЧ, А.В. О формировании межполушарного взаимодействия в онтогенезе. В: Тезисы докладов Международной конференции памяти А.Р. Лурии / Под ред. Е.Д. Хомской и др. Москва: Российское психо-логическое общество, 1997, с.85-86.
16. СЕМЕНОВИЧ, А.В., ЦЫГАНОК, А.А. Нейропсихологический подход к типологии онтогенеза. В: Нейро-психология сегодня / Под ред. Е.Д. Хомской. Москва: Изд-во МГУ, 1995, с.170-171.
17. СИМЕРНИЦКАЯ, Э. Г. Доминантность полушарий. Москва: Медия, 2012. 47 с. ISBN 978-5-458-39284-6 18. СПРИНГЕР, С., ДЕЙЧ, Г. Левый мозг, правый мозг / Пер. с англ. Москва: Мир, 1983. 256 c.
Prezentat la 19.11.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
9
APRECIEREA NIVELULUI ELEMENTELOR MINERALE DE CALCIU,
MAGNEZIU ŞI FIER LA SPORTIVII–ÎNOTĂTORI DE PERFORMANŢĂ
PÂNĂ LA ŞI DUPĂ EFORT FIZIC EFECTUAT PE VELOERGOMETRU
Inga DELEU
Universitatea de Stat din Moldova
În acest articol este studiat nivelul elementelor minerale (calciu, magneziu, fier) la sportivii–înotători de performanţă
(fete şi băieţi) înainte şi după efort fizic efectuat pe veloergometru.
În rezultatul testărilor efectuate nivelul elementelor minerale de calciu şi magneziu la majoritatea sportivilor–înotă-
tori până la efort fizic este sub limitele normei, iar nivelul elementului mineral de fier este în limitele normei.
Rezultatele obţinute după efortul fizic efectuat pe veloergometru denotă o scădere esenţială a nivelului de calciu şi
magneziu şi o creştere a nivelului de fier în sângele sportivilor–înotători testaţi.
Cuvinte-cheie: sportiv, înot, elemente minerale, efort fizic, veloergometru, sânge, sportivi de performanţă.
ESTIMATION OF THE MINERAL ELEMENTS OF CALCIUM, MAGNESIUM
AND IRON AT ELITE SWIMMERS BEFORE AND AFTER PHYSICAL EFFORT ON THE BICYCLE
This paper deals with the studying of the levels of mineral elements (calcium, magnesium, iron) at elite swimmers
(boys and girls) before and after physical effort on the bicycle.
As a result of our research it was found out that the level of calcium and magnesium is decreased before the physical
effort at the majority of the swimmers tested, while the level of iron is at a normal degree.
The results obtained after physical effort on bicycle denotes of a further decrease of the level of calcium and magne-
sium and the increase of iron level in the blood of the tested sportsmen.
Keywords: sportsmen, swimmer, mineral elements, physical effort, bicycle, blood, elite swimmers.
Introducere
Înotul este considerat unul dintre cele mai vechi şi mai populare sporturi. Această probă sportivă a apărut
demult, odată cu istoria omului, ca o deprindere necesară asigurării existenţei. Intrarea în apă se impunea ca refugiu în faţa incendiilor, în căutarea hranei sau pentru apărarea vieţii. Pe lângă aceste motivaţii practice,
omenirea a fost întotdeauna fascinată de apă, ca de o forţă necunoscută. Existenţa omului l-a constrâns să înveţe înotul. În anul 2500 î.e.n, ca dovadă a răspândirii înotului serveşte un papirus egiptean, găsit de arheo-
logi, în care apar oameni înotând ce foloseau o tehnică asemănătoare cu cea de craul din zilele noastre [5].
Cultura indiană caracteriza înotul ca mijloc de menţinere a sănătăţii corpului şi a minţii. Referinţe despre
primele metode de învăţare a înotului apar la grecii antici şi la romani. Importanţa acordată înotului este con-
firmată de existenţa bazinelor de înot în gimnazii, unde grecii învăţau înotul. Metodica învăţării, asemănătoa-
re cu cea din zilele noastre, constă în folosirea plutelor şi a mănunchiurilor de trestii, legate în jurul corpului
pentru a favoriza plutirea. Deşi înotul nu figura printre sporturile din cadrul Jocurilor Olimpice ale Greciei antice, primele competiţii
de înot apar în sec. II î. e. n., fapt specificat de istoricul grec Pausanias în lucrarea „Descrierea Greciei” [5,1].
În jurul anului 1200 î.e.n., romanii au perfecţionat modelul grec de bazine, prin construirea termelor, clă-
diri care cuprindeau saune şi bazine cu apă caldă pentru înot şi apă rece pentru sărituri. Termele din vechile cetăţi aveau o capacitate de înot pentru circa 1600 de persoane. Aici aveau loc întreceri, la care participau
atât vârstnicii, cât şi tinerii. În secolul IV e. n. se construiesc până la 850 de terme gigantice [1]. În perioada Renaşterii, înotul este apreciat la un nivel înalt, şi anume: ca mijloc de educaţie fizică şi mora-
lă a tineretului. Alături de călărie, aruncarea lăncii, scrimă şi vânătoare, înotul este dezvoltat de asemenea ca un mijloc de obţinere a titlului de cavaler.
În anul 1837 apare înotul sportiv, în Anglia – „patria înotului modern”. Este anul în care la Londra apare
primul club de înot „National Swimming Association”, fondat de John Strachan. În Londra existau la acea dată 8 bazine de înot. Deja în anul 1840 înotul a fost introdus în instituţiile de învăţământ, iar în anul 1858 în
Australia a avut loc primul Campionat Mondial la înot. În anul 1889 s-au desfăşurat, la Viena, primele cam-pionate europene care au inclus două probe: de 60 m şi 1500 m, amândouă câştigate de austrieci.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
10
La prima ediţie a Jocurilor Olimpice moderne, anul 1896, în program este inclus şi înotul. Cu doisprezece ani mai târziu, în anul 1908, este înfiinţată Federaţia Internaţională de Nataţie pentru Amatori (FINA), care reglementează desfăşurarea competiţiilor de înot, şi anume: stabileşte un regulament internaţional de înot şi elucidează regulile de stabilire a recordurilor. În anul 1951, în Bucureşti, se construieşte primul bazin acope-rit. Ulterior, înotul s-a dezvoltat rapid în toate ţările. Apar înotători cu renume şi recorduri mondiale [8].
Înotul este considerat medicament de selecţie, prin efectele sale miraculoase asupra întregului organism. Este acceptat la toate vârstele, deoarece poate fi practicat atât de copiii cu o vârstă fragedă, cât şi de oamenii în etate. Atunci când facem exerciţii pe pământ, precum alergatul sau mersul, trebuie să ne luptăm cu forţa întregii noastre greutăţi, presiunea exercitată asupra oaselor şi articulaţiilor fiind foarte ridicată.
Starea de plutire în apă acţionează ca un scut protector în jurul corpului. Apa preia tensiunea în exces exercitată asupra oaselor, articulaţiilor şi muşchilor. În timpul înotului, toate grupele musculare sunt puse în mişcare simultan, provocând organismul la un antrenament total.
Se favorizează poziţia corectă pe apă a capului şi corpului, care, asociată cu caracterul simetric şi ciclic al mişcărilor, duce la obţinerea atitudinii corecte a corpului. Acţionează pozitiv asupra mobilităţii coloanei ver-tebrale, întărind şi musculatura trunchiului. Eliberarea coloanei vertebrale de sub greutatea corpului, asociată cu activitatea intensificată a muşchilor respiratori, au o influenţă profilactică, dar şi terapeutică în combaterea deformaţiilor coloanei vertebrale. Înotul este considerat un exerciţiu cu risc scăzut de traumare, dacă este co-rect practicat şi este sportul ideal pentru cei care suferă de osteoporoză, artroză sau de alte boli articulare. Acesta este unul dintre motivele pentru care înotul este recomandat persoanelor supraponderale, femeilor în-sărcinate sau celor care se recuperează după diverse afecţiuni ale aparatului locomotor [6].
Mişcările efectuate în timpul înotului dezvoltă musculatura inimii, respectiv a atriilor şi ventriculelor. Fortificarea muşchiului cardiac influenţează în mod favorabil activitatea inimii şi circulaţia sangvină în gene-ral. Datorită poziţiei orizontale care se capătă în apă şi efortului fizic depus, sângele pompat de inimă ajunge mult mai repede la extremităţi, decât în cazul aflării în poziţie verticală.
În timpul înotului presiunea sistolică scade, iar cea diastolică creşte, adică se îmbunătăţeşte tensiunea arterială diferenţiată, se măreşte elasticitatea vaselor sangvine şi viteza de propulsare a sângelui prin vase. Activând circulaţia sângelui, înotul împiedică depunerea de substanţe grase (colesterină) pe pereţii vaselor sangvine, combătând astfel ateroscleroza [6, 9].
Sub influenţa înotului au loc modificări în compoziţia sângelui. Numărul eritrocitelor se măreşte, ceea ce ne sugerează despre un nivel mai înalt de captare şi transportare a oxigenului către celule, dar şi despre un proces mai efectiv de apărare, datorită creşterii numărului de leucocite.
Înotul acţionează prielnic şi asupra sistemului respirator; astfel, creşte capacitatea de respiraţie. Datorită
presiunii exercitate de apă asupra corpului, şi anume: asupra cutiei toracice, plămânii depun mai mult efort
pentru a funcţiona normal. Elasticitatea cutiei toracice se îmbunătăţeşte şi, ca rezultat, creşte capacitatea vita-
lă a plămânilor. Metabolismul gazos la nivelul alveolelor pulmonare se intensifică. În pereţii alveolelor se
formează mai multe capilare, astfel că organismul poate absorbi un procent mai mare de oxigen din acelaşi
volum de aer. Cu toate că volumul oxigenului inhalat creşte, frecvenţa cardiacă nu se măreşte [3].
Un alt beneficiu este faptul că înotul stimulează sistemul nervos, ceea ce duce la creşterea activităţii cere-
brale. De asemenea, presiunea exercitată de apă asupra corpului este percepută precum un masaj uşor şi rela-
xant ce duce la eliminarea de serotonină. Este de folos pentru oamenii care suferă de depresie. Unii consideră
înotul un mijloc de meditaţie şi eliminare a stresului. Psihologii americani susţin că 70% din cei care practică
acest sport trăiesc intens emoţiile plăcute. Studiile arată că înotătorii au, în general, tensiunea mai mică, iar
nivelul depresiei sau al furiei este, de asemenea, scăzut.
Prin acţiunea uşor excitantă, înotul se recomandă în unele tulburări ale glandelor cu secreţie internă, în
special ale glandei tiroide, prin accelerarea arderii tisulare. Acest efect justifică înotul terapeutic prescris în
obezitate, când metabolismul este încetinit. Potrivit specialiştilor, un minut de înot valorează cât patru minute
de mers. Doctorul Jean Pierre de Mondenard, autorul unui studiu în domeniu, a făcut următorul calcul ştiinţi-
fic: pentru îndepărtarea unui kilogram de grăsime sunt necesare peste 96 ore de marş, 18 ore de alergare sau
8 ore de nataţie [3,6].
Sportul dat favorizează activitatea sistemul digestiv, prin faptul că, înotând, se măreşte lucrul tuturor muş-
chilor, inclusiv al celor abdominali, şi, ca rezultat, creşte peristaltica intestinală. Procesul de digerare şi asi-
milare a hrănii se îmbunătăţeşte de pe urmele creşterii activităţii fermentative. Înotul este binevenit, mai ales,
în cazul constipaţiilor.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
11
Un rol important este acordat temperaturii apei, care exercită o influenţă pozitivă asupra organismului în întregime. Mediul acvatic constituie pentru organismul uman un factor de călire alături de aer şi soare. Este
cunoscut faptul că apa are temperatura aproximativ cu 2oC mai joasă decât cea a mediului înconjurător, ceea
ce influenţează termoreglarea organismului. Comasarea celor 3 factori duce la creşterea rezistenţei generale a organismului şi la menţinerea unei stări optime de sănătate [9].
După o activitate îndelungată (şcolară sau profesională), organismul, are nevoie de o perioadă de refacere, pentru întărirea stării de sănătate şi cu scopul reluării activităţii de zi cu zi. În cazul refacerii, înotul este spor-
tul cel mai potrivit şi efectiv. Totuşi, fiind o probă sportivă foarte benefică pentru organism, trebuie să ţinem seama şi de unele precau-
ţii, ca de exemplu: nu se poate de înotat după ce aţi luat masa şi, invers, nu se poate de înotat dacă nu aţi mâncat nimic 5 ore; nu se poate de intrat brusc în apă rece după o expunere îndelungată la soare sau după
efort fizic intens. Pentru a ajunge la performanţe sportive, practicarea profesională a înotului trebuie să fie coordonată de
către antrenor. Însă, un lucru foarte important pentru evaluarea sportivilor pe plan profesional este alimenta-ţia raţională şi corectă a sportivilor.
Lucrarea dată este consacrată studiului unor indici biochimici la înotătorii de performanţă, până la efort fizic şi după efort efectuat pe veloergometru.
Scopul cercetărilor efectuate rezidă în testarea sportivilor-înotători de performanţă (fete şi băieţi) după nivelul elementelor minerale extrem de importante, şi anume: nivelul calciului, magneziului şi ferului, până
la efort fizic şi după efort efectuat pe veloergometru. La cercetările efectuate au participat 10 sportivi–înotători de performanţă (5 fete şi 5 băieţi). Calificarea
sportivă a înotătorilor testaţi variază de la maestru în sport până la maestru în sport de clasă internaţională.
În procesul de colectare şi prelucrare a materialului experimental au fost analizate datele despre anamneza sportivă şi indicii de bază ai sportivilor-înotători, precum şi dinamica lor pe parcursul anilor de antrenament.
Metode de cercetare: Pentru testarea sportivilor-înotători a fost folosită metoda biochimică de determi-nare a nivelului de calciu, magneziu şi fier în serul sangvin. Testările au fost efectuate la analizatorul biochi-
mic semiautomat Mindray BA - 88A. Datele obţinute au fost prelucrate statistic.
Rezultate şi discuţii
Alimentaţia raţională constituie unul dintre factorii de echilibru al organismului, asigurând printr-un metabolism normal o bună dezvoltare a acestuia. În urma alimentaţiei corecte se măreşte capacitatea de adap-
tare la efort şi la condiţiile mediului înconjurător, asigurându-se o stare de sănătate perfectă. Pentru realizarea performanţelor sportive de mare valoare sunt importante principiile alimentaţiei îndestulate cu toate compo-
nentele alimentare necesare. Situaţiile specifice, de exemplu, în cazul antrenamentelor sportive intense, tre-buie tratate cu grijă deosebită în ceea ce priveşte dieta pe care o solicită. În acest caz, se selectează o dietă
nutriţională semnificativă pentru sportivi [2]. Este evident faptul că obiceiurile alimentare ale unui sportiv pot determina în mod decisiv calitatea execu-
tării unui exerciţiu fizic. Substanţele mineralele sunt foarte necesare pentru organismul omului, dar mai ales pentru organismul unui sportiv de performanţă. Ele prezintă componente metabolizate în decursul activităţii
vitale a organismului. Fiind ingerate odată cu hrana, constituie elementele de structură a scheletului şi a ţesu-
turilor moi, dar şi factori ce reglează numeroase funcţii fiziologice. Anumiţi ioni au în organism locuri speci-ale de depozitare, de unde pot fi utilizaţi în perioadele în care aportul lor în alimente este insuficient.
Organismul omului, ca şi majoritatea organismelor animale, are proprietatea de a menţine aproximativ constantă concentraţia ionilor minerali în lichidele corpului, asigurând astfel aşa-numitul ,,mediul intern”
constant. Reglarea concentraţiei sărurilor în organism joacă un rol deosebit de important în procesele fiziolo-gice, decizie pentru buna funcţionare a organelor şi ţesuturilor. Ele contribuie la reglarea activităţii celulare
în raport cu necesităţile organismului, cum ar fi reglarea echilibrului acido-bazic, contracţia musculară, sti-mularea sau inhibarea activităţii enzimelor, excitabilitatea nervoasă, coagularea sângelui, transportul oxige-
nului etc. Elementele minerale, numite bioelemente, se regăsesc în aproximativ toate celulele şi participă activ la menţinerea homeostaziei generale. Apariţia unui dezechilibru acut al acestor minerale poate duce la
efecte deosebit de grave asupra sănătăţii [7]. Un interes deosebit pentru organismul unui sportiv reprezintă conţinutul mineralelor de calciu, magneziu
şi fier. Printre elementele care intră în organismul uman, calciul ocupă, în ceea ce priveşte cantitatea, locul 5,

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
12
după elementele de bază: oxigenul, hidrogenul, carbonul şi azotul. Calciul este considerat ,,regele bioelemen-telor” în organismul uman. 99% din cantitatea de calciu se găseşte în oase şi în ţesuturile tari şi 1% în sânge,
în celule şi în ţesuturile moi. Calciul este elementul care întăreşte întreg organismul. În formă ionică, calciul
intră şi iese cu uşurinţă din celula umană. Ionul de Ca2+
, penetrat în celulă, duce după sine un lanţ de ele-mente nutritive. Calciul neionizat se asimilează foarte greu, iar pentru a fi asimilat de organism el numaide-
cât trebuie să treacă în formă ionică. Procesul de trecere a calciului neionizat în formă ionică necesită o can-titate mare de energie şi acid gastric.
Celulele produc permanent energie şi în rezultatul acestor procese se formează elementele nocive – rezi-
duuri, de natură acidă. Calciul este considerat elementul primordial în eliminarea acestor toxine. Prin elimi-
narea resturilor acide, calciul ionizat susţine pH-ul sângelui în stare slab alcalină, ceea ce este foarte impor-
tant pentru menţinerea vieţii, deoarece coborârea pH-ului sângelui până la 6,8 finisează cu decesul. Deci, una
dintre funcţiile principale ale calciului este menţinerea organismului în stare semialcalină, adică menţinerea
vieţii [2,7].
Calciul joacă un rol deosebit în procesul de trecere a oxigenului în interiorul celulelor. În anul 1932 Otto
Warburg a primit Premiul Nobel în chimie pentru dovada adusă, precum că procesul de dezvoltare a canceru-
lui este un proces anaerob. Stoparea accesului de oxigen, în cazul insuficienţei de calciu, face lichidele din
organism acide.
În cazul când folosim cafea, băuturi nealcoolizate dulci, pH-ul cărora este de doar 2,2-2,5, adică pH la
care viaţă nu există, lichidele din interiorul organismului uman se acidifică. Organismul caută permanent re-
zerve de substanţe alcaline, pentru neutralizarea cantităţii mari de acizi, iar rezervele sunt oasele, dinţii, părul.
Putem spune că în cazul alimentării cu produse, care provoacă acidificarea organismului, el pierde o cantitate
foarte mare de calciu [4].
Calciul este necesar pentru coagularea normală a sângelui, contractarea şi relaxarea muşchilor inimii şi a
muşchilor scheletici, pentru reglarea funcţiei fermentative şi, parţial, pentru secreţia insulinei. Un rol impor-
tant îi revine acestui element în transmiterea impulsurilor nervoase, iar insuficienţa lui provoacă iritarea ner-
vilor. După părerea specialiştilor, o cantitate suficientă de calciu, în perioada tinereţii, menţine organismul
tânăr în decursul multor ani.
Calciul se conţine în lapte şi în produsele lactate, în gălbenuşul de ou, conopidă, seminţele de susan, in,
tofu; în ierburile uscate de cimbru, busuioc, mărar, salvie, rozmarin, oregano, pătrunjel; în legume, aşa ca:
soia, fasolea, mazărea, vânăta, rodia. Însă, pentru a fi asimilat şi pentru a-şi exercita funcţiile, calciul are
nevoie de câteva ajutoare, adică de alte vitamine şi minerale, ca vitaminele D, A, C şi magneziu.
Lipsa de calciu se simte prin senzaţii de amorţeală sau furnicături la nivelul membrelor, precum şi con-
tracturile musculare. Formele mai avansate ale lipsei de calciu sunt osteoporozele, cariile dentare, unghii cu
puncte albe, oboseală generală a organismului, palpitaţii, stare de iritabilitate psihică, senzaţia de lipsă de aer,
insomnia [4].
Calciul este deosebit de important pentru organismul sportivilor, deoarece în timpul antrenamentului ei
pierd o cantitate enormă de calciu prin transpiraţie. Norma elementului mineral de calciu în serul sangvin
este de 2,1–2,6 mmol/l.
Un alt element necesar organismului uman, dar mai ales sportivilor, este magneziul. Funcţia lui primară
constă în participarea la dezvoltarea conductibilităţii şi contractilităţii muşchilor. Magneziul intervine în
reacţiile fiziologice (metabolismul glucidelor, lipidelor şi proteinelor) şi în activităţile enzimatice, în permea-
bilitatea celulară, coagularea sangvină. Mai mult de jumătate din cantitatea totală de magneziu se găseşte în
oase, un sfert se găseşte în muşchi, iar restul se repartizează în inimă, ficat, rinichi, tubul digestiv şi în plas-
ma sangvină. În cazul în care nivelul de magneziu scade sub normă, tabloul clinic al bolii este caracterizat
prin hiperexcitabilitate, spasmofilie şi astenie, dureri de cap, ameţeli, tremurături, insomnie, senzaţie de fur-
nicături sau amorţeală a membrelor, cârcei, scăderea tensiunii arteriale, aritmie cardiacă, palpitaţii cardiace,
reacţii alergice frecvente, scăderea rezistenţei la infecţii, hipoglicemie ş.a. [2,4 ].
Surse naturale de magneziu sunt: cerealele intergale (porumb, grâu, ovăs, orz, secară), nucile, alunele,
smochinele, migdalele, merele, legumele verzi, laptele, ouăle, crustaceele, ciocolata.
Magneziul se asimilează foarte bine atunci când este asociat cu vitamina B6, motiv pentru care consuma-
rea zilnică de cereale integrale, care sunt bogate în vitaminele din complexul B, prezintă un tratament foarte
bun. Alimentaţia cu multă carne împiedică asimilarea magneziului, fixând ireversibil magneziul intestinal.
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
13
Consumul de alcool provoacă lipsă de magneziu, deoarece acesta va fi eliminat masiv prin diureză. Necesita-
tea de magneziu este mult mai mare în cazul sportivilor. Norma elementului mineral de magneziu în serul
sangvin este de 0,66–1,07 mmol/l.
Fierul prezintă al treilea element mineral testat. Elementul dat este necesar pentru formarea hemoglobinei,
mioglobinei şi a unor enzime. Cu ajutorul fierului se asigură nutriţia mucoaselor, secreţia gastrică, respiraţia
celulară. Fierul ajută la procesul de creştere, măreşte rezistenţa la îmbolnăviri, mai ales la infecţii, transportă
diferite substanţe chimice implicate în metabolismul energetic, are acţiune antioxidantă, asigură funcţia re-
productivă. Norma elementului mineral de fier este de 9,0–30,4 mmol/l.
Semnele carenţei de fier sunt: anemie, iritabilitate, slăbiciune, oboseală, ameţeală, dureri de cap (mai ales
în zona frunţii). În cazul unei lipse grave de fier, apare o astenie marcată, pierderea luciului şi solidităţii un-
ghiilor, uscăciunea gurii cu apariţia unor leziuni, dureri la înghiţire, senzaţie de lipsă de aer, palpitaţii, tulbu-
rări grave de ciclu menstrual [4,7].
Fierul se conţine în ficat, ouă, lapte de vacă, iaurt, caşcaval, caş, banane, spanac, avocado, asparagus, broc-
coli, ciuperci, rădăcină de pătrunjel, stafide şi caise uscate, pâine/cereale integrale, fasole, mazăre, arahide ş.a.
În ciuda dietei foarte bogate în carne a omului modern, cazurile de anemie fieriprivă sunt tot mai frecvente,
afectând 20% din femei, 3% din bărbaţi, 50% din gravide şi aproape 15% din adolescenţi. Cauza anemiei nu
este aportul insuficient de fier, ci deficienţele în asimilaţie provocate de intoxicarea colonului prin consumul
de proteine animale, perturbarea echilibrului endocrin prin consumarea de alimente cu aditivi sintetici, consu-
mul de cafea, ceai negru, abuz medicamentos [2].
Foarte importantă pentru organismul înotătorilor a fost aprecierea nivelului de elemente minerale (calciu,
magneziu şi fier) din serul sangvin la fetele–înotătoare şi la băieţii-înotători de performanţă.
Tabelul 1
Nivelul elementelor minerale de calciu, magneziu şi fier la fetele–înotătoare până la efort fizic
Nr.
d/o
Numele, prenumele Ca
(mmol/l)
Mg
(mmol/l)
Fe
(mmol/l)
1 P. T., 15 ani, m/s 1,94 0,91 18,9
2 T. E., 18 ani, m/s 1,87 0,44 14,8
3 C. T., 16 ani, m/s 1,72 0,34 14,2
4 D.G., 14 ani, m/s 1,64 0,56 13,7
5 S. I., 13 ani, m/s 1,40 0,48 14,7
6 M ± m 1,71 ± 0,08 0,54 ± 0,08 15,26 ± 0,92
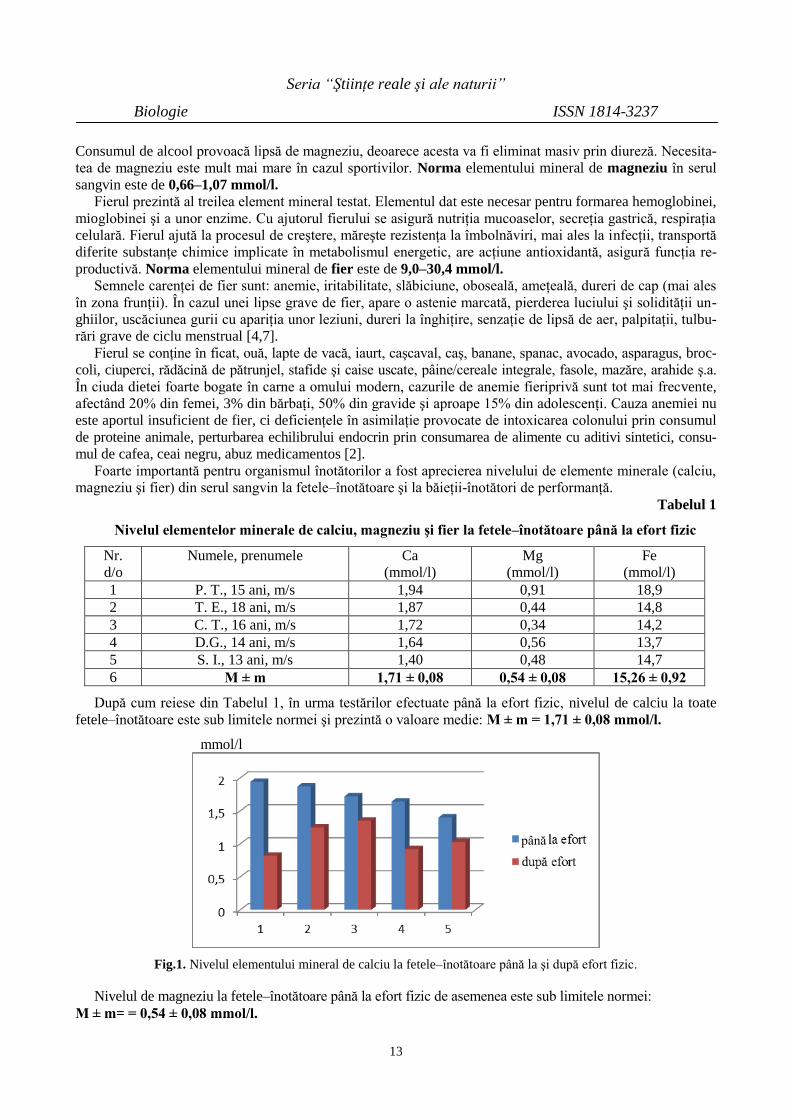
După cum reiese din Tabelul 1, în urma testărilor efectuate până la efort fizic, nivelul de calciu la toate
fetele–înotătoare este sub limitele normei şi prezintă o valoare medie: M ± m = 1,71 ± 0,08 mmol/l.
mmol/l
Fig.1. Nivelul elementului mineral de calciu la fetele–înotătoare până la şi după efort fizic.
Nivelul de magneziu la fetele–înotătoare până la efort fizic de asemenea este sub limitele normei:
M ± m= = 0,54 ± 0,08 mmol/l.
până

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
14
mmol/l
Fig.2. Nivelul elementului mineral de magneziu la fetele–înotătoare până la şi după efort fizic.
Nivelul fierului la fetele–înotătoare este în limitele normei cu o valoare medie de M ± m = 15,26 ± 0,92 mmol/l.
mmol/l
Fig.3. Nivelul elementului mineral de fier la fetele–înotătoare până la şi după efort fizic.
La băieţii – înotători de performanţă nivelul elementului mineral de calciu pînă la efort fizic este puţin sub
limitele normei, iar la un sportiv înotător (C.E.) prezintă norma.
Tabelul 2
Nivelul elementelor minerale de calciu, magneziu şi fier la băieţii-înotători până la efort fizic
Nr.
d/o
Numele, prenumele Ca,
mmol/l
Mg,
mmol/l
Fe,
mmol/l
1 A. D., 17 ani, m/s 1,65 0,38 22,1
2 C. E., 17 ani, m/s 2,18 0,93 27,2
3 N. V., 17 ani, m/s 1,71 0,34 17,1
4 B. D., 16 ani, m/s 1,98 0,53 17,2
5 A. D., 18 ani, m/s 1,34 0,42 16,8
6 M ± m 1,77 ± 0,13 0,52 ± 0,09 20,08 ± 0,91
Deci, valoarea medie de calciu la băieţii–înotători este: M ± m = 1,77 ± 0,13 mmol/l.
mmol/l
Fig.4. Nivelul elementului mineral de calciu la băieţii–înotători până la şi după efort fizic.
până
până
până
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
15
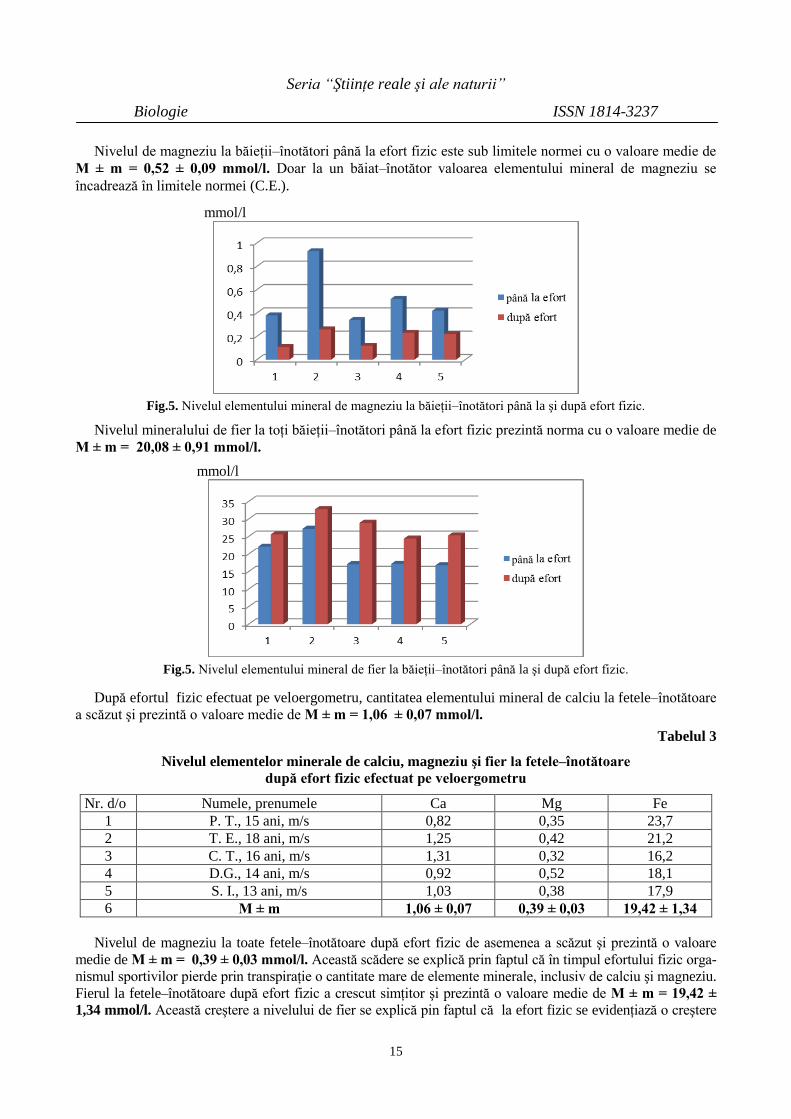
Nivelul de magneziu la băieţii–înotători până la efort fizic este sub limitele normei cu o valoare medie de
M ± m = 0,52 ± 0,09 mmol/l. Doar la un băiat–înotător valoarea elementului mineral de magneziu se
încadrează în limitele normei (C.E.).
mmol/l
Fig.5. Nivelul elementului mineral de magneziu la băieţii–înotători până la şi după efort fizic.
Nivelul mineralului de fier la toţi băieţii–înotători până la efort fizic prezintă norma cu o valoare medie de
M ± m = 20,08 ± 0,91 mmol/l.
mmol/l
Fig.5. Nivelul elementului mineral de fier la băieţii–înotători până la şi după efort fizic.
După efortul fizic efectuat pe veloergometru, cantitatea elementului mineral de calciu la fetele–înotătoare
a scăzut şi prezintă o valoare medie de M ± m = 1,06 ± 0,07 mmol/l.
Tabelul 3
Nivelul elementelor minerale de calciu, magneziu şi fier la fetele–înotătoare
după efort fizic efectuat pe veloergometru
Nr. d/o Numele, prenumele Ca Mg Fe
1 P. T., 15 ani, m/s 0,82 0,35 23,7
2 T. E., 18 ani, m/s 1,25 0,42 21,2
3 C. T., 16 ani, m/s 1,31 0,32 16,2
4 D.G., 14 ani, m/s 0,92 0,52 18,1
5 S. I., 13 ani, m/s 1,03 0,38 17,9
6 M ± m 1,06 ± 0,07 0,39 ± 0,03 19,42 ± 1,34
Nivelul de magneziu la toate fetele–înotătoare după efort fizic de asemenea a scăzut şi prezintă o valoare
medie de M ± m = 0,39 ± 0,03 mmol/l. Această scădere se explică prin faptul că în timpul efortului fizic orga-
nismul sportivilor pierde prin transpiraţie o cantitate mare de elemente minerale, inclusiv de calciu şi magneziu.
Fierul la fetele–înotătoare după efort fizic a crescut simţitor şi prezintă o valoare medie de M ± m = 19,42 ±
1,34 mmol/l. Această creştere a nivelului de fier se explică pin faptul că la efort fizic se evidenţiază o creştere
până
până

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
16
a numărului de elemente sangvine, inclusiv de eritrocite, din cauza necesităţii mult mai mari în oxigen a orga-
nismului. Odată cu creşterea numărului de eritrocite creşte şi cantitatea de hemoglobină, ca urmare se măreşte
şi cantitatea de fier în sânge, element component al hem-ului (partea anorganică a hemoglobinei).
După efortul fizic efectuat pe veloergometru cantitatea de calciu din serul sangvin la băieţii–înotători a
scăzut esenţial şi prezintă o valoare medie de M ± m = 0,65 ± 0,07 mmol/l.
Tabelul 4
Nivelul elementelor minerale de calciu, magneziu şi fier la băieţii-înotători
după efort fizic efectuat pe veloergometru
Nr.
d/o
Numele, prenumele Ca Mg Fe
1 A. D., 17 ani, m/s 0,60 0,11 25,6
2 C. E., 17 ani, m/s 0,64 0,26 32,8
3 N. V., 17 ani, m/s 0,48 0,12 28,9
4 B. D., 16 ani, m/s 0,98 0,23 24,4
5 A. D., 18 ani, m/s 0,56 0,22 25,3
6 M ± m 0,65 ± 0,07 0,18 ± 0,001 27,4 ± 1,55
Aceeaşi situaţie se stabileşte şi la nivelul mineralului de magneziu, unde cantitatea medie a scăzut până la
o valoare de M ± m = 0,18 ± 0,001 mmol/l. În cazul mineralului de fier, după efort fizic, la băieţii–înotători,
precum şi la fetele-înotătoare, s-a stabilit o situaţie inversă, adică cantitatea de fier din serul sangvin s-a mărit
esenţial. Aceasta demonstrează încă odată că este depus un efort de o intensitate înaltă, în urma căruia
organismul necesită o cantitate avansată de oxigen, care este distribuită prin intermediul hemoglobinei.
Concluzii
1. Substanţele minerale, împreună cu proteinele, carbohidraţii, grăsimile şi vitaminele, sunt foarte necesare
pentru o bună funcţionare a proceselor fiziologice şi biochimice din organism, care stau la baza tuturor
funcţiilor vitale ale organismului. Unele dintre cele mai importante fiind elementele minerale de calciu,
magneziu şi fier.
2. În urma aprecierii nivelului de calciu şi magneziu până la efort fizic, atât la fetele-înotătoare, cât şi la
băieţii-înotători, s-a detectat un nivel sub limitele normei. Nivelul de fier la toţi sportivii-înotători (băieţi
şi fete) până la efort fizic se încadrează în limitele normei.
3. După efortul fizic cantitatea de calciu şi de magneziu a scăzut esenţial, în urma transpiraţiei, iar nivelul
fierului a crescut, ca rezultat al necesităţii avansate în oxigen.
4. Conform datelor actuale, majoritatea cazurilor de disfuncţii la nivelul schimbului de substanţe minerale la
sportivi pot fi clasificate ca profesionale sau profesional asociate, legate de eforturile fizice şi psihoemo-
ţionale foarte ridicate. Deci, efortul fizic duce la pierderea unor elemente minerale şi, ca rezultat, se cere
creşterea venitului de substanţe minerale ingerate odată cu hrana.
Bibliografie:
1. BOTNARENCO, F., RÎŞNEAC, B. ŞARPOV, T. Înotul. Chişinău: Lumina, 1991. 172 p.
2. DRĂGAN, I., STROESCU, V. Medicaţia în efort fizic. Bucureşti: Editis, 1993. 117 p.
3. FIELDER, P., FETESCU, S. Înot. Iaşi: Universitatea ,,Alexandru Ioan Cuza”, 1996. 242 p.
4. HARALAMBIE, Gh. Biochimie şi sport. Bucureşti: Teora, 1966. 231 p.
5. MUREŞAN, E., PARASCHIV, I.C. Înotul utilitar–aplicativ şi tehnici de vâslit. Bucureşti: Editura Fundaţiei România
de Mâine, 2002. 68 p.
6. PENTIA, D. Îndrumar metodic de înot. Timişoara: Universitatea Timişoara, 1994. 37 p.
7. VAJIALĂ, G.E. Curs de biochimia efortului. Bucureşti: Editura Fundaţiei România de Mâine, 1996. 152 p.
8. БУЛГАКОВА, Н. Плавание. Москва: Астрель, 2005. 157 с.
9. Плавание / Под общей редакцией профессора Н. Булгаковой. Москва: Физкультура и спорт, 2001. 400 с.
Prezentat la 20.10.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
17
ОСОБЕННОСТИ АЗОТИСТОГО ОБМЕНА У ЛИЦ
С ПСИХИЧЕСКОЙ ДИЗОНТОГЕНИЕЙ
Анна ЛЕОРДА, Светлана ГАРАЕВА, Влада ФУРДУЙ,
Галина РЕДКОЗУБОВА, Галина ПОСТОЛАТИ, Нина КОВАРСКАЯ
Институт физиологии и санокреатологии АН Молдовы
PARTICULARITĂŢILE METABOLISMULUI AZOTAT LA PERSOANELE
CU DISONTOGENEZĂ PSIHICĂ În articol sunt investigate particularităţile metabolismului aminoacizilor liberi la persoanele cu disontogeneză psihică.
Sunt determinaţi, prin metoda cromatografiei lichide cu schimb de ioni, indicii metabolismului azotat în sângele şi în urina copiilor cu paralizie cerebrală infantilă. La copiii cu paralizie cerebrală infantilă a fost depistată o sporire conside-rabilă a coeficientului raportului aminoacizilor de excitaţie/inhibiţie, precum şi hipoaminoacidemie şi hipoaminoaci-durie. Acestea sunt însoţite de o disbalanţă pronunţată a aminoacizilor cu conţinut de sulf, ceea ce permite utilizarea lor în calitate de markeri nespecifici ai nivelului de sănătate. Rezultatele obţinute sunt discutate în cadrul conceptului de sănătate psihică de pe poziţiile abordării sanocreatologice a formării dirijate a sănătăţii mentale a omului.
Cuvinte-cheie: sănătate mentală, disontogeneză psihică, sanocreatologie, paralizie cerebrală infantilă, metabolism azotat, aminoacizi cu conţinut de sulf, aminoacizi de excitaţie, aminoacizi de inhibiţie.
FEATURES OF NITROGEN METABOLISM IN INDIVIDUALS WITH
MENTAL DIZONTOGENY
The article investigates the characteristics of the exchange of free amino acids in individuals with mental dizontogeny. There were detected the data of nitrogen metabolism in the serum and urine of children with cerebral palsy morning by ion exchange HPLC. It is shown a significant increase in the coefficient of the braking/excitatory amino acids ratio, hipoaminoacidemia and hipoaminoaciduria children with cerebral palsy. This is accompanied by severe imbalance of the sulfur amino acids, which allows using them as non-specific marker level of health.
Results are discussed in terms of the concept of mental health positions of sanocreatology approach to the formation of a focused mental health.
Keywords: mental health, mental dizontogeniya, sanocreatology, cerebral palsy, nitrogen metabolism, sulfur, exciting, brake amino acids.
Введение Концепция психического здоровья, разработанная в Институте физиологии и санокреатологии
АНМ под руководством академика Ф.И. Фурдуя, раскрывает новые физиологически обоснованные подходы к разработке научных основ и методов целенаправленного формирования психического здо-ровья человека. Согласно этой концепции, психическое здоровье реализуется нейрофизиологической интегральной и нейрохимической системами мозга, детерминированными генетической программой развития организма, а также психофункциональными и оценочно-исполнительными системами, фор-мирующимися в онтогенезе под влиянием факторов окружающей среды [12, 13]. Иначе говоря, ста-новление и развитие психического здоровья зависит от взаимодействия организма с окружающей средой, степени и длительности воздействия экологических и социальных факторов, от возрастного периода, в котором они оказали свое влияние.
Одним из следствий воздействия вышеуказанных факторов являются нарушения соподчиненности функций жизненно важных органов и систем жизнеобеспечения организма как единого целого, инте-грального взаимодействия функций различной модальности [14, 15,16] и координационной деятель-ности анализаторов. Эти нарушения приводят к ограничению восприятия на уровне ощущений [2, 8], что указывает на наличие, наряду с другими дисфункциями, выраженной сенсорной депривации, в результате чего может развиться психический дизонтогенез [1]. Общей моделью психической дизон-тогении на почве нарушений со стороны опорно-двигательной системы может служить детский цере-бральный паралич (ДЦП), развивающийся в результате поражения по разным причинам головного и спинного мозга на ранних стадиях внутриутробного развития плода и в процессе родов [19]. Для детей с ДЦП характерно отставание в формировании тех или иных психических процессов (внимания, вос-приятия, памяти, мышления), что тормозит психическое развитие ребенка в целом [4, 6, 19].

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
18
Согласно концепции феноменологии психического здоровья, составляющая его структура пред-
ставлена нейрофизиологическими, эмоционально-чувственными, коммуникативно-поведенческими и
личностно-смысловыми реакциями. Формирование и поддержание психического здоровья происхо-
дит за счет мобильной модификации нейрональных метаболических процессов (синтеза и транслока-
ции мембранных и структурных белков, медиаторов и секреции гормонов, катехоламинов), электро-
химических реакций в структурных элементах ЦНС (определяющих возбудимость, проводимость,
формирование энграмм), стабилизации метаболических процессов, определяющих длительные плас-
тические преобразования в синапсах, и др. [13]. В настоящее время сформулирована также концепция
дисбаланса медиаторно-модуляторных субсистем как одного из основных патогенетических факторов
формирования полиморфизма двигательных нарушений [4]. В механизмы патогенеза ДЦП вовлечены
медиаторные субсистемы головного мозга, в частности дофаминовая, холинэргическая, серотонинэр-
гическая, ГАМК-эргическая и глутаматная [5, 9]. Многочисленные данные свидетельствуют о том,
что нейромедиаторные аминокислоты и их рецепторы принимают участие в формировании фунда-
ментальных процессов нервной деятельности, в развитии процессов мышления, обучения, памяти и
др. [5, 7]. Нарушения обмена аминокислот в организме являются одной из причин развития дисфунк-
ций нервной системы, провоцирующих возникновение ряда нервных и психических заболеваний,
особенно в детском возрасте [7, 9, 10]. Исходя из этого, нами была поставлена задача выявить особен-
ности азотистого обмена у детей с ДЦП и его влияние на дизонтогенез психического здоровья.
Материалы и методы исследования
Исследованы показатели азотистого обмена (АО), в частности – свободных аминокислот (САК) в
сыворотке крови и утренней моче у детей, ранжированных в 2 группы (до 2 лет и 4-10 лет) с установ-
ленным диагнозом ДЦП. Показатели АО у здоровых детей этого же возраста практически не разни-
лись, что позволило объединить их в одну контрольную группу. Исследование проб проводилось ме-
тодом жидкостной хроматографии на аминокислотном анализаторе ААА 339М. Полученные данные
обработаны с помощью критерия Стъюдента.
Результаты и их обсуждение
Полученные результаты определения продуктов АО в сыворотке крови обследованных детей
отражены на таблице 1.
Таблица 1
Сравнительное содержание показателей АО в сыворотке крови детей
контрольной группы и с ДЦП (мкмоль/100мл)
Показатели азотистого обмена Контрольная
группа
Дети
до 2 лет
Дети
4 - 10 лет
Цистеиновая кислота 0,79±0,05 1,69±0,29* 2,08±0,35*
Таурин 13,48±0,48 6,28±1,19* 8,05±1,69*
Аспарагиновая кислота 4,77±0,26 2,18±0,41* 1,97±0,39*
Треонин 12,63±0,76 5,32±1,28* 6,43±1,35*
Серин 11,57±0,55 10,00±2,20* 9,88±1,78
Аспарагин 11,10±0,65 4,45±0,89* 4,57±1,01*
Глутаминовая кислота 17,71±1,16 9,10±2,09* 9,54±1,72*
Глутамин 46,56±2,25 25,26±4,30* 22,38±4,48*
α-аминоадипиновая кислота 0,37±0,03 0,30±0,06* 0,44±0,09
Пролин 18,01±1,59 9,17±1,83* 13,45±2,69
Глицин 18,48±1,56 17,43±3,66 19,64±4,13
Аланин 24,95±1,88 17,32±3,81* 26,48±5,56
Цитруллин 2,12±0,26 1,63±0,37* 2,76±0,55
α-аминомасляная кислота 1,11±0,11 1,01±0,18 2,07±0,43*
Валин 15,96±1,51 11,42±1,71* 14,45±2,60
Цистеин 2,54±0,32 5,51±0,61* 8,88±1,51*
Гомоцистеин 0,59±0,03 0,27±0,06* 1,96±0,45*
Метионин 1,86±0,42 1,01±0,21* 1,09±0,19*

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
19
Изолейцин 6,63±0,42 2,40±0,43* 3,31±0,60*
Лейцин 11,88±1,34 4,43±0,97* 6,90±1,31
Тирозин 7,99±0,50 5,68±1,02* 3,84±0,92*
Фенилаланин 8,44±0,56 2,61±0,52* 4,12±0,91*
γ-аминомасляная кислота 0,28±0,03 0,18±0,04* 0,28±0,06
Этаноламин 1,83±0,08 1,46±0,10* 3,98±0,92*
Триптофан 4,08±0,39 2,31±0,23* 2,68±0,46*
Орнитин 6,60±0,42 3,96±0,51* 3,43±0,65*
Лизин 24,10±1,65 8,94±1,43* 7,64±0,84*
Гистидин 18,24±1,26 5,27±1,11* 7,38±1,70*
Аргинин 11,37±0,98 5,95±1,31* 6,06±1,27*
Мочевина 377,47±13,74 103,86±21,81* 205,82±43,22*
Аммиак 19,10±2,60 59,54±11,31* 44,68±7,15*
САК 305,98±17,87 129,69±22,05* 154,02±29,26*
Сумма показателей азотистого обмена 702,55±35,34 239,08±43,04* 315,04±63,01*
Заменимые аминокислоты 163,66±8,27 79,38±17,46* 90,42±19,89*
Незаменимые аминокислоты 115,17±8,28 38,54±9,25* 47,20±10,86*
Иммуноактивные аминокислоты 105,57±7,51 50,93±10,19* 66,24±11,92*
Гликогенные аминокислоты 88,35±6,02 49,07±10,31* 61,63±9,24*
Кетогенные аминокислоты 63,11±5,56 19,72±3,94* 21,12±4,01*
Протеиногенные аминокислоты 278,83±22,84 117,93±24,76* 137,62±9,63*
Серосодержащие аминокислоты 19,24±1,38 9,16±1,83* 14,91±1,49
К тормозные/возбуждающие САК 1,21±0,08 1,76±0,30* 1,97±0,36*
* Р < 0,05
Анализ данных таблицы выявил достоверное и значимое пониженное содержание показателей АО
в обеих возрастных группах детей с ДЦП по сравнению с группами здоровых детей в 2,9 и 2,2 раза
соответственно. При этом обнаружено однонаправленное изменение содержания конечных продуктов
АО: уровень мочевины в 1 группе (до 2 лет) снижен в 3,6 раза, во 2 группе (4-10 лет) в 1,8 раза; в то
же время уровень аммиака значительно повышен: в 1 группе в 3,1 раза, во 2 группе – в 2,3 раза. Так,
соотношение мочевина/аммиак в крови у детей с ДЦП обеих групп резко снижено (в 11,3 и 4,3 раза
соответственно). Это указывает на затруднение утилизации аммиака мочевиной, накопление кото-
рого, в свою очередь, как известно, влияет на клетки ЦНС, вызывая отек нейроглии и дисфункцию
астроцитов.
Пул САК в сыворотке крови в обеих возрастных группах детей с ДЦП по сравнению с группами
сравнения достоверно снижен в 2,4 и 2,0 раза. Наиболее значительно падает уровень таурина (в 2,1 и
1,7 раза в 1 и 2 группах соответственно), аспартата (в 2,2 и 2,4 раза), аспарагина (в 2,5 и 2,4 раза),
глутамата (в 2,1 и 1,9 раза), глутамина (в 1,8 и 2,1 раза), фенилаланина (в 3,2 и 2,0 раза), лизина (в 2,7
и 3,2 раза), гистидина (в 3,5 и 2,5 раза), метионина (в 1,8 и 1,7 раза), изолейцина (в 2,8 и 2,0 раза),
лейцина (в 2,7 и 1,7 раза), тирозина (в 1,4 и 2,1 раза), триптофана (в 1,8 и 1,5 раза), аргинина (в 1,9 и
1,9 раза), орнитина (в 1,7 и 1,9 раза). В то же время достоверно повышена концентрация цистеиновой
кислоты (в 2,1 и 2,6 раза) и цистина (в 2,2 и 3,5 раза).
Количественный анализ различных функциональных групп аминокислот выявил ряд их изменений.
Так, содержание САК с разветвленной цепью в крови детей с ДЦП значительно снижено (в 1,9 и 1,4
раза соответственно). Эти незаменимые аминокислоты активно участвуют в синтезе белка, особенно
в мышечной ткани, стимулируют поступление других САК в мышцы, в синтезе гормона роста, акти-
визируют обмен инсулина и метаболитов цикла Кребса, а также играют важную роль в энергетике и
метаболизме нервных клеток [8]. Наконец, эти САК активируют глутаматдегидрогеназу, обеспечиваю-
щую нормальное проведение нервных импульсов, поэтому их недостаток может вызывать мышечную
гипотрофию при ДЦП.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
20
Особое значение в азотистом метаболизме имеют дикарбоновые аминокислоты, содержание кото-
рых при ДЦП достоверно снижено (аспартата в 2,2 и 2,4 раза соответственно и глутамата в 1,9 раза в
обеих группах). Глутамат и аспартат принимают участие в обезвреживании аммиака [17], наряду с
глюкозой являются источником АТФ для головного мозга [5]. Глутамат является самостоятельным
нейромедиатором в ряде отделов головного и спинного мозга [5], повышает проницаемость клеток
для ионов калия, в частности – для скелетных мышц. Поэтому недостаток дикарбоновых аминокислот
может также сказываться как на проведении импульсов в мышечной ткани, так и на ее метаболизме,
вызывая нарушение согласованности функций опорно-двигательной системы.
Таким образом, одним из возможных начальных звеньев двигательной патологии при ДЦП могут
выступать нарушения глутаматергической системы и субординационных влияний, связанных с явле-
ниями нейротрансмиттерной дисрегуляции возбудимости сегментарного аппарата спинного мозга [11].
Нарушение обмена глутамата ведет к накоплению в митохондриях нейронов ионов кальция, что
способствует нарушению тканевого дыхания и процессов окислительного фосфорилирования [9].
Анализ соотношения тормозные/возбуждающие САК выявил значительные превышения (в 1,5 и
1,6 раза соответственно) по сравнению с группами контроля. Действительно, установлено, что на-
рушение уровня ГАМК и дофамина в ликворе больных с ДЦП коррелирует со степенью поражения
интегральных связей в ЦНС [14].
В последние годы в патогенезе заболеваний нервной системы большое значение придается арги-
нину как участнику цикла Кребса и как источнику окиси азота в организме. Действительно, концен-
трация аргинина в крови детей с ДЦП снижена (в 1,9 и 1,9 раза соответственно). Как избыток, так и
недостаток окиси азота, возникающие в силу разных причин и в первую очередь – из-за дефицита
аргинина, могут быть значимыми в процессе формирования и развития ЦНС [3].
Наконец, следует отметить выраженный дисбаланс серосодержащих САК в крови детей с ДЦП.
Так, если содержание цистеиновой кислоты (в 2,1 и 2,6 раза) и цистеина (в 2,2 и 3,5 раза) повышено,
то концентрация таурина (в 2,1 и 1,7 раза) и метионина (в 1,8 и 1,7 раза) наоборот снижена в обеих
возрастных группах. Ранее было показано [15], что содержание и соотношение серосодержащих
аминокислот играет важнейшую роль в поддержании функционального состояния всех основных
органов и систем организма. Нарушение обмена серосодержащих аминокислот приводит к многооб-
разным патологическим последствиям для организма (повреждениям стенок артерий, клеток голов-
ного мозга, структуры ДНК, нарушениям внутриутробного развития и т.д.). Это свидетельствует о
неспецифическом влиянии серосодержащих САК на функциональное состояние организма. Их коли-
чественные изменения во многом определяют уровень здоровья, в частности психического, и могут
быть использованы в качестве его маркеров. Подтверждается санокреатологический принцип необхо-
димости единого подхода к оценке морфофункционального состояния различных органов и систем.
Результаты определения содержания продуктов АО в утренней моче детей различного возраста с
ДЦП и здоровых детей отражены в таблице 2.
Таблица 2
Сравнительное содержание показателей АО в моче детей
контрольной группы и с ДЦП (мг/100мл)
Показатели азотистого обмена Контрольная
группа
Дети
до 2 лет
Дети
4 - 10 лет
Цистеиновая кислота 2,41±0,33 2,77±0,53* 4,10±0,86*
Таурин 8,63±0,89 1,03±0,19* 10,42±2,08*
Аспарагиновая кислота 1,95±0,16 1,92±0,40 1,73±0,19
Гидроксипролин 4,33±0,49 3,10±0,59* 3,00±0,69*
Треонин 1,09±0,14 1,35±0,30* 1,14±0,14
Серин 5,63±0,54 3,37±0,38* 4,50±0,81*
Аспарагин 1,01±0,11 0,91±0,21 2,69±0,43*
Глутаминовая кислота 0,93±0,09 0,43±0,09* 1,13±0,19*
Глутамин 8,25±1,19 9,82±1,77* 16,86±3,20*
α-аминоадипиновая кислота 0,43±0,06 0,45±0,09 0,20±0,04*

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
21
Пролин 2,30±0,29 1,70±0,32* 3,21±0,67*
Глицин 8,23±1,28 9,28±1,48* 12,23±1,96*
Аланин 4,55±0,50 3,73±0,63* 6,37±0,96
Цитруллин 0,30±0,03 0,41±0,08* 0,76±0,14*
α-аминомасляная кислота 0,28±0,03 0,30±0,06 0,23±0,04
Валин 1,24±0,14 0,41±0,09* 0,74±0,15*
Цистеин 3,07±0,48 2,05±1,27* 2,03±0,41*
Гомоцистеин 0,28±0,05 0,22±0,05* 0,49±0,10*
Метионин 0,41±0,05 0,59±0,12* 1,15±0,21*
Цистатионин 0,30±0,05 0,39±0,07* 0,29±0,04
Изолейцин 0,72±0,08 0,76±0,12 0,48±0,08*
Лейцин 1,10±0,20 0,28±0,04* 0,42±0,09*
Тирозин 1,10±0,15 2,06±0,37* 2,92±0,53*
Фенилаланин 0,54±,08 0,88±0,15* 1,73±0,35*
β- аланин 0,55±0,09 0,41±0,08* 2,14±0,28*
β-аминомасляная кислота 0,60±0,09 0,29±0,04* 0,71±0,15
γ-аминомасляная кислота 0,28±0,03 0,27±0,06 0,50±0,10*
Этаноламин 1,25±0,18 0,55±0,13* 0,53±0,09*
Триптофан 0,53±0,08 1,01±0,19* 1,69±0,35*
Орнитин 0,54±0,10 0,63±0,13* 0,74±0,18*
Лизин 1,85±0,26 2,82±0,51* 2,22±0,44*
Гистидин 5,21±1,12 12,58±1,89* 19,66±4,33*
1-метилгистидин 2,00±0,22 4,28±0,73* 4,29±0,73*
3-метилгистидин 1,50±0,23 2,44±0,39* 4,94±0,94*
Аргинин 1,48±0,20 1,12±0,23* 1,15±,023*
Мочевина 614,29±141,90 284,36±62,56* 524,65±120,67
Аммиак 24,82±3,58 9,71±1,85* 18,59±2,42*
САК 73,79±12,49 48,77±9,75* 77,93±17,14
Сумма показателей азотистого обмена 712,90±158,19 237,67±54,67* 463,31±106,56*
Заменимые аминокислоты 37,01±4,40 23,73±4,27* 35,71±5,41
Незаменимые аминокислоты 14,16±1,65 13,95±2,65 19,55±4,11
Иммуноактивные аминокислоты 20,26±2,65 10,84±1,95* 15,35±3,38*
Гликогенные аминокислоты 22,68±3,58 13,61±2,86* 17,42±3,14*
Кетогенные аминокислоты 5,83±0,82 5,12±1,02* 7,74±1,47*
Протеиногенные аминокислоты 51,17±5,34 37,69±8,29* 55,26±9,95
Серосодержащие аминокислоты 14,81±2,17 4,73±0,80* 12,25±2,57
* Р < 0,05
Сравнительный анализ содержания САК в моче у здоровых детей и с ДЦП также выявил значитель-
ные разнонаправленные изменения. Для детей до 2 лет с ДЦП характерна выраженная гипоаминоаци-
дурия (суммарное содержание САК снижено в 1,5 раз. При этом наиболее значительно снижено со-
держание серосодержащих, иммуноактивных и гликогенных САК (в 3,1, 1,9 и 1,7 раз соответственно).
У детей старшей группы имеет место лишь незначительное снижение содержания иммуноактивных
и гликогенных САК в 1,3 раза. Это может свидетельствовать о возрастном развитии адаптивных
механизмов. Содержание конечных продуктов АО в моче детей обеих возрастных групп снижено:
аммиака – в 2,6 и 1,3 раза соответственно, мочевины – в 2,0 и 1,2 раза соответственно.
У детей до 2 лет достоверно и значительно снижено содержание таурина в 8,3 раза, лейцина в 3,9
раза, этаноламина в 2,3 раза, валина в 3,0 раза, оксипролина в 1,4 раза, серина в 1,7 раза, глутамата в
2,2 раза, пролина в 1,4 раза, аланина в 1,2 раза, аргинина в 1,3 раза. У детей второй возрастной группы

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
22
с ДЦП снижено содержание только лейцина в 2,6 раза, цистеина в 1,5 раза, валина в 1,7 раза, изолей-
цина в 1,5 раза, этаноламина в 2,4 раза.
Ряд концентраций САК в моче детей с ДЦП повышен. Так, у детей до 2 лет достоверно повышен
уровень цитруллина в 1,4 раза, метионина в 1,4 раза, цистатионина 1,3 раза, тирозина в 1,9 раза, фенил-
аланина в 1,6 раза, триптофана в 1,9 раза, лизина в 1,5 раза, гистидина в 2,4 раза, 1-метилгистидина в
2,1 раза, 3-метилгистидина в 1,6 раза. У детей старшей группы достоверно повышен уровень аспара-
гина в 2,7 раза, глутамина в 2,0 раза, пролина в 1,4 раза, глицина в 1,5 раза, аланина в 1,4 раза, ци-
труллина в 2,5 раза, гомоцистеина в 1,8 раза, метионина в 2,8 раза, тирозина в 2,7 раза, фенилаланина
в 3,2 раза, триптофана в 3,2 раза, орнитина в 1,4 раза, гистидина в 3,8 раза, 1-метилгистидина в 2,1 раза,
3-метилгистидина в 3,3 раза.
Далее нам представилось важным соотнести данные о концентрациях САК в сыворотке крови и в
моче детей с ДЦП и здоровых детей тех же возрастов.
Из данных, приведенных в таблицах 1 и 2, можно сделать вывод, что для детей с ДЦП характерен
недостаток как в сыворотке крови, так и в моче иммуноактивных, гликогенных и серосодержащих
аминокислот. Это свидетельствует о недостаточном их поступлении в кровь или о нарушениях всасы-
вания в ЖКТ при ДЦП.
У детей с ДЦП на фоне сниженного содержания изолейцина, тирозина, фенилаланина, трипто-
фана, орнитина, лизина, гистидина в плазме крови наблюдается повышенное их выведение с мочой.
Отдельно следует отметить значительный уровень аммиака в сыворотки крови детей с ДЦП на
фоне сниженной его концентрации в моче.
Наконец, в организме таких детей имеет место дисбаланс концентраций серосодержащих САК как
в крови, так и в моче. Необходимо отметить накопление гомоцистеина и в крови и в моче у детей стар-
шей группы, что может свидетельствовать об усугублении с возрастом нарушения обмена серосодер-
жащих аминокислот. Гомоцистеин, как известно [15], является предиктором развития нарушений
функциональной активности сердечно-сосудистой системы.
Как повышенное, так и сниженное выведение продуктов азотистого обмена из организма может
свидетельствовать об изменениях функциональной активности почек.
Таким образом, при ДЦП имеют место значительные изменения азотистого обмена на фоне нару-
шений ряда функциональных систем организма, что обусловливает развитие психической дизонтоге-
нии. Приведенные выше данные подтверждают санокреатологическую концепцию, согласно которой
нарушение психического здоровья, как и саногенное его становление, будучи интегративным процес-
сом высшей нервной деятельности, предопределяется программой развития организма, реализуемой
в онтогенезе за счет генетически детерминированных нейрофизиологической интегральной и нейро-
химической систем мозга, а также психофункциональных и оценочно-исполнительных систем, фор-
мирующихся за счет действия психологических факторов внешней среды [12, 13].
Выводы
1. Нарушение метаболизма свободных аминокислот позволяет выявить формирование психической
дизонтогении на модели ДЦП и расширяет представление о биохимическом компоненте его патогенеза.
2. Значительное увеличение коэффициента соотношения тормозные/возбуждающие САК у детей
с ДЦП практически не зависит от возраста и свидетельствует о наличии у них стойкого торможения в
ЦНС, вызывающего отставание психического развития.
3. Для детей с ДЦП характерны выраженная гипоаминоацидемия и снижение показателей азотис-
того обмена в моче, при этом наиболее значительно снижена концентрация серосодержащих, иммуно-
активных и гликогенных САК.
4. ДЦП сопровождается выраженным дисбалансом серосодержащих САК, что позволяет исполь-
зовать их в качестве неспецифического маркера уровня здоровья.
Библиография:
1. ЛЕВЧЕНКО, И.Ю. Психологические особенности подростков с детским церебральным параличом. Москва:
Альфа, 2001. 160 с.
2. МАМАЙЧУК, И.И. Нейропсихологические исследования гностических процессов у детей с различными
формами детского церебрального паралича. В: Журнал невропатологии и психиатрии, 1992, №4, с.49-56.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
23
3. МАРКОВ, Х.М. Роль оксида азота в патогенезе болезней детского возраста. В: Российский вестник перина-
тологии и педиатрии, 2000, №4, с.43-47.
4. МАСТЮКОВА, Е.М. Дети с церебральным параличом. В: Специальная психология, 2003, №2, с.89-92.
5. РАЕВСКИЙ, К.С. Возбуждающие аминокислоты, глутаматные рецепторы и патология ЦНС. В: Патологи-
ческая физиология и экспериментальная терапия, 1990, № 1, с.3-9.
6. РЕПИНА, Н.В. Основы клинической психологии: Учебное пособие. Ростов-на-Дону: Феникс, 2003. 480 с.
7. СКОВОРЦОВ, И.А. Неврология развития: руководство для врачей. Москва: Лит-терра. 2008. 544 с.
8. СОЛОДКОВ, А.С. Физиология человека: Учебное пособие. Москва: Терра-спорт. 2001. 520 с.
9. СТОРОЖУК, В.М. Участие глутаматных внутрикорковых связей в условнорефлекторной деятельности. В:
Нейрофизиология, 2000, т.24, № 6, с.701-712.
10. СУРКИНА, И.Д. Некоторые аспекты аминокислотного обмена при нервных и психических заболеваниях
(метаболизм триптофана). В: Журнал невропатологии и психиатрии, 1995, № 8, с.65-69.
11. УАКАСОВА, Г.М. Возможности метаболической терапии. Молекулярные основы патогенеза заболеваний
нервной системы. В: Вестник КАСУ, 2007, №3, с.187-190.
12. ФУРДУЙ, Ф.И., ЧОКИНЭ, В.К., ФУРДУЙ, В.Ф. и др. Психическое здоровье с позиций санокреатологии и
системогенеза. В: Известия Академии наук Молдовы. Науки о жизни, 2012, №2(317), с.4-11.
13. ФУРДУЙ, Ф.И., ЧОКИНЭ, В.К. Психосанокреатологическая концепция. Генетически и онтогенетически
обусловленные системы, участвующие в формировании и поддержании психического здоровья. В: Известия
Академии наук Молдовы. Науки о жизни, 2012, №3(318), с.4-23.
14. ФУРДУЙ, Ф.И. Санокреатология – новая отрасль биомедицины, призванная приостановить биологическую
деградацию человека. В: Стресс, адаптация, функциональные нарушения и санокреатология, Кишинев,
1999, с.36-42.
15. ФУРДУЙ, Ф.И., ЧОКИНЭ, В.К., ПАВАЛЮК, П.П. и др. Научные основы создания физиологически обосно-
ванного образа жизни. В: The Bulletin of the European Postgraduate Centre of Acupuncture and Homeopathy,
Chisinau, 2000, №4, р.26-40.
16. ФУРДУЙ, Ф.И. Проблемы стресса и преждевременной биологической деградации человека. Санокреато-
логия, еѐ настоящее и будущее. В: Современные проблемы физиологии и санокреатологии, Кишинев, 2005,
с.16-36.
17. ЦЫМБАЛЮК, В.И., ВАСИЛЬЕВА, И.Г., ПИЧКУР, Л.Д. и др. Содержание нейромедиаторов в субарахнои-
дальном ликворе у больных детским церебральным параличом и влияние на него трансплантации ЭНТ. В:
Українский нейрохiрургiчний журнал, 2001, №1, с.51-54.
18. ЧОКИНЭ, В.К., ГАРАЕВА, С.Н., НЕВОЯ, А.В. и др. Серосодержащие аминокислоты в диагностике, целе-
направленном поддержании и формировании здоровья. В: Известия Академии наук Молдовы. Науки о жизни,
2011, №3(315), с.15-35.
19. ШИПИЦЫНА, Л.М. Психология детей с нарушениями опорно-двигательного аппарата. Москва: Владос,
2004. 368 с.
20. BALAZS, R. Excitatory amino acid transmission in Health and Disease. Binding, Hardback Book Condition. 2005.
369 p.
Работа выполнена в рамках Институционально проекта № 11.817.09.01.F. „Elaborarea bazei ştiinţifice
a sănătăţii psihice şi identificarea nivelelor de expresie a ei”.
Prezentat la 12.09.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
24
INFLUENŢA CITRATULUI DE FIER ŞI A INTENSITĂŢII LUMINII
ASUPRA PRODUCTIVITĂŢII SPIRULINEI ŞI CONŢINUTULUI
DE CAROTENOIZI ŞI FIER ÎN BIOMASĂ
Valentina BULIMAGA, Liliana ZOSIM, Maria PISOV, Olga OLAN
Universitatea de Stat din Moldova
A fost studiată influenţa citratului de fier şi a intensităţii luminii asupra productivităţii spirulinei şi conţinutului de carotenoizi şi fier în biomasă. S-a stabilit că aplicarea combinată a unor astfel de factori, cum sunt intensitatea luminii şi suplimentarea citratului de fier la mediul nutritiv, condiţionează majorarea conţinutului de carotenoizi şi fier în biomasa de spirulină la cultivare în decurs de 10 zile.
Cuvinte-cheie: Spirulina platensis, intensitatea luminii, termen de cultivare, productivitate, carotenoizi, citrat de fier.
THE INFLUENCE OF IRON CITRATE AND LIGHT REGIM ON SPIRULINA PRODUCTIVITY
AND CAROTENOIDS AND IRON CONTENT IN BIOMASS The influence of iron citrate and light intensity on the spirulina productivity, carotenoids and iron content in
biomass have been studied. It was established that the combined application of such factors as light intensity and iron citrate administration
conducted to the increasing of carotenoids and iron content in spirulina biomass at cultivation during 10 days. Keywords: Spirulina platensis, light intensity, period of cultivation, productivity, carotenoids, iron citrate.
Introducere Majoritatea cercetărilor efectuate pe parcursul ultimelor decenii asupra algelor şi cianobacteriilor, în special
asupra spirulinei, sunt îndeosebi cercetări ficobitehnologice ce vizează utilizarea ei în calitate de supliment alimentar şi pentru obţinerea unor preparate polivalente cu efecte curative înalte.
Cultivarea spirulinei în condiţii controlate asigură obţinerea biomasei ecologic pure care poate servi ca sursă eficientă nu doar a unui şir de substanţe bioactive preţioase, dar şi a unor microelemente esenţiale, precum zincul, seleniul, cuprul şi, în special, fierul. Dezvoltarea în continuare a aspectelor fundamentale şi aplicative ale acestor cercetări îşi păstrează actualitatea, fiind nemijlocit legate de perfecţionarea procedeelor de cultivare a spirulinei, plasând în prim-plan elaborarea unor modele noi de dirijare eficientă a fotobiosintezei în direcţia sporirii producerii de biomasă şi obţinerii principiilor bioactive cu calităţi pronosticate [1,2,8,10,11].
Carotenoizii reprezintă un grup de substanţe bioactive cu proprietăţi antioxidante pronunţate. Datorită pre-zenţei legăturilor duble conjugate, carotenoizii leagă oxigenul singlet şi inhibă formarea radicalilor liberi. Joacă un rol important în combaterea aterosclerozei, împiedică oxidarea lipoproteinelor de mică densitate (coles-terolul LDL), reducând riscul declanşării şi producerii infarctului miocardic, posedă proprietăţi metabolice, imunostimulatoare şi antioxidante, ceea ce denotă perspectiva utilizării lor în profilaxia şi tratamentul complex al cancerului, maladiilor cardiovasculare şi infecţioase [3,4].
Fierul este unul dintre bioelementele esenţiale ale multor procese biochimice şi sisteme enzimatice. Fiind con-vertit în compuşi bioorganici, fierul se poate include mai eficient în metabolismul de biosinteză celulară, în mă-sura necesităţilor fiziologo-metabolice ale organismului. Deficienţa nutriţională şi carenţa fierului în organismul uman are drept consecinţă anemia fieriprivă – una dintre cele mai stringente probleme ale medicinii contem-porane, nesoluţionate la momentul actual, dar şi apariţia defectelor în răspunsul imun al organismului [6].
Având în vedere importanţa obţinerii unor produse bogate în principii bioactive cu efect antioxidant şi anti-anemic, prezintă interes studiul în complex al influenţei unor parametri de cultivare a spirulinei (intensitatea luminii), durata cultivării şi a sursei de fier, suplimentate la mediul nutritiv în diverse concentraţii asupra pro-ductivităţii şi acumulării carotenoizilor şi fierului la Spirulina platensis.
Scopul lucrării a constat în stabilirea condiţiilor optime de obţinere a unei cantităţi înalte de biomasă de spirulină îmbogăţite cu carotenoizi şi fier la cultivarea spirulinei în prezenţa citratului de fier.
Material şi metode Obiectul de studiu – tulpina cianobacteriei Spirulina platensis CNM-CB-11- (CYANOPHYTA) depozitate
în Colecţia Naţională de Microorganisme Nepatogene al IMB al AŞM. Pentru cultivarea spirulinei a fost uti-

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
25
lizat mediul nutritiv mineral SP-1 cu o compoziţie echilibrată a macro- şi micronutrienţilor necesari creşterii şi dezvoltării spirulinei. Cultivarea s-a efectuat în baloane Erlenmeyer a câte 100 ml cu 50 ml suspensie de spirulină, timp de 144 ore pentru prima variantă şi de 216 ore pentru varianta a doua, la iluminarea 3500 lx şi 4500 lx.
Productivitatea spirulinei a fost determinată conform metodei descrise în [7,9]. Determinarea fierului în biomasă a fost efectuată prin metoda colorimetrică bazată pe reacţia Fe(III) cu
rodanura de potasiu [14]. Concentraţia pigmenţilor în soluţii alcoolice a fost determinată prin metoda spectrometrică propusă de
H.Lichtenthaler şi C.Buschmann [5]. Analiza statistică a valorilor obţinute în trei serii de determinări ale productivităţii, conţinutului de carote-
noizi, clorofila a şi fier a fost realizată prin metodele propuse de Maximov şi Dospehov [12,13].
Rezultate şi discuţii În mod standard spirulina în proporţii industriale este cultivată pe mediul Zarouk sau cu unele modificări
ale lui cu o componenţă echilibrată de macro- şi microelemente. Administrarea în mediul de cultivare a can-tităţilor excesive sau insuficiente de elemente chimice, precum şi expunerea culturii la acţiunea unor factori fizici poate provoca dezechilibru în sistemul fotosintetic, diviziunea celulelor, dereglarea proceselor de creş-tere. Din cele menţionate rezultă importanţa selectării condiţiilor de cultivare, a compuşilor şi a concentraţii-lor utilizate în supliment la mediul de cultivare în aşa mod, încât sa nu fie perturbate procesele vitale.
Un parametru esenţial la cultivarea spirulinei în proporţii industriale prezintă productivitatea, ai cărei indici caracterizează în linii generale starea culturii. Rezultatele cercetărilor au demonstrat că la cultivare în regim de iluminare 3500 lx, indiferent de termenul de cultivare, suplimentarea citratului de fier are acţiune pozitivă asupra productivităţii (Fig.1).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0
15
30
45
60
75 0
15
30
45
60
75 0
15
30
45
60
75 0
15
30
45
60
75
iluminare 3500 lx
cultivare 7 zile
iluminare 3500 lx
cultivare 10 zile
iluminare 4500 lx
cultivare 7 zile
iluminare 4500 lx
cultivare 10 zile
Pro
du
cti
vit
ate
, g
/l
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Fe %
BA
U
Productivitatea, g/l Fe, % BAU
Fig.1. Influenţa citratului de fier şi a intensităţii luminii asupra productivităţii şi conţinutului de fier
la cultivarea spirulinei în decurs de 7 şi 10 zile.
La spirulina cultivată 7 şi 10 zile la iluminarea 4500 lx, prezenţa citratului influenţează nesemnificativ asupra productivităţii, manifestându-se o tendinţă de diminuare a ei cu creşterea concentraţiei citratului de fier. La ambele regimuri de iluminare productivitatea spirulinei cultivate în decurs de 10 zile este mai înaltă cu 21-26% faţă de cea cultivată timp de 7 zile, fără suplimentarea citratului de fier şi cu 14-25% în cazul suplimentării cu citrat. Aşadar, valori maxime ale productivităţii spirulinei – 1,79 g/l au fost atestate la spiru-lina cultivată 10 zile în regim de iluminare 4500 lx cu suplimentarea a 15 mg/l citrat de fier (Fig.1).

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
26
În ce priveşte acumularea fierului în biomasa de spirulină, se poate afirma că conţinutul de fier variază în
dependenţă de termenul de cultivare, regimul de iluminare şi concentraţia citratului de fier administrat în
mediul de cultivare. Conţinutul de fier acumulat în biomasă se majorează concomitent cu creşterea concen-
traţiei citratului de fier administrat în mediul de cultivare, pentru toate cazurile studiate (Fig.1). Totuşi, valori
cantitative mai înalte au fost semnalate în variantele experimentale obţinute la cultivare timp de 10 zile.
Astfel, conţinutul maxim de fier se acumulează în biomasa cultivată în decurs de 10 zile cu suplimentarea
mediului de cultivare cu concentraţie maximă de citrat de fier (75 mg/l), valorile cantitative întrecând de 8,9 ori
valoarea martorului, ceea ce constituie 634 mg% din biomasă. Asupra conţinutului de fier influenţează şi re-
gimul de iluminare, astfel încât la administrarea compusului în concentraţie maximă (75 mg/l) în regim de
iluminare de 4500 lx conţinutul de fier este de 2,51 ori mai înalt decât în proba obţinută la cultivare în regim
de iluminare de 3500 lx.
Rezultatele cercetărilor influenţei citratului de fier şi a intensităţii luminii asupra conţinutului de carotenoizi
şi clorofilă în biomasa de spirulină, cultivată în decurs de 7 şi 10 zile, demonstrează că un rol important, ca şi
în cazul conţinutului de fier, revine atât intensităţii luminii, cât şi duratei de cultivare (Fig.2).
0
5
10
15
20
25
30
0 m
g/l
15 m
g/l
30 m
g/l
45 m
g/l
60 m
g/l
75 m
g/l
0 m
g/l
15 m
g/l
30 m
g/l
45 m
g/l
60 m
g/l
75 m
g/l
0 m
g/l
15 m
g/l
30 m
g/l
45 m
g/l
60 m
g/l
75 m
g/l
0 m
g/l
15 m
g/l
30 m
g/l
45 m
g/l
60 m
g/l
75 m
g/l
iluminare 3500 lx
cultivare 7 zile
iluminare 3500 lx
cultivare 10 zile
iluminare 4500 lx
cultivare 7 zile
iluminare 4500 lx
cultivare 10 zile
Clo
rofi
la a
, m
g/g
0
1
2
3
4
5
6
7
C
aro
ten
oiz
i, m
g/g
Clorofila a mg/g Carotenoizi mg/g
Fig.2. Influenţa citratului de fier şi a intensităţii luminii asupra conţinutului de carotenoizi
şi de clorofilă a la cultivarea spirulinei în decurs de 7 şi 10 zile.
În prezenţa citratului de fier în concentraţii de 15-75 mg/l şi la intensitatea luminii 4500 lx, conţinutul de
carotenoizi sporeşte cu 7-37% faţă de proba-martor, obţinută la cultivare în regim de iluminare de 3500 lx, fără
suplimentarea citratului de fier. În biomasa cultivată timp de 10 zile fără adaos de citrat de fier la 4500 lx se
atestă un spor nesemnificativ al conţinutului de carotenoizi (cu 9%) faţă de biomasa cultivată la 3500 lx.
Un spor maxim al conţinutului de carotenoizi (cu 63%) a fost înregistrat în biomasa de spirulină cultivată
în decurs de 10 zile în regim de iluminare intensă (4500 lx) în prezenţa citratului de fier în concentraţie optimă
de 30 mg/l, comparativ cu proba cultivată în decurs de 7 zile fără administrarea citratului de fier. Rezultatele
cercetărilor efectuate au scos în evidenţă că aplicarea combinată a unor astfel de factori, cum este intensi-
tatea luminii şi citratul de fier, condiţionează majorarea conţinutului de carotenoizi şi de fier în biomasa de
spirulină la cultivare în decurs de 10 zile. A fost stabilit că concentraţia optimă a citratului de fier pentru
acumularea maximă a fierului este de 75 mg/l, iar pentru un conţinut maxim de carotenoizi – de 30 mg/l.
Aşadar, pentru obţinerea biomasei de spirulină îmbogăţite cu carotenoizi şi fier poate fi utilizat mediul
de cultivare SP-1, cu suplimentarea la cultura de spirulină a citratului de fier în concentraţii de 30-75 mg/l
şi efectuarea cultivării în decurs de 10 zile, la intensitatea luminii 4500 lx.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
27
Concluzii
1. La cultivarea spirulinei în regim de iluminare 3500 lx, indiferent de termenul de cultivare (7 sau 10 zile),
suplimentarea citratului de fier în concentraţii de 15-75 mg/l manifestă acţiune stimulatoare asupra pro-
ductivităţii. Valori maxime ale productivităţii spirulinei (1,79 g/l) au fost atestate la spirulina cultivată pe
mediu cu adaos de citrat de fier (15 mg/l), timp de 10 zile, în regim de iluminare 4500 lx.
2. Conţinutul de fier în biomasă creşte cu majorarea concentraţiei şi asupra acumulării lui influenţează atât
regimul de iluminare, cât şi durata de cultivare, valorile lui cantitative fiind de 2,51 ori mai înalte la 4500 lx,
comparativ cu proba obţinută la cultivare timp de 7 zile în regim de iluminare 3500 lx. Valori cantitative
maxime (0,634% Fe din biomasă) au fost înregistrate în variantele experimentale obţinute la cultivare în
decurs de 10 zile, la iluminarea 4500 lx, în prezenţa citratului de fier (75 mg/l).
3. Un spor maxim al conţinutului de carotenoizi (cu 63%) a fost înregistrat în biomasa de spirulină cultivată
în decurs de 10 zile în regim de iluminare intensă (4500 lx) şi în prezenţa citratului de fier în concentraţie
optimă de 30 mg/l.
Bibliografie:
1. CHIRIAC, T. Biotehnologia cultivării spirulinei şi obţinerii produselor cu conţinut prognozat de zinc şi principii
bioactive valoroase: Autoref. tezei de dr. în şt. biologice. Chişinău, 2003. 23 p.
2. CIUMAC, D. Studiul modificării componenţei biochimice a cianobacteriei Spirulina platensis la cultivarea în
prezenţa compuşilor coordinativi ai Cr(III): Autoref. tezei de dr. şt. biologice. Chişinău, 2008. 24 p.
3. El-BAKY, H., El- BAZ, F., El-BAROTY, G. Production of carotenoids from marine algae and its evaluation as self
food colorants and lowering cholesterol agents. In: American –Euroasian J. Agr. And Food Sci., 2007, vol.2, no.6,
p.792-800. ISSN 1818-6769
4. JASWIR, D., NOVIENDRI, R., HASRINI, F. Carotenoids: Sources, medicinal properties and their application in food
and nutraceutical industry. In: Journal of Medicinal Plants Research, 2011, vol.33, p.7119-7131. ISSN 1996-0875
5. LIHTENTHALER, H., BUSCHMANN, C. Chlorophylls and carotenoids: Measurement and characterization by UV-
VIS spectroscopy. In: Current Protocols in Food Analytical Chemistry. 2001. F4.3.1-F4.3.8. ISBN 978-0471142911
6. PUYFOULHOUX, G., ROUANET, J., BESANÇON, P., BAROUX, B., BACCOU, J., CAPORICCIO, B. Iron
Availability from Iron-Fortified Spirulina by an in Vitro Digestion Caco-2 Cell Culture Model. In: J. Agric. Food
Chem., 2001, vol.49, no.3, p.1625–1629.
7. RUDIC, V. Aspecte noi ale biotehnologiei moderne. Chişinău: Ştiinţa, 1993. 139 p. ISBN 978-5376018293
8. RUDIC, V. Ficobiotehnologie – cercetări fundamentale şi realizări practice. Chişinău: Elena V.I., 2007. 364 p.
ISBN 978-9975989251
9. RUDIC, V. ş. a. Metode de investigaţie în ficobiotehnologie. Chişinău: CE USM, 2002. 61 p. ISBN 9975-70-254-6
10. ZOSIM, L. Biotehnologia cultivării spirulinei şi obţinerii produselor cu conţinut prognozat de fier şi alte substanţe bioactive
valoroase: Autoref. tezei de dr. şt. biologice. Chişinău, 2007. 22 p.
11. АЛЕШКО-ОЖЕВСКИЙ, Ю., ЗИЛОВА, И., МАЗО, В. Spirulina platensis – перспективный пищевой источник
эссенциальных микроэлементов. B: Вестник новых медицинских технологий, 2002, том. 9, №.1, с.3-10.
12. ДОСПЕХОВ, Б. Методика полевого опыта. Москва: Агропромиздат, 1985. 351 с.
13. МАКСИМОВ, В.Н. Многофакторный эксперимент в биологии. Москва: Изд-во МГУ, 1980. 278 с.
14. ФИЛИППОВИЧ, Ю., ЕГОРОВА, Т., СЕВАСТЬЯНОВА, Г. Практикум по общей биохимии. Москва: Просвещение,
1982, с.284-286.
Notă: Cercetările au fost întreprinse în cadrul Proiectului Instituţional 11.817.08.44A finanţat de către Consiliul
Suprem pentru Ştiinţă şi Dezvoltare Tehnologică al AŞM.
Prezentat la 15.09.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
28
CULTIVAREA ALGELOR CHLORELLA VULGARIS BEIJER, SYNECHOCYSTIS SALINA
WISL., PHORMIDIUM FOVEOLARUM (MONT.) GOM. ŞI TRIBONEMA VIRIDE PASCH.
PE MEDII CU ADAOS DE APE REZIDUALE ORĂŞENEŞTI
Natalia DONŢU
Universitatea de Stat din Moldova
Pentru cultivarea speciilor de alge (Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum şi Tribonema
viride) au fost utilizate mediile cu adaos de ape reziduale de la Staţia de Epurare Biologică, în concentraţii de la 1 la
50%. Pentru fiecare specie au fost stabilite concentraţiile optimale ale mediilor de cultivare. Cea mai înaltă cantitate de
biomasă a fost obţinută după 10-15 zile de cultivare. Pentru specia Chlorella vulgaris cel mai favorabil s-a dovedit a fi
mediul cu adaos de 5% (17,55 g/l), pentru Synechocystis salina mediul cu adaos de 20% (16,56 g/l), pentru Phormidium
foveolarum – mediul cu adaos de 10% (3,36 g/l), iar pentru Tribonema viride – mediul cu adaos de 30% (2,10g/l).
Cuvinte-cheie: alge, ape reziduale, mediu de cultivare, mediu optimal, biomasă.
CULTIVATION OF THE ALGAE CHLORELLA VULGARIS BEIJER, SYNECHOCYSTIS SALINA WISL.,
PHORMIDIUM FOVEOLARUM (MONT.) GOM. AND TRIBONEMA VIRIDE PASCH IN MEDIA WITH
ADDED MUNICIPAL WASTEWATERS
For the cultivation of algae species (Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum and Tribonema viride) were used media with added of wastewater from Biological Wastewater Treatment Plant in the range of 1 to 50%. For each species were set to its optimum cultivation media. The best results were obtained a fter 10-15 days of cultivation. The highest amount of biomass of the species Chlorella vulgaris was at the 5% medium (17.545 g/l), for the species Synechocystis salina peak was on average with 20% (16.556 g/l), Phormidium foveolarum – medium with added 10% (3.36 g/l), and for the species Tribonema viride – the 30% medium (2.1 g/l). Common to all species was the medium with added to 10% of the waste water, but the obtained results vary from case to case.
Keywords: algae, wastewaters, cultivation medium, biomass, optimal medium.
Introducere Apele reziduale se caracterizează printr-un conţinut înalt de elemente biogene, care pot contribui la mări-
rea productivităţii algelor, considerent din care ele pot servi în calitate de medii de cultivare [7,9]. Efluenţii de apă provenite de la staţiile de epurare conţin o cantitate mare de nutrienţi (azot, fosfor ş.a.) şi deversarea lor directă în bazinele acvatice poate duce la eutrofizarea acestora [6,8]. Pentru diminuarea nivelului de eutro-fizare se poate implementa tratarea „terţiară” cu ajutorul algelor [1,4]. Biomasa algală crescută pe ape rezi-duale poate să conţină cantităţi suficiente de principii biologic active şi chiar să depăşească uneori cantitatea lor din unele plante de cultură [2,5,13].
Speciile de alge se dezvoltă pe diferite concentraţii de ape reziduale. De exemplu, speciile cianofite din genurile Phormidium, Oscillatoria, Synechocystis, Nostoc, Spirulina ş.a. se dezvoltă în mediile cu adaos de ape reziduale cu concentraţiile de la 5 până la 25% [2]. Clorofitele sunt mai tolerante la ape reziduale, spre deosebire de cianofite. Unele specii din genurile Chlorella, Scenedesmus, Dunaliella, Kirchneriella etc. pre-feră mediile cu adaos de ape reziduale de la 1 până la 40% [3,11]. Algele macrofite, conform datelor de ultimă oră din literatură, se dezvoltă foarte bine în mediile cu adaos de 10% şi 25% ape reziduale, iar limita de tole-ranţă faţă de adaos de ape reziduale este de 50%. Odată cu creşterea concentraţiei de ape reziduale este inhi-bată dezvoltarea algelor şi se reduce la minimum capacitatea de asimilare a azotului şi fosforului din apă [10].
Scopul cercetărilor noastre a fost de a stabili concentraţiile de ape reziduale optime pentru dezvoltarea speciilor Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum şi Tribonema viride şi de a stabili cantitatea maximală de biomasa acumulată de fiecare specie în parte.
Material şi metode
Cercetările au fost efectuate în LCŞ „Algologie” al USM. Ca obiect de cercetare au servit speciile de alge
Synechocystis salina, Chlorella vulgaris, Phormidium foveolarum şi Tribonema viride. Primele trei specii se
întâlnesc predominant în bazinele Staţiei de Epurare Biologică a mun. Chişinău, cu excepţia celei din urmă,
care preferă bazinele cu apă mai puţin poluată din treptele finale de epurare. Speciile de alge au fost selectate

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
29
în culturi algologic pure şi cultivate în condiţii de laborator pe medii minerale, care prezentau probele-martor,
şi pe mediile cu adaos de ape reziduale de diferit procentaj. Apa reziduală, utilizată în experimentele pro-
puse, a fost prelevată din colectorul de la ieşirea din Staţia de Epurare Biologică. Rezultatele obţinute au fost
prelucrate statistic [14].
Rezultate şi discuţii
Unul dintre scopurile propuse în experienţele de cultivare a fost stabilirea concentraţiilor optimale de ape
reziduale orăşeneşti. Pentru fiecare specie apa reziduală utilizată la cultivare a fost prelevată separat şi în zile
diferite, de aceea există mici diferenţe în compoziţia chimică. Cantitatea ionului NH4+ variază de la 52,3 până
la 79,7 mg/l în proba de 100%, iar cantitatea ionului nitrat oscilează între 0,5 şi 0,66 mg/l. Concentraţia
ionului de nitrit în toate seriile de experimente este cuprinsă între 0,039 şi 0,11 mg/l. Concentraţia ionului de
fosfat a avut valori cuprinse între 0,12 şi 0,83 mg/l.
Culturile algale de Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum şi Tribonema viride
au fost crescute iniţial timp de 10 zile pe medii cu adaos de 1, 5, 10, 15, 20, 25, 30, 40, 50, 75 şi 100% ape
reziduale colectate de la ieşirea din Staţia de Epurare Biologică. În variantele cu adaos de 75 şi 100% ape
reziduale nu a supravieţuit nicio specie inoculată, cauza fiind concentraţia înaltă de substanţe chimice. În
fiecare caz concret ce ţine de specie au fost alese câte trei tipuri de medii cu adaos de ape reziduale, pe care
fiecare din culturile de alge a obţinut valori mari ale cantităţii de biomasă.
Din datele prezentate în Figura 1 putem constata că pentru cultura de Chlorella vulgaris cele mai optimale
medii de creştere sunt cele cu adaos de 1, 5 şi 10%. Microalga cianofită Synechocystis salina, spre deosebire
de chlorelă, preferă mediile cu concentraţii mai mari de substanţe chimice, şi anume: acelea cu 10, 15 şi 20%
ape reziduale orăşeneşti. Deşi este o algă filamentoasă, pentru creşterea şi dezvoltarea speciei Phormidium
foveolarum pot fi utilizate aceleaşi medii ca şi pentru cultura de Synechocystis salina. Tribonema viride este
o specie de talie mai mare şi, probabil, de aceea se dezvoltă bine şi pe concentraţii mai mari de ape reziduale.
Cele mai bune rezultate au fost stabilite în variantele cu adaos de 10, 30 şi 50% ape reziduale orăşeneşti, unde
cantitatea de biomasă a variat de la 1,70 până la 2,00 g/l.
Fig.1. Cantitatea de biomasă (g/l) de Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum şi
Tribonema viride crescute pe medii cu adaos de ape reziduale (pentru speciile Chlorella vulgaris şi
Synechocystis salina rezultatele sunt micşorate de 10 ori).
Un alt scop al cercetărilor noastre a fost de a evidenţia mediul optimal pe care aceste culturi de alge pot
produce cea mai mare cantitate de biomasă algală şi perioada optimă de cultivare. Pentru realizarea lui am
efectuat experienţe de cultivare timp de 20 zile cu tulpinile de alge sus-menţionate.
Pentru specia Chlorella vulgaris toate trei medii selectate au fost favorabile pentru dezvoltare. Însă, din
datele prezentate în Figura 2, observăm că cea mai mare valoare a cantităţii de biomasă algală a fost stabilită
în mediul cu adaos de 5% ape reziduale (17,55 ± 1,72 g/l) în a 15-a zi de cultivare. Această cantitate de bio-
masă obţinută este de 1,63 ori mai mare decât în proba-martor (mediul mineral Tamia).
În variantele cu 1 şi 10% adaos de ape reziduale cantitatea de biomasă a crescut treptat până în a 15-a zi
de cultivare şi depăşeşte cantitatea de biomasă obţinută în urma cultivării pe mediul Tamia.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
30
Fig.2. Cantitatea de biomasă (g/l) de Chlorella vulgaris crescută pe medii cu adaos de ape reziduale orăşeneşti.
La cultivarea algei Synechocystis salina cu biomasa iniţială de 2,86 ± 0,66 g/l, cea mai înaltă producti-
vitate a fost obţinută, ca şi în cazul precedent, la a 15-a zi, doar că în mediile cu concentraţia de 20% ape
reziduale – 16,56 ± 3,70 g/l (Fig.3). Cele mai mici valori au fost constatate la cea de a 5-a zi de cultivare în
varianta cu adaos de 15% ape reziduale – 4,76 ± 0,68 g/l biomasă algală.
Fig.3. Cantitatea de biomasă (g/l) de Synechocystis salina crescută pe medii cu adaos de ape reziduale orăşeneşti.
În proba-martor pe mediul Bristol cantităţile de biomasă au fost mai mici, spre deosebire de probele cu adaos
de ape reziduale. Cantitatea maximă de biomasă este acumulată la a 15-a zi, fiind estimată la 6,68 ± 2,52 g/l.
Specia Phormidium foveolarum a fost cultivată pe medii asemănătoare cu cele pe care a fost cultivată
specia Synechocystis salina, însă rezultatele obţinute diferă. Datele prezentate în Figura 4 atestă o oscilare a
cantităţii de biomasă algală acumulate în urma cultivării speciei Phormidium foveolarum pe aceste tipuri de
medii nutritive cu adaos de ape reziduale. Cele mai mari cantităţi de biomasă algală au fost obţinute tot la a
15-a zi de cultivare, însă după 20 de zile cantitatea de biomasă scade. Cantitatea maximală de biomasă a fost
stabilită în mediul nutritiv cu adaos de 10% ape reziduale – 3,36 ± 0,035 g/l, ceea ce este cu 40% mai mult faţă
de cantitatea maximală obţinută pe mediul nutritiv chimic Gromov 6 (2,00 ± 0,048 g/l). Cele mai mici cantităţi
de biomasă au fost stabilite în primele zile de cultivare, datorită perioadei de adaptare la aceste condiţii.
Fig.4. Cantitatea de biomasă (g/l) de Phormidium foveolarum crescută
pe medii cu adaos de ape reziduale orăşeneşti.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
31
În variantele cu adaos de 15 şi 20% ape reziduale variaţia cantităţii de biomasă nu a fost radicală. Valorile
ei au crescut treptat până la a 15-a zi de cultivare şi au depăşit evident cantitatea de biomasă obţinută de pe
proba-martor.
Asemănător ca şi la specia Phormidium foveolarum, pe mediul cu adaos de 10% ape reziduale s-au obţi-
nut rezultate înalte la cultivarea speciei Tribonema viride (Fig.5). Analizând cantitatea de biomasă acumulată
în perioada de cultivare, putem constata că, spre deosebire de celelalte specii implementate în experienţele de
cultivare, cea mai mare cantitate a fost depistată la a 10-a zi în lotul cu 30% ape reziduale (2,10 ± 0,03 g/l).
Fig.5. Cantitatea de biomasă (g/l) de Tribonema viride crescută
pe medii cu adaos de ape reziduale orăşeneşti.
În lotul cu 50% ape reziduale rezultatele obţinute au fost mai slabe, în unele cazuri chiar şi faţă de proba-
martor (mediul chimic Tamia), fapt ce se datorează dezvoltării intense a multor specii de clorofite pe lângă
cultura inoculată.
Deşi comun pentru cele patru specii algale a fost mediul de cultivare cu 10% adaos de ape reziduale şi
perioada optimală de 10-15 zile, rezultatele au fost diferite: pentru specia Chlorella vulgaris cantitatea de
biomasă a crescut cu 71,68% faţă de cantitatea inoculată; în cazul speciei Synechocystis salina – cu 67,91%.
Rezultat maximal a fost obţinut la cultivarea speciei Phormidium foveolarum (88,10%), iar rezultat minimal –
la cultivarea speciei Tribonema viride (45,65%).
Concluzii 1. Pentru cultivarea speciilor Chlorella vulgaris, Synechocystis salina, Phormidium foveolarum şi
Tribonema viride pot fi utilizate mediile cu adaos de ape reziduale în intervalul 1-50%, iar perioada de
10-15 zile este optimală pentru cultivarea acestor specii.
2. În urma cultivării speciei Chlorella vulgaris cele mai bune rezultate au fost obţinute în mediul cu 5%
adaos de ape reziduale (17,55 g/l), iar pentru specia Synechocystis salina cantitatea maximală de
16,56 g/l a fost stabilită în mediul cu adaos de 20%. La cultivarea speciilor filamentoase Phormidium
foveolarum şi Tribonema viride cele mai bune rezultate au fost obţinute în probele cu mediul de 10%
şi, respectiv, de 30% ape reziduale.
3. Comun pentru toate speciile a fost mediul cu adaos de 10% ape reziduale, însă rezultatele obţinute diferă
de la caz la caz. Cele mai bune rezultate au fost stabilite la cultivarea speciei Phormidium foveolarum,
iar cantitatea minimă a fost constatată în variantele cu inoculat de Tribonema viride.
Bibliografie:
1. CONG–FA, Y., ZHONG–YANG, D., KE–CHANG, Zh. Growth of Chlorella pyrenoidosa in wastewater from
Cassava ethanol fermentation. In: World J. Microbiol. Biotechnol., 2008, no.24, p.2919-2925.
2. GLUCK, S. Producing fuels and chemicals from algae and the intersection of the water industry. In: Water
Environment Federation proceeding of the Water Environment Federation WEFTEC, 2009, p.7916-7922.
3. GONZALEZ, C., MARCINIAK, J., VILLAVERDE, S., GARCIA–ENCINA, P. and MUÑOZ, R. Microalgae – based
processes for the biodegradation of pretreated piggery wastewaters. In: Applied microbiology and biotechnology,
2008, vol.80, p.891-898.
4. GONZÁLEZ, L.E., CAÑIZAREZ, R. and BAENA, S. Efficiency of ammonia and phosphorus removal from a
Colombian agroindustrial wastewater by the microalgae Chlorella vulgaris and Scenedesmus dimorphus. In: Journal
Bioresource Technology, 1997, no.60, p.259-262.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
32
5. MULBRY, W., KONDRAD, H., BUYER, J. Treatment of dairy and swine manure effluents using freshwater
algae: fatty acid content and composition of algal biomass at different manure loading rates. In: Journal of Applied
Phycology, 2007, no.109, p.18-924.
6. PARK, J., CRAGGS, R., SHILTON, A. Wastewater treatment high rate algal ponds for biofuel production. In:
Bioresource Technology, 2011, no.102 , p.35-42.
7. QUN, W., ZHIQUAN, H, GENBAO, L, BO, X., HAO, S., MEIPING, T. Removing nitrogen and phosphorus from
simulated wastewater using algae biofilm techinique. In: Environ. Sci. Engin., 2008, no.2(4), p.4467-451.
8. SRIRAM, S., SEENIVASAN, R. Microalgae Cultivation in Wastewater for Nutrient Removal. In: J. Algal Biomass
Utln, 2012, no.3 (2), p.9-13.
9. ŞALARU, V.M., ŞALARU, V.V. Utilizarea apelor reziduale de la complexele zootehnice în scopul obţinerii bio-
masei furajere. În: Studia Universitatis. Seria „Ştiinţe ale naturii”, 2008, nr.1, p.47-55.
10. TROFIM, A. Evaluarea stării ecologice a râului Cogâlnic şi elaborarea metodelor de epurare a apei / Autoref.
tezei de doct. în biol. Chişinău, 2013. 30 p.
11. VALSAMMA, J., AMMINI, J. Algae in the assessment of industrial wastewater holdings ponds. In: Journal
Water, Air and Soil Pollution, 2001, no.132, p.251-261.
12. ДЁГЕ, А. Альгофлора сточных вод живодноводческих комплексов и ее значение в процессах биологической
очистки / Дисс. канд. биол. наук. Кишинев, 1983. 280 с.
13. МАКАРОВА, Е.И., ОТУРИНА, И.П., СИДЯКИН, А.И. Прикладные аспекты применения микроводорослей –
обитателей водных экосистем. В: Экосистемы, их оптимизация и охрана, 2009, вып. 20, c.120-133.
14. СИРЕНКО, Л.А., САКЕВИЧ, А.И., ОСИПОВ, Л.Ф. Методы физиолого–биохимического исследования водо-
рослeй в гидробиологической практике. Киев: Наукова думка, 1975. 248 с.
Prezentat la 04.10.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
33
ROLUL UNOR ALGE CIANOFITE AZOTFIXATOARE
ÎN REZOLVAREA PROBLEMEI ALIMENTARE
Vasile ŞALARU, Valentina BULIMAGA, Victor ŞALARU,
Alina TROFIM, Liliana ZOSIM, Maria PISOV
Universitatea de Stat din Moldova
Este prezentată o sinteză a rezultatelor cercetărilor din ultimele decenii privind rolul algelor cianofite, în primul rând
al celor azotfixatoare, în calitate de biofertilizatori şi amelioratori ai solului şi influenţa lor asupra plantelor de cultură.
Sunt evaluate unele metode de obţinere a inoculului de alge cainofite azotfixatoare şi discutate unele tehnologii de imo-
bilizare a lor. Biofertilizatorii sunt componente importante ale managementului integrat al nutrienţilor. Ei sunt rentabili
din punct de vedere economic şi ecologic şi, totodată, pot servi ca sursă de substanţe biologic active şi ca sursă de pro-
duse alimentare. Algele cianofite reprezintă una dintre sursele regenerabile pentru a suplimenta sau substitui îngrăşă-
mintele chimice şi agenţii de biocontrol în sisteme agricole durabile. Aceste îngrăşăminte biologice potenţiale pot juca
un rol-cheie în productivitatea plantelor de cultură, precum şi în protejarea mediului înconjurător, fiind ecologice şi cu
costuri mai convenabile pentru fermieri.
Cuvinte-cheie: alge cianofite azotfixatoare, biofertilizatori, tehnologii de cultivare ecologice.
THE ROLE OF NITROGEN FIXING CYANOPHYTE ALGAE
IN THE SOLVING OF NUTRITION PROBLEM
The review of research results in recent decades on the role of cyanophyte algae, primarily nitrogen fixing algae,
as biofertilizers and soil improvers and their influence on crop plants are presented. Some methods for obtaining algal
inoculum of nitrogen fixing cyanophyte algae are evaluated and some technologies of their immobilization are discussed.
Biofertilizers are the important components of integrated management of nutrients, being economically profitable and
environmentally friendly, serving as a source of biologically active substances and sources of food. Cyanophyte algae
are one of the renewable sources to supplement or substitute chemical fertilizers and biocontrol agents in sustainable
agricultural systems. The potential biological fertilizer can play a key role in the productivity of crop plants, as well as
protecting of the environment, being ecological and cost-efficiently for farmers.
Keywords: nitrogen fixing cyanophyte algae, biofertilizers, technologies of cultivation, environmentally friendly.
Introducere
Una dintre cele mai alarmante probleme cu care se confruntă azi societatea este epuizarea catastrofală a resurselor naturale, în primul rând epuizarea produselor alimentare. Cu toate că, în linii generale, cantitatea
produselor alimentare sporeşte din an în an, totuşi pe glob se simte un deficit în alimentaţia populaţiei. Încă în anul 1977 acad. Е.К. Федоров scria că circa o treime din populaţia de pe glob suferă de foame cronică, iar
anual zeci de milioane de oameni mor din cauza insuficienţei de hrană [67]. Circa 70 la sută din cei care mor de foame sunt copii.
În scopul diminuării decalajului dintre creşterea numărului populaţiei şi posibilităţile de producere a pro-duselor alimentare oamenii de ştiinţă selectează noi soiuri productive de plante şi animale, elaborează tehno-
logii avansate în fito- şi zootehnie, propun metode noi de lucrare a solului, valorifică stepele şi pădurile, transformându-le în terenuri arabile etc. Cu toate acestea, deficitul de produse alimentare, obţinute din surse
tradiţionale, creşte din an în an.
În acest context apare necesitatea căutării unor noi surse netradiţionale de obţinere a produselor alimen-tare. O sursă incontestabilă de produse alimentare prezintă algele, care conţin în biomasa sintetizată, în de-
pendenţă de specie, până la 65-75% proteină comestibilă. Pe lângă aceasta, biomasa algală conţine până la 25-40% glucide, 10-40% lipide şi o gamă largă de cele mai diverse principii biologic active necesare orga-
nismului uman. Algele pot contribui direct la sporirea produselor alimentare prin utilizarea nemijlocită a biomasei algale şi, indirect, prin aplicarea algelor în calitate de stimulatori biologici la cultivarea plantelor
agricole şi la creşterea animalelor domestice, fiind utilizate totodată şi în calitate de fertilizatori biologici ai solului, ca sursă de remaniere a solurilor degradate etc. Toate acestea se referă în primul rând la algele ciano-
fite (Cyanophyta), care, pe lângă particularităţile menţionate mai sus, caracteristice pentru alge în general , posedă şi un şir de alte însuşiri valoroase, ca: organisme azotofixatoare, indicatori ai calităţii mediului

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
34
ambiant, producători de cele mai diverse substanţe biologic active, printre care şi multe toxine. Până în pre-zent sunt descrise şi pasportizate cca 2000 specii de alge cianofite. În condiţiile Moldovei au fost depistate
cca 350 specii de cianofite acvatice şi edafice în proporţii de aproximativ 1:1 [48,63,68]. Cianofitele sunt
cele mai primitive organisme vegetale pe Pământ, a căror vârstă depăşeşte 3,4 mild. ani, de aceea se întâlnesc şi se acomodează uşor la cele mai extremale condiţii de viaţă [66]. Tocmai din aceste considerente studierea
acestor organizme prezintă un deosebit interes atât teoretic, cât şi aplicativ. În continuare vom expune în cele mai generale linii rezultatele cercetărilor asupra rolului unor alge cianofite, în temei al celor azotofixatoare,
în sporirea fertilităţii solului şi obţinerii produselor alimentare, deficitul cărora devine din an în an tot mai evident.
Algele cianofite prezintă o grupă de procariote fotosintetice, care manifestă proprietăţi de ameliorare a
fertilităţii solului, servesc ca producători de substante biologic active, precum şi ca biosorbenţi ai metalelor
grele (bioremediere) [15]. Aceste organisme au nevoie de lumina soarelui, ca o sursă de energie pentru fixa-
rea carbonului şi azotului. De aceea, ele posedă un potenţial valoros care poate fi valorificat, fiind utilizate
în calitate de biofertilizatori, ceea ce va reduce cu mult cheltuielile de producere a culturilor agricole [37].
Potenţialul ameliorator al biofertilizatorilor este asigurat nu doar de algele cianofite heterocistice, care se pot
dezvolta de sine stătător, dar şi în simbioză cu specii de plante sau bacterii [13]. Algele cianofite s-au mani-
festat ca biofertilizatori potriviţi pentru sistemele de cultivare a orezului, fiind principalele componente ale
ecosistemelor, care menţin umiditatea pentru culturile de orez şi servesc în calitate de cele mai ieftine surse
de biofertilizatori naturali [23,47]. Totodată, cianofitele azotfixatoare pot fi utilizate nu doar la cultivarea ore-
zului, ci şi a multor altor culturi.
Fixarea azotului este un fenomen global important, prin care azotul molecular, unul dintre componentele
cele mai abundente ale atmosferei Pământului, este într-o formă mult mai redusă, adecvată pentru integrarea
în sistemele vii. În mare parte, fixarea azotului se realizează prin mijloace biologice, fiind asigurată de acti-
vitatea microorganismelor [7,38,44]. Acest proces este consumator de energie şi în reacţia biologică de fixare
a azotului este implicat un complex de enzime, numite nitrogenaze, care sunt extrem de sensibile la prezenţa
oxigenului [5,40,43]. Astfel, majoritatea microorganismelor participă la acest proces de fixare a azotului nu-
mai în condiţii adecvate anaerobe sau microaerobe.
Cu toate acestea, unele microorganisme diazotrofice fixatoare de azot au avantajul de a asimila azotul în
medii aerobe. Remarcabile printre acestea sunt procariotele fotosintetizante sau algele cianofite, considerate
a fi progenitori de cloroplaste. Algele cianofite efectuează atât fotosinteza oxigenului, cât şi fixarea azotului
sensibilă la oxigen, servind astfel ca o platformă pentru captarea energiei solare şi utilizarea ei în procesele
metabolice din celule [53]. Printre tulpinile fixatoare de azot, algele cianofite filamentoase au fost intens stu-
diate pentru contribuţia lor la ciclul azotului în ecosistemele acvatice şi terestre [17,29,32,64].
Unele dintre aceste tulpini filamentoase dezvoltă celule speciale, numite heterocişti, care permit segrega-
rea spaţială a fotosintezei de fixarea azotului. Aceşti heterocişti au, de asemenea, rate mai mari de consum de
oxigen, ceea ce duce practic la un mediu anoxic propice pentru enzima nitrogenaza [4]. Spre deosebire de
acestea, algele cianofite non-heterocistice duc lipsă de compartimente speciale, libere de oxigen, şi de multe
ori este nevoie de incubare în condiţiile microaerobe sau anaerobe pentru fixarea azotului [39]. Cu toate
acestea, unele tulpini de alge cianofite non-heterocistice pot asimila azot în condiţii aerobe. Acestea includ
unele specii de filamentoase, astfel ca Cyanothece [21,24,35].
Al doilea nutrient important după azot pentru plante şi microorganisme este fosforul (P). Cele mai multe
sisteme acvatice au resurse limitate, în cazul în care P şi N sunt adesea nutrienţii primari limitatori. Micro-
organismele acvatice pentru supravieţuire menţin net creşterea populaţiei cu un nivel de resurse mai mici
decât cele impuse de alte specii [52]. Algele sunt în special adaptate pentru a curăţa de poluanţi mediul în
care se dezvoltă, prin schimbări structurale, de depozitare sau prin eficientizarea utilizării resurselor [54].
Structura celulei algale implică adaptări biochimice şi fiziologice, fiind apte de a elimina substanţe pentru
sporirea disponibilităţii nutrienţilor.
În procesul metabolismului algele cianofite elimină în mediul înconjurător o serie de substanţe extracelu-
lare, care influenţează creşterea şi dezvoltarea plantelor de cultură [30,31,62]. Aceste alge aduc beneficii plan-
telor prin promovarea creşterii producerii de vitamine, aminoacizi, polipeptide, substanţe antibacteriene şi
antifungice, care exercită biocontrolul asupra fitopatogenilor. Un rol aparte revine excreţiei de către algele
cianofite a polimerilor, în special a exopolizaharidelor, care ameliorează structura solului, dar şi a exoenzime-

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
35
lor, precum şi producerea şi furnizarea de hormoni de creştere a plantelor [25]. Aşadar, efectul benefic al
algelor asupra dezvoltării plantelor de cultură nu poate fi limitat la capacitatea lor de a remedia doar azotul
atmosferic, dar, de asemenea, şi de a elibera substanţe bioactive.
Cercetările recente vin să confirme cu noi date importanţa algelor ca sursă de materie organică în sol
[15,16,50]. Materia organică produsă de alge, precum şi produsele de descompunere a lor, acţionează ca agent
de legătură pentru textura solului; astfel, creşte conţinutul de humus în sol, făcându-l mai fertil pentru creşte-
rea plantelor. Acumularea humusului este de asemenea importantă pentru retenţia hidratării [6].
Efectul unei vaste varietăţi de substanţe extracelulare produse de către alge, inclusiv de algele cianofite,
joacă un rol important în habitatele din apă şi, totodată, are un rol important în îmbunătăţirea germinării seminţe-
lor şi creşterii plantelor superioare [12,47]. Acestea pot fi substanţele bogate în azot, inclusiv aminoacizii [59],
vitamina B12 şi biotina [31], precum şi hormoniii de creştere, inclusiv: auxinele [61], citokininele [56], gibe-
relinele sau acidul giberelic [51,44], antibioticele, algicidele, toxinele, acizii organici şi alţi compuşi bioactivi
pentru utilizare în farmaceutică [30].
Algele cianofite pot fi cei mai importanţi agenţi de fixare a azotului în multe soluri agricole [20,41,42,55],
precum şi în ameliorarea calităţii lor [57]. Fertilizarea cu alge cianofite a fost utilizată pentru prima dată la
culturile de orez [62]. Ele sunt utilizate pe scară largă în câmpurile de orez din diverse ţări, pentru consolidarea
fertilităţii solului prin intermediul procesului de fixare a azotului biologic (aşa-numita algalizare) în loc de
îngrăşăminte bogate în azot [20,42]. Biofertilizatorii sunt susceptibili de a-şi asuma un rol semnificativ la
substituirea sau completarea îngrăşămintelor chimice pentru ameliorarea aportului de nutrienţi în culturile de
cerealiere, pentru a exclude costurile ridicate ale îngrăşămintelor, precum şi în protecţia mediului ambiant de
poluare şi în majorarea productivităţii culturilor agricole [1,49]. Importanţa lor ca fixatori de azot în câmpu-
rile de culturi agricole a fost studiată de către mai mulţi cercetători [2,19,20,42]. Utilizarea algelor cianofite
ca biofertilizatori pentru câmpurile de orez este foarte promiţătoare, dar limitată, din cauza fluctuaţiei în cali-
tatea şi cantitatea de inocul şi atributele fiziologice în diverse regiuni agro-ecologice.
În mai multe regiuni ale Indiei, câmpurile de orez sunt fertilizate de Azolla – ferigă de apă care se înmul-
ţeşte rapid şi conţine ca simbiot alga cianofută Anabaena, care fixează azotul atmosferic, contribuind astfel la
creşterea evidentă a roadei. În genere, algele joacă un rol economic important în sporirea fertilităţii solului şi
în îmbunătăţirea condiţiilor de creştere a plantelor de cultură. Mecanismele utilizate pentru a stimula creş-
terea plantelor include: biofertilizarea (creşterea ofertei de substanţe nutritive minerale la plante), controlul
biologic (eliminarea vătămătorilor plantelor, inclusiv patogeni microbieni, insecte şi buruieni), producerea şi
furnizarea hormonilor de creştere a plantelor [25,44,51,56,60]. Tehnicile de biofertilizare cu utilizarea alge-
lor cianofite sunt recomandate pentru creşterea ratei de germinare a seminţelor şi ameliorarea parametrilor de
creştere a plantelor [14,44,47,49].
Încă din primele cercetări asupra algelor cianofite în unele regiuni ale Indiei, Gupta şi colabolatorii (1964)
au observat că ele accelerează germinarea seminţelor şi creşterea răsadului. În plus, s-a observat că atât randa-
mentul, cât şi calitatea boabelor au fost îmbunătăţite, privind conţinutul de proteine. Inocularea algelor ciano-
fite fixatoare de azot reprezintă o sursă alternativă şi durabilă de azot pentru creşterea productivităţii plante-
lor de cultură. Inoculul mixt de Nostoc, Anabaena, Westiellopsis, Aulosira şi Scytonema a fost folosit în studii
de teren în 2005 [34]. Pentru comparaţie a fost utilizat separat şi inoculul de Anabaena. Inoculul mixt supli-
mentat cu N: P: K (30:20:20 kg/ha) s-a dovedit a fi cel mai eficient tratament pentru productivitatea plantelor
de cultură.
Algele cianofite diazotrofice heterocistice posedă capacitatea de a forma asociaţii cu plantele superioare
şi de a produce substanţe de stimulare a creşterii. Cele mai multe dintre lucrările referitoare la inoculanţi de
alge cianofite au fost axate pe culturile de orez, iar alte culturi au fost mai puţin studiate.
Experimentele efectuate de noi în ultimii ani demonstrează că algalizarea solului cu alge azotfixatoare în
condiţii de seră stimulează evident productivitatea plantelor leguminoase (date nepublicate).
A fost raportat un studiu de evaluare a potenţialului a trei tulpini de alge cianofite din rizosfera de grâu şi
a influenţei lor asupra creşterii acestei plante în experimentele de cultivare a culturii în ghivece. Tratamentele
aplicate cu cele trei tulpini în combinare cu 1/3 de N + P + K a dat rezultate echivalente statistic în compa-
raţie cu aplicarea dozei totale de îngrăşăminte chimice, în ceea ce priveşte randamentele de cereale [19].
A fost cercetată şi influenţa inoculului de Nostoc asupra creşterii culturii de porumb în ghivece. Suspensia
tulpinii de Nostoc la o concentraţie de 1 g (greutate uscată) pe litru a fost uniform turnată în cantitate de 6 g/m2,

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
36
respectiv, peste 2 tipuri de sol în ghivece, curând după germinarea porumbului şi s-a efectuat cultivarea în
decurs de 6 săptămâni. În ambele cazuri s-a observat creşterea conţinutului de N în sol, comparativ cu ghive-
cele cu soluri netratate cu Nostoc. A fost observat şi un spor al randamentului biomasei vegetale uscate cu 40
şi 49%, respectiv, pentru fiecare tip de sol [27].
Cercetările efectului inoculatului a două alge cianofite Anabaena oryzae şi Nostoc ellipsoidum asupra cul-
turii de tomate a scos în evidenţă că atât suspensia, cât şi filtratele obţinute din aceste două alge (tratate sau
netratate termic) n-au influenţat semnificativ asupra germinării seminţelor de tomate. Totuşi, filtratul din Ana-
baena supus tratării termice s-a dovedit a fi mai eficient decât suspensia celor două alge şi filtratul de Nostoc.
Un efect mai semnificativ a fost observat asupra creşterii răsadului de tomate în sol nisipos cu inoculul de
Anabaena în combinare cu amelioratorul sintetic pe bază de poliacriamidă de sodiu – Broadleaf-P4 0,2%
W/W, manifestată prin majorarea lungimii rădăcinii şi a mugurilor, precum şi a masei lor verzi şi uscate [2].
Cercetările ulterioare [44] au demonstrat rolul benefic al unor substanţe bioactive eliberate de algele ciano-
fite asupra ratei de germinare a seminţelor de Senna alexandrina, fiind evaluate şi modificări metabolice la
această plantă medicinală. Autorii menţionaţi au raportat că exometaboliţii produşi de Nostoc piscinale şi de
N. muscorum au majorat rata de germinare a seminţelor de Senna până la 100 şi 90%, respectiv, după 60 de
ore. De asemenea, s-a depistat că exudatele algale conţin concentraţii variabile de acid abscisic (ABA), acid
giberelic (GA3), indol acid acetic (IAA) şi alţi metaboliţi, care ar putea fi implicate în calitate de agenţi alelo-
chimici. Ei au constatat, de asemenea, o creştere semnificativă a proteinelor şi glucidelor solubile totale, la
tratare cu exudate algale, în special cu cele de N. piscinale şi N. muscorum.
Au fost studiate efectele alelopatice ale exudatelor de N. muscorum, N. piscinale şi Anabaena fertilissima
asupra unor componente biochimice ale plantei comestibile Cynara cardunculus [44]. Analiza cantitativă a
exudatelor algelor cianofite au evidenţiat prezenţa fitohormonilor, aminoacizilor, compuşilor solubili în apă
cu conţinut de azot şi a zaharurilor reducătoare. Datele obţinute au evidenţiat o creştere semnificativă în tota-
lul de zăharuri solubile şi a conţinutului de proteine în seminţele germinate tratate cu diferite exudate algale.
În plus, tratamentul cu exudate algale stimulează rata de germinare a seminţelor după 96 de ore.
Cu toate că algele cianofite pot servi şi în calitate de agenţi de biocontrol al acţiunii dăunătorilor asupra
plantelor, ele au fost mai puţin utilizate în acest scop. Principalii agenţi biologici care au fost studiaţi pentru
controlul patogenic la plante sunt fungiile şi bacteriile. Încă în 1995 a fost efectuată o analiză a datelor referi-
toare la evaluarea potenţialului cianofitelor şi al altor alge pentru utilizarea lor în controlul biologic al bacte-
riilor patogene la plante [22]. Caire şi colaboratorii [8] au demonstrat că extractul apos de N. muscorum în
concentraţii diluate demonstrează eficienţa lui în controlul biologic. În ultimele decenii, diferiţi cercetători au
studiat posibilitatea înlocuirii pesticidelor chimice cu componente naturale extrase din diferite plante şi surse
microalgale pentru utilizare în calitate de agenţi insecticidici [Nassar et al., 1999], agenţi acaricidici [3,11] şi
agenţi fungici [16,45,61]. Aceste substanţe naturale posedă şi acţiune letală asupra dăunătorilor şi au rolul de
a ocroti mediul ambiant. Intensificarea activităţii de cercetare, privind controlul biologic, este dictată de ne-
cesitatea de a elabora o schemă conceptuală a gestionării integrate a dăunătorilor, ca o abordare acceptabilă
de control al afectării ecosistemului, iar realizarea acestui control biologic trebuie să devină una dintre com-
ponentele de bază în practicile de gestionare a dăunătorilor.
Algele cianofite joacă un rol important în menţinerea biotei solului fără dăunători printr-un management
adecvat, care permite asigurarea culturilor cu nutrienţi şi menţinerea sănătăţii solului, ce reprezintă un sistem
dinamic, în care componentele fizice, chimice şi biotice sunt într-o stare de echilibru. Aplicarea insecticidelor
chimice fără a ţine seama de alte componente ale solului dereglează acest echilibru, ceea ce afectează produc-
tivitatea solului. Insecticidele exercită frecvent efecte inhibitoare sau stimulatoare asupra creşterii sau altor
activităţi ale microorganismelor, fie în cultură pură sau în câmp. A fost demonstrată şi activitatea insecticidă
a algelor cianofite asupra a 4 insecte, ceea ce indică la posibilitatea de utilizare a lor în calitate de agenţi în
lupta cu insectele [33].
Cianofitele, în special cele azotfixatoare, reprezintă microorganisme importante care contribuie la fertili-
tatea solului. Aceste organisme joacă un rol evident în acest sistem prin furnizarea constantă de azot fixat şi
alte efecte benefice. Cea mai mare parte a algelor din sol şi a microalgelor acvatice este sensibilă la insecticide
datorită faptului că algele sunt angajate în fotosinteză [50], iar multe insecticide interferează cu acest proces.
Algele cianofite pot fi încorporate în sol nu doar ca materie organică, ci şi ca sursă de enzime, deoarece
acestea produc fosfatazele alcaline şi acide extracelulare, care sunt active în soluţie sau sunt situate în spaţiul

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
37
periplasmatic al peretelui celular. Atât biomasa, cât şi exopolizaharidele încorporate în sol, induc o promo-
vare a creşterii altor microorganisme şi sporirea activităţii enzimelor din sol, care participă la eliberarea de
substanţe nutritive necesare plantelor [8].
Un rol nu mai puţin important revine algelor cianofite şi în bioremedierea solului, datorită capacităţii lor
înalte de biosorbţie a metalelor grele. La fel ca şi alte microorganisme, algele cianofite sunt capabile să acu-
muleze ionii metalici din mediul ambiant [36], inclusiv radionucleidele [9]. Mecanismele de rezistenţă a
algelor cianofite la metale grele implică: factori de mediu, mecanisme specifice şi non-specifice de protecţie
a celulei, dezvoltate ca răspuns al celulei la impactul toxic al metalului asupra speciei. Dintre mecanismele
intracelulare de protecţie un rol important ocupă mecanismul de biosorbţie a metalelor grele bazat pe schimbul
de ioni în peretele celular exterior, acoperit cu teci mucilaginoase care se comportă ca o „vacuolă externă”.
Proprietăţile de legare a metalului se datorează prezenţei grupărilor anionice, în special a grupărilor carboxil,
identificate în polimerul capsular. Acest grup de microorganisme ar putea avea o eficienţă mai mare în bio-
sorbţie, în timpul creşterii lor în medii poluate. Această legare a metalelor sporeşte conţinutul de materie
organică în crustă [10], îmbunătăţeşte rezistenţa solului la eroziunea provocată din cauza vântului şi a apei.
Importanţa microorganismelor în consolidarea stabilităţii agregatelor de sol este bine cunoscută, fiind demon-
strată agregarea sporită în cazul când solurile au fost inoculate cu alge sau cu alge cianofite [26]. Astfel, este
evident rolul algelor în protecţia solului şi în dezvoltarea plantelor de cultură.
Algele cianofite sunt componente importante ale ecosistemelor aride şi semiaride. Distribuirea lor în na-
tutră indică starea de sănătate a mediului. În ultimii ani se investighează posibilitatea de folosire a algelor
cianofite ca umidificator biologic al solului, deoarece teaca gelatinoasă, care acoperă celula sau trihomii, are
capacitatea de a acumula vaporii de apă din atmosferă.
Astfel, algele cianofite sunt principalii agenţi fotosintetici prezenţi în solurile din întreaga lume, rolul eco-
logic al cărora este inestimabil şi nu este încă pe deplin definit. Dintre efectele lor asupra sistemului plantă/
sol pot fi evidenţiate următoarele [50]:
Excreţia de acizi organici care majorează disponibilitatea şi absorbţia fosforului.
Furnizarea de azot prin fixarea azotului biologic.
Sporirea cantităţii de materie organică din sol.
Eliminarea în procesul metabolismului a substanţelor bioactive extracelulare, care influenţează creşte-
rea şi dezvoltarea plantelor vasculare. Acestea sunt: reglatorii de creştere a plantelor (PGRs), vitami-
nele, aminoacizii, polipeptidele, substanţele antibacteriene sau antifungice, care exercită biocontrolul
asupra fitopatogenilor, precum şi polimeri, în special exopolizaharide şi exoenzime, care în comun
influenţează pozitiv structura şi fertilitatea solului.
Formarea crustei şi stabilizarea agregării solului prin formarea de agregate a polizaharidelor extrace-
lulare.
Acumulatori de ioni metalici prezenţi în mediul ambiant.
În continuare sunt expuse rezultatele unor cercetări referitoare la metodele de cultivare a algelor cianofite
pentru obţinerea inoculului pentru aplicare în calitate de biofertilizatori.
Cultivarea biomasei de alge cianofite ca biofertilizatori şi metode de inoculare a lor în sol. Pentru culti-
varea în aer liber a biomasei algelor cianofite ca sursă de biofertilizatori este necesară utilizarea unor tulpini
specifice regionale. Conform datelor prezentate de unii cercetători, un amestec de 5 sau 6 tulpini regionale
aclimatizate de alge cianofite este utilizat în general pentru inoculul iniţial (starter). La cultivarea, de exemplu,
a orezului sunt utilizate următoarele metode de obţinere a inoculului de alge cianofite: a) metoda bazinului
(rezervorului); b) metoda gropii căptuşite cu polietilenă; c) metoda de teren şi d) metoda pepinierei [58].
Metoda bazinului (rezervorului) presupune pregătirea cuvelor permanente sau tăvilor de tablă zincată de
anumite dimensiuni (2 m x 3 m x 23 cm), care pot fi modificate în funcţie de cantitatea de material produs.
În dependenţă de suprafaţă, în bazin se introduc 8-10 kg de sol şi se amestecă minuţios cu 200 g superfosfat.
Tăvile sunt umplute cu apă într-un strat de 5-15 cm, în funcţie de condiţiile locale şi de rata de evaporare.
pH-ul solului este păstrat la valoarea neutră – 7,0, prin adăugare de var. În scopul de a preveni infectarea cu
insecte dăunătoare, la acest sol se adaugă orice insecticid corespunzător, de preferinţă carbofuran (3% granule).
Cultura pură de alge cianofite azotfixatoare este adusă din laborator şi introdusă în apa limpezită din tavă sau
cuvă, după ce solul s-a stabilizat, apoi se expune la soare, în aer liber. Se adaugă apă intermitent pentru a
compensa pierderea datorată evaporării. În timpul verii inoculatul creşte viguros şi formează un covor gros în

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
38
decurs de 7-10 zile. La această etapă, udarea este oprită, astfel încât covorul algal se usucă la soare şi for-
mează fulgi, care sunt colectaţi şi stocaţi pentru utilizare pe teren. Fulgii sunt colectaţi şi utilizaţi ca inocul
pentru înmulţirea ulterioară. Procesul poate fi repetat de trei sau patru ori cu sol proaspăt, cu adaos de super-
fosfat şi insecticide, după cum e descris mai sus. Două recolte de fulgi de alge cianofite, astfel produse, dă
aproximativ 3-5 kg de material activ, suficient pentru a biofertiliza, de exemplu, un hectar de câmp de orez
sau alte culturi [46,58].
Metoda gropii căptuşite cu polietilenă este cea mai potrivită pentru fermierii mici şi cei mijlocii la prepa-
rarea algelor ca biofertilizatori. În câmp sunt pregătite gropi mici, căptuşite cu folii groase de polietilenă în
care se cultivă algele azotfixatoare menite pentru algalizarea solului.
Metoda de teren este o versiune a metodei rezervorului, însă la o scară mai largă, folosită pentru a genera
cantităţi mari de material algal biofertilizator. Câmpul de producţie este împărţit în parcele, care sunt inun-
date cu un strat subţire de apă deasupra solului. Se utilizează superfosfat şi acelaşi insecticid, la fel ca în me-
toda rezervorului. Apoi, pe suprafaţa apei se presoară inoculatul de alge cianofite. Nivelul apei este menţinut
întotdeauna intermitent. Pe vreme însorită, în cazul în care solul este argilos, termenul optim de creştere a
algelor cianofite va fi de două săptămâni. După formarea unui covor des plutitor, udarea este oprită şi fulgii
algali după uscare la soare sunt colectaţi şi depozitaţi în saci pentru a fi utilizaţi de fermieri. Procesul poate
fi repetat pe aceleaşi parcele de 2-3 ori, doar cu adaos de superfosfat şi insecticide, dar fără suplimentarea
inoculului. Prin această metodă se atinge un randament mediu de biofertilizare algală de 16-30 kg pe un teren
de 40 m2
la fiecare recoltare. Adoptarea acestei metode în unele regiuni din India a oferit fermierilor o pro-
ducţie record de 15,6 tone boabe de orez /ha [58].
Cultivarea biomasei de alge cianofite se face prin intermediul uneia dintre metodele descrise mai sus, care
include următoarele etape:
Se pregătesc rezervoare cimentate, tăvi din foi inox sau de plastic sau gropi căptuşite cu polietilenă,
aliniate într-o zonă deschisă. Lăţimea rezervoarelor sau a gropilor să nu fie mai mare de 1,5 m. Acest
lucru va facilita manipularea corectă a culturii.
Se transferă 4-5 kg sol (colectate de pe teren deschis) şi se adaugă 100 g de superfosfat. În groapă la
acest amestec se adaugă apă până la o înălţime de aproximativ 10 cm. Se amestecă cu var pentru a ajusta
valoarea pH-ului până la 7. Se adaugă 2 ml de insecticid (de exemplu, malathion) pentru a proteja cul-
tura de ţânţari. Se amestecă bine şi se lasă să se sedimenteze particulele de sol.
Când apa se limpezeşte, se presară 100 g de inocul pe suprafaţa apei. La temperatura de 35-40°C dez-
voltarea algelor cianofite atinge cota maximă. În această perioadă permanent se menţine nivelul de apă
la aproximativ 10 cm.
După uscare, culturile de alge din sol se vor obţine separat sub formă de fulgi. În timpul verii biomasa
algală produsă este de circa 1 kg/m2
de suprafaţă. Ea este colectată sub formă de pulbere, păstrată în
saci de polietilenă, sigilaţi şi livraţi către fermieri.
Fulgii de alge pot fi folosiţi ca inocul iniţial, în cazul repetării procedurii de cultivare.
Prin metoda de pepinieră se recomandă producerea biofertilizatorilor algali împreună cu plantele de cul-
tură, răsădite în pepiniere. 15-20 kg de material de alge cianofite sunt uşor puse la biofertilizare eficientă pe
un teren de 1,5 ha. Astfel, pământul nu este utilizat exclusiv doar pentru producţia de biofertilizatori algali,
însă este folosit mai eficient [30].
Biofertilizatorii sunt componente importante ale managementului integrat al nutrienţilor. Ei sunt rentabili
din punct de vedere economic şi ecologic şi pot servi totodată ca sursă de substanţe nutritive pentru plante.
Algele cianofite reprezintă una dintre sursele natural regenerabile pentru a suplimenta sau substitui îngrăşă-
mintele chimice şi agenţii de biocontrol în sisteme agricole durabile. Aceste îngrăşăminte biologice potenţiale
vor juca un rol-cheie în productivitatea şi durabilitatea solului, precum şi în protejarea mediului înconjurător,
fiind ecologice şi cu costuri mai eficiente pentru fermieri.
Imobilizarea cianobacteriilor într-o matrice solidă, cum ar fi spuma de poliuretan pentru producerea de
amoniac, maximizarea productivităţii în câmpurile agricole şi fixarea azotului endofitic în câmpurile cu grâu
sunt considerate tehnologii potenţiale pentru viitor [18,19].
Este propusă şi imobilizarea tulpinii de Nostoc ANTH pe alginat de Na, care a demonstrat păstrarea viabi-
lităţii culturii până la 3 ani, chiar şi în cazul când inoculul de Nostoc în alginat era lăsat dehidratatat timp
de 3-4 luni, apoi se efectua periodic hidratarea lui cu mediu de cultură. În experienţe de laborator a fost

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
39
demonstrată şi capacitatea nostocului imobilizat pe alginat de a forma asociaţii cu tulpinile de grâu. Totuşi,
din cauza costului majorat al alginatului acest suport ar putea fi utilizat doar pentru păstrarea culturii viabile
de Nostoc şi mai puţin pentru fertilizare [28]. În scopul imobilizării algelor cianofite pentru păstrare şi utilizare
ca biofertilizatori ar putea sluji suporturile naturale ieftine, biodegradabile, cum sunt paiele sau alte resturi
vegetale.
Aşadar, algele cianofite existente în sol sunt rezerve potenţiale importante pentru agricultura ecologică,
servind ca biofertilizatori (pentru algalizarea solului) sau amelioratori ai solului [65]. Aplicarea lor în practică
rămâne însă limitată din cauza cunoştinţelor incomplete privind ecologia microbiană a solului, ecofiziologia,
formula produsului şi tehnologiile de utilizare. Primul pas spre o agricultură ecologică constituie selectarea
suşelor de alge cianofite din diferite regiuni ale ţării şi separarea lor în culturi pure. Următoarea etapă constă
în elaborarea tehnologiilor de cultivare şi de obţinere a biomasei pentru algalizarea solului. Este necesară şi
testarea ulterioară a inocului ca biofertilizator la cultivarea anumitor plante pe soluri concrete. Etapa finală
în acest proces va constitui elaborarea tehnologiei de obţinere a lor pentru sporirea productivităţii culturilor
agricole. Producerea biofertilizatorilor în proporţii mici şi mijlocii prin tehnologii prietenoase mediului ar
diminua consumul de îngrăşăminte azotate şi ar deschide noi perspective în asigurarea populaţiei cu alimente
ecologice şi benefice pentru sănătatea populaţiei.
Bibliografie:
1. AHMED, A.A.S. Cyanobacterial application for the improvement of soil fertility: M. Sc. Thesis, Botany Dept.,
Fac. of Science, Beni-Suef University, Egypt, 2009.
2. ALI AL-KHIAT, S.H. Effect of Cyanobacteria as a Soil Conditioner and Biofertilizer on Growth and Some Bioche-
mical Characteristics of Tomato (Lycopersicon esculentum L.) Seedlings: Thesis for Degree of Master of Science
(M. Sc.) Microbiology (Algae), 2006.
3. AMER, S.A.A., MOHAMED, S.M., EL-GENGAIHI, S et.al. Acaricidal activity of lipoidal matter of different plant
extracts against the two-spotted mite. In: Tetranychus urticae koch. Insect. Sci. Appl., 2000, vol.20, no.3, p.191-194.
4. BERGMAN, B., GALLON, J.R., RAI, A.N., et.al. N2 fixation by non-heterocystous cyanobacteria. În: FEMS
Microbiol. Rev., 1997, vol.19, p.139-185.
5. BERMAN-FRANK, I., CHEN, Y., GERCHMAN, Y. et.al. Inhibition of nitrogenase by oxygen in marine cyano-
bacteria controls the global nitrogen and oxygen cycles, 2005, vol.2, p.261-273.
6. BOLYSHEV, N., NOVICHKOVA-IVANOVA, L. The algal flora of initial stages of soil formation on blanket
loam under the condition of a lysimeter. In: Bot. Zurn. (Moscow & Leningrad), 1978, vol.63, p.690-696.
7. BURRIS, R., ROBERTS, G. Biological nitrogen fixation. In: Annal Rev. Nutr., 1993, vol.13, p.317-335.
8. CAIRE, G., DE CANO, M., PALMA., R. et.al Changes in soil enzyme activities following additions of cyanobac-
terial biomass and exopolysaccharide. In: Soil Biology & Biochemistry, 2000, vol.32, no.13, p.1985-1987.
9. CECAL, A., RUDIC, V., GULEA, A. et al. Recuperation of uranil ions from effluents by means of microbiological
collectors. In: Waste Management, 1997, vol.17, no.1, p.97-99.
10. DANIN, A., BAR-OR, Y., YISRAELI, T. The role of Cyanobacteria instabilisation of sand dunes in southern
Israel. In: Ecol. Mediterr., 1989, vol.15, p.55-64.
11. DUKE, S., DAYAN, F.E., RIMANDO, A.M. Chemicals from nature for weed management. In: Weed Sci., 2002,
vol.50, no.2, p.138-151.
12. EL-AYOUTY, Y. Soil inoculation valuable by blue-green algae and their effects on yield attributes of different
rice varieties. Proc., 6th
Egyptian. November 24-26, Bot. Con. Cairo Univ., 1998, vol.11, p.221-330.
13. EL-ZEKY, M., EL-SHAHAT, R., METWALY, G.et.al. Using Cyanobacteria or Azolla as alternative nitrogen
sources for rice production. In: J. Agric. Mansoura Univ., 2005, vol.30, no.9, p.5567-5577.
14. GUPTA, A., LATA, K. Effect of algal growth hormones on the germination of paddy seeds. In: Hidrobiologia,
1964, vol.24, no.1-3, p.430-434.
15. IBRAHEEM, I. Cyanobacteria as alternative biological conditioners for bioremediation of barren soil. In: Egyptian
J. Phycol., 2007, vol.8, p.99-116.
16. IBRAHEEM, I., ABDEL-RAOUF, N. Allelopathic activity of some local cyanobacterial extra-metabolites against
some pathogenic bacteria. In: Egypt. J. Phycol., 2007, vol.8, p.120-122.
17. KANEKO, T., NAKAMURA, Y., WOLK, C., et.al. Complete genomic sequence of the filamentous nitrogen-fixing
cyanobacterium Anabaena sp. strain PCC 7120. In: DNA Res., 2001, vol.8, p.205-213.
18. KANNAIYAN, S. Biotechnology of Biofertilizers. Editor Spriger, 2002. 375 p. ISBN: 9781402002199
19. KARTHIKEYAN, N., PRASANNA, R., LATA, N. et al. Evaluating the potential of plant growth promoting cyano-
bacteria as inoculants for wheat. In: European Journal of Soil Biology. 2007, vol.43, no.1, p.23-30.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
40
20. KAUSHIK, B. Use of cyanobacterial biofertilizer in rice cultivation: A Technology Improvement. In: G.SUBRA-
MANIAN, B.D. KAUSHIK, G.S. VENKATARAMAN. Cyanobacterial Biotechnology. Editor Science Publs. Inc.
USA; 1998, p.211-222.
21. KOMÁREK, J., CEPÁK, V. Cytomorphological characters supporting the taxonomic validity of Cyanothece
(Cyanoprokaryota). In: Plant. Syst. Evol., 1998, vol.210, no.1-2, p.25-39.
22. KULIK, M. The potential for using Cyanobacteria (blue-green algae) and algae in the biological control of plant
pathogenic bacteria and fungi. In: Eur. J. Plant. Pathol., 1995, vol.101, no.6, p.585-599. ISSN 0929-1873
23. LADHA, J., REDDY, P. Nitrogen fixation in rice systems: state of knowledge and future prospects. In: Plant Soil.,
2003, vol.252, p.151-167.
24. LIBERTON, M., AUSTIN JR, I., BERG, R. et al. Unique thylakoid membrane architecture of a un cellular N2-fixing
cyanobacterium revealed by electron tomography. In: Plant. Physiol., 2011, vol.155, no.1, p.1656-1666.
25. LUGTENBERG, B.J., DE WEGER, L., BENNETT, J. Microbial stimulation of plant growth and protection from
disease. In: Current Opinion in Microbiology, 1991, vol.2, no.3, p.457-464.
26. LUTZOW, M., KOGEL-KNABNER, I., EKSCHMITT, K. et al. Stabilization of organic matter in temperate soils:
mechanisms and their relevance under different soil conditions. In: European Journal of Soil Science, 2006, vol.57,
no.4, p.426-445.
27. MAQUBELA, M.P., MNKENI, P.N.S., MALAM-ISSA, O. Nostoc cyanobacterial inoculation in South African
agricultural soils enhances soil structure, fertility and maize growth. In: Plant. Soil., 2009, vol.315, p.79-92.
28. MAYASHREE, B. Entrapped cyanobacteria: Implications for biotechnology. In: Indian Journal of Biotechnology,
2005, vol.4, no.4, p.209-215.
29. MEEKS, J., ELHAI, J., THIEL, T. et al. An overview of the genome of Nostoc punctiforme, a multicellular, sym-
biotic cyanobacterium. In: Photosynth., Res., 2001, vol.70, p.85-106.
30. METTING, B., PYNE, J.W. Biologically active compounds from microalgae. In: Enzyme Microbiol. Technol.,
1986, vol.8, no.7, p.386-394.
31. MISRA, S., KAUSHIK, B. Growth promoting substances of Cyanobacteria. I. Vitamin and their influence. In: FAO
Soils. Bull., 1989, vol.46, no.1, p.1-52.
32. MULLIGAN, M., HASELKORN, R. Nitrogen-fixation (nif) genes of the cyanobacterium Anabaena sp. strain PCC
7120: the nzjB-fdxN-nifS-nzjU operon. In: J. Biol. Chem., 1989, vol.264, p.19200-19207.
33. NASSAR, M.M.I., HAFEZ, S., NAGATY, I. et al. Insecticidal activity of Cyanobacteria against four insects, two
of medical importance and two agricultural pests with reference to the action on albino mice. In: Egyptian Soc.
Parasitol., 1999, vol.29, no.3, p.939-949.
34. PAUDEL, Y.P., PRADHAN, S., PANT, B. et al. Role of blue green algae in rice productivity.In: Agric. Biol. J. N.
Am., 2012, vol.3, no.8, p.332-335. ISSN Online: 2151-7525
35. PORTA, D., RIPPKA, R., HERNÁNDEZ-MARINÉ, M. Unusual ultrastructural features in three strains of Cyanothece
(cyanobacteria). In: Arch. Microbiol., 2000, vol.173, p.154-163.
36. RANA, L., CHHIKARA, S., DHANKHAR, R. Assessment of Growth Rate of Indigenous Cyanobacteria in Metal
Enriched Culture Medium. In: Asian J. Exp. Biol. Sci., 2013, vol.4, no.3, p.465-471.
37. RAOUF, N., AL-HOMAIDAN, A., IBRAHEEM, I. Agricultural importance of algae. In: African Journal of
Biotechnology, 2012, vol.11(54), p.11648-11658.
38. RAYMOND, J., SIEFERT, J., STAPLES, C. The natural history of nitrogen fixation. In: Mol. Biol. Evol., 2004,
vol.21, p.541-554.
39. RIPPKA, R., DERUELLES, J., WATERBURY, J. et al. Generic assignments, strain histories and properties of pure
cultures of cyanobacteria. In: J. Gen. Microbiol., 1979, vol.111, p.1-61.
40. ROBSON, R., POSTGATE, J. Oxygen and hydrogen in biological nitrogen fixation. In: Annual Rev. Microbiol.,
1980, vol.34, p.183-207.
41. RODRIGO, V., EBERTO, N. Seasonal changes in periphyton nitrogen fixation in a protected tropical wetland. In:
Biol. Fertil. Soils., 2007, vol.43, p.367-372.
42. ROGER, P.-A. Rice field cyanobacteria: ecology, contribution to soil fertility and practical utilisation. In: MOROT-
GAUDRY J.F. (ed.) Nitrogen assimilation by plants: physiological, biochemical and molecular aspects. Enfield,
New Hamshire: Science Publ., 2001, p.199-226. ISBN 2-7380-0716-3
43. RUBIO L., LUDDEN, P. Biosynthesis of the iron-molybdenum cofactor of nitrogenase. In: Annual Rev.
Microbiol., 2008, vol.62, p.93-111.
44. SAFFAN, E., MOHAMED, A. Response of Senna alexandrina to cyanobacterial exudates. In: Az. J. Pharm. Sci.,
2001, vol.27, p.266-279.
45. SAFONOVA, E., REISSER, W. Growth promoting and inhibiting effects of extracellular substances of soil micro-
algae and cyanobacteria on Escherichia coli and Micrococcus luteus. In: Phycol. Res., 2005, vol.53, p.189-193.
46. SAHU, D., PRIYADARSHANI, I., RATH, B. Cyanobacteria - as potential biofertilizer. CIBTech. In: Journal of
Microbiology, 2012, vol.1, no.2-3, p.20-26. ISSN 2319-3867

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
41
47. ŞALARU, V.V., ŞALARU, V.M. Procedee de tratare a seminţelor de castraveţi înainte de semănare: Brevet de
invenţie, MD 2998, BOPI 2006, nr.3, p.22-23.
48. ŞALARU, V.M., ŞALARU, V.V., ANDRONIC, E. şi al. Unele rezultate ale studierii algoflorei efectuate în USM
în ultimii 10 ani. În: Analele Ştiinţifice ale USM, Seria „Ştiinţe chimico-biologice”, 2006, p.35-45.
49. ŞALARU, V.V., ŞALARU, V.M., CHICU, N., DUDNICENCO, T. Algele şi rolul lor în sporirea fertilităţii solului.
În: Serviciul agrochimic la 4 decenii de afirmare, 2004, p.113-121.
50. SHARMA, R., KHOKHAR, M.K., JAT, R.L. et al. Role of algae and cyanobacteria in sustainable agriculture
system. In: Wudpecker Journal of Agricultural Research, 2012, vol.1, no.9, p.381-388. ISSN 2315-7259
51. SHEN-RUI, Z., SHEN, R.Z. A broad spectrum biopesticide type biofertilizer anaerobic fermentation effluent and
plant adverse resistance. In: Acta. Agr. Shanghai, 1997, vol.13, no.2, p.89-96.
52. SILKE, R., VIGDIS, T., FRIDA, L. Nitrogen availability decreases prokaryotic diversity in sandy soils. In: Biol.
Fertil. Soils., 2007, vol.43, p.449-459.
53. SIMPSON, F., BURRIS, R. A nitrogen pressure of 50 atmospheres does not prevent evolution of hydrogen by
nitrogenase. In: Science, 1984, vol.224, p.1095-1097.
54. SINGH, N., DHAR, D. Nitrogen and phosphorous scavenging potential in microalgae. In: India J. Biotechnol.,
2007, vol.6, p.52-56.
55. STEWART, W. Algal fixation of atmospheric nitrogen. In: Pl. Soil., 1970, vol.32, p.555-588.
56. STRICK, W., STADEN, J., VAN-STADEN, J. Screening of some South African seaweeds for cytokinin-like activity.
In: South Afr. J. Bot., 1997, vol.63, no.3, p.161-164.
57. SUBHASHINI, D., KAUSHIK, B. Amelioration of sodic soils with blue-green algae. In: Australian Journal of Soil
Research, 1981, vol.19, no.3, p.361-366.
58. VAISHAMPAYAN, A., SINHA, R.P., HADER, D.-P. et al. Cyanobacterial biofertilizers in rice agriculture. In:
Botanical Review, 2001, vol.67, no.4, p.453-516.
59. VARGA, L., SZIGETI, J., ORDOG, V. Effect of Spirulina platensis biomass enriched with trace elements on com-
binations of starter culture strains employed in the dairy industry. In: Milchwissen-Schaft, 1999, vol.54, no.5, p.247-
248.
60. VENKATARAMAN, G. Blue-green algae for rice production-a manual for its promotion on rice plant. In: Proc.
Indian Nat. Sci., Acad., Part-B, Biol. Sci., 1981, vol.55, no.4, p.295-300.
61. VOLK, R., FURKERT, F. Antialgal, antibacterial and antifungal activity of two metabolites produced and excreted
by cyanobacteria during growth. In: Microbiol. Res., 2006, vol.161, p.180-186.
62. WATANABE, A., ITO, R., KONISHI, C. Effect of nitrogen fixing blue green algae on the growth of rice plants.
In: Nature, 1951, vol.168, p.748-749.
63. WWW.alge.md
64. ZEHR, J.P. Nitrogen fixation by marine cyanobacteria. In: Trends in microbiology, 2011, vol.19, no.4, p.162-173.
65. ZIMMERMAN, W.J. Microalgal biotechnology and applications in agriculture. In the book: Soil microbial
ecology: applications in agricultural and environmental management. Editor Metting, F.B., Jr., 1992, p.457-479,
ISBN 0-8247-8737-4
66. ВИНОГРАДОВА, О.Н. Синезеленые водоросли экстремальных местообитаний: Автореферат дис. на соиск.
учёной степени доктора биол. наук. Киев, 2013.
67. ФЕДОРОВ, Е.К. Экологический кризис и социальный прогресс. Москва: Гидрометеоиздат. 1977. 175 с.
68. ШАЛАРЬ, В.М., НАБЕРЕЖНЫЙ, А.И. Роль сине-зеленых водорослей в водоемах Молдавской ССР. В:
Экология и физиология сине-зеленых водорослей. Москва-Ленинград, 1965.
Notă: Lucrarea este publicată cu susţinerea financiară a Proiectului instituţional 11.817.08.44A
Prezentat la 12.09.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
42
INFLUENŢA UNOR COMPUŞI COORDINATIVI AI Cu(II) ASUPRA
PRODUCTIVITĂŢII ŞI CONŢINUTULUI DE PIGMENŢI FOTOSINTETICI LA
MICROALGA DUNALIELLA SALINA CNM-AV-02
Liliana ZOSIM, Cezara BIVOL, Ludmila BATÎR*,
Daniela ELENCIUC**
, Svetlana DJUR*,Olga OLAN
Universitatea de Stat din Moldova
*Institutul de Microbiologie şi Biotehnologie al Academiei de Ştiinţe a Moldovei
**Universitatea Academiei de Ştiinţe a Moldovei
A fost studiată influenţa compuşilor coordinativi ai Cu(II) şi a intensităţii luminii asupra productivităţii şi conţinu-
tului de carotenoizi şi clorofilă a în biomasa de dunalielă. S-a constatat că intensitatea luminii, temperatura şi adaptarea
prealabilă la noul mediu de nutriţie folosit (mediul organo-mineral MD, preparat în baza lichidului cultural al spirulinei)
joacă un rol determinant în iniţierea şi parcurgerea fazei exponenţiale de dezvoltare a dunalielei.
Cuvinte-cheie: Dunaliella salina, mediu organo-mineral, biomasă, productivitate, pigmenţi fotosintetici, compuşi
coordinativi ai Cu(II), sinteză orientată.
THE INFLUENCE OF SOME COORDINATION COMPOUNDS OF Cu (II) ON PRODUCTIVITY AND
PHOTOSYNTHETIC PIGMENT CONTENT IN MICROALGA DUNALIELLA SALINA CNM-AV-02
The dunaliella productivity, content of carotenoids and chlorophyll a in the dunaliella biomass at cultivation in the
presence of some Cu(II) coordination compounds at the different light intensity was determined. The light intensity,
temperature and prior adaptation to the new nutrition medium (the organic-mineral medium MD, prepared on the basis
of spirulina cultural liquid) plays a determinant factor in the initiation and crossing of dunaliella's exponential grown
phase.
Keywords: Dunaliella salina, organic-mineral medium, biomass, productivity, photosynthetic pigments, coordination
compounds of Cu(II), directed synthesis.
Introducere
Dintre toate speciile genului Dunaliella, microalga Dunaliella salina prezintă interes şi avantaj practic în cultivarea industrială a microalgelor datorită conţinutului sporit de carotenoizi, care depăşeşte conţinutul
acestora din plantele superioare şi poate constitui până la 10% din biomasă. Pigmenţii carotenoizii sunt pe larg utilizaţi în industria alimentară în calitate de coloranţi naturali, în industria farmaceutică, în practica me-
dicinală, dar şi în alimentaţia terapeutico-profilactică la tratarea diferitelor maladii. De asemenea, dunaliela
este considerată a fi potenţial producător de glicerol, care poate depăşi 30% din substanţa uscată a celulei, proprietate valoroasă pentru sursele alternative de carburanţi în industria energetică [3,10,11,17].
D. salina prezintă interes atât sub aspect evolutiv, cât şi fiziologic, ca obiect-model de cercetare a mecanis-melor de osmoreglare, manifestând şi rezistenţă la factori limitanţi de mediu (iradiere puternică, deshidratare,
prezenţa substanţelor toxice sau a metalelor grele etc.) [1,2,7,9]. Investigaţiile precedente întreprinse în cadrul LCŞ ,,Ficobiotehnologie” (USM şi AŞM) demonstrează că
condiţiile de stres (absenţa sau excesul unor nutrienţi, modificarea regimului de temperatură, iluminare şi ira-diere, salinitate înaltă, modificarea valorii pH), precum şi administrarea unor compuşi coordinativi ai metale-lor de tranziţie joacă un rol determinant în procesele de creştere, dezvoltare şi de acumulare a substanţelor bioactive în biomasa dunalielei. Iar datorită metabolizării de către D. salina a microelementelor din structura compuşilor coordinativi biomasa microalgei capătă proprietăţi calitative noi (imunostimulatoare, regenera-toare, citoprotectoare etc.), proprietăţi valoroase în obţinerea unor produse polifuncţionale ecologice, cu pro-prietăţi unicale [5,6,13,14].
Tendinţele biotehnologiei moderne impune obţinerea produselor în ciclu închis prin reutilizarea deşeurilor industriale [12,16,21]. În acest context, dunaliela reprezintă un obiect atractiv de studiu în vederea cultivării
pe deşeuri lichide de la producerea în special a biomasei altor microalge. Scopul lucrării constă în evaluarea productivităţii şi conţinutului de pigmenţi fotosintetici în biomasa
microalgei D. salina cultivate în prezenţa unor compuşi coordinativi ai Cu(II).
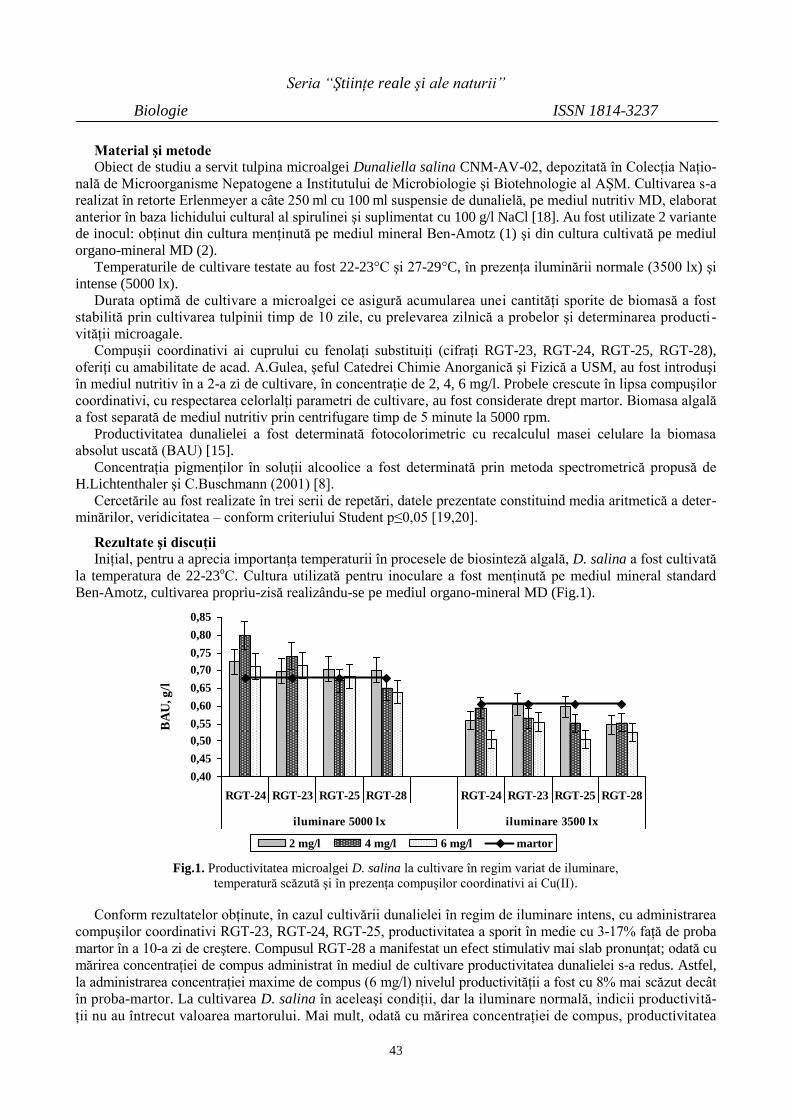
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
43
Material şi metode Obiect de studiu a servit tulpina microalgei Dunaliella salina CNM-AV-02, depozitată în Colecţia Naţio-
nală de Microorganisme Nepatogene a Institutului de Microbiologie şi Biotehnologie al AŞM. Cultivarea s-a realizat în retorte Erlenmeyer a câte 250 ml cu 100 ml suspensie de dunalielă, pe mediul nutritiv MD, elaborat
anterior în baza lichidului cultural al spirulinei şi suplimentat cu 100 g/l NaCl [18]. Au fost utilizate 2 variante de inocul: obţinut din cultura menţinută pe mediul mineral Ben-Amotz (1) şi din cultura cultivată pe mediul
organo-mineral MD (2). Temperaturile de cultivare testate au fost 22-23°C şi 27-29°C, în prezenţa iluminării normale (3500 lx) şi
intense (5000 lx).
Durata optimă de cultivare a microalgei ce asigură acumularea unei cantităţi sporite de biomasă a fost stabilită prin cultivarea tulpinii timp de 10 zile, cu prelevarea zilnică a probelor şi determinarea producti-
vităţii microagale. Compuşii coordinativi ai cuprului cu fenolaţi substituiţi (cifraţi RGT-23, RGT-24, RGT-25, RGT-28),
oferiţi cu amabilitate de acad. A.Gulea, şeful Catedrei Chimie Anorganică şi Fizică a USM, au fost introduşi în mediul nutritiv în a 2-a zi de cultivare, în concentraţie de 2, 4, 6 mg/l. Probele crescute în lipsa compuşilor
coordinativi, cu respectarea celorlalţi parametri de cultivare, au fost considerate drept martor. Biomasa algală a fost separată de mediul nutritiv prin centrifugare timp de 5 minute la 5000 rpm.
Productivitatea dunalielei a fost determinată fotocolorimetric cu recalculul masei celulare la biomasa absolut uscată (BAU) [15].
Concentraţia pigmenţilor în soluţii alcoolice a fost determinată prin metoda spectrometrică propusă de H.Lichtenthaler şi C.Buschmann (2001) [8].
Cercetările au fost realizate în trei serii de repetări, datele prezentate constituind media aritmetică a deter-minărilor, veridicitatea – conform criteriului Student p≤0,05 [19,20].
Rezultate şi discuţii
Iniţial, pentru a aprecia importanţa temperaturii în procesele de biosinteză algală, D. salina a fost cultivată
la temperatura de 22-23oC. Cultura utilizată pentru inoculare a fost menţinută pe mediul mineral standard
Ben-Amotz, cultivarea propriu-zisă realizându-se pe mediul organo-mineral MD (Fig.1).
0,40
0,45
0,50
0,55
0,60
0,65
0,70
0,75
0,80
0,85
RGT-24 RGT-23 RGT-25 RGT-28 RGT-24 RGT-23 RGT-25 RGT-28
iluminare 5000 lx iluminare 3500 lx
BA
U, g
/l
2 mg/l 4 mg/l 6 mg/l martor
Fig.1. Productivitatea microalgei D. salina la cultivare în regim variat de iluminare,
temperatură scăzută şi în prezenţa compuşilor coordinativi ai Cu(II).
Conform rezultatelor obţinute, în cazul cultivării dunalielei în regim de iluminare intens, cu administrarea
compuşilor coordinativi RGT-23, RGT-24, RGT-25, productivitatea a sporit în medie cu 3-17% faţă de proba
martor în a 10-a zi de creştere. Compusul RGT-28 a manifestat un efect stimulativ mai slab pronunţat; odată cu
mărirea concentraţiei de compus administrat în mediul de cultivare productivitatea dunalielei s-a redus. Astfel,
la administrarea concentraţiei maxime de compus (6 mg/l) nivelul productivităţii a fost cu 8% mai scăzut decât
în proba-martor. La cultivarea D. salina în aceleaşi condiţii, dar la iluminare normală, indicii productivită-
ţii nu au întrecut valoarea martorului. Mai mult, odată cu mărirea concentraţiei de compus, productivitatea

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
44
dunalielei a scăzut în medie cu 6-16% sub nivelul martorului. Cel mai semnificativ efect stimulator asupra
productivităţii l-a manifestat compusul RGT-24, la a cărui administrare în concentraţie de 4 mg/l, în regim de
iluminare intensă, productivitatea a fost cu 17,8% mai înaltă decât în proba-martor. Evaluarea în dinamică a creşterii culturii de dunaliela pe mediul organo-mineral suplimentat cu compusul
coordinativ RGT-24 timp de 10 zile a evidenţiat faptul că durata Lag-fazei de creştere este prelungită şi du-rează 72 ore (Fig.2). Fenomenul dat este caracteristic culturilor ce se află în proces de adaptare la condiţiile
nespecifice sau noi de viaţă. În cazul experienţei efectuate, celulele de dunaliela au fost inoculate pe mediul organo-mineral MD direct din mediul lor tradiţional de cultivare (mediul Ben-Amotz), ceea ce presupune ini-
ţial adaptarea culturii la condiţiile nespecifice de existenţă. Plus la aceasta, suplimentarea compusului coordi-nativ a putut favoriza instalarea unui stres determinat de prezenţa în mediul de cultivare a unor concentraţii
înalte de cupru.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1 zi 2 zi 3 zi 5 zi 7 zi 10 zi
Durata cultivării, zile
BA
U,
g/l
iluminare 5000 lx
iluminare 3500 lx
Fig.2. Dinamica productivităţii dunalielei cultivate pe mediul organo-mineral
suplimentat cu RGT-24 în concentraţie de 4 mg/l.
Intensitatea luminii joacă un rol determinant în iniţierea şi parcurgerea fazei exponenţiale de dezvoltare a du-
nalielei. Odată intrată în faza exponenţială, cultura intens iluminată evoluează mai rapid, cantitatea de biomasă
sporeşte pe parcursul a 10 zile de creştere de 4 ori (de la 0,20 g/l la 0,8 g/l), ceea ce este de 1,34 ori mai mult de-
cât în cazul cultivării dunalielei pe acelaşi mediu, în prezenţa aceluiaşi compus, dar în regim de iluminare obişnuit.
Cercetările au continuat cu studiul efectului aceloraşi compuşi coordinativi ai Cu(II) asupra productivităţii
şi duratei de cultivare a dunalielei la temperatură înaltă de 27-29°C şi utilizarea inoculului crescut prealabil
10 zile pe mediul organo-mineral MD. Rezultatele obţinute au indicat influenţa temperaturii sporite, de 27-29°C,
asupra productivităţii dunalielei cultivate atât în prezenţa compuşilor coordinativi, cât şi în lipsa lor (Fig.3 şi 4).
0,75
1,00
1,25
1,50
1,75
2,00
2,25
2,50
RGT-
24
RGT-
23
RGT-
25
RGT-
28
RGT-
24
RGT-
23
RGT-
25
RGT-
28
iluminare 5000 lx iluminare 3500 lx
BA
U, g
/l
2mg/l 4mg/l 6mg/l martor
Fig.3. Productivitatea microalgei D. salina la cultivare în regim variat de iluminare,
temperatură înaltă şi în prezenţa compuşilor coordinativi ai Cu(II).
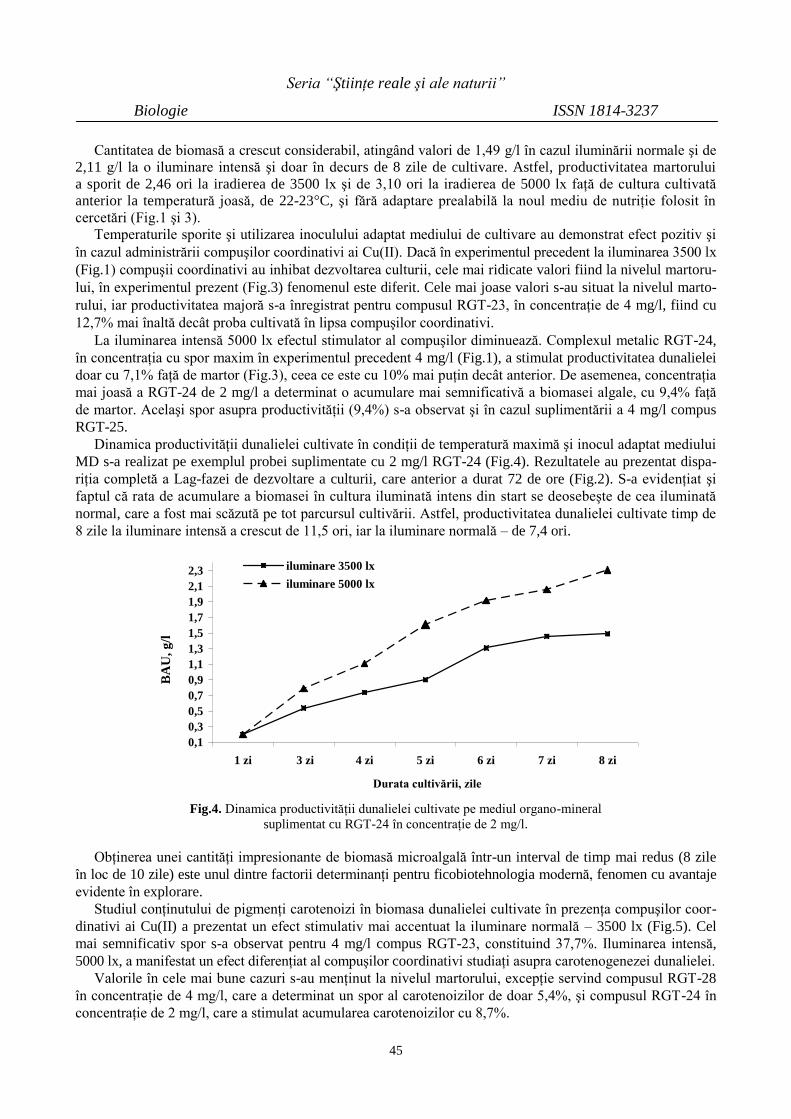
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
45
Cantitatea de biomasă a crescut considerabil, atingând valori de 1,49 g/l în cazul iluminării normale şi de
2,11 g/l la o iluminare intensă şi doar în decurs de 8 zile de cultivare. Astfel, productivitatea martorului
a sporit de 2,46 ori la iradierea de 3500 lx şi de 3,10 ori la iradierea de 5000 lx faţă de cultura cultivată
anterior la temperatură joasă, de 22-23°C, şi fără adaptare prealabilă la noul mediu de nutriţie folosit în
cercetări (Fig.1 şi 3).
Temperaturile sporite şi utilizarea inoculului adaptat mediului de cultivare au demonstrat efect pozitiv şi
în cazul administrării compuşilor coordinativi ai Cu(II). Dacă în experimentul precedent la iluminarea 3500 lx
(Fig.1) compuşii coordinativi au inhibat dezvoltarea culturii, cele mai ridicate valori fiind la nivelul martoru-
lui, în experimentul prezent (Fig.3) fenomenul este diferit. Cele mai joase valori s-au situat la nivelul marto-
rului, iar productivitatea majoră s-a înregistrat pentru compusul RGT-23, în concentraţie de 4 mg/l, fiind cu
12,7% mai înaltă decât proba cultivată în lipsa compuşilor coordinativi.
La iluminarea intensă 5000 lx efectul stimulator al compuşilor diminuează. Complexul metalic RGT-24,
în concentraţia cu spor maxim în experimentul precedent 4 mg/l (Fig.1), a stimulat productivitatea dunalielei
doar cu 7,1% faţă de martor (Fig.3), ceea ce este cu 10% mai puţin decât anterior. De asemenea, concentraţia
mai joasă a RGT-24 de 2 mg/l a determinat o acumulare mai semnificativă a biomasei algale, cu 9,4% faţă
de martor. Acelaşi spor asupra productivităţii (9,4%) s-a observat şi în cazul suplimentării a 4 mg/l compus
RGT-25.
Dinamica productivităţii dunalielei cultivate în condiţii de temperatură maximă şi inocul adaptat mediului
MD s-a realizat pe exemplul probei suplimentate cu 2 mg/l RGT-24 (Fig.4). Rezultatele au prezentat dispa-
riţia completă a Lag-fazei de dezvoltare a culturii, care anterior a durat 72 de ore (Fig.2). S-a evidenţiat şi
faptul că rata de acumulare a biomasei în cultura iluminată intens din start se deosebeşte de cea iluminată
normal, care a fost mai scăzută pe tot parcursul cultivării. Astfel, productivitatea dunalielei cultivate timp de
8 zile la iluminare intensă a crescut de 11,5 ori, iar la iluminare normală – de 7,4 ori.
0,1
0,3
0,5
0,7
0,9
1,1
1,3
1,5
1,7
1,9
2,1
2,3
1 zi 3 zi 4 zi 5 zi 6 zi 7 zi 8 zi
Durata cultivării, zile
BA
U,
g/l
iluminare 3500 lx
iluminare 5000 lx
Fig.4. Dinamica productivităţii dunalielei cultivate pe mediul organo-mineral
suplimentat cu RGT-24 în concentraţie de 2 mg/l.
Obţinerea unei cantităţi impresionante de biomasă microalgală într-un interval de timp mai redus (8 zile
în loc de 10 zile) este unul dintre factorii determinanţi pentru ficobiotehnologia modernă, fenomen cu avantaje
evidente în explorare.
Studiul conţinutului de pigmenţi carotenoizi în biomasa dunalielei cultivate în prezenţa compuşilor coor-
dinativi ai Cu(II) a prezentat un efect stimulativ mai accentuat la iluminare normală – 3500 lx (Fig.5). Cel
mai semnificativ spor s-a observat pentru 4 mg/l compus RGT-23, constituind 37,7%. Iluminarea intensă,
5000 lx, a manifestat un efect diferenţiat al compuşilor coordinativi studiaţi asupra carotenogenezei dunalielei.
Valorile în cele mai bune cazuri s-au menţinut la nivelul martorului, excepţie servind compusul RGT-28
în concentraţie de 4 mg/l, care a determinat un spor al carotenoizilor de doar 5,4%, şi compusul RGT-24 în
concentraţie de 2 mg/l, care a stimulat acumularea carotenoizilor cu 8,7%.
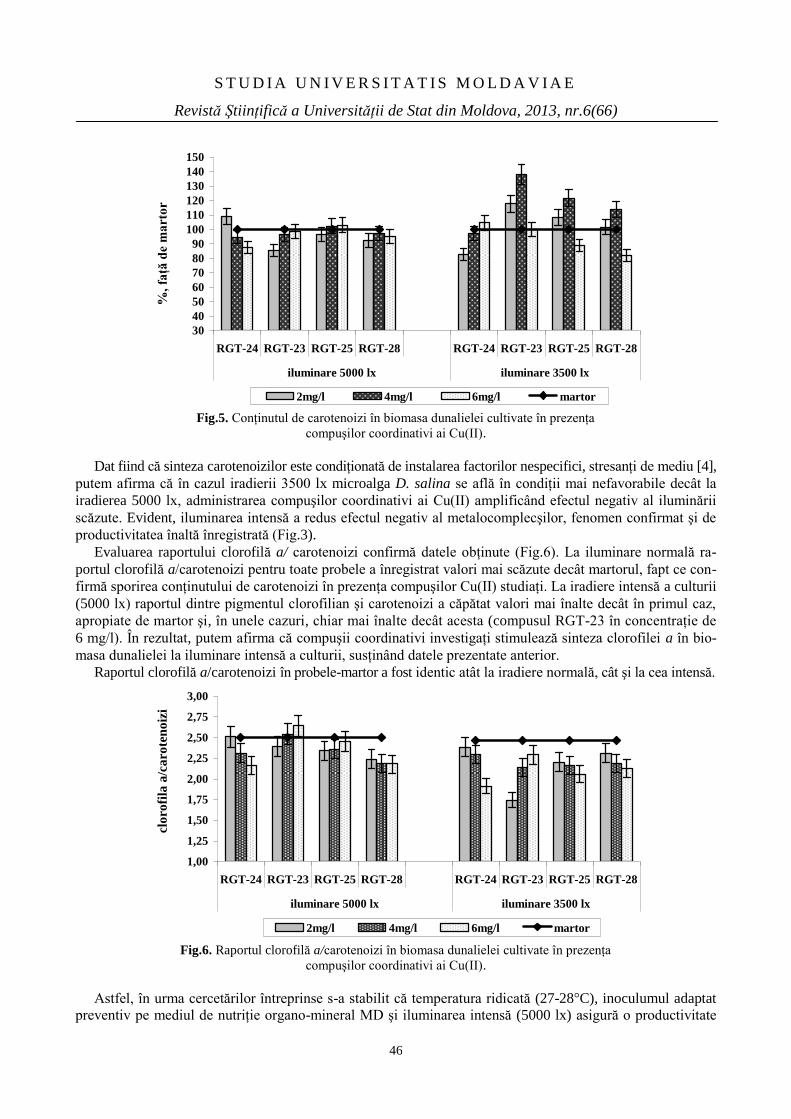
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
46
30
40
50
60
70
80
90
100
110
120
130
140
150
RGT-24 RGT-23 RGT-25 RGT-28 RGT-24 RGT-23 RGT-25 RGT-28
iluminare 5000 lx iluminare 3500 lx
%,
faţă
de
ma
rto
r
2mg/l 4mg/l 6mg/l martor
Fig.5. Conţinutul de carotenoizi în biomasa dunalielei cultivate în prezenţa
compuşilor coordinativi ai Cu(II).
Dat fiind că sinteza carotenoizilor este condiţionată de instalarea factorilor nespecifici, stresanţi de mediu [4],
putem afirma că în cazul iradierii 3500 lx microalga D. salina se află în condiţii mai nefavorabile decât la
iradierea 5000 lx, administrarea compuşilor coordinativi ai Cu(II) amplificând efectul negativ al iluminării
scăzute. Evident, iluminarea intensă a redus efectul negativ al metalocomplecşilor, fenomen confirmat şi de
productivitatea înaltă înregistrată (Fig.3).
Evaluarea raportului clorofilă a/ carotenoizi confirmă datele obţinute (Fig.6). La iluminare normală ra-
portul clorofilă a/carotenoizi pentru toate probele a înregistrat valori mai scăzute decât martorul, fapt ce con-
firmă sporirea conţinutului de carotenoizi în prezenţa compuşilor Cu(II) studiaţi. La iradiere intensă a culturii
(5000 lx) raportul dintre pigmentul clorofilian şi carotenoizi a căpătat valori mai înalte decât în primul caz,
apropiate de martor şi, în unele cazuri, chiar mai înalte decât acesta (compusul RGT-23 în concentraţie de
6 mg/l). În rezultat, putem afirma că compuşii coordinativi investigaţi stimulează sinteza clorofilei a în bio-
masa dunalielei la iluminare intensă a culturii, susţinând datele prezentate anterior.
Raportul clorofilă a/carotenoizi în probele-martor a fost identic atât la iradiere normală, cât şi la cea intensă.
1,00
1,25
1,50
1,75
2,00
2,25
2,50
2,75
3,00
RGT-24 RGT-23 RGT-25 RGT-28 RGT-24 RGT-23 RGT-25 RGT-28
iluminare 5000 lx iluminare 3500 lx
clo
rofi
la a
/ca
rote
no
izi
2mg/l 4mg/l 6mg/l martor
Fig.6. Raportul clorofilă a/carotenoizi în biomasa dunalielei cultivate în prezenţa
compuşilor coordinativi ai Cu(II).
Astfel, în urma cercetărilor întreprinse s-a stabilit că temperatura ridicată (27-28°C), inoculumul adaptat
preventiv pe mediul de nutriţie organo-mineral MD şi iluminarea intensă (5000 lx) asigură o productivitate

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
47
mai sporită la microalga D. salina decât în prezenţa temperaturii 22-23°C, inoculului neadaptat şi la ilumi-
nare de 3500 lx. Aceşti factori determină şi adaptarea culturii la prezenţa compuşilor coordinativi ai Cu(II) în
mediu, diminuând efectul lor inhibitor. S-a evidenţiat compusul RGT-24 în concentraţie de 2 mg/l, asigurând
un spor al biomasei de 9,4% şi de carotenoizi de 8,7%.
Concluzii
1. Intensitatea luminii, temperatura şi adaptarea prealabilă la noul mediu de nutriţie folosit (mediul organo-
mineral MD, preparat în baza lichidului cultural al spirulinei) joacă un rol determinant în iniţierea şi
parcurgerea fazei exponenţiale de dezvoltare a dunalielei.
2. Dintre compuşii coordinativi ai Cu(II) cercetaţi productivitate majoră s-a înregistrat la iluminarea
intensă (5000 lx) pentru compusul RGT-24, în concentraţie de 2 mg/l, fiind cu 9,4% mai înaltă decât
proba cultivată în lipsa compuşilor coordinativi.
3. Conţinutul de pigmenţi carotenoizi în biomasa dunalielei cultivate în prezenţa compuşilor coordinativi
ai Cu(II) a prezentat un efect stimulativ mai accentuat faţă de proba-martor la iluminare normală (3500 lx).
Cel mai semnificativ spor s-a observat pentru 4 mg/l compus RGT-23, constituind 37,7%.
Bibliografie:
1. ABD EL-BAKY, H., HUSSEIN, M., EL-BAROTY, G. Algal extracts improve antioxidant defense abilities on salt
tolerance of wheat plant irrigated with sea water. In: African Journal of Biochemistry Research, 2008, no.2(7),
p.151-164. ISSN 1996-0778
2. AVRON, M., BEN-AMOTZ, A. Dunaliella: physiology, biochemistry and biotechnology. Boca Raton: CRC Press,
1992. 240 p. ISBN 978-0849366475
3. BEN-AMOTZ, A. Industrial production of microalgae cell-mass and secondary products – major industrial species
Dunaliella. In: A.RICHMOND, ed. Handbook of microalgal culture. Oxford: Blakwell, 2004, p.273-280.
4. BHOSALE, P. Environmental and cultural stimulants in the production of carotenoids from microorganisms. In:
Applied Microbiology and Biotechnology, 2004, no.63(4), p.351-361. ISSN 0175-7598
5. BIVOL, C. Parametri morfologici şi biochimici ai microalgei verzi Dunaliella salina la cultivare pe medii minerale
şi organo-minerale. În: Studia Universitatis. Seria „Ştiinţe ale naturii”, 2008, nr.2(12), p.32-35. ISSN 1857-1735
6. BIVOL, C. ş.a. Acţiunea unor compuşi coordinativi ai Fe(III) cu α-aminoacizi asupra conţinutului pigmenţilor
fotosintetici ai microalgei Dunaliella salina CNM-AV-02. În: Buletinul Academiei de Ştiinţe a Moldovei. Seria
„Ştiinţele vieţii”, 2012, nr.2(317), p.149-154. ISSN 1857-064X
7. BOROWITZKA, M., SIVA, C. The taxonomy of the genus Dunaliella (Chlorophyta, Dunaliellales) with emphasis
on the marine and halophilic species. In: Journal of Applied Phycology, 2007, no.19(5), p.567-590. ISSN 0921-8971
8. LIHTENTHALER, H.K., BUSCHMANN, C. Chlorophylls and carotenoids: Measurement and characterization by
UV-VIS spectroscopy. In: Current Protocols in Food Analytical Chemistry. 2001. F4.3.1-F4.3.8.
ISBN 978-0471142911
9. PASQUALETTI, M. et al. Salinity and nitrate concentration on the growth and carotenoids accumulation in a strain
of Dunaliella salina (Chlorophyta) cultivated under laboratory conditions. In: Transitional Waters Bulletin, 2010,
no.4(2), p.94-104. ISSN 1825-229X
10. PRASANNA, R. et al. Potentials and applications of algal pigments in biology and industry. In: Acta Botanica
Hungarica, 2007, no.49(1-2), p.131-156. ISSN 0236-6495
11. RAJA, R. et al. Exploitation of Dunaliella for β-carotene production. In: Applied Microbiology and Biotechnology,
2007, no.74(3), p.517-523. ISSN 0175-7598
12. RICHMOND, A., HU, Q. Handbook of microalgal culture. In: Applied phycology and biotechnology. Oxford:
Blakwell, 2013. 736 p. ISBN 978-0470673898
13. RUDIC, V. Aspecte noi ale biotehnologiei moderne. Chişinău: Ştiinţa, 1993. 139 p. ISBN 978-5376018293
14. RUDIC, V. Ficobiotehnologie – cercetări fundamentale şi realizări practice. Chişinău: Elena V.I., 2007. 364 p.
ISBN 978-9975989251
15. RUDIC, V. ş. a. Metode de investigaţie în ficobiotehnologie. Chişinău: CE USM, 2002. 61 p. ISBN 9975-70-254-6
16. ŞALARU, V. ş. a. Biomasa algală – sursă alternativă de energie, produse alimentare nonpoluante şi substanţe bio-
logic active. În: Studia Universitatis. Seria „Ştiinţe ale naturii”, 2007, no.7, p.196-200. ISSN 1857-1735
17. TAFRESHI, A., SHARIATI, M. Pilot culture of three strains of Dunaliella salina for β-carotene production in open
ponds in the central region of Iran. In: Journal of Microbiology and Biotechnology, 2006, no.22(9), p.1003-1006.
ISSN 1017-7825

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
48
18. UNIVERSITATEA DE STAT DIN MOLDOVA. Procedeu de cultivare a microalgei verzi Dunaliella salina:
brevet MD nr.3780, Inventatori RUDIC, V., BULIMAGA, V., BIVOL, C. CIB A01G33/00. Publ. 31.12.2008,
BOPI nr.12/08.
19. ДОСПЕХОВ, Б. Методика полевого опыта. Москва: Агропромиздат, 1985. 351 с.
20. МАКСИМОВ, В. Многофакторный эксперимент в биологии. Москва: МГУ, 1980. 278 с.
21. ТРЕНКЕНШУ, Р., ГЕВОРГИЗ, Р., БОРОВКОВ, А. Основы промышленного культивирования дуналиеллы
солоноводной (Dunaliella salina Teod.). Севастополь: ЭКОСИ-Гидрофизика, 2005. 103 с.
Notă: Cercetările au fost întreprinse în cadrul Proiectului independent pentru tinerii cercetători 13.819.18.07A,
2013-2014, finanţat de către Consiliul Suprem pentru Ştiinţă şi Dezvoltare Tehnologică al AŞM.
Prezentat la 15.09.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
49
OPTIMIZAREA MATEMATICĂ A MEDIULUI DE CULTURĂ PENTRU
PRODUCEREA ß-GLUCANILOR LA TULPINA
SACCHAROMYCES CEREVISIAE CNMN-Y-20
Natalia CHISELIŢA, Agafia USATÎI, Elena MOLODOI,
Nadejda EFREMOVA, Ludmila FULGA, Tamara BORISOVA
Institutul de Microbiologie şi Biotehnologie al AŞM
În acest studiu sunt reflectate rezultatele optimizării mediului de cultivare pentru Saccharomyces cerevisiae CNMN-
Y-20 în vederea sporirii conţinutului de β-glucani. Evidenţierea ingredientelor mediului de cultivare cu efect stimulator (cum ar fi sursa de carbon şi acetatul de zinc) a fost efectuată aplicând algoritmul Yates. Concentraţiile componentelor mediului de cultură optimizat sunt, g L
-1: zaharoză – 37,0; (NH4)2SO4 – 3,0; MgSO4•7H2O – 0,7; NaCl – 0,5; Ca(NO3)2 –
0,4; KH2PO4 – 1,0; acetat de zinc – 0,00816; autolizat de drojdii – 10 ml. Mediul optimizat utilizat pentru cultivarea tul-pinii de drojdie se caracterizează prin obţinerea a 29,47±0,52% β-glucani în biomasa uscată, sau cu 31,8% mai mult decât mediul iniţial.
Cuvinte-cheie: Saccharomyces cerevisiae, β-glucani, medii de cultură, optimizare.
THE MATHEMATICAL OPTIMIZATION OF CULTURE MEDIUM FOR
Β- GLUCANES PRODUCTION BY SACCHAROMYCES CEREVISIAE CNMN-Y-20 This paper contains results of optimization of culture medium for Saccharomyces cerevisiae CNMN-Y-20 with aim
to increase β-glucans content. The selection of components of culture medium (carbon sources, zinc acetate) with sti-mulatory effect was carried out by applying the Yates algorithm. Concentrations of optimized culture medium compo-nents g L
-1: saccharose – 37,0; (NH4)2SO4 – 3,0; MgSO4•7H2O – 0,7; NaCl – 0,5; Ca(NO3)2 – 0,4; KH2PO4 – 1,0; zinc
acetate – 0,00816; yeast autolysate – 10 ml. Optimized medium used for cultivation of yeast strain is characterized by obtaining 29,47 ± 0.52% β-glucans per dry biomass or 31,8% more than the original medium.
Keywords: Saccharomyces cerevisiae, β-glucans, culture media, optimization.
Introducere Interes comercial prezintă polizaharidele peretelui celular al levurilor, care pot fi utilizate în industria ali-
mentară, farmaceutică, cosmetică. Ţinând cont de siguranţa în cadrul de sănătate publică, până în prezent doar glucanii şi manoproteinele levurilor Saccharomyces cerevisiae au fost acceptate pentru de a fi utilizate pe larg.
β-glucanii au o semnificaţie biomedicală deosebită, datorită activităţii lor antivirale şi antibacteriene [17], imunomodulatoare şi imunostimulatoare [8,22], anticancerigene [20,21,25]. Derivaţii sulfataţi ai β-glucanilor au demonstrat in vitro activitate antiproliferativă impotriva celulelor sarcomei 180 (S-180) [23]. Mai mult ca atât, β-glucanii prezintă substanţa activă de bază la elaborarea noilor vaccinuri anticancerigene polivalente, care sunt actualmente în curs de dezvoltare [16]. În combinaţie cu alte substanţe biologic active β-glucanii au ca-pacitatea de a reduce conţinutul toxinelor din tractul digestiv, mecanismul acestui proces fiind bazat pe comple-xarea toxinelor cu catena polizaharidică cu ajutorul legăturilor de hidrogen şi Van der Waals [24].
Glucanii intră în structura stratului interior al peretelui celular al levurilor genului Saccharomyces [11]. În general, structura şi dinamica compoziţiei chimice a peretelui celular sunt legate de schimbările în starea fizio-logică a celulei şi de reacţia la influenţa factorilor externi. Arhitectura peretelui unei celule şi mecanismele res-ponsabile de sinteza componentelor acestuia pot fi controlate de componenţa mediului de cultură [10,12,13].
Pentru o bună dezvoltare şi derulare a proceselor metabolice pentru tulpinile de drojdie, este necesar ca în mediul de cultură să fie incluse surse de carbon, azot, alţi factori de creştere în concentraţii specifice tulpinii [3]. În calitate de sursă de carbon utilizată pentru fermentare şi pentru a spori randamentul producerii de glucani de către S. cerevisiae pot fi nominalizate glucoza, zaharoza, lactoza, fructoza, maltoza, manoza, amidonul [7]. Glucoza este cotată ca cea mai importantă sursă pentru biosinteza glucanilor şi mananilor [6]. În calitate de sursă de azot organic pentru fermentare se aplică peptona şi extractul de levuri, combinate cu surse de azot anorganic, cum sunt sulfatul de amoniu, azotatul de potasiu, cazeina [2,15].
Reieşind din faptul că compoziţia biomasei de levuri ar putea fi modificată în mod semnificativ prin inter-
mediul mediului de nutriţie şi prin condiţiile de cultivare, pentru producţia înaltă de glucani este important a
optimiza aceşti factori pentru producătorii identificaţi.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
50
Scopul cercetărilor. Optimizarea mediului de cultură pentru producerea maximală a β-glucanilor la tulpina de levuri Saccharomyces cerevisiae CNMN-Y-20.
Material şi metode
Obiect de cercetare – tulpina de levuri Saccharomyces cerevisiae CNMN-Y-20 selectată ca producător de
β-glucani [4]. Medii de cultură. S-a utilizat mediul pentru însămânţare must de bere [1], mediile de fermentare Rieder, g L
-1:
30,0 – glucoză, 3,0 – (NH4)2SO4, 0,7 – MgSO4•7H2O, 0,5 – NaCl, 0,4 – Ca(NO3)2, 1,0 – KH2PO4, 10 ml auto-lizat de levuri, apă potabilă 1 L, pH – 5,0-6,0 [1], YPD, g L
-1: 20,0 – peptonă, 20,0 – glucoză, 10 ml extract de
levuri, apă potabilă 1 L, pH – 5,5 [14], mediul M-4086, g L-1
: 3,0 – (NH4)2SO4, 0,7 – MgSO4•7H2O, 0,5 – NaCl, 0,4 – Ca(NO3)2, 1,0 – KH2PO4, 20,0 – melasă, 5,0...10,0 mg – clorură de tricloracetat de zinc, 10 ml
autolizat de levuri, apă potabilă 1 L, pH – 5,0 [5].
Condiţii de fermentare. Mediul de fermentare a fost însămânţat cu 5 ml inocul (2x106 celule/ml), turbidita-
tea se măsoară ca densitate optică (DO) prin spectrofotometrie cu utilizarea lungimii de undă λ=600 nm [18].
Cultivarea tulpinii de drojdii s-a realizat în baloane Erlenmeyer cu capacitate de 1 L ce conţine 0,2 L mediu de cultură, pe agitator rotativ (200 rpm), la temperatura de 25
oC, durata de cultivare în profunzime 120 ore.
Metode de investigaţie. Biomasa levuriană s-a determinat gravimetric, prin centrifugare la 3000 g, timp de 15 min., resuspendarea depozitului de celule cu apă distilată sterilă, centrifugare repetată. Celulele au fost co-
lectate, uscate la 105oC timp de 5 ore în etuvă, apoi biomasa de levuri a fost cântărită la balanţa AQT-250
(2005) [14]. Carbohidraţii totali în biomasa de levuri s-au determinat la spectrofotometrul T60 VIS Spectropho-
tometer, la lungimea de undă 620 nm cu utilizarea reactivului antron şi a D-glucozei în calitate de standard [9]. Conţinutul de β-glucanii a fost determinat gravimetric conform procedeului prezentat în [19]. Conţinutul de
oxigen dizolvat în mediu s-a măsurat cu oximetrul portabil – Oxi-315i/SET 2B10-0011 (2008). Valorile pH-ului mediului au fost determinate cu pH-316i MeBketten WTW, Germania (2008). Optimizarea componenţei
mediului de nutriţie s-a efectuat conform metodelor de planificare matematică a experienţelor [26].
Analiza statistică a rezultatelor s-a efectuat computerizat cu calcularea erorilor standard pentru valorile re-
lative şi medii, cu ajutorul setului de programe Statistica 7.
Rezultate şi discuţii
Pentru selectărea unui mediu nutritiv adecvat pentru optimizare au fost studiate particularităţile de creşte-
re a tulpinii şi conţinutul de β-glucani la cultivare pe diferite medii nutritive. Rezultatele obţinute sunt pre-
zentate în Tabelul 1.
Tabelul 1
Conţinutul de biomasă celulară şi de β-glucani la cultivarea tulpinii
Saccharomyces cerevisiae CNMN-Y-20 pe diferite medii
Tipul de mediu Biomasa uscată, g/L β-glucani, % la B.U.
YPD 4,59±1,06 19,06±0,09
Rieder 2,09±0,03 22,36±0,57
M - 4086 3,97±0,04 12,61±0,45
După cum reflectă rezultatele obţinute în cercetările întreprinse, un nivel maximal al cantităţii de bioma-
să se obţine la cultivarea tulpinii pe mediile YPD şi M-4086. Mediul Rieder nu asigură o creştere suficientă a
biomasei celulare, dar este eficient din punctul de vedere al acumulării în peretele celular a cantităţii maxi-
male de β-glucani (22,36 ± 0,57% la B.U.). Conform rezultatelor prezentate în tabelul de mai sus, drept fac-
tori iniţiali pentru optimizarea mediului de fermentaţie au fost alese unele elemente ale mediului Rieder.
Optimizarea mediului Rieder în scopul obţinerii unei cantităţi sporite de β-glucani a fost efectuată în câ-
teva etape consecutive. Primul pas a constituit determinarea dependenţei acumulării β-glucanilor de concen-
traţia unui factor pe fonul stabilităţii celorlalte componente ale mediului. Cercetările în direcţia dată au per-
mis a stabili zonele ce redau dependenţa productivităţii culturii de concentraţia factorului.
Rezultatele experienţelor monofactoriale pentru compuşii mediului de nutriuţie au fost supuse analizei
statistice şi regresionale. După prelucrarea rezultatelor au fost evidenţiaţi pentru mediul Rieder doi factori
esenţiali: zaharoza şi acetatul de zinc ((CH3COO)2Zn), care pot influenţa semnificativ biosinteza β-glucanilor.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
51
Retultatele acestor investigaţii au stat la baza optimizării matematice a mediului de cultură pentru levura
Saccharomyces cerevisiae CNMN-Y-20 în vederea obţinerii conţinutului maximal de β-glucani.
Optimizarea mediului a fost efectuată în câteva etape consecutive: experienţa conform planului „Experi-
enţa factorială fracţionată (EFF22)”, în timpul căreia se determină direcţia varierii concentraţiei factorilor
(spre mărirea sau micşorarea concentraţiei) şi experienţa conform planului „Mişcarea pe gradient”, în timpul
căreia se alege cea mai reuşită combinaţie a factorilor esenţiali şi neesenţiali.
Evidenţierea factorilor cu efecte pozitive în procesul de acumulare a β-glucanilor a fost efectuată în câteva
trepte, utilizând metoda balanţei aleatoare. La fiecare treaptă au fost selectate efectele cu valoare esenţială.
Iniţial a fost elaborat planul experienţei factoriale fracţionate EFF 22 (Tab.2).
Tabelul 2
Informaţia iniţială despre factori
Componenta mediului Codifi-
carea
Concentraţia componentelor Pasul de
variaţie,
λ(i) Minimă
- 1
Baza
0
Maximă
+ 1
Zaharoza, g L-1
X1 30 35 40 5
Acetat de zinc, mg L-1
X2 10 15 20 5
Obţinând rezultatele experienţei EFF 22
au fost calculaţi coeficienţii de regresie după algoritmul Iets [26]
(Tab.3).
Tabelul 3
Calcularea coeficienţilor de regresie după algoritmul Iets
Planul experienţei
Rezultate experimentale
(Y) β-glucani, % la S.U.
Calculul efectelor Coeficientul
liniar de regresie, b(i) Pasul 1 Pasul 2
X1 X2
- - 20,82 Y1+Y2 = 44,91 Y1+Y2 + Y3+Y4 = 96,41 24,10 + - 24,09 Y3+Y4 = 51,5 -Y1+Y2 - Y3+Y4 = - 1,93 - 0,482 - + 28,35 Y2-Y1 = 3,27 -Y1-Y2 + Y3+Y4 = 6,59 1,647
+ + 23,15 Y4-Y3 = - 5,27 Y1-Y2 - Y3+Y4 = - 8,47 - 2,117
Pe baza coeficienţilor de regresie calculaţi a fost alcătuită ecuaţia de regresie, care are expresia:
Y= 24,10 - 0,482 X1 + 1,647 X2 – 2,117 X1X2.
În corespundere cu ecuaţia dată a fost montată experienţa „Mişcarea pe gradient”, conform planului din
Tabelul 4.
Tabelul 4
Planul experienţelor „Mişcarea pe gradient”
Valorile Factorii Biomasa
uscată,
g L-1
β-glucani,
% la B.U. Zaharoză,
X1
Acetat de zinc,
X2
Coeficient liniar de
regresie, b(i) - 0,482 1, 647
Unităţi de variaţie, λ(i) 5 5
b(i) · λ(i) - 2,41 8,235
Coeficient de
proporţionalitate, Ki 1
b(i) · λ(i)x2/ b(i) · λ(i)x1
- 3,42
Pasul pantei maximale,
H(i) 2 -3,42 ·2=-6,84
Baza experimentului 35 15

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
52
Concentraţia g L-1
mg L-1
1 35,0 15,0 1,72±0,13 23,59±0,46
2 37,0 8,16 1,73±0,01 29,47±0,52
3 39,0 1,4 1,73±0,08 29,25±0,08
4 41,0 - 5,52 1,91±0,01 29,16±0,61
5 43,0 - 12,36 1,97±0,01 25,86±0,73
Conform datelor din Tabelul 4, nivelul maximal de sinteză a β-glucanilor ( 29,47 ± 0,52% la B.U.) pentru
tulpina Saccharomyces certevisiae CNMN-Y-20 este determinat de următoarea componenţă cantitativă a me-
diului nutritiv, g L-1
: zaharoză – 37,0; (NH4)2SO4 – 3,0; MgSO4•7H2O – 0,7; NaCl – 0,5; Ca(NO3)2 – 0,4;
KH2PO4 – 1,0; acetat de zinc – 0,00816; autolizat de drojdii –10 ml; apă potabilă – 1 L, pH – 5,0-6,0, care a
fost numit convenţional R-ZZ.
Astfel, prin analiza regresională au fost determinate concentraţiile optime ale sursei de carbon (zaharo-
zei) şi ale acetatului de zinc în componenţa mediului nutritiv pentru obţinerea β-glucanilor la tulpina Saccha-
romyces certevisiae CNMN-Y-20. Cercetările efectuate au finalizat cu elaborarea unui nou mediu de nutriţie –
R-ZZ pentru cultivarea levurii Saccharomyces cerevisiae CNMN-Y-20, care permite obţinerea până la 29,47
± 0,52% β-glucani, faţă de 22,36 ± 0,57% pe mediul martor Rieder, ceea ce este cu 31,8% mai mult faţă de
mediul iniţial. Comparativ cu mediile YPD sau M - 4086 (pe care tulpina sintetizează 19,06 ± 0,09%,
respectiv, 12,61 ± 0,45% β-glucani), eficienţa mediului optimizat R-ZZ este net superioară.
Concluzii
1. La cultivarea tulpinii Saccharomyces cerevisiae CNMN-Y-20 pe mediile nutritive utilizate în prezent, mai
convenabil pentru biosinteza β-glucanilor poate fi considerat mediul Rieder, iar pentru dezvoltarea şi
multiplicarea levurii – mediile YPD şi M–4086.
2. Optimizarea compoziţiei mediului Rieder cu aplicarea metodei matematice de planificare a experienţelor
a condus la elaborarea unui mediu nutritiv eficient care asigură obţinerea cu 31,8% mai mult β-glucani
comparativ cu mediul iniţial.
Bibliografie:
1. ANGHEL, I., VASSU, T., SEGAL, B. et. al. Biologia şi tehnologia drojdiilor. Bucureşti: Editura Tehnică, 1993.
vol.3. 308 p. 2. ANGHEL, I., VOICA, C., TOMA, N. şi.al. Biologia şi tehnologia drojdiilor. Bucureşti: Editura Tehnică, 1991,
vol.2. 385 p.
3. AGUILAR-USCANGA, B., FRANCOIS, J.M. A study of the yeast cell wall composition and structure in response
to growth conditions and mode of cultivation. In: Letters in Applied Microbiology, 2003, vol.37, p.268-274. 4. Brevet de invenţie. 4048 B1, MD, C12N 1/16 Tulpină de drojdie Saccharomyces cerevisiae – sursă de β-glucani // Chiseliţa
O., Usatîi A., Taran, N., Rudic, V., Chiseliţa, N., Adajuc V. (MD). Cererea depusă 2010.02.11, BOPI nr. 6/2010.
5. Brevet de invenţie. 4086 MD, C12N 1/16, C 12 F 7/64 Mediu nutritiv pentru cultivarea tulpinii de drojdie
Saccharomyces cerevisiae CNMN-Y-20 // Chiseliţa O., Usatîi A., Chiseliţa, N., Gulea Aurelian. (MD). Cererea depu-
să 2010.09.08, BOPI nr.12/2010.
6. BELINCHÓN, M.M., GANCEDO, J.M. Glucose controls multiple processes in S. cerevisiae. through diverse combi-
nations of signaling pathways. In: FEMS Y. Res., 2007, vol.7(6), p.808–818. 7. BERTHELS, N.J., et al. Discrepancy in glucose and fructose utilisation during fermentation by S. cerevisiae wine yeast
strains. In: FEMS Y. Res., 2004, vol.4(7), p.683-689. 8. CHAUNG, H.C., HUANG, T.C., YU, J.H. et. al. Immunomodulatory effects of beta-glucans on porcine alveolar
macrophages and bone marrow haematopoietic cell-derived dendritic cells. In: Vet. Immunol. Immunopathol. 2009,
vol.131(3-4), p.147-157. 9. DEY, P., HARBORNE, J. Methods in Plant Biochemistry. Carbohydrats Academic Press, 1993, vol.2, 529 p. 10. KLIS, F.M., et al. Dynamics of cell wall structure in S. cerevisiae. In: FEMS Microbiol. Rev., 2002, vol.26, p.239-256.
11. KOGAN, G., KOCHER, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. In: Live-
stock Science, 2007, vol.109, no.1-3, p.161-165.
12. LATGÉ, J.-P. The cell wall: a carbohydrates armour for the fungal cell. In: Mol. Microbiol., 2007, vol.66(2), p.279-290. 13. LESAGE, G., BUSSEY, H. Cell Wall Assembly in S. cerevisiae. In: Microbiol. and Mol. Biol. Rev., 2006, vol.70(2),
p.317-343.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
53
14. HONG-ZHI, Liu, QIANG, Wang, YUAN-YUN, Liu et. al. Statistical Optimization of Culture Media and Conditions
for Production of Mannan by Saccharomyces cerevisiae. In: Biotechnology and Bioprocess Engineering, 2009,
vol.14, p.577-583.
15. PARROU, J., et al. Dynamic responses of reserve carbohydrates metabolism under carbon and nitrogen limitation in S.
cerevisiae. In: Yeast, 1999, vol.15, p.191-203. 16. RAFAEL, O, AFRICA, G., MARTÍN-LOMAS, M., PENADÉS, S. Preparation of multifunctional glyconanoparticles as a
platform for potential carbohydrate-based anticancer vaccines. In: Carbohydrate Research, 2007, 342(3-4), p.448-459. 17. RONDANELLI, M., OPIZZI, A., MONTEFERRARIO, F. The biological activity of beta-glucans. In: Minerva Med.
2009, 100(3), p.237-245. 18. RUIZ-GOMEZ, M.J., PRIETO-BARCIA, M.I., RISTORI-BOGAJO, E. et. al. Static and 50 Hz magnetic fields of
0.35 and 2.45 mT have no effect on the growth of Saccharomyces cerevisiae. In: Bioelectrochemistry, 2004, vol.64,
p.151–155.
19. THAMMAKITI, S., SUPHANTHARIKA, M., PHAESUWAN, T. et. al. Preparation of spent brewer's yeast β-glucans
for potential applications in the food industry. In: International Journal of Food Science&Technology, 2004, vol.39(1),
p.21-29. 20. TUBIANA, M. Prevention of cancer and the dose-effect relationship: the carcinogenic effects of ionizing radiations.
In: Cancer Radiother, 2009, vol.13(4), p.238-258. 21. VOLMAN, J.J., MENSINK, R.P., RAMAKERS, J.D., et al. Dietary (1-->3),(1-->4)-beta-D-glucans from oat activate
nuclear factor-kappaB in intestinal leukocytes and enterocytes from mice. In: Nutr Res., 2010, vol.30(1), p.40-48. 22. VOLMAN, J.J., RAMAKERS, J.D., PLAT, J. Dietary modulation of immune function by beta-glucans. In: Physiol.
Behav., 2008, vol.23/94(2), p.276-284. 23. XIAOHUA, W., LINA, Z. Physicochemical properties and antitumor activities for sulfated derivatives of lentinan.
In: Carbohydrate Research. 2009, 344(16), p.2209-2216. 24. YIANNIKOURIS, A., FRANÇOIS, J., POUGHON, L., et al. Influence of pH on Complexing of Model β-D-Glucans
with Zearalenone. In: Journal of Food Protection, 2004, 67(12), p.2741-2746. 25. YOON TAEK, J., KIM TACK, J., LEE, H. et.al. Anti-tumor metastatic activity of β-glucan purified from mutated
Saccharomyces cerevisiae. In: International Immunopharmacology, 2008, 8(1), p.36-42. 26. МАКСИМОВ, В. Многофакторный эксперимент в биологии. Москва: МГУ, 1980. 280 с.
Prezentat la 05.03.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
54
EFFECT OF BIOSTIMULATOR “FERTILEADER GOLD”
ON DEVELOPMENT OF SUNFLOWER PLANTS
Aliona GLIJIN, Maria JOIŢA-PĂCUREANU*,
Adriana ACCIU, Maria GORCEAG, Maria DUCA
University of Academy of Sciences of Moldova *National Agricultural Research and Development Institute, Fundulea, Romania
The aim of the research was to determine the influence of foliar nutrition with “Fertileader Gold” on growth indices
of sunflower plants under field conditions. Results showed that foliar application of mentioned biostimulant increased the growth rate with about 6% compared to control. The highest positive influence of the fertilizer was on the head diameter (about 28% at stage 70 days after planting), comparing with plant height. This study may help the integrated nutrient management for oil seed crops especially the sunflower, and biostimulators as “Fertileader Gold” can be beneficial to improve growth and development of treated sunflower.
Keywords: sunflower, fertilizer, biostimulator, plant height, head diameter.
EFECTUL BIOSTIMULATORULUI „FERTILEADER GOLD”
ASUPRA DEZVOLTĂRII PLANTELOR DE FLOAREA-SOARELUI
Scopul cercetării a inclus studierea influenţei nutriţiei foliare cu „Fertileader Gold” asupra parametrilor de creştere a plantelor de floarea-soarelui în condiţii de câmp. Rezultatele obţinute au demonstrat că aplicarea acestui biostimulator a indus majorarea ratei de creştere cu cca 6%. O influenţă mai mare a exercitat asupra diametrului calatidiului, care a înre-gistrat (la a 70-a zi de la cultivare) valori mai mari cu cca 28% faţă de control. Aceste rezultate pot fi utile în managemen-tul nutriţiei integrate pentru plantele de cultură oleaginoase, în special floarea-soarelui, iar biostimulatorul „Fertileader Gold” este recomandat pentru a fi utilizat la stimularea creşterii şi dezvoltării acestei culturi.
Cuvinte-cheie: floarea-soarelui, fertilizant, biostimulator, înălţimea plantei, diametrul calatidiului.
Introduction
Continuous application of chemical fertilizers causes soil health problems even if applied in balanced
proportion [26]. It has been established that organic material improves soil health and phyto-availability of
the nutrients by increasing organic matter contents in the soil and improving the soil texture [16,1].
Considering the environmental and health problems arising from chemical fertilizers usage, much attention
has been drawn to the application of biological fertilizers in agriculture. Generally, the biostimulants are the organic substances used in low concentrations, involved in physiolo-
gical processes of growth and plant development, which produce favorable effects, both quantitative and qualitative on crops while reducing losses due to transport and storage products [2].
The biostimulants and their combinations significantly affect growth processes and some growth attributes
and agronomic characters as well as yield and its components [5].
Application of fertilizers having nutrients like nitrogen, phosphorous and potash can increase sunflower
growth and yield substantially [17,21,10,8,22]. All these combine to create improved agronomic performance
and as the result, improved profitability for the grower. Plant mineral nutrition is a vital area of research to understand the mineral needs of plants, especially crops,
and increase crop yields. Plants require both macronutrients (C, H, O, N, P, S, K, Mg, and Ca) and micronu-trients (Fe, Mn, Zn, Cu, B, Mo, Cl, and Ni) in specific concentrations for their growth and development [14].
Nutritional elements can be used as foliar application. Nutritional spray on plants can decrease the delay between absorption and consumption of elements by plants that are very important for fast growth stages of plants [24].
Foliar feeding or foliar fertilization includes the application of necessary plant nutrients to the above ground, living plant parts. Its purpose is not to replace soil fertilization, but rather to supplement plant nutrient needs during short and/or critical growth stages.
Considering the economic importance and yield potential of the crop, this study was conducted to investi-gate the effects of biostimulant “Fertileader Gold” application, which is supplemented with nitrogen, boron and molybdenum on growth indices of Helianthus annuus L. under field conditions.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
55
Material and methods
Fifteen commercial sunflower hybrids (Sandrina, ITC-06, ITC-09, Procera 2, Super 5, Super 3, Super 1,
Performer, Alex, Novi Sad 1, Novi Sad 2, Novi Sad 3, Limagrain 3, Limagrain 4, Limagrain 5) were used in
our research. Seeds were obtained from National Agricultural Research and Development Institute, Fundulea,
Romania and other European Sunflower Seed Producers.
The investigations were carried out at Institute of Genetics and Plant Physiology of Academy of Sciences of
Moldova (Latitude 47° 0′ 0″ N, Longitude 28° 55′ 0″ E and Altitude 39 m above sea level). The experimental
non-irrigated field (0,5 hectare) was prepared for the “International Symposium on Broomrape (Orobanche
pp.) in Sunflower”, Chisinau, Republic of Moldova, August 25-27, 2011. The experimental design included
two sub-blocks with and without foliar nutrition (control plants).
For each hybrid was planted 4 rows with 26 m length. The distance between rows was 70 cm and between
plants was 35 cm. Seeds were sown to a depth of 5 – 8 cm. Crop was planted manually in last week of April
of 2011.
Plant height and head diameter were recorded on 80 randomly selected plants in each plot at different
development stages: 30, 40, 50, 60, 70 days after planting and at harvest.
The applied biofertilizer was “Fertileader Gold” (1%), produced by S. C. Timac AGRO România S. R. L.
This foliar biostimulant contain the patented SEACTIV base, molecular complex ofIso-pentényl adenine (IPA)
extracted from marine seaweeds [25], and supplemented with fertilizing elements such as nitrogen, boron
and molybdenum.
According to the Producer Fertileader's powerful biostimulant reduces environmental stress, increases
nutrient movement, and increases chlorophyll activity in crop to maximize yield potential. Also, this product
boost the potential of normal plant physiological processes, like root development, photosynthesis and mineral
movement within the plant.
The fertilizer was directly sprayed on the plants at vegetative stage (4-8 leaves).
Obtained data were statistically analyzed using standard methods [27].
Results and discussions
Growth and development of sunflower are a combination of many events at many different levels, from
biophysical and biochemical to tissue and organ levels. A few reports are available on the influence of organic
fertilization on the phasic development of sunflower. Hocking and Steer [13] and Blanchet et al. [7] stated
that rate of development and growth of both vegetative (leaves) and reproductive (florets and seed) organs
are very much influenced by scarcity of biofertlizer nutrition.
Variety is an important factor considering plant height. Under the present study, plant height was signifi-
cantly influenced by different genotypes of sunflower at different days after planting (DAP). Results showed
that the hybrids Alex and Novi Sad 2 were evident for highest plant height at all growth stages. The tallest
plant at 30; 40 and 50 DAP were 100,7; 133,3 and 145,0 cm respectively was obtained with Super 5 (Fig.1).
Fertilizer is the most important factor for achieving best yield of crop. Plant height was significantly in-
fluenced by “Fertileader Gold” application under the present study (Fig.1). It is evident that plant height was
the highest after treatment at different growth stages of almost varieties of sunflower. The highest plant height
at fertilized hybrids was 154,0 cm (Limagrain 4); 194,3 cm (Novi Sad 2) and 210,7 cm (Alex) at 30, 40 and
50 DAP respectively. The maximum increasing (24,8%; 15,5% and 11,7%) by fertilizer was induced at Super
5 genotype at 30, 40 and 50 DAP respectively. The hybrids Performer, Alex, Novi Sad 3 and Limagrain 4
showed no significant effects on this trait at all studied stages. Comparing dependence of fertilizer influence
on growth stages, it was demonstrated that it increase growth rate with about 6% at all three studied stages.
However, applying biostimulator “Fertileader Gold” tended to increase plant heights of sunflower cultivars
and the effect of it application was significant in most genotypes. Our research data concur with the literature
[15,19,20].
The data taken for plant height at harvest are shown in figure 1. At the control genotypes the highest plant
height were observed at two hybrids: Novi Sad 2 (200,93 cm) and Alex (200,58 cm). The minimum plant
height recorded in control was attested at sunflower genotypes Sandrina (142,48 cm) and Super 5 (142,63 cm).
Moreover, all treated plants had more height than non-treated plants. Application of “Fertileader Gold”,
increased the plant height by 12,65% (ITC-06) and 11,54% (Super 5), compared to control respectively
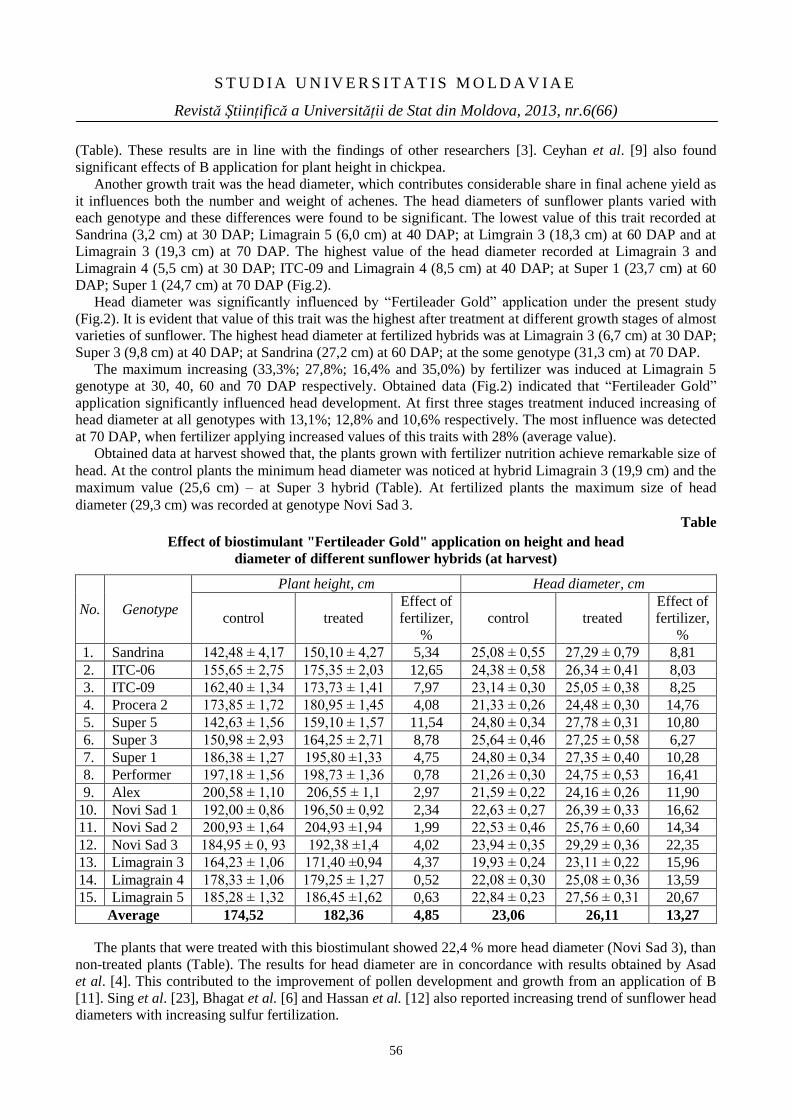
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
56
(Table). These results are in line with the findings of other researchers [3]. Ceyhan et al. [9] also found
significant effects of B application for plant height in chickpea.
Another growth trait was the head diameter, which contributes considerable share in final achene yield as
it influences both the number and weight of achenes. The head diameters of sunflower plants varied with
each genotype and these differences were found to be significant. The lowest value of this trait recorded at
Sandrina (3,2 cm) at 30 DAP; Limagrain 5 (6,0 cm) at 40 DAP; at Limgrain 3 (18,3 cm) at 60 DAP and at
Limagrain 3 (19,3 cm) at 70 DAP. The highest value of the head diameter recorded at Limagrain 3 and
Limagrain 4 (5,5 cm) at 30 DAP; ITC-09 and Limagrain 4 (8,5 cm) at 40 DAP; at Super 1 (23,7 cm) at 60
DAP; Super 1 (24,7 cm) at 70 DAP (Fig.2).
Head diameter was significantly influenced by “Fertileader Gold” application under the present study
(Fig.2). It is evident that value of this trait was the highest after treatment at different growth stages of almost
varieties of sunflower. The highest head diameter at fertilized hybrids was at Limagrain 3 (6,7 cm) at 30 DAP;
Super 3 (9,8 cm) at 40 DAP; at Sandrina (27,2 cm) at 60 DAP; at the some genotype (31,3 cm) at 70 DAP.
The maximum increasing (33,3%; 27,8%; 16,4% and 35,0%) by fertilizer was induced at Limagrain 5
genotype at 30, 40, 60 and 70 DAP respectively. Obtained data (Fig.2) indicated that “Fertileader Gold”
application significantly influenced head development. At first three stages treatment induced increasing of
head diameter at all genotypes with 13,1%; 12,8% and 10,6% respectively. The most influence was detected
at 70 DAP, when fertilizer applying increased values of this traits with 28% (average value).
Obtained data at harvest showed that, the plants grown with fertilizer nutrition achieve remarkable size of
head. At the control plants the minimum head diameter was noticed at hybrid Limagrain 3 (19,9 cm) and the
maximum value (25,6 cm) – at Super 3 hybrid (Table). At fertilized plants the maximum size of head
diameter (29,3 cm) was recorded at genotype Novi Sad 3.
Table
Effect of biostimulant "Fertileader Gold" application on height and head
diameter of different sunflower hybrids (at harvest)
No. Genotype
Plant height, cm Head diameter, cm
control treated
Effect of
fertilizer,
%
control treated
Effect of
fertilizer,
%
1. Sandrina 142,48 ± 4,17 150,10 ± 4,27 5,34 25,08 ± 0,55 27,29 ± 0,79 8,81
2. ITC-06 155,65 ± 2,75 175,35 ± 2,03 12,65 24,38 ± 0,58 26,34 ± 0,41 8,03
3. ITC-09 162,40 ± 1,34 173,73 ± 1,41 7,97 23,14 ± 0,30 25,05 ± 0,38 8,25
4. Procera 2 173,85 ± 1,72 180,95 ± 1,45 4,08 21,33 ± 0,26 24,48 ± 0,30 14,76
5. Super 5 142,63 ± 1,56 159,10 ± 1,57 11,54 24,80 ± 0,34 27,78 ± 0,31 10,80
6. Super 3 150,98 ± 2,93 164,25 ± 2,71 8,78 25,64 ± 0,46 27,25 ± 0,58 6,27
7. Super 1 186,38 ± 1,27 195,80 ±1,33 4,75 24,80 ± 0,34 27,35 ± 0,40 10,28
8. Performer 197,18 ± 1,56 198,73 ± 1,36 0,78 21,26 ± 0,30 24,75 ± 0,53 16,41
9. Alex 200,58 ± 1,10 206,55 ± 1,1 2,97 21,59 ± 0,22 24,16 ± 0,26 11,90
10. Novi Sad 1 192,00 ± 0,86 196,50 ± 0,92 2,34 22,63 ± 0,27 26,39 ± 0,33 16,62
11. Novi Sad 2 200,93 ± 1,64 204,93 ±1,94 1,99 22,53 ± 0,46 25,76 ± 0,60 14,34
12. Novi Sad 3 184,95 ± 0, 93 192,38 ±1,4 4,02 23,94 ± 0,35 29,29 ± 0,36 22,35
13. Limagrain 3 164,23 ± 1,06 171,40 ±0,94 4,37 19,93 ± 0,24 23,11 ± 0,22 15,96
14. Limagrain 4 178,33 ± 1,06 179,25 ± 1,27 0,52 22,08 ± 0,30 25,08 ± 0,36 13,59
15. Limagrain 5 185,28 ± 1,32 186,45 ±1,62 0,63 22,84 ± 0,23 27,56 ± 0,31 20,67
Average 174,52 182,36 4,85 23,06 26,11 13,27
The plants that were treated with this biostimulant showed 22,4 % more head diameter (Novi Sad 3), than
non-treated plants (Table). The results for head diameter are in concordance with results obtained by Asad
et al. [4]. This contributed to the improvement of pollen development and growth from an application of B
[11]. Sing et al. [23], Bhagat et al. [6] and Hassan et al. [12] also reported increasing trend of sunflower head
diameters with increasing sulfur fertilization.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
57
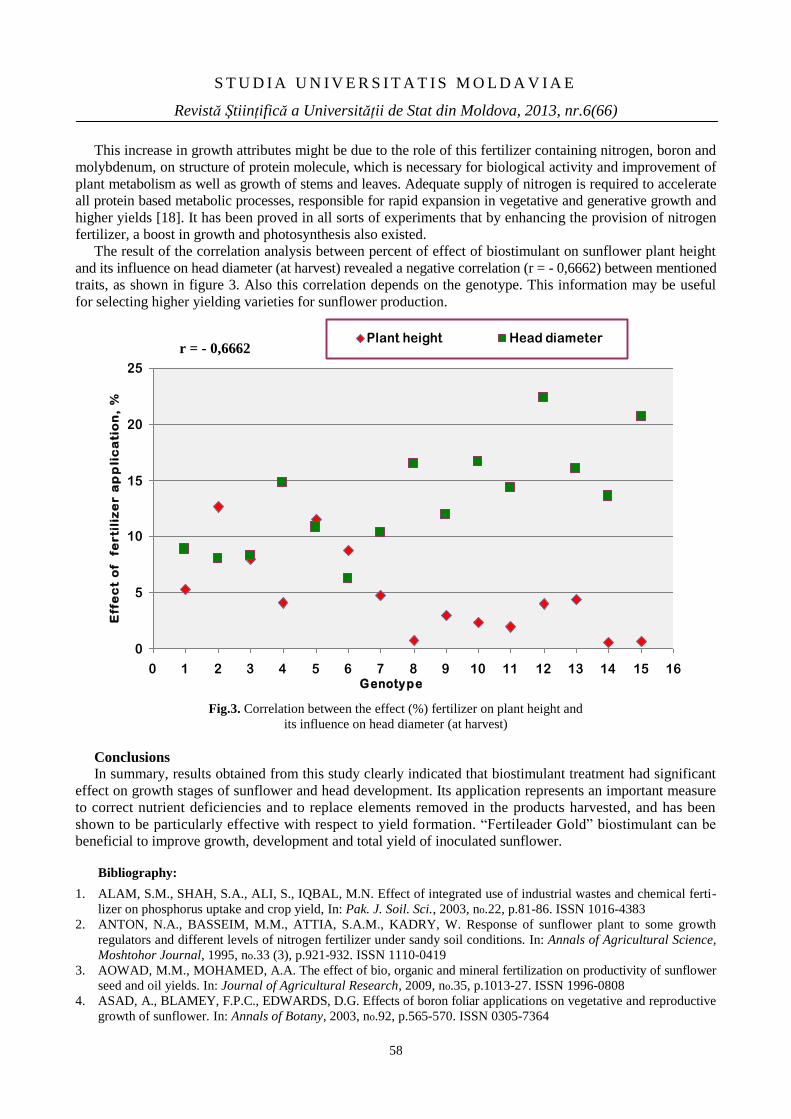
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
58
This increase in growth attributes might be due to the role of this fertilizer containing nitrogen, boron and
molybdenum, on structure of protein molecule, which is necessary for biological activity and improvement of
plant metabolism as well as growth of stems and leaves. Adequate supply of nitrogen is required to accelerate
all protein based metabolic processes, responsible for rapid expansion in vegetative and generative growth and
higher yields [18]. It has been proved in all sorts of experiments that by enhancing the provision of nitrogen
fertilizer, a boost in growth and photosynthesis also existed.
The result of the correlation analysis between percent of effect of biostimulant on sunflower plant height
and its influence on head diameter (at harvest) revealed a negative correlation (r = - 0,6662) between mentioned
traits, as shown in figure 3. Also this correlation depends on the genotype. This information may be useful
for selecting higher yielding varieties for sunflower production.
0
5
10
15
20
25
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16Genotype
Eff
ec
t o
f f
erti
liz
er a
pp
lic
ati
on
, %
Plant height Head diameter
Fig.3. Correlation between the effect (%) fertilizer on plant height and
its influence on head diameter (at harvest)
Conclusions In summary, results obtained from this study clearly indicated that biostimulant treatment had significant
effect on growth stages of sunflower and head development. Its application represents an important measure
to correct nutrient deficiencies and to replace elements removed in the products harvested, and has been
shown to be particularly effective with respect to yield formation. “Fertileader Gold” biostimulant can be
beneficial to improve growth, development and total yield of inoculated sunflower.
Bibliography:
1. ALAM, S.M., SHAH, S.A., ALI, S., IQBAL, M.N. Effect of integrated use of industrial wastes and chemical ferti-
lizer on phosphorus uptake and crop yield, In: Pak. J. Soil. Sci., 2003, no.22, p.81-86. ISSN 1016-4383
2. ANTON, N.A., BASSEIM, M.M., ATTIA, S.A.M., KADRY, W. Response of sunflower plant to some growth
regulators and different levels of nitrogen fertilizer under sandy soil conditions. In: Annals of Agricultural Science,
Moshtohor Journal, 1995, no.33 (3), p.921-932. ISSN 1110-0419
3. AOWAD, M.M., MOHAMED, A.A. The effect of bio, organic and mineral fertilization on productivity of sunflower
seed and oil yields. In: Journal of Agricultural Research, 2009, no.35, p.1013-27. ISSN 1996-0808
4. ASAD, A., BLAMEY, F.P.C., EDWARDS, D.G. Effects of boron foliar applications on vegetative and reproductive
growth of sunflower. In: Annals of Botany, 2003, no.92, p.565-570. ISSN 0305-7364
r = - 0,6662

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
59
5. BAKHT, J. Physiology, phenology and yield of sunflower (autumn) as affected by npk fertilizer and hybrids.
Pakistan Journal of Botany, 2010, no.42, p.1902-1908. ISSN 0556-3321
6. BHAGAT, G.J., HAMID, A., BONDE, S.P., GIRI, M.D., SAJIDI, M. Yield quality concentration, uptake and
availability of N, phosphorus and sulfur of sunflower. (Helianthus annus L.) as influenced by irrigation and sulphur
levels. In: Crop Research, 2005, no.30(1), p.30-36, ISSN 0264-3049
7. BLANCHET, R., GELFII, N., PUECH, J. Alimentation azotée surface foliaire et formation du tournesol. In: Agro-
chimica, 1987, no.31, p.233-243. ISSN 0002-1857
8. CECHIN, I., FUMIS, T. Effect of nitrogen supply on growth and photosynthesis of sunflower plants in the greenhouse.
Plant Science, 2004, no.166, p.1379-1380. ISSN 0168-9452
9. CEYHAN E., ÖNDER M., HARMANKAYA M., GEZGIN S. Response of chickpea cultivars to application of boron in
boron-deficient calcareous soils. In: Commun. Soil Science and Plant Analysis, 2007, no.38, p.1-19. ISSN 0010-3624
10. DE VARENNES, A., MELO-ABREU, J.P, FERREIRA, M.E. Predicting the concentration and uptake of nitrogen,
phosphorus and potassium by field grown green beans under non-limiting conditions. In: European Journal of
Agronomy, 2002, no.17, p.63-72. ISSN 1161-0301
11. DELL, B., HUANG, L.B. Physiological response of plants to low boron. In: Plant Soi, 1997, no.193, p.103-120.
ISSN 0168-9452
12. HASSAN, F.U., HAKIM, S.A., MUNAF, A., QADIR, G., AHMAD, S. Response of sunflower (Helianthus annuus L.)
to sulphur and seasonal variations. In: International Journal of Agriculture, 2007, no.9(3), p.499-503. ISSN 2227-670
13. HOCKING, P.J., STEER, B.T. Distribution of N during growth of sunflower (Helianthus annuus L.). In: Annals of
Botany. 1983, no.51, p.787-799. ISSN 0305-7364
14. HORST M. Introduction, definition, and classification of mineral nutrients. In: Mineral nutrition of higher plants.
2nd
eds. Academic Press, San Diego, 1995, cap. 1, p.1-5. ISBN 0124735428
15. HUSSEIN, M.A., EL-HATTAB, A.H., AHMED, A.K. Effect of plant spacing and nitrogen levels on morphological
characters, seed yield and quality in sunflower (Helianthus annuus L.). In: Journal of Agronomy and Crop Science,
1980, no.149, p.148-156. ISSN 1439-037
16. IBRAHIM, M., AHMAD, N., RASHID, A., SAEED, M. Use of press – mud as source of phosphorus for sustainable
agriculture. In: Proc. Symp. “Role of Phosphorus in Crop Production”, NFDC, Pakistan, 1992, p.293-301.
ISBN 13-9780070610286
17. KHO, R.M. On crop production and the balance of available resources. In: Agriculture, Ecosystems & Environment
Journal, 200, no.80, p.71-85. ISSN 0167-8809
18. LAWLOR, D.W. Carbon and nitrogen assimilation in relation to yield mechanisms are the key to understanding
production systems. In: The Journal of Experimental Botany, 2002, no.53, p.773-787. ISSN 0022-0957
19. MAHAL, S., UPPAL, H.S., MANKOTIA, B.S. Performance of spring sunflower (Helianthus annuus L.) under different
levels of soil moisture regime and nitrogen. In: Environment and Ecology, 1998, no.16, p.599-602. ISSN 0970-0420
20. POONIA, K.L. Effect of planting geometry, nitrogen and sulfur on growth and yield of sunflower (Helianthus
annuus L.). In: J. Ecol. Physiol., 2000, no.3, p.59-71. ISSN 0972-0413
21. PRASAD, P.V., SATYANARAYAMA, V., MURTHY, V.R., BOOTE, K.J. Maximizing yields in rice ground cropping
sequence through integrated nutrient management. In: Field Crops Research, 2002, no.75, p.9-21. ISSN: 0378-4290
22. SADRAS, V.O. The N: P stoichiometry of cereal, grain legume and oilseed crops. In: Field Crop Research, 2006,
no.95, p.13-29. ISSN 0378-4290
23. SING, A., SING, S.P., KATYAR, R.S., SING, P.P. Response of nitrogen and sulphur on economic yield of sunflower
(Helianthus annuus L.) under sodic soil condition. In: Indian Journal of Agricultural Sciences, 2000, no.70 (8),
p.536-537. ISSN 0019-5022
24. TAIZ, L., ZEIGER, E. Plant Physiology. Sinauer Associates Press. 5 edition. 2010. 782 p. ISBN 978-0-87-893565-9
25. YVIN, J-C. New approach of the mode of action of seaweed extracts in agriculture. Compterendu de l’Académied’
agriculture de France, 1994, no.80 (2), p.103-112. ISSN/ISBN 0989-6988
26. ZIA, M.S., MANN, R.A., ASLAM, M., KHAN, M.A. and HUSSAIN F. The role of green manuring in sustaining
rice – wheat production. In: Proc. Symp. “Integrated Plant Nutrition Management”, NDFC, Islamabad, Pakistan,
2000, p.130-149. ISBN 968-6127-18-6
27. ДОСПЕХОВ, А. Методы полевого опыта. Москва: Агропромиздат, 1985. 351 p. УДК 631.5
Acknowledgement. We express our appreciation to the National Agricultural Research and Development Institute,
Fundulea, Romania and other European Sunflower Seed Producers: “Limagrain”, “Syngenta”, “Nidera”, “Novi Sad
institute”, “Quality Crops”, “ITC Seeds” and “Procera” for providing sunflower hybrid seeds.
We would like to thank S.C. Timac AGRO România S.R.L. which provided us the Fertilizer used in our experiments.
Prezentat la 19.11.2013
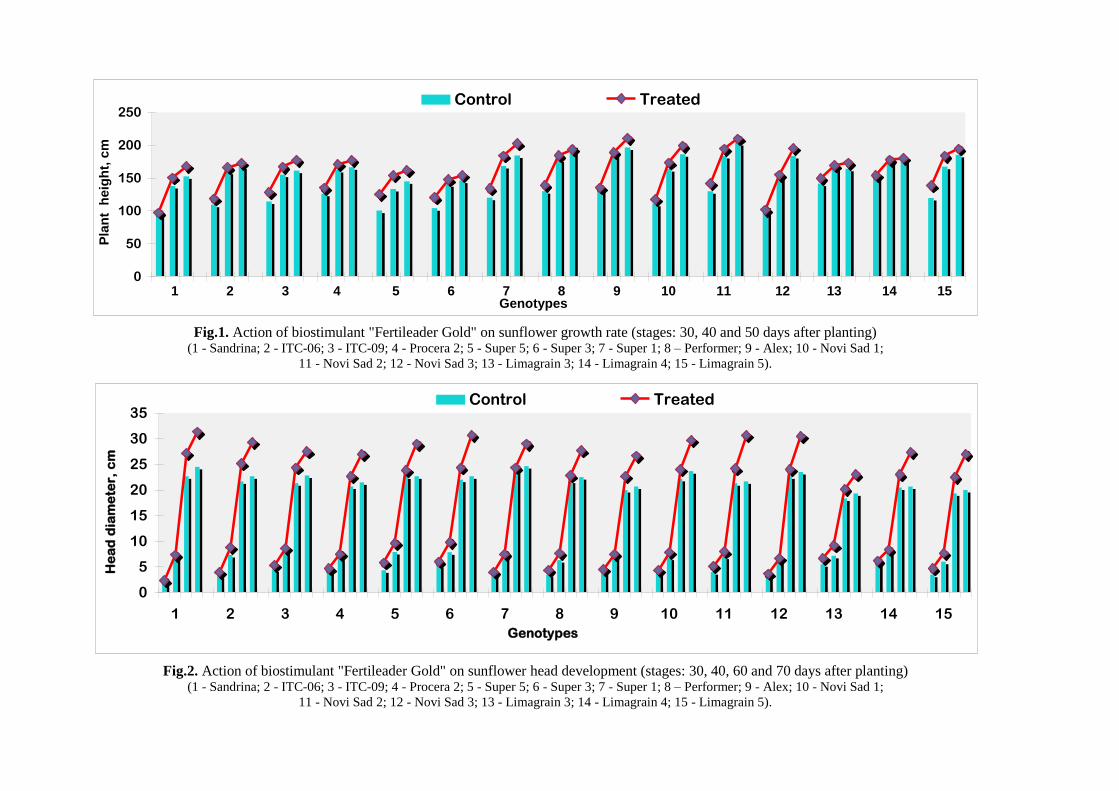
0
50
100
150
200
250
Genotypes
Pla
nt
heig
ht,
cm
Control Treated
Fig.1. Action of biostimulant "Fertileader Gold" on sunflower growth rate (stages: 30, 40 and 50 days after planting) (1 - Sandrina; 2 - ITC-06; 3 - ITC-09; 4 - Procera 2; 5 - Super 5; 6 - Super 3; 7 - Super 1; 8 – Performer; 9 - Alex; 10 - Novi Sad 1;
11 - Novi Sad 2; 12 - Novi Sad 3; 13 - Limagrain 3; 14 - Limagrain 4; 15 - Limagrain 5).
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Genotypes
He
ad
dia
me
ter,
cm
Control Treated
Fig.2. Action of biostimulant "Fertileader Gold" on sunflower head development (stages: 30, 40, 60 and 70 days after planting) (1 - Sandrina; 2 - ITC-06; 3 - ITC-09; 4 - Procera 2; 5 - Super 5; 6 - Super 3; 7 - Super 1; 8 – Performer; 9 - Alex; 10 - Novi Sad 1;
11 - Novi Sad 2; 12 - Novi Sad 3; 13 - Limagrain 3; 14 - Limagrain 4; 15 - Limagrain 5).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
60
ИЗМЕНЧИВОСТЬ И НАСЛЕДОВАНИЕ НЕКОТОРЫХ ПРИЗНАКОВ В
F3- И F4- ПОКОЛЕНИЯХ НУТА (CICER ARIETINUM L.)
Дмитрий КУРШУНЖИ, Анатолий ГАНЯ
Институт генетики и физиологии растений АН Молдовы VARIABILITATEA ŞI EREDITATEA UNOR CARACTERE ÎN GENERAŢIILE F3 ŞI F4 DE NĂUT
(CICER ARIETINUM L.) Au fost studiate ereditatea şi variabilitatea unor caractere în generaţiile F3 (11 familii) и F4 (47 de familii) din combi-
naţia hibridă ♀MDI0432×♂MDI02419. În ambele generaţii numărul familiilor segregante după pigmentaţia seminţelor era superior celor ce n-au segregat. Segregarea după caracterul tipul seminţelor s-a depistat, mai ales, în generaţia F3. S-a evidenţiat o variabilitate mai pronunţată a indicelui masa 100 de boabe în familiile pigmentate, iar ereditatea lui s-a caracterizat printr-un gradient spre forma parentală cu o valoare mai redusă a caracterului. A fost depistată ereditatea intermediară a caracterului înălţimea plantei. După indicele durata perioadei de vegetaţie, de rând cu variaţia lui în limitele formelor parentale, a fost stabilită o transgresiune negativă.
Cuvinte-cheie: ereditate, variabilitate, năut, caractere, populaţii.
VARIABILITY AND INHERITANCE OF SOME TRAITS IN GENERATIONS F3 AND F4 OF CHICKPEA
(CICER ARIETINUM L.)
Inheritance and variability of some traits have been studied in populations F3 (11 families) and F4 (47 families) of chickpea from intraspecies hybrid combination of ♀MDI0432×♂MDI02419. In both generations the number of families segregated by seed pigmentation prevailed over those with no segregation. Segregation by type of seeds trait was observed mostly in F3 generation. The higher variability of such trait as weight of 100 seeds was revealed in pigmented families and its inheritance was characterized by gradient with regard to parental form with lower value of said trait. Intermediate inheritance of plant height trait was observed. Negative transgression was noted by such parameter as duration of vegetation period along with its variability within parental forms.
Keywords: inheritance, variability, chickpea, traits, populations.
Сокращения и пояснения: с/с – светлосемянные; с/п – субпопуляция; desi-тип – пигментированные, мелкие или
среднего размера, угловатые семена; kabuli-тип – светлосемянные, среднего или крупного размера семена; gulabi-тип – гороховидные семена различного размера и пигментации.
Введение Нут (Cicer arietinum L.) является важнейшей зернобобовой культурой в мировом сельскохозяйст-
венном производстве. Его выращивают не менее чем в 37 странах Азии, Африки, Европы, Америки и Австралии на общей площади 11,55 млн. га [13]. Общее производство семян составляет 10,46 млн. тонн. Особую ценность в качестве пищевого продукта данная культура приобретает в регионах с не-достаточным увлажнением [14,21].
Несмотря на то, что история целенаправленной исследовательской работы по нуту насчитывает многие десятилетия, ряд вопросов, касающихся повышения его продуктивности и устойчивости в условиях расширения зон его возделывания, а также новых требований рынка, находятся на стадии активных поисков и новых решений. Для получения значительного спектра доступной генотипиче-ской изменчивости используют различные подходы: межвидовую гибридизацию с дикими родичами нута, индуцированный физический и химический мутагенез, внутривидовые скрещивания с привле-чением географически отдаленных генотипов. При селекции на высокую продуктивность, устойчи-вость к абиотическим и биотическим стрессам, большое значение имеет создание разнообразия исход-ного материала [24,25].
Изучению наследования пигментации семян нута посвящены ряд работ [2,4,6,7,15,17]. На осно-вании результатов расщепления в F2 поколениях сделан вывод о вовлечении от 1 до 5 генов в формиро-вание признака. Ряд авторов отмечают, что факт обнаружения разного количества генов может быть связан с использованием родительских форм c различной генетической структурой.
Ряд исследователей [5,9,11,16,18] указывают на то, что в наследовании признака типа (формы)
семян выявлен 2-генный контроль признака. Cowda C.L.L. и др. [8] отмечают широкий диапазон рас-
щепления по форме, размеру и пигментации семян в desi-kabuli комбинациях в F2 и F3 поколениях.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
61
В их работе говорится о формировании промежуточного (gulabi) типа семян, и в серии комбинаций в
количественном отношении он был на 2-м месте после desi-типа семян, а соотношение может варьиро-
вать в зависимости от комбинации скрещивания.
Salimath P.M. и Patil S.S. [20] изучали наследование и варьирование 5-ти признаков (высоты расте-
ний, количества семян в бобе, количества бобов на растении, массы 100 семян, массы семян на рас-
тении) в F3 и F4 поколениях. Авторы отмечают высокое и среднее значения наследования для массы
100 семян и количества семян в бобе соответственно и низкие значения в наследовании остальных
признаков. Варьирование (коэффициент вариации) признака высота растений – 12,2 и 11,3, массы
100 семян – 25,0 и 21,6 в F3- и F4- поколениях соответственно. Средние значения высоты растений
(31,2 и 35,5 см) и массы 100 семян (15,8 и 15,7 г) существенно не различаются между поколениями.
Генетические изучения 2-х скрещиваний [3] показали, что мелкие семена частично доминируют
над крупными и признак определяют несколько факторов – 12 и 15, для 1-ой и 2-ой гибридной ком-
бинации соответственно. В диаллельном анализе с включением 5 генотипов нута выявлено, что гены,
контролирующие мелкие семена, доминируют над генами, контролирующими крупные семена [19]. В
работе [12] исследовали 2 рекомбинантные инбредные линии, отобранные из популяций, полученных
при скрещивании kabuli-типа генотипа S95362 и 2-х образцов desi-типа – Howzat и ICC3996, которые
были испытаны в 2-х природных условиях. Показано, что размер семян контролируется 2-мя компле-
ментарными генами и линии с семенами малых размеров доминируют над крупными. Sharma и др. [22]
отмечают возможность влияния материнского эффекта на размер семян в desi×kabuli скрещиваниях;
в F2 поколении не выявлено расщепления по главным генам, а непрерывная вариация указывает на
количественный характер наследования. У нута время до цветения контролируется 2-мя главными
генами наряду с другими полигенами. Позднее цветение доминирует над ранним цветением для обоих
главных генов с взаимодействиями в большинстве случаев по аддитивно-аддитивному типу [1].
Доминирование и аддитивно-аддитивные генные взаимодействия в 1 комбинации, аддитивные и
аддитивно-аддитивные эффекты генов во 2-ой комбинации были определены для вегетационного
периода, а гены, определяющие раннее созревание, доминируют над поздним [10].
Материал и методы
В 2012 г. были изучены 11 и 47 семей F3 и F4 поколений гибридной комбинации ♀MDI0432×♂MDI02419
по ряду признаков – пигментации и типу семян, массе 100 сямян и высоте растений, периоду цветения
и созревания. В результате проведенных оценок отмечены наиболее продуктивные семьи. Опыт
осуществлялся в одном блоке, согласно методам полевого опыта [23]. Посев проводили 24-25 марта
по схеме 60×10 см. Для отборов с пигментированными семенами через каждые 7 рядов размещали
родительскую форму MDI 02432 и сорт Botna, для светлосемянных отборов – родительскую форму
MDI 02419 и сорт Ichel для сравнительного анализа и контроля продуктивности. Для признака высота
растений проанализировано 34-36 растений, для массы 100 семян использовали 6-8 повторений для
каждого варианта обоих признаков.
Результаты и обсуждение
Родительские формы MDI0432 и MDI02419 различались по ряду признаков, основные из которых
представлены в таблице 1.
Таблица 1
Различия по основным (маркерным) признакам между родительскими формами
Признаки P1 MDI 02432 P2 MDI 02419
Пигментация семян чѐрные светлые
Пигментация цветка пурпурный белый
Тип семян desi kabuli
Высота растений средние высокие
Тип куста полуштамбовый штамбовый
Период вегетации среднеспелый позднеспелый

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
62
Пигментация семян. В поколении F3 из 9-и пигментированных семей в 8-и (С41,С45,С48,С53,
С54,С55,С66,С67) отмечены расщепления с формированием чѐрного, коричневого, комбинирован-
ного и светлого пигментов. В семье С59 расщепление не отмечено, все семена были коричневыми
(рис.1). В популяции F4 из 24-х пигментированных семей в 14-ти отмечены расщепления на пигменты
предыдущего поколения, а также выявлены светло-жѐлтые, оранжевые и коричневые пигменты с
красным оттенком. У остальных 10-ти семей расщепления не обнаружены – семена коричневые с
оттенком или без него (рис.2). В обоих поколениях расщепление по пигменту разделялось на 2 или
3 класса в различных соотношениях. Во всех семьях преобладали пигментированные семена, при
наличии или отсутствии светлых семян. Таким образам, в F4-поколении увеличивается число семей,
гомозиготных по пигменту.
Рис.1. Расщепление по пигментации семян в популяции F3 нута.
Рис.2. Характер расщепления по пигментации семян в популяции F4.
Все семьи в поколении F4, полученные от светлосемянных отборов, образованных при расщепле-
нии F2-поколения, являются светлосемянными. Семьи, полученные от светлосемянных отборов, при
расщеплении в F3-поколении были светлыми или светлыми с оранжевым оттенком.
Тип семян. В пигментированных семьях F3-поколения преобладали типы desi и gulabi. Так, по
этим типам обнаружены расщепления в семьях С41, С48, С55, С59, С69, в то время как в семьях С45,
С53, С54, С66, С67 расщепления не выявлены либо они проходили по типу desi или gulabi. В F4 поко-
лении все с/с семьи, включительно с оранжевым оттенком, были gulabi- или kabuli-типа и не расщепля-
лись. Две семьи со светло-кофейными семенами характеризовались гороховидным типом, в третьей –
выявлено расщепление в соотношении 12 gulabi:1 desi. В пигментированных семьях F4-поколения
преобладал гороховидный тип, в семье С155 отмечен desi-тип, в семьях С188 и С189 – промежуточ-
ный между gulabi- и kabuli-типом. В F4-поколении преобладал промежуточный (gulabi) тип семян. В
данном случае наследование и изменчивость по пигментации и типу семян согласуются с данными,
опубликованными Cowda C.L.L. и др. [8].
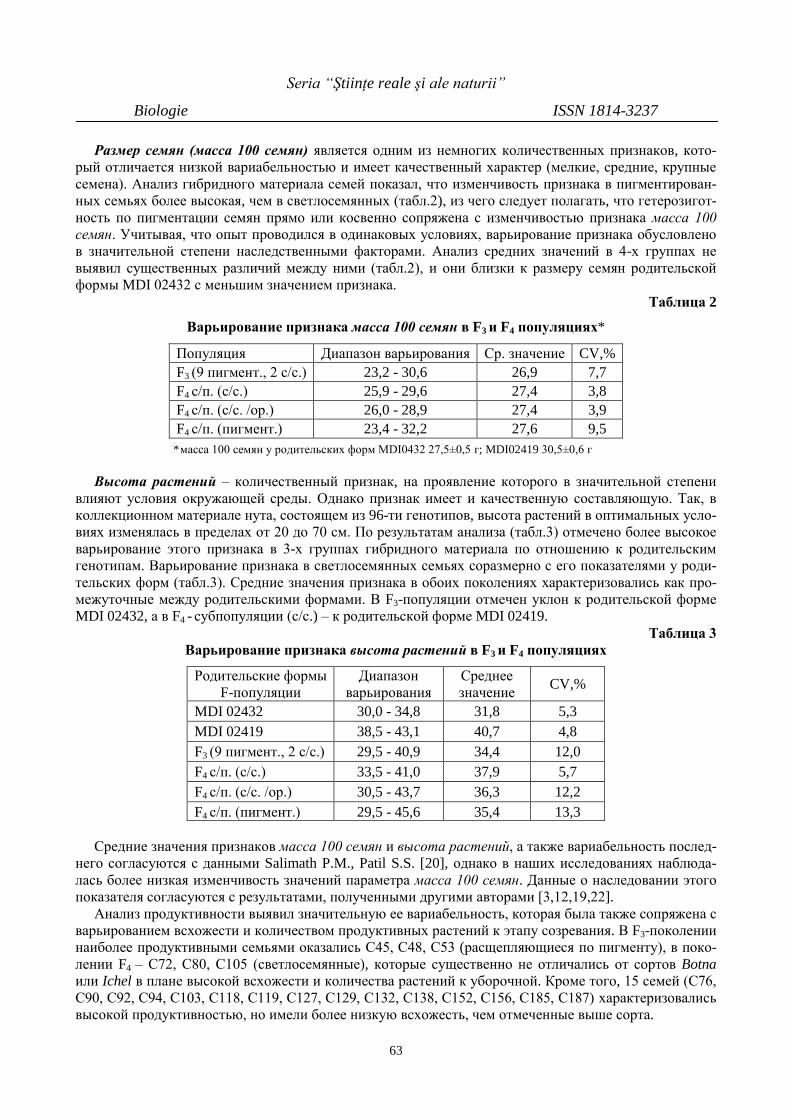
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
63
Размер семян (масса 100 семян) является одним из немногих количественных признаков, кото-
рый отличается низкой вариабельностью и имеет качественный характер (мелкие, средние, крупные
семена). Анализ гибридного материала семей показал, что изменчивость признака в пигментирован-
ных семьях более высокая, чем в светлосемянных (табл.2), из чего следует полагать, что гетерозигот-
ность по пигментации семян прямо или косвенно сопряжена с изменчивостью признака масса 100
семян. Учитывая, что опыт проводился в одинаковых условиях, варьирование признака обусловлено
в значительной степени наследственными факторами. Анализ средних значений в 4-х группах не
выявил существенных различий между ними (табл.2), и они близки к размеру семян родительской
формы MDI 02432 с меньшим значением признака.
Таблица 2
Варьирование признака масса 100 семян в F3 и F4 популяциях*
Популяция Диапазон варьирования Ср. значение CV,%
F3 (9 пигмент., 2 с/с.) 23,2 - 30,6 26,9 7,7
F4 с/п. (с/с.) 25,9 - 29,6 27,4 3,8
F4 с/п. (с/с. /ор.) 26,0 - 28,9 27,4 3,9
F4 с/п. (пигмент.) 23,4 - 32,2 27,6 9,5
* масса 100 семян у родительских форм MDI0432 27,5±0,5 г; MDI02419 30,5±0,6 г
Высота растений – количественный признак, на проявление которого в значительной степени
влияют условия окружающей среды. Однако признак имеет и качественную составляющую. Так, в
коллекционном материале нута, состоящем из 96-ти генотипов, высота растений в оптимальных усло-
виях изменялась в пределах от 20 до 70 см. По результатам анализа (табл.3) отмечено более высокое
варьирование этого признака в 3-х группах гибридного материала по отношению к родительским
генотипам. Варьирование признака в светлосемянных семьях соразмерно с его показателями у роди-
тельских форм (табл.3). Средние значения признака в обоих поколениях характеризовались как про-
межуточные между родительскими формами. В F3-популяции отмечен уклон к родительской форме
MDI 02432, а в F4 - субпопуляции (с/с.) – к родительской форме MDI 02419.
Таблица 3
Варьирование признака высота растений в F3 и F4 популяциях
Родительские формы
F-популяции
Диапазон
варьирования
Среднее
значение CV,%
MDI 02432 30,0 - 34,8 31,8 5,3
MDI 02419 38,5 - 43,1 40,7 4,8
F3 (9 пигмент., 2 с/с.) 29,5 - 40,9 34,4 12,0
F4 с/п. (с/с.) 33,5 - 41,0 37,9 5,7
F4 с/п. (с/с. /ор.) 30,5 - 43,7 36,3 12,2
F4 с/п. (пигмент.) 29,5 - 45,6 35,4 13,3
Средние значения признаков масса 100 семян и высота растений, а также вариабельность послед-
него согласуются с данными Salimath P.M., Patil S.S. [20], однако в наших исследованиях наблюда-
лась более низкая изменчивость значений параметра масса 100 семян. Данные о наследовании этого
показателя согласуются с результатами, полученными другими авторами [3,12,19,22].
Анализ продуктивности выявил значительную ее вариабельность, которая была также сопряжена с
варьированием всхожести и количеством продуктивных растений к этапу созревания. В F3-поколении
наиболее продуктивными семьями оказались С45, С48, С53 (расщепляющиеся по пигменту), в поко-
лении F4 – С72, С80, С105 (светлосемянные), которые существенно не отличались от сортов Botna
или Ichel в плане высокой всхожести и количества растений к уборочной. Кроме того, 15 семей (С76,
С90, С92, С94, С103, С118, С119, С127, С129, С132, С138, С152, С156, С185, С187) характеризовались
высокой продуктивностью, но имели более низкую всхожесть, чем отмеченные выше сорта.
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
64
Период цветения. Установлено, что популяции F3 и F4 отличались по степени вариации данного
признака. В F3-популяции и F4-с/популяции (пигментированной) отмечено перекрывание родитель-
ских форм на 1-2 дня в начале и по окончании периода цветения (табл.4). В F4-с/популяции (с/с и
с/с/ор.) обнаружено увеличение показателей признака на 5 дней (окончание цветения) по отношению
к родительской форме MDI 02419 (табл.4). Характер наследования периода цветения исследованного
материала согласуется с известным аддитивно-аддитивным типом [1].
Таблица 4
Периоды цветения и созревания родительских форм
и гибридного материала F3- и F4-популяций
Родительские формы
F-популяции
Период
цветения
Родительские формы
F-популяции
Период
созревания
MDI 02432 с 1-2 по 17-18.06 MDI 02432 13 - 14.07
MDI 02419 с 3 по 19-20.06 MDI 02419 19 - 21.07
F3 (9 пигмент., 2 с/с.) с 30.05-3.06 по 15-21.06 F3 9 - 20.07
F4 с/п. (с/с., с/с./ ор.) с 3-4 по 20-25.06 F4 с/п. (св. , св./ ор.) 18 - 22.07
F4 с/п. (пигмент.) с 30.05-4.06 по 16-22.06 F4 с/п. (пигмент.) 8 - 20.07
Период созревания. Анализ данного показателя в гибридных популяциях выявил более высокое
варьирование по сравнению с периодом цветения. В F3-популяции и F4-с/популяции (пигментирован-
ной) отмечен значительный диапазон изменчивости признака с 8-9 по 20 июля (табл.4) и несколько
более сжатый по срокам в F4-с/популяции (св. и св./ор.) – 18-22 июля. При сравнении периода созре-
вания пигментированного гибридного материала с родительскими формами определѐнный сдвиг на-
блюдался в направлении более раннего созревания. Необходимо также отметить, что продолжитель-
ность вегетационного периода у нута находится в прямой корреляции с размерами семян генотипов.
Так, в F3-популяции коэффициент корреляция (r) между указанными выше признаками составил
0,53, а в F4-с/популяции (пигментированной) – 0,66. Продолжительность вегетационного периода у
родительских форм MDI 02432 равнялась 84 дням, MDI 02419 – 89 дням, а у гибридного материала
варьировала от 76-77 дней у C181, C67, C68 и до 90 дней у C83, C101, C103. При анализе F3- и F4-поко-
лений гибридной комбинации ♀MDI0432×♂MDI02419 по изученным признакам выявлена определѐн-
ная изменчивость и гетерогенность гибридного материала.
Выводы
1. По признакам типа и пигментации семян в F3-поколении выявлена более высокая гетерогенность,
чем в F4-поколении.
2. Отмечена более значительная вариабельность признака масса 100 семян в пигментированном
гибридном материале, а характер наследования этого показателя приближался по значению к
родительской форме.
3. Выявлен промежуточный характер наследования признака высота растений в обоих поколениях.
4. Период созревания в пигментированных семьях оказался более вариабельным, чем в светлосе-
мянных, а наследование признака проходило в направлении более раннего созревания.
5. Установлена средняя корреляционная связь между признаками массa 100 семян и продолжи-
тельность вегетационного периода.
Библиография:
1. ANBESSA, Y., WARKENTIN, T., VANDERBERG, A. and BALL R. Inheritance of time to flowering in chickpea
in a short-season temperature environment. In: Heredity. 2006, vol. 97, no.1, p.55-61.
2. ARGIKAR, G.P., D'CRUZ, R.D. Inheritance of foliage cotyledon and testa colour in Сicer. In: Indian Journal of
Genetics and Plant Breeding. 1962, vol. 22, no.3, p.241-243.
3. ATHWAL, D.S., SANDRA, G.S. Inheritance of seed size and seed number per pod in Cicer. In: Indian Journal of
Genetics and Plant Breeding. 1967, vol.27, p.21-33.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
65
4. AZIZ, M.A., SHAH, S., ASGHAR, M. Inheritance studies in Pulses. I. Seed color inheritance in gram (Cicer
arietinum L.). In: Proc. Pak. Sci. Conf. 1960, vol.12, p.72-73.
5. BALASUBRAMANIAN, R. Inheritance of characters in gram. Foliage colour and rough seed-coat. In: Madras
Agricultural Journal. 1937, vol.25, p.207-208.
6. BALASUBRAMANIAN, R. Inheritance of seed-coat colour in gram. In: Indian J. Agric. Sci. 1952, vol. 21, p.239-
243.
7. BRAR, H.S., ATHWAL, D.S. Identification of gene controlling seed colour in Cicer arietinum L. In: Indian
Journal of Genetics and Plant Breeding. I970, vol.30, no.3, p.690-703.
8. COWDA, C.L.L., RAO, B.V., CHOPRA, S. Utility of desi×kabuli crosses in chickpea improvement. In: International
Chickpea Newsletter. 1987, vol.15, p.4-6.
9. D’CRUZ, R., TENDULKAR, A.V. Genetics studies in Bengal gram (Cicer arietinum L.) Double pod ×White
flower gram. II. In: Research Journal of Mahatma Phule Krishi Vidyapeeth, Rahuri, India. 1970, vol.1, p.121-127.
10. GIRASE, V.S. and DESHMUKH, R.B. Gene action for yield and its components in chickpea. In: Indian J. Genetics
and Plant Breeding. 2000, v.60, no.2, p.185-189.
11. HOSSAIN, S., FORD, R., MCNEIL, D., PITTOCK, C., PANOZZO, J. F. Development of selection tool for seed
shape and QTL analysis of seed shape with other morphological trails for selective breeding in chickpea. In:
Australian Journal Crop Science. 2010, vol.4, no.4, p.278-288.
12. HOSSAIN, S., FORD, R., MCNEIL, D., PITTOCK, C., PANOZZO J. F. Inheritance of seed size in chickpea
(Cicer arietinum L.) and identification of QTL based in 100-seed weight and seed size index. In: Australian Journal
Crop Science. 2010, vol.4, nr.2, p.126-135.
13. http://faostat.fao.org/default.aspx
14. KATERJI, N., VAN HOORN, J.W., HAMDY, A. et al. Response to soil salinity of two chickpea varieties differing
in drought tolerance. In: Agricultural Water Management. 2001, vol.50, no.2, p.83-96.
15. MEENA, H. S., KUMAR, J., YADAV, S.S. Inheritance of seed colour in chickpea (Cicer arietinum). In: Indian
Journal of Genetics and Plant Breeding. 2004, vol.62, no.2, p.151-152.
16. MORE, D. C., D'CRUZ, R. Genetics studies in Bengal gram (Cicer arietinum L.) V.D.-70-10 crossed with white
flowered white grained-II. In: Maharashtra Agric. Univ. 1976, vol.1, p.11-14.
17. NIKNEJAD, M., KHOSH-KHUI, M. The relationship of seed color inheritance to flower color in chickpeas. In:
J. Heredity. 1972, vol.63, p.155-156.
18. PHADNIS, B.A. Inheritance of evenness of seed surface in Bengal gram (Cicer arietinum L.). In: Nagpur Agric.
Coll. Mag. 1977, vol.49, p.1-6.
19. RASTOGI, K.B. Genetic analysis of seed size in chickpea. In: Ind. J. Agric. Sci. 1979, vol.49, p.42-44.
20. SALIMATH, P.M., PATIL, S.S. Genetic stady in F3 and F4 generations of chickpea. In: Indian Journal of Genetics
and Plant Breeding. 1990, vol.50, no.4, p.378-381.
21. SAXENA, N.P. Status of chickpea in the Mediterranean basin. In: Present and future prospects of chickpea crop
production and improvement in the Mediterranean countries. In: Option Mediterraneennes. 1990, vol.9, Ser.A,
p.17-24.
22. SHARMA, S., UPADHYAYA, H.D., COWDA, C.L. L., KUMAR, S., SINGH, S. Genetic analysis for seed size in
three crosses of chickpea (Cicer arietinum L.). In: Can. J. Plant Sci. 2013, vol.93, no.3, p.387-395.
23. ДОСПЕХОВ, Б.А. Методика полевого опыта. Москва: Колос, 1968. 335 c.
24. КРИВЧЕНКО, В.И., ФАДЕЕВА, Т.С., БУРЕНИН, В.И. Генетика культурных растений: зернобобовые,
овощные, бахчевые. Л: Агропромиздат, 1990. 287 с.
25. СОБОЛЕВ, Н.А. Генетика зерновых бобовых культур. Орѐл: Труд, 1972. 166 с.
Prezentat la 16.09.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
66
VARIABILITATEA GENETICĂ INTRASPECIFICĂ LA SALVIA SCLAREA L.
Rodica MARTEA, Irina DOROŞ, Alexandru MANOLE
Universitatea Academiei de Ştiinţe a Moldovei
Analiza spectrelor electroforetice obţinute în rezultatul PCR cu primerul RAPD - A3 pentru 28 genotipuri de salvie
a permis constatarea unei heterogenităţi înalte a materialului ameliorativ studiat, care reflectă structura genotipică com-
plexă a liniilor şi hibrizilor incluşi în cercetare. La nivelul genomului de salvie, cea mai înaltă frecvenţă a fost relevată
în cazul fragmentelor A3550, A3750, A3700 şi A31050. În acelaşi timp, o frecvenţă scăzută s-a atestat în cazul ampliconilor
A3400, A3650, A3800 şi A31450. În cadrul cercetării au fost evidenţiaţi 4 ampliconi (A3400, A3450, A3650 şi A31450) specifici
hibridului [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57S5 x 0 - 21S4)F8] F2 şi ampliconul A31750 specific hibridu-
lui [(V-24-86 809 S3 x 0-33 S6)F7 x Cr.p. 160 S11] F1, care ulterior pot fi utilizaţi în elaborarea markerilor moleculari.
Cuvinte-cheie: diversitate genetică, RAPD-A3, Salvia sclarea L.
INTRASPECIFIC GENETIC VARIABILITY OF SALVIA SCLAREA L.
Analysis of electrophoretic spectrum of RAPD - A3 primer for 28 genotypes of sage, has revealed a high hetero-
geneities, reflecting genotypic structure lines and hybrids included in the research. The highest frequency was observed
for A3550, A3750, A3700 şi A31050 amplicons, also, the low frequency in the case of A3400, A3650, A3800 şi A31450 amplicons.
The [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57S5 x 0 - 21S4)F8] F2 reveals 4 specific amplicons (A3400, A3450,
A3650 şi A31450) and one specific amplicons - A31750 for [(V-24-86 809 S3 x 0-33 S6)F7 x Cr.p. 160 S11] F1, which can
then be used to develop molecular markers.
Keywords: genetic diversity, RAPD-A3, Salvia sclarea L.
Introducere
Polimorfismul genetic reprezintă diferenţele genetice care stau la baza variaţiilor în cadrul speciilor. În
acest aspect, organisme cu genotipuri individuale, care se deosebesc după o serie de calităţi distincte, coexistă
în cadrul unei populaţii, în calitate de membri egali. În practică este dificil să se cunoască frecvenţa cu care
este menţinută o anumită alelă prin mutaţie. Se consideră că există un polimorfism, dacă cea mai comună
alelă pe un locus are o frecvenţă 99%.
Utilizarea markerilor moleculari în ameliorare, iniţial a proteinelor, iar mai târziu a ADN-ului, s-a dovedit a
fi utilă în estimarea variabilităţii genetice [1]. Un marker genetic este un caracter cu transmitere ereditară cu
alele uşor de cunoscut, care implică un polimorfism ce poate fi uşor detectabil [8]. Se consideră că markerii
polimorfici sunt cei mai valoroşi. Astfel, markerii moleculari au o importanţă deosebită în aprecierea compo-
nenţei ereditare a organismului şi reprezintă principala forţă motrice a ameliorării.
Analiza unui număr considerabil de loci în diferite regiuni ale genomului poate fi realizată prin analiza
RAPD (Random Amplified Polymorphic DNA), care reprezintă o varietate a reacţiei PCR, cu utilizarea de
primeri arbitrari. În general, se folosesc primeri cu 10 nucleotide şi cu un conţinut în GC de cel puţin 50%.
Optimizarea protocolului RAPD poate fi o problemă dificilă; modificarea componentelor de reacţie sau a
programului utilizat la amplificare are consecinţe imprevizibile [10].
Tehnica RAPD reprezintă o soluţie relativ simplă şi rapidă de relevare a variabilităţii genetice. RAPD este
utilizat pentru screening-ul primar al materialului iniţial de ameliorare [11] în scopul identificării de markeri
moleculari, cu perspectivă în selecţia caracterelor de interes economic [4,5]. În ultimul timp, metoda a fost
folosită cu succes şi în evidenţierea polimorfismului intraspecific.
Markerii RAPD au un caracter dominant când sunt cercetaţi în descendenţă. Aceasta rezultă din faptul că
un fragment amplificat este prezent în gel (ca o alelă dominantă A) sau absent (ca alelă recesivă a). În analiza
descendenţilor, un fragment este observat în stare homozigotă (analog cu AA), fiind amplificat atât ADN-ul
de la un părinte, cât şi de la celălalt, sau în stare heterozigotă (analog cu Aa), când este amplificat numai
ADN-ul provenit de la unul din părinţi, celălalt prezentând un polimorfism caracterizat prin absenţa fragmen-
tului. În stare heterozigotă, absenţa fragmentului de la unul din părinţi este mascată de prezenţa benzii dato-
rată fragmentului amplificat de la celălalt părinte. În cazul markerilor codominanţi (în care fiecare părinte
posedă o bandă distinctă, pe care celălalt nu o posedă), starea heterozigotă (AB) în descendenţă (prezenţa
ambelor benzi în F1) poate fi deosebită de stările homozigote (AA) şi (BB) [2]. Сercetările au demonstrat că

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
67
95% din fragmentele RAPD se comportă ca markeri dominanţi (un singur fragment amplificat distinctiv la
unul sau la ambii părinţi şi descendenţi) şi sub 5% se comportă ca markeri codominanţi [12].
În lucrarea de faţă s-a propus ca scop relevarea particularităţilor genetice pentru un şir de linii consangvi-
nizate şi hibrizi de Salvia sclarea L. din colecţia de plante medicinale şi aromatice a IGFPP, AŞM, prin apli-
carea metodei RAPD-PCR.
Material şi metode
Materialul de studiu reprezintă 13 hibrizi de Salvia sclarea L. şi formele parentale ale acestora [6]. Geno-
tipurile au fost oferite de Centrul Genetica şi Ameliorarea Plantelor Aromatice şi Medicinale de la Institutul
de Genetică, Fiziologie şi Protecţie a Plantelor, AȘM.
Extracţia ADN-ului din frunzuliţe de salvie s-a realizat după metoda standard (CTAB). Amplificarea a
fost efectuată cu primerul RAPD - A3 (5’ CCCGTCAGCA 3’) [7]. Aceasta s-a realizat întru-un mediu de 15 µl,
cu compoziţia: 50 ng ADN, 200 µlM dNTP de fiecare tip, 0,4 - 0,6 µlM primer, 1 unităţi/pe reacţie Dream
Taq DNA Polymerase, în soluţie tampon corespunzătoare şi 2,5 mM MgCl2 [7].
Pentru amplificare a fost utilizat următorul program [6]:
predenaturare – 3 minute la 95oC, urmată de 35 de cicluri, fiecare incluzând etapele:
denaturare – 45 secunde la 95oC
fixarea primerilor – 1 minut la 34-36oC
extensie – 1 minut la 72oC
extensia finală – 5 minute la 72oC.
Ampliconii s-au evidenţiat prin electroforeză în gel de agaroză 1,4% şi au fost notaţi conform primerului,
asociat cu dimensiunea ampliconului.
Rezultate şi discuţii
În procesul de obţinere a hibrizilor cu productivitate înaltă, evaluarea polimorfismului genetic prezintă o
importanţă deosebită în scopul aprecierii diversităţii genetice şi descrierii formelor parentale. Analiza RAPD
a formelor cercetate a permis constatarea heterogenităţii materialului ameliorativ după numărul de locus-uri,
dimensiune şi conţinut. Benzile polimorfe depistate la formele investigate, în baza primerilor arbitrari testaţi,
pot fi analizate în scopul elaborării unor markeri moleculari pentru identificarea acestor genotipuri [4].
Dintre cele 28 genotipuri cercetate, primerul A3 a pus în evidenţă produse ale amplificării în cazul a
18 genotipuri, 9 hibrizi şi 10 forme parentale (Tab.1).
Tabelul 1
Rezultatele amplificării cu RAPD-A3 la formele cercetate
Nr.
d/o
Primerul
A3
Nr.
ben
zi
Benzi atestate
2100
2000
1750
1600
1500
1450
1050
950
850
800
750
700
650
550
500
450
400
350
FORME PARENTALE
1. (V-24-86 809 S3 x 0-33 S6)F7 11 1 1 0 1 1 0 1 1 0 0 1 1 0 1 1 0 0 1
2. (M-55+130S4 x (K-44 x L-15)F2 x
0-47)F6 8 0 1 0 1 0 0 0 1 0 0 1 1 0 1 1 0 0 1
3. (0-57S5 x 0-21S4)F8 6 0 0 0 0 0 0 1 1 1 0 1 1 0 1 0 0 0 0
4. Cr.p. 11 S11 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0
5. Cr.p. 160 S11 6 0 0 0 0 0 0 1 0 1 0 0 1 0 1 1 0 0 1
6. (K-36 x 0-41)F20-19)F1 x L-15)B5 3 0 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0
7. (M-69 429-82 S3 x 0-40 S5)F7 9 0 0 0 1 1 1 1 0 1 0 0 1 0 0 1 1 0 1
8. (K-50)F5 x S 1122(102+113)F2 x
K-43)F4 12 0 1 0 0 1 1 1 1 1 0 0 1 1 0 1 1 1 1
9. (K-36 x 0-41)F2 x 0-19)F1 x 0-22)B4 x
L-15) 7 0 0 0 1 0 0 1 0 1 0 0 1 1 0 1 1 0 0
10. (S-1122 528 S3 x (Rubin x S-786)F1 x
(0-33 S3 x L-15)F7)F7 10 0 1 0 0 1 0 1 0 1 0 0 1 1 0 1 1 1 1

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
68
HIBRIZI
1. [S.s.Turkmen/N S7 x (K-36 x0-41) F2 x 0-19)F1 x 0-22) B4 x L-15)F8] F1
7 1 0 1 0 0 0 1 1 0 1 1 0 0 1 0 0 0 0
2. [S.s.Turkmen/N S7 x (S-1122 528 S3 x K-50) F1 x 0-48 )F6] F1
6 1 0 1 0 0 1 0 1 0 0 1 0 0 1 0 0 0 0
3. [S.s.Turkmen/N S7 x (Rubin x S1122
9S3)F1 x (0-56 x V-24) F1)] F7 5 1 0 1 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0
4.
[(V-24-86 809 S3 x 0-33 S6)F7 x
(S1122 528S3 x S.s.Tian-Shan/sud) F5 x S.s.Tian-Shan/sud)B5 ]F1
6 1 0 1 0 1 0 0 1 0 0 1 0 0 1 0 0 0 0
5. [ (V-24-86 809 S3 x 0-33 S6)F7 x Cr.p.
160 S11] F1 2 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0
6. [M-69 655 S9 x (K-36 x0-41)F20-
19)F1 x L-15)B5] F1 5 1 0 0 0 1 0 0 0 1 0 1 0 0 1 0 0 0 0
7. [Cr.p. 11 S11 x (S.s.Turkmen/N S7] F1 2 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0
8. [(K-36 x 0-41) F2 x 0-19)F1 x 0-22) B4 x L-15)] F7 x (M-44S4 x L-15) F1 x L-15)]
B6] F2 2 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0
9. [(K-50)F5 x S 1122(102+113)F2 x
K-43)F4 x (0-57S5x0-21S4)F8] F2 2 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0
Numărul cel mai mare de ampliconi, în cadrul liniilor parentale, s-a constatat la (K-50)F5 x S 1122(102+113)F2 x K-43)F4 , în cazul hibrizilor la [S.s.Turkmen/N S7 x (K-36 x 0-41) F2 x 0-19)F1 x 0-22) B4 x L-15)F8]F1. Astfel, dintre fragmentele analizate, cea mai mare frecvenţă o prezintă ampliconii A3550, A3700, A3750, A3950 şi A31050.
Un număr scăzut de fragmente de amplificare este atestat la hibrizii [(V-24-86 809 S3 x 0-33 S6)F7 x
Cr.p. 160 S11]F1, [M-69 655 S9 x (K-36 x 0-41)F20-19)F1 x L-15)B5]F1 şi [(K-50)F5 x S 1122(102+113)F2 x
K-43)F4 x (0-57 S5 x 0-21 S4)F8]F2. Aceste rezultate ar putea fi explicate prin faptul că majoritatea regiuni-lor amplificate de primerul A3 prezintă un caracter de tip codominant [3].
O particularitate deosebită a fost determinată de apariţia a două benzi specifice (800 pb): prima în cazul hibridului [S.s.Turkmen/N S7 x (K-36 x 0-41)F2 x 0-19)F1 x 0-22) B4 x L-15)F8] F1 şi a doua bandă speci-fică (400 pb) prezentă la forma parentală (K-36 x 0-41)F2 x 0-19)F1 x 0-22) B4 x L-15) F8, care ulterior pot fi utilizaţi în elaborarea markerilor moleculari.
Rezultate complexe integrale au fost relevate pentru 3 grupuri genetice, care includ hibrizii: [(V-24-86
809 S3 x 0-33 S6)F7 x Cr.p. 160 S11]F1, [M-69 655 S9 x (K-36 x 0-41)F20-19)F1 x L-15)B5]F1, [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57 S5 x 0-21 S4)F8]F2 şi formele lor parentale. Acestea au fost cercetate sub aspectele privind polimorfismul genetic, moştenirea produselor de amplificare, precum şi distanţa genetică.
Polimorfismul genetic. Profilul hibrizilor de salvie şi al formelor parentale ale acestora, obţinut cu pri-merul A3, a prezentat 18 ampliconi, care au fost puşi în evidenţă de spectre electroforetice heterogene atât după dimensiune, care se includ în limita de valori ale greutăţii moleculare 350-2100 pb, cât şi după intensi-tatea fluorescenţei, aceasta sugerând cantităţi diferite ale produsului de amplificare (Tab.1).
În baza analizei spectrelor electroforetice se constată că un nivel înalt al diversităţii genetice este atestat în cazul genotipului matern (V-24-86 809 S3 x 0-33 S6)F7, M-69 655 S9 şi patern (Cr.p. 160 S11, (K-36 x
0-41)F20-19)F1 x L-15)B5, (0-57 S5 x 0-21S4)F8) – criteriu important în formarea combinaţiilor hibride de perspectivă.
Rezultatele pentru hibridul [(V-24-86 809 S3 x 0-33 S6)F7 x Cr.p. 160 S11] F1 şi formele parentale pre-zintă 13 ampliconi cu masa moleculară de 350-2100 pb – linia maternă relevă 11 benzi, forma paternă – 6 benzi şi F1 – 2 ampliconi. A fost obţinut un fragment specific de 1750 pb.
Analiza RAPD a hibridului [M-69 655 S9 x (K-36 x 0-41)F20-19)F1 x L-15)B5] F1 şi a formelor parentale a determinat prezenţa a 11 ampliconi cu masa moleculară cuprinsă între 450 şi 2100 pb. Linia maternă pre-zintă 7 fragmente, un total de 3 benzi sunt în cazul formei paterne, hibridul – 5 ampliconi. La linia paternă lipsesc fragmentele de 160, 650, 500 şi 450 pb, prezente la genotipul matern, iar la forma F1 lipsesc compo-nenţii electroforetici de 1050 şi 700 pb, prezenţi la părinţi.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
69
Hibridul [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57 S5 x 0-21 S4)F8] F2 şi formele sale parentale
se caracterizează prin 14 componenţi polinucleotidici cu dimensiuni de la 350 la 2000 pb, linia maternă re-
levă 12 benzi, forma paternă – 6 benzi şi 2 ampliconi – hibridul analizat. La linia paternă au lipsit ampliconii
de 2000, 1500, 1450, 650, 500, 450, 400 şi 350 pb, prezenţi la genotipul matern, iar la forma hibridă au lipsit
fragmentele de 1050, 950, 850 şi 700 pb prezente la părinţi. S-au evidenţiat 4 ampliconi specifici: A3400, A3450,
A3650 şi A31450.
Moştenirea ampliconilor. Produsele amplificării ADN evidenţiate în spectrele electroforetice obţinute au
fost studiate în descendenţă, fiind evidenţiat modul de moştenire a fragmentelor identificate.
Cadrul de moştenire pentru hibridul [(V-24-86 809 S3 x 0-33 S6)F7 x Cr.p. 160 S11] F1 şi a formelor pa-
rentale (Tab.2.) evidenţiază la genotipuruile formelor parentale ale hibridului 5 ampliconi comuni de 1050,
700, 550, 500 şi 350 pb, dintre care niciunul nu este moştenit de hibrid.
Tabelul 2
Moştenirea produselor amplificării ADN la salvie, cu A3
♀ ♂ F1 [(V-24-86 809 S3 x 0-33
S6)F7 x Cr.p. 160 S11] F1
M-69 655 S9 x (K-36 x 0-41)
F20-19)F1 x L-15)B5] F1
[(K-50)F5 x S 1122
(102+113)F2 x K-43)F4 x
(0-57 S5 x 0-21 S4)F8] F2
+ + - 5 2 4
- - + 1 4 0
+ - + 1 0 0
- + + 0 0 2
+ - - 5 4 8
- + - 1 0 0
+ + + 0 1 0
Linia paternă prezintă lipsa fragmentului A3750, prezent la forma maternă, care a fost moştenit în F1. De
asemenea, în cazul liniei paterne au lipsit ampliconii de 2100, 2000, 1600 şi 1500 pb, prezenţi la genotipul
matern. În acelaşi timp, hibridul a moştenit ampliconul A3850 de la forma paternă, acest fragment nefiind
observat la linia maternă. De asemenea, se relevă ampliconul A31750, care nu este prezent la niciuna dintre
formele parentale, dar care se manifestă la forma hibrid.
În cazul hibridului [M-69 655 S9 x (K-36 x 0-41)F20-19)F1 x L-15)B5] F1 s-au identificat 2 fragmente de
1050 şi 700 pb prezente la ambele forme parentale, care nu au fost moştenite de hibrid. În acelaşi timp, frag-
mentul de 850 pb prezent la ambele forme parentale este moştenit şi de hibrid, care evidenţiază 4 fragamente
noi ce lipsesc la părinţi, şi anume: A32100, A31500, A3750 şi A3550.
Analiza RAPD a hibridului [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57 S5 x 0-21 S4)F8] F2 şi a
formelor parentale (Tab.2) arată lipsa a două fragmente – A3550 şi A3750 – la linia maternă, acestea fiind pre-
zente la forma paternă, care au fost moştenite de forma hibridă. În acelaşi timp, 4 fragmente – A3700, A3850,
A3950 şi A31050 – prezente la părinţi nu au fost remarcate la hibridul analizat.
Distanţa genetică. Rezultatele obţinute cu primerul RAPD-A3 au relevat un polimorfism pronunţat, de-
terminat atât de genotip, cât şi de modul de transmitere în descendenţă a ampliconilor. Astfel, pentru descrie-
rea complexă a formelor de salvie cercetate, au fost calculate matricele similarităţii genetice.
În continuare, în baza instrumentului Unweighted Pairwise Group Method with Arithmetic mean – UPGMA
(http://genomes.urv.cat/UPGMA/index. php) [9], matricele au fost examinate prin analiza clusteriană. UPGMA
permite construcţia dendrogramelor prin combinarea genotipurilor (grupurilor de genotipuri) în baza valori-
lor distanţei genetice.
Datele obţinute au fost utilizate la elaborarea dendrogramelor de repartiţie a genotipurilor de salvie în baza
produselor amplificării (Fig.1).
Metoda de analiză se bazează pe calcularea coeficientului Jaccard (Tanimoto).
Studiul relaţiilor intraspecifice în baza ampliconilor evidenţiaţi prin RAPD-PCR la hibrizii de salvie a
relevat prezenţa a 2 clustere de bază (Fig.1).

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
70
A B
Fig.1. Dendrograma de repartiţie a genotipurilor de salvie în baza produselor
amplificării cu primerul RAPD-A3: (A) formele parentale, (B) hibrizi:
c. (V-24-86 809 S3 x 0-33 S6)F7
e. (M-55+130S4 x (K-44 x L-15)F2 x 0-47)F6
f. (0-57S5 x 0-21S4)F8
h. (S1122 528S3 x S.s.Tian-Shan/sud)F5 x S.s.Tian-
Shan/sud)B5
i. Cr.p. 11 S 11
j. Cr.p. 160 S11
k. (K-36 x 0-41)F20-19)F1 x L-15)B5
l. (M-69 429-82 S3 x 0-40 S5)F7
m. (K-50)F5 x S 1122(102+113)F x K-43)F4
n. (K-36 x 0-41)F2 x 0-19)F1 x 0-22)B4 x L-15)
o. (S-1122 528 S3 x (Rubin x S-786)F1 x (0-33 S3 x
L-15)F7)F7
A. [S.s.Turkmen/N S7 x (K-36 x0-41) F2 x 0-19)F1 x 0-22)
B4 x L-15)F8] F1
B. [S.s.Turkmen/N S7 x (S-1122 528 S3 x K-50) F1 x
0-48 )F6] F1
D. [(V-24-86 809 S3 x 0-33 S6)F7 x (Rubin x S1122
9S3)F1 x (0-56 x V-24)F1)F7 ] F1
E. [(V-24-86 809 S3 x 0-33 S6)F7 x (S1122 528S3 x
S.s.Tian-Shan/sud) F5 x .S.s.Tian-Shan/sud)B5 ]F1
G. [M-69 655 S9 x (S-1122 528 S3 x (Rubin x S-786)F1 x
(0-33 S3 x L-15) F7) F7] F1
I. [M-69 655 S9 x (M-69 429-82 S3 x 0-40 S5)F7] F1
K. [(K-36 x 0-41) F2 x 0-19)F1 x 0-22) B4 x L-15)] F7 x
(M-44S4 x L-15) F1 x L-15)] B6] F2
L. [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57S5 x
0-21S4)F8] F2
Primul claster este caracterizat prin prezenţa următorilor hibrizi: [S.s.Turkmen/N S7 x (K-36 x 0-41) F2 x
0-19)F1 x 0-22) B4 x L-15)F8] F1, [S.s.Turkmen/N S7 x (S-1122 528 S3 x K-50) F1 x 0-48 )F6] F1 şi [(V-
24-86 809 S3 x 0-33 S6)F7 x (Rubin x S1122 9S3)F1 x (0-56 x V-24)F1)F7 ] F1. Aceştia prezintă o similari-
tate genetică de 70-80%.
Clasterul 2 include [M-69 655 S9 x (M-69 429-82 S3 x 0-40 S5)F7] F1, [(K-36 x 0-41) F2 x 0-19)F1 x
0-22) B4 x L-15)] F7 x (M-44S4 x L-15) F1 x L-15)] B6] F2 şi [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x
(0-57S5 x 0-21S4)F8] F2, care prezintă o similaritate genetică de 100% (Fig.1). În acelaşi timp, doi dintre hibrizi [(V-24-86 809 S3 x 0-33 S6)F7 x (Rubin x S1122 9S3)F1 x (0-56 x V-
24)F1)F7 ] F1 şi [(V-24-86 809 S3 x 0-33 S6)F7 x (S1122 528S3 x S.s.Tian-Shan/sud) F5 x S.s.Tian-Shan/
sud)B5 ]F1 sunt evidenţiaţi în afara grupurilor obţinute (Fig.1).
Rezultatele pentru liniile parentale de salvie au relevat prezenţa a 3 clustere (Fig.1).
Hibridul de prima generaţie [M-69 655 S9 x (S-1122 528 S3 x (Rubin x S-786)F1 x (0-33 S3 x L-15) F7)
F7] a demonstrat asemănări înalte cu formele sale parentale, ♀ V-24-86 809 S3 x 0-33 S6)F7 şi ♂ (S1122
528S3 x S.s.Tian-Shan/sud)F5 x S.s.Tian-Shan/sud)B5, care se evidenţiază prin distanţă genetică cea mai
mare – 0,909, fiind situate la cea mai mare distanţă.
Hibridul [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57 S5 x 0-21 S4)F8] F2 prezintă asemănări înalte
cu formele sale parentale, ♀ (K-50)F5 x S 1122(102+113)F2 x K-43)F4 şi ♂ (0-57S5x0-21S4)F8. Acestea pre-
zintă o distanţă genetică de 0,7 şi sunt evidenţiate în clustere diferite.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
71
Concluzii
Analiza spectrelor electroforetice ale ampliconilor RAPD-PCR la 28 genotipuri de salvie cu primerul
RAPD-A3 a permis relevarea unei heterogenităţi înalte a materialului ameliorativ studiat.
Cel mai mare număr de fragmente de amplificare este pus în evidenţă la forma parentală (K-50)F5 x S 1122
(102+113)F2 x K-43)F4 , în cazul hibrizilor la [S.s.Turkmen/N S7 x (K-36 x 0-41) F2 x 0-19)F1 x 0-22) B4 x
L-15)F8]F1.
Cel mai mic număr de fragmente este atestat la hibrizii [(V-24-86 809 S3 x 0-33 S6)F7 x Cr.p. 160 S11]F1,
[M-69 655 S9 x (K-36 x0-41)F20-19)F1 x L-15)B5]F1 şi [(K-50)F5 x S 1122(102+113)F2 x K-43)F4 x (0-57
S5 x 0-21 S4)F8]F2. Aceste rezultate ar putea fi explicate prin faptul că majoritatea regiunilor amplificate de
primerul A3 prezintă un caracter de tip codominant.
Au fost stabiliţi 4 ampliconi RAPD (A3400, A3450, A3650 şi A31450) specifici hibridului [(K-50)F5 x S 1122
(102+113)F2 x K-43)F4 x (0-57S5 x 0 - 21S4)F8] F2 şi un amplicon – A31750, specific hibridului [(V-24-86
809 S3 x 0-33 S6)F7 x Cr.p. 160 S11] F1. Aceştia pot fi utilizaţi ulterior la elaborarea markerilor moleculari.
La nivelul genomului de salvie sunt evidenţiaţi ampliconii A3550, A3750, A3700 şi A31050 cu cea mai înaltă
frecvenţă. Ampliconii A3400, A3650, A3800 şi A31450 prezintă frecvenţă scăzută, sugerând în acelaşi timp asupra
unui grad înalt de specificitate genotipică.
Bibliografie:
1. ARIF, I.A., BAKIR, M.A., KHAN, H.A. et al. A Brief Review of Molecular Techniques to Assess Plant Diversity,
Int. J. Mol. Sci. 2010, no.11, p.2079-2096.
2. CARŞAI, T. C., VLAIC, A., COȘIER, V., BÂLTEANU, V.A. Cercetări privind polimorfismul la locusul genei
leptinei în scopul aplicării selecţiei asistate de markeri genetici la taurine. ISBN 978-606-92029-6-8
3. DEVOS, K.M., GALE, M.D. The use of random amplified polymorphic DNA in wheat. In: Theor. Appl. Genet.,
1992, vol.84, p.567-572.
4. DUCA, M., CĂPĂŢANĂ, A., BARBACAR, N. Moştenirea ampliconilor ADN la diverse genotipuri de floarea-
soarelui. În: Buletinul Academiei de Ştiinţe a Moldovei. Seria „Ştiinţele vieţii”, 2006, nr.2, p.58-65.
5. DUCA, M., PORT, A., LEVIŢCHI, A. Characteristics of RAPD markers in breeding of Cucumis sativus L. In:
Roumanian Biotechnological Letters, 2008, vol.13, no.4, p.3843-3850.
6. MARTEA, R. Evaluarea diversităţii genetice la Salvia sclarea L. în baza markerilor moleculari. În: Simpozionul
ştiinţific internaţional „Agricultura modernă – realizări şi perspective” 9-11 octombrie, 2013, Chişinău. Lucrări
ştiinţifice, 2013, vol.39, p.93. ISBN 978-9975-64-250-7. [631/635+574]:378.663(478-25)(082)=135.1=111=161.1 U 56
7. MARTEA, R., PORT, A., GONCEARIUC, M., DUCA, M. Estimarea diveristăţii genetice la Salvia sclarea L.
În: Simpozionul naţional cu participare înternaţională „Biotehnologii avansate – realizări şi perspective”, 24-
25 octombrie, 2013, Chişinău. Teze, 2013, p.164. ISBN 978-9975-56-7-111-2
8. NIU, L., MANTRI, N., LI, G. et al. Array-based techniques for fingerprinting medicinal herbs. In: Chinese Medicine,
2011, nr.6, p.18.
9. REGUANT, C., BORDONS, A. Typification of Oenococcus oeni strains by multiplex RAPD-PCR and study of
population dynamics during malolactic fermentation. In: J. Appl. Microbiol., 2003, no.95, p.344-353.
10. SAEZ, R., SANZ, Y., TOLDRA, F. PCR-based fingerprinting techniques for rapid detection on animal species in
meat product. In: Meat. Science, 2004, no.66, p.659-665.
11. WELSH, J., MCCLELLAND, M. Fingerprinting genomes using PCR with arbitrary primers. In: Nucleic Acids
Res., 1990, vol.18, p.7213-7218.
12. WILLIAMS, J.G.K., KUBELIK, A.R., LIVAK, K.J., et al. DNA polymorphisms amplified by arbitrary primers are
useful as genetic markers, Nucleic Acids Res, 1990, vol.18, nr.22. p.6531-6535.
Sincere mulţumiri şi sentimente de recunoştinţă conducătorului ştiinţific Maria DUCA, profesor universitar,
academician, pentru sprijinul ştiinţific şi moral. Deosebite mulţumiri doamnelor Angela PORT, dr., conf. univ., şi
Tatiana ŞESTACOVA, drd., pentru indicaţiile şi sugestiile metodice utile în realizarea experienţelor.
Mulţumiri doamnei profesor Maria GONCEARIUC şi colectivului Centrului Genetică şi Ameliorarea Plantelor
Aromatice şi Medicinale, IGFPP, AŞM, pentru materialul biologic pus la dispoziţie.
Prezentat la 05.09.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
72
CONTROLUL GENETIC AL MEIOZEI LA PLANTE
Victoria NECHIFOR
Universitatea Academiei de Ştiinţe a Moldovei
Meioza este un proces biologic fundamental al eucariotelor cu reproducere sexuată. În această lucrare vom revizui progresele realizate în ultimii ani pentru izolarea genelor meiotice la diferite specii de plante. Înţelegerea modului în care acest proces este reglat ar putea ajuta amelioratorii de a crea noi combinaţii de alele, care ar produce hibrizi de cali-tate superioară. Cunoaşterea proceselor meiotice la plante, care în prezent nu sunt complet elucidate, în viitor va oferi informaţii importante pentru utilizarea apomixisului şi a poliploidizării sexuate.
Cuvinte-cheie: meioză, control genetic, recombinare, crossing-over, Arabidopsis thaliana, mutanţi meiotici, expresia genelor.
GENETIC CONTROL OF MEIOSIS IN PLANTS Meiosis is a fundamental biological process for sexual reproduction in eukaryotes. In this paper, we review the pro-
gress made in recent years for isolating meiotic genes at different plant species. Understanding how this process is regu-lated might help plant breeders to create new allele combinations that could produce superior hybrids. The knowledge of meiotic processes in plants, which at date are not completely understood, in future, will have important implications for the exploitation of apomixis and sexual polyploidization.
Keywords: meiosis, genetic control, recombination, crossing-over, Arabidopsis thaliana, meiotic mutants, gene expression.
Meioza (de la grec. meiosis – reducere, micşorare) reprezintă un proces-cheie în ciclul de viaţă al tuturor
eucariotelor cu reproducere sexuată, diviziune celulară în urma căreia din celulele somatice diploide se for-
mează celule sexuale haploide. Meioza determină constanţa numărului de cromozomi de-a lungul generaţii-
lor la organismele ce se înmulţesc sexuat, asigură recombinarea genetică şi măreşte variabilitatea genetică a
organismelor.
Drojdiile (Saccharomyces cerevisiae), musculiţa-de-oţet (Drosophila melanogaster), nematoda de pământ
(Caenorhabditis elegans), arabidopsisul (Arabidopsis thaliana) sunt organisme model utilizate pe larg pentru
cercetarea meiozei.
Numeroase cercetări ale meiozei la diferite specii de plante şi insecte au fost realizate la nivel citologic
[3,57,69]. În ultimul deceniu au apărut publicaţii cu privire la analiza moleculară şi funcţională a genelor
care controlează meioza la drojdii [45,58,59], şoareci [65,76], tomate şi porumb [8] etc.
S-au constatat diferite niveluri de omologie a genelor-cheie implicate în meioza la drojdii, nematoda de
pământ, drozofilă şi mamifere [33]. În acelaşi timp, au fost puse în evidenţă şi diferite mecanisme genetice
de formare a gameţilor la fiecare dintre organismele studiate [20,21]. Până în prezent a fost înregistrat un nu-
măr mare de gene care afectează premeioza, meioza şi postmeioza [35]. S-a realizat clonarea unor transcripţi
specifici din antere la tomate, au fost izolaţi şi analizaţi transcripţii a două gene necunoscute (cmg1 şi cmg2)
la porumb [9].
Astfel, s-a constatat că procesul meiotic se desfăşoară conform unui mecanism genetic strict, însă încă
puţin cunoscut, informaţia privind etapele de reglare a meiozei la plante fiind foarte fragmentară.
Un rol important în reglarea şi derularea meiozei au genele implicate în iniţierea meiozei, conjugarea şi
recombinarea cromozomilor, citochineză. Fiecare eveniment (formarea complexelor sinaptonemale, a nodu-
rilor recombinaţionale, crosing-over-ul etc.) se află sub controlul unui grup de gene specializate. Analiza
mutaţiilor, care afectează meioza la diferite specii de organisme, a permis identificarea a cel puţin şapte etape,
care se află sub control genetic:
• iniţierea meiozei sau tranziţia de la mitoză la meioză;
• conjugarea şi procesul de sinapsis al cromozomilor omologi;
• recombinarea cromozomilor omologi;
• formarea chiasmelor;
• disjuncţia cromozomilor omologi şi prima diviziune a celulei;
• iniţierea diviziunii meiotice secundare;
• a doua diviziune celulară.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
73
Studiile moleculare ale acestor gene prezintă un interes deosebit, deoarece permit a înţelege mai profund
mecanismele de iniţiere şi realizare treptată a meiozei.
Primele gene meiotice la plante au fost izolate la crin – specie din famila Liliaceae, remarcată prin dimen-
siuni relativ mari ale organelor de reproducere şi o meioză de lungă durată [13,40,41]. Ulterior, la sfârşitul
anilor '90 ai sec. XX, s-a obţinut o colecţie mare de mutanţi premeiotici şi meiotici la Arabidopsis [49]. Au
fost izolate primele gene implicate în recombinare [25,39] şi clonate primele gene meiotice [6,10,11,16,26,64].
Au fost identificate gene cu expresie diferenţiată în diverse stadii ale meiozei [22].
Au fost creaţi mutanţi privind genele asociate formării sporilor masculi – microsporogeneză [32,73], spo-
rilor femeli – macrosporogeneză [14,66] sau ambelor procese [23,26]. Astfel, forma mutantă de Arabidopsis
sporocyteless/nozzle (spl/nzz) se caracterizează prin incapacitatea celulelor arhesporale de a se diferenţia în
micro- şi megaspori [63,73]. Gena SPL codifică un factor reglator de transcripţie, esenţial în dezvoltarea spo-
rociţilor, omolog cu factorii de transcripţie MADS box. Genele AML1–5 la Arabidopsis cu un efect pleiotro-
pic accentuat asupra dezvoltării plantelor sunt similare cu gena Mei2, având un rol major în iniţierea meiozei
la drojdia Schizosaccharomyces pombe, fapt ce demonstrează că o parte din mecanismul de iniţiere a meiozei
este unul conservat [36].
În ultimii ani, identificarea şi studierea mutaţiilor meiotice s-a extins şi asupra altor specii de plante (Tab.1),
precum porumb (Zea mays) [56], orez (Oryza sativa) [53], grâu (Triticum aestivum) [27], ierburi tropicale
(Brachiaria brizantha) [46], tomate (Solanum Lycopersicum) [31], petunie (Petunia Hibrida) [21], secară
(Secale cereale) [80], proces stimulat şi de secvenţierea genomurilor plantelor superioare [24,34]. Evoluţia
ciclului celular este controlată de cicline, care interacţionează şi activează kinazele ciclin dependente (CDKs),
contribuind la realizarea tranziţiilor treptate prin intermediul ciclului de fosforilare [38].
Tabelul 1
Gene premeiotice şi meiotice (profaza I) la diferite specii de plante [22,49]
Gena Speciaa Fenotip mutant Omologia
genei
Funcţia genei Referinţe
SPL/NZZ A.t. Nediferenţierea
celulelor arhesporale
Factori de
transcripţie
MADS box
Diferenţierea celu-
lelor arhesporale în
microsporocite
Schiefthaler
et al.,1999
MEL1 O.s. Microsporocit anor-
mal, blocarea meiozei
Familia
Argonaute
Tranziţia mitoză–
meioză Nonomura
et al., 2007
AML1-5 A.t. Agregarea şi frag-
mentarea cromozo-
milor
SpMei2 Organizarea
cromatinei Kaur et al.,
2006
SWI1/
DYAD
A.t. Absenţa formării
(AEs)c, sinapsei şi
recombinării
cromozomilor
–b
Iniţierea meiozei Agashe et al.,
2002
SYN/DIF A.t. Condensarea nere-
gulată şi fragmenta-
rea cromozomilor
REC8/
RAD21
Coeziunea
cromatidelor surori
în meioză
Chelysheva
et al., 2005
MMD/DUET A.t. Degenerarea
microsporilor
–b
Derularea
microsporogenezei Yang et al.,
2003
Ph1 T.a. Dereglări în recom-
binarea omoloagă a
cromozomilor
Heterocromat
ină
Recombinarea omo-
loagă a cromozomi-
lor
Martinez-
Perez et al.,
2001
PHS1 Z.m. Conjugarea neomo-
loagă a cromozomilor
–b
Recunoaşterea omo-
loagă a cromozomilor
Pawlowski
et al., 2004
PAIR1 O.s. Absenţa sinapsei
cromozomilor şi
formarea bivalenţilor
–b
Formarea bivalen-
ţilor
Nonomura
et al., 2004a
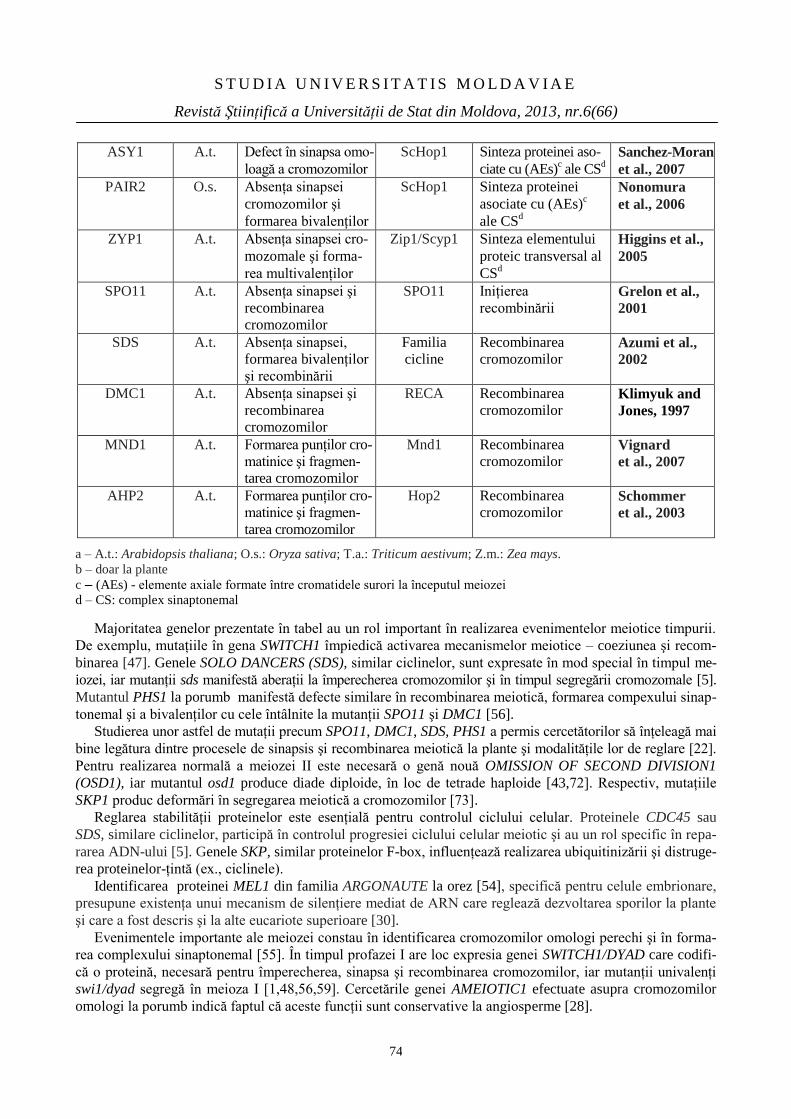
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
74
ASY1 A.t. Defect în sinapsa omo-
loagă a cromozomilor
ScHop1 Sinteza proteinei aso-
ciate cu (AEs)c ale CS
d
Sanchez-Moran
et al., 2007
PAIR2 O.s. Absenţa sinapsei
cromozomilor şi
formarea bivalenţilor
ScHop1 Sinteza proteinei
asociate cu (AEs)c
ale CSd
Nonomura
et al., 2006
ZYP1 A.t. Absenţa sinapsei cro-
mozomale şi forma-
rea multivalenţilor
Zip1/Scyp1 Sinteza elementului
proteic transversal al
CSd
Higgins et al.,
2005
SPO11 A.t. Absenţa sinapsei şi
recombinarea
cromozomilor
SPO11 Iniţierea
recombinării Grelon et al.,
2001
SDS A.t. Absenţa sinapsei,
formarea bivalenţilor
şi recombinării
Familia
cicline
Recombinarea
cromozomilor Azumi et al.,
2002
DMC1 A.t. Absenţa sinapsei şi
recombinarea
cromozomilor
RECA Recombinarea
cromozomilor Klimyuk and
Jones, 1997
MND1 A.t. Formarea punţilor cro-
matinice şi fragmen-
tarea cromozomilor
Mnd1 Recombinarea
cromozomilor Vignard
et al., 2007
AHP2 A.t. Formarea punţilor cro-
matinice şi fragmen-
tarea cromozomilor
Hop2 Recombinarea
cromozomilor Schommer
et al., 2003
a – A.t.: Arabidopsis thaliana; O.s.: Oryza sativa; T.a.: Triticum aestivum; Z.m.: Zea mays.
b – doar la plante c – (AEs) - elemente axiale formate între cromatidele surori la începutul meiozei
d – CS: complex sinaptonemal
Majoritatea genelor prezentate în tabel au un rol important în realizarea evenimentelor meiotice timpurii.
De exemplu, mutaţiile în gena SWITCH1 împiedică activarea mecanismelor meiotice – coeziunea şi recom-
binarea [47]. Genele SOLO DANCERS (SDS), similar ciclinelor, sunt expresate în mod special în timpul me-
iozei, iar mutanţii sds manifestă aberaţii la împerecherea cromozomilor şi în timpul segregării cromozomale [5].
Mutantul PHS1 la porumb manifestă defecte similare în recombinarea meiotică, formarea compexului sinap-
tonemal şi a bivalenţilor cu cele întâlnite la mutanţii SPO11 şi DMC1 [56].
Studierea unor astfel de mutaţii precum SPO11, DMC1, SDS, PHS1 a permis cercetătorilor să înţeleagă mai
bine legătura dintre procesele de sinapsis şi recombinarea meiotică la plante şi modalităţile lor de reglare [22].
Pentru realizarea normală a meiozei II este necesară o genă nouă OMISSION OF SECOND DIVISION1
(OSD1), iar mutantul osd1 produce diade diploide, în loc de tetrade haploide [43,72]. Respectiv, mutaţiile
SKP1 produc deformări în segregarea meiotică a cromozomilor [73].
Reglarea stabilităţii proteinelor este esenţială pentru controlul ciclului celular. Proteinele CDC45 sau
SDS, similare ciclinelor, participă în controlul progresiei ciclului celular meiotic şi au un rol specific în repa-
rarea ADN-ului [5]. Genele SKP, similar proteinelor F-box, influenţează realizarea ubiquitinizării şi distruge-
rea proteinelor-ţintă (ex., ciclinele).
Identificarea proteinei MEL1 din familia ARGONAUTE la orez [54], specifică pentru celule embrionare,
presupune existenţa unui mecanism de silenţiere mediat de ARN care reglează dezvoltarea sporilor la plante
şi care a fost descris şi la alte eucariote superioare [30].
Evenimentele importante ale meiozei constau în identificarea cromozomilor omologi perechi şi în forma-
rea complexului sinaptonemal [55]. În timpul profazei I are loc expresia genei SWITCH1/DYAD care codifi-
că o proteină, necesară pentru împerecherea, sinapsa şi recombinarea cromozomilor, iar mutanţii univalenţi
swi1/dyad segregă în meioza I [1,48,56,59]. Cercetările genei AMEIOTIC1 efectuate asupra cromozomilor
omologi la porumb indică faptul că aceste funcţii sunt conservative la angiosperme [28].
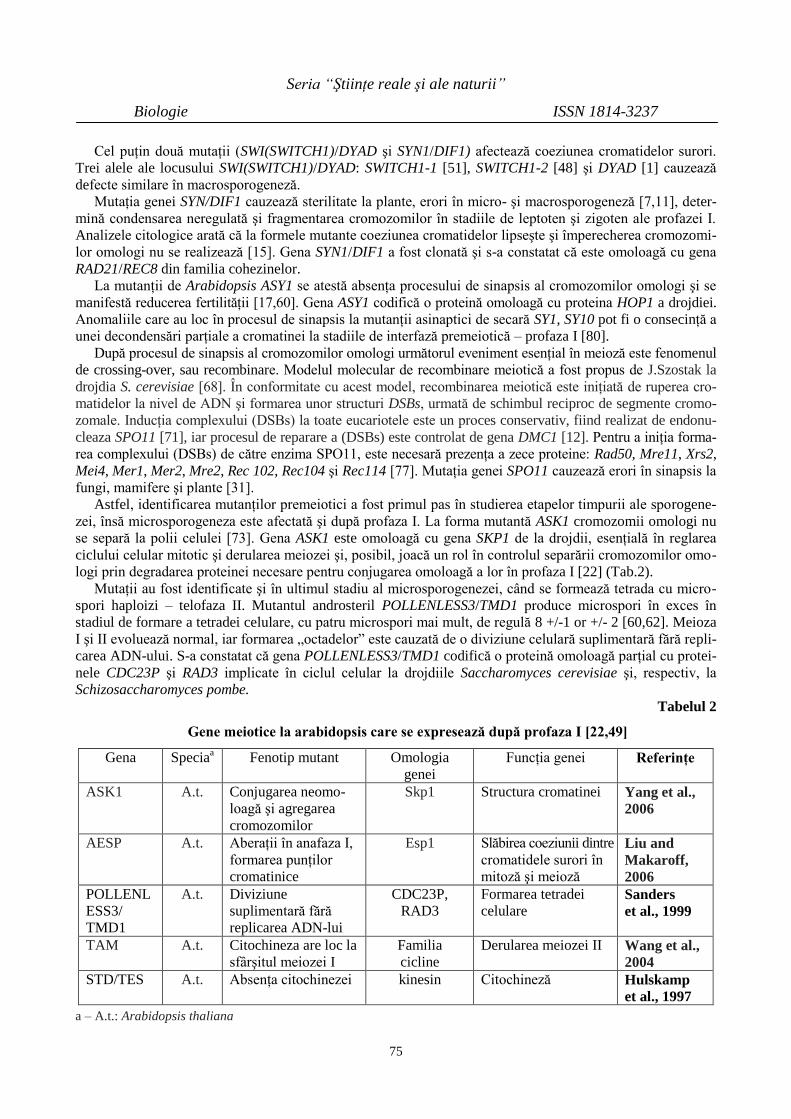
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
75
Cel puţin două mutaţii (SWI(SWITCH1)/DYAD şi SYN1/DIF1) afectează coeziunea cromatidelor surori.
Trei alele ale locusului SWI(SWITCH1)/DYAD: SWITCH1-1 [51], SWITCH1-2 [48] şi DYAD [1] cauzează
defecte similare în macrosporogeneză.
Mutaţia genei SYN/DIF1 cauzează sterilitate la plante, erori în micro- şi macrosporogeneză [7,11], deter-
mină condensarea neregulată şi fragmentarea cromozomilor în stadiile de leptoten şi zigoten ale profazei I.
Analizele citologice arată că la formele mutante coeziunea cromatidelor lipseşte şi împerecherea cromozomi-
lor omologi nu se realizează [15]. Gena SYN1/DIF1 a fost clonată şi s-a constatat că este omoloagă cu gena
RAD21/REC8 din familia cohezinelor.
La mutanţii de Arabidopsis ASY1 se atestă absenţa procesului de sinapsis al cromozomilor omologi şi se
manifestă reducerea fertilităţii [17,60]. Gena ASY1 codifică o proteină omoloagă cu proteina HOP1 a drojdiei.
Anomaliile care au loc în procesul de sinapsis la mutanţii asinaptici de secară SY1, SY10 pot fi o consecinţă a
unei decondensări parţiale a cromatinei la stadiile de interfază premeiotică – profaza I [80].
După procesul de sinapsis al cromozomilor omologi următorul eveniment esenţial în meioză este fenomenul
de crossing-over, sau recombinare. Modelul molecular de recombinare meiotică a fost propus de J.Szostak la
drojdia S. cerevisiae [68]. În conformitate cu acest model, recombinarea meiotică este iniţiată de ruperea cro-
matidelor la nivel de ADN şi formarea unor structuri DSBs, urmată de schimbul reciproc de segmente cromo-
zomale. Inducţia complexului (DSBs) la toate eucariotele este un proces conservativ, fiind realizat de endonu-
cleaza SPO11 [71], iar procesul de reparare a (DSBs) este controlat de gena DMC1 [12]. Pentru a iniţia forma-
rea complexului (DSBs) de către enzima SPO11, este necesară prezenţa a zece proteine: Rad50, Mre11, Xrs2,
Mei4, Mer1, Mer2, Mre2, Rec 102, Rec104 şi Rec114 [77]. Mutaţia genei SPO11 cauzează erori în sinapsis la
fungi, mamifere şi plante [31].
Astfel, identificarea mutanţilor premeiotici a fost primul pas în studierea etapelor timpurii ale sporogene-
zei, însă microsporogeneza este afectată şi după profaza I. La forma mutantă ASK1 cromozomii omologi nu
se separă la polii celulei [73]. Gena ASK1 este omoloagă cu gena SKP1 de la drojdii, esenţială în reglarea
ciclului celular mitotic şi derularea meiozei şi, posibil, joacă un rol în controlul separării cromozomilor omo-
logi prin degradarea proteinei necesare pentru conjugarea omoloagă a lor în profaza I [22] (Tab.2).
Mutaţii au fost identificate şi în ultimul stadiu al microsporogenezei, când se formează tetrada cu micro-
spori haploizi – telofaza II. Mutantul androsteril POLLENLESS3/TMD1 produce microspori în exces în
stadiul de formare a tetradei celulare, cu patru microspori mai mult, de regulă 8 +/-1 or +/- 2 [60,62]. Meioza
I şi II evoluează normal, iar formarea „octadelor” este cauzată de o diviziune celulară suplimentară fără repli-
carea ADN-ului. S-a constatat că gena POLLENLESS3/TMD1 codifică o proteină omoloagă parţial cu protei-
nele CDC23P şi RAD3 implicate în ciclul celular la drojdiile Saccharomyces cerevisiae şi, respectiv, la
Schizosaccharomyces pombe.
Tabelul 2
Gene meiotice la arabidopsis care se expresează după profaza I [22,49]
Gena Speciaa Fenotip mutant Omologia
genei
Funcţia genei Referinţe
ASK1 A.t. Conjugarea neomo-
loagă şi agregarea
cromozomilor
Skp1 Structura cromatinei Yang et al.,
2006
AESP A.t. Aberaţii în anafaza I,
formarea punţilor
cromatinice
Esp1 Slăbirea coeziunii dintre
cromatidele surori în
mitoză şi meioză
Liu and
Makaroff,
2006
POLLENL
ESS3/
TMD1
A.t. Diviziune
suplimentară fără
replicarea ADN-lui
CDC23P,
RAD3
Formarea tetradei
celulare Sanders
et al., 1999
TAM A.t. Citochineza are loc la
sfârşitul meiozei I
Familia
cicline
Derularea meiozei II Wang et al.,
2004
STD/TES A.t. Absenţa citochinezei kinesin Citochineză Hulskamp
et al., 1997
a – A.t.: Arabidopsis thaliana

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
76
În urma citochinezei, în meioza secundară, dintr-o diadă cu celule haploide rezultă patru celule haploide
ce formează o tetradă celulară. La mutanţii Stud (STD) / tetraspore (TES) se produc diviziuni meiotice nor-
male, însă e absentă citochineza. Polenul formelor mutante este mai mare decât cel de tip sălbatic, deoarece
microsporii sunt tetranucleaţi, iar dezvoltarea lor postmeiotică se poate desfăşură relativ normal. Nucleele
pot trece printr-o mitoză completă, ceea ce duce la formarea unor grăuncioare de polen cu un număr variabil
de nuclee. Dacă nucleii microsporilor fuzionează înainte de prima diviziune mitotică, în loc de nuclei norma-
li haploizi se formează microspori cu nuclei poliploizi [32,67].
Fig.1. Expresia genelor meiotice mutante la arabidopsis la diferite stadii ale meiozei [22].
Astfel de erori în microsporogeneză, ca realizarea citochinezei după telofaza I şi producerea diadelor cu
două celule fiice la sfârşitul primei diviziuni nucleare, se constată şi la mutanţii TAM (tardy asynchronous
meiosis). Se presupune că proteina TAM reglează desfăşurarea normală a ciclului celular, asigurând tranziţia
de la stadiul G2 al interfazei la mitoză şi a diviziunilor mitotice ale celulei-mamă a polenului [43].
Concluzii
Progresele înregistrate în studierea meiozei la Arabidopsis stimulează cercetarea genelor asociate meiozei
şi la plantele de cultură. Studierea controlului genetic al meiozei la plante va contribui la înţelegerea comple-
tă a mecanismelor micro- şi macrosporogenezei, va deschide noi perspective pentru manipularea biotehnolo-
gică a reproducerii la plante, va permite amelioratorilor să creeze noi variante de gene pentru hibrizii de cali-
tate superioară sau să folosească genele meiotice în programe ce vizează apomixisul şi poliploidizarea.
Bibliografie:
1. AGASHE, B., PRASAD, CK., SIDDIQII. Identification and analysis of DYAD: a gene required for meiotic chro-
mosome organization and female meiotic progression in Arabidopsis. In: Development 129, 2002, p.3935-3943.
2. ANDERSEN, L.K., STACK, S.M. Nodules associated with axial cores and synaptonemal complexes during
zygotene in Psilotum nudum. In: Chromosoma, 1988, vol.97, p.96-100.
3. ANDRONIC, L. Evidence of meiotic recombination in virus infected tomato. În: Buletinul AŞM, 2010, nr.2 (311), p.8-15.
4. ARMSTRONG, S.J., SANCHEZ-MORAN, E., FRANKLIN, F.C. Cytological analysis of Arabidopsis thaliana
meiotic chromosomes. In: Methods in Molecular Biology, 2009, p.131-145.
5. AZUMI, Y., LIU, D., ZHAO, D., LI, W., WANG, G., HU, Y., MA, H. Homolog interaction during meiotic pro-
phase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. In: EMBO Journal,
2002, vol.21, p.3081-3095.
6. BAI, X., PEIRSON, B.N., DONG, F., XUE, C., MAKAROFF, C.A. Isolation and characterization of SYN1, a
RAD21 - like gene essential for meiosis in Arabidopsis. In: Plant Cell, 1999, vol.11, p.417-430.
7. BAI, S., GUO, A., HE, K., LIU, D. et al: DATF: a database of Arabidopsis transcription factors. In : Bioinformatics
applications note, 2005, vol. 21 no.10, p.2568-2569.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
77
8. BARBACARU, N. Analize genomice-moleculare la plantele de cultură (tomate, porumb). În: Primul Simpozion
Naţional „Implicaţii ştiinţifice şi bioetice în analiza şi manipularea genomului”. Centrul de analize genomice şi
citogenice moleculare. Bucureşti, 2000, p.42-76.
9. BARBACARU, N., CEMORTAN, I. Izolarea unor transcripţi specifici din gametofitul mascul la porumb. În: Mate-
rialele Simpozionului ,,Ingineria genetică şi biotehnologia". Chişinău, 1998.
10. BHATT, A.M., CANALES, C. AND DICKISON, H.G. Plant meiosis: the means to 1N. In: Trends in Plant Science,
2001, vol.6, p.114-121.
11. BHATT, A.M., LISTER, C., PAGE, T., FRANSZ, P., FINDLAY, K., et al: The DIF1 gene of Arabidopsis is requi-
red for meiotic chromosome segregation and belongs to the REC8/RAD21 cohesin gene family. In: Plant J., 1999,
vol.19, p.463-472.
12. BISHOP, D.K., PARK, D., XU, L., KLECKNER, N. DMC1: a meiosis-specific yeast homolog of E. coli RecA required
for recombination, synaptonemal complex formation, and cell cycle progression. In: Cell, 1992, vol.69, p.439-456.
13. BOUCHARD, R.A., Characterization of expressed meiotic prophase repeat transcript clones of Lilium: meiosis-spe-
cific expression, relatedness, and affinities to small heat shock protein genes. In: Genome, 1990, vol.33, p.68-79.
14. BYZOVA, M.V., FRANKEN, J., AARTS, M.G.M, ALMEIDA-ENGLER, J., ENGLER, G., MARIANO, C., VAN
LOOKEREN CAMPAGNE, M., ANGENENT, G.C. Arabidopsis Sterile Apetala, a multifunctional gene regulating
inflorescence, flower, and ovule development. In: Gene Dev., 1999, vol.13, p.1002-1014.
15. CAI, X., DONG, F., EDELMANN, R.E., MAKAROFF, C.A. The Arabidopsis SYN1 cohesin protein is required for
sister chromatid arm cohesion and homologous chromosome pairing. J. In: Cell Sci., 2003, vol.116, p.2999-3007.
16. CARYL, A.P, JONES, G.H. AND FRANKLIN, F.C.H. Dissecting plant meiosis using Arabidopsis thaliana
mutants. In: Journal of Experimental Botany, 2003, vol.54, p.25-38.
17. CARYL, A.P., ARMSTRONG, S.J., JONES, G.H., FRANKLIN, F.C.H. A homologue of the yeast HOP1 gene is
inactivated in the Arabidopsis meiotic mutant asy1. In: Chromosoma, 2000, vol.109, p.62-71.
18. CHELYSHEVA, L., DIALLO, S., VEZON, D., et al. AtREC8 and AtSCC3 are essential to the monopolar orienta-
tion of the kinetochores during meiosis. In: Journal of Cell Science, 2005, vol.118, p.4621-4632.
19. CHELYSHEVA, L., GENDROT, G., VEZON, D., DOUTRIAUX, M.P., MERCIER, R., GRELON, M. Zip4/Spo22
is required for class I CO formation but not for synapsis completion in Arabidopsis thaliana. In: PLoS Genet., 2007,
vol.3, p.83.
20. CNUDDE, F., GERATS, T. Meiosis: inducing variation by reduction. In: Plant Biol., 2005, vol.7, p.321-341.
21. CNUDDE, F., HEDATALE, V., DE JONG, H., PIERSON, E.S., RAINEY, D.Y., et al. Changes in gene expression
during male meiosis in Petunia hybrida. In: Chromosome Research, 02/2006; 14(8), p.919-932.
22. CONSIGLIO, F., CONICELLA, C., MONTI, L. Highlights of meiotic genes in Arabidopsis thaliana. In: African
Journal of Biotechnology , 2003, vol.2(12), p.516-520.
23. COUTEAU, F., BELZILE, F., HORLOW, C., GRANDJEAN, O., VEZON, D., DOUTRIAUX, M.P. Random chro-
mosome segregation without meiotic arrest in both male and female meiocytes of a dmc1 mutant of Arabidopsis. In:
Plant Cell, 1999, vol.11, p.1623-1634.
24. DABING, Z., XUE, L. Cytological analysis and genetic control of rice anther development. In: Journal of Genetics
and Genomics, 2011, vol.38, p.379-390.
25. DOUTRIAUX, M.P., COUTEAU, F., BERGOUNIOUX, C., WHITE, C. Isolation and characterisation of the
RAD51 and DMC1 homologs from Arabidopsis thaliana. In: Mol. Gen. Genet., 1998, vol.257, p.283-291.
26. GRELON, M., VEZON, D., GENDROT, G., PELLETIER, G. AtSPO11-1is necessary for efficient meiotic recom-
bination in plants. In: EMBO J., 2001,vol.20, p.589-600.
27. GRIFFITHS, S., SHARP, R., FOOTE, T.N., BERTIN, I., WANOUS, M., et al. Molecular characterization of Ph1 as
a major chromosome pairing locus in polyploid wheat. In: Nature, 2006, vol.439, p.749-752.
28. Harrison, C., Alvey, E., Henderson, I. Meiosis in flowering plants and other green organisms. In: Journal of Experi-
mental Botany, 2010, vol.61, no.11, p.2863-2875.
29. HIGGINS, J.D., SANCHEZ-MORAN, E., ARMSTRONG, S.J., JONES, G.H., FRANKLIN, F.C. The Arabidopsis
synaptonemal complex protein ZYP1 is required for chromosome synapsis and normal fidelity of crossing over. In:
Genes Dev., 2005, vol.19, p.2488-2500.
30. HOLMES, R.J., COHEN, P.E. Small RNAs and RNAi pathways in meiotic prophase I. In: Chrom. Res., 2007,
vol.15, p.653-665.
31. HUANYU, QIAO. Meiotic recombination and synapsis in wild-type and asynaptic mutants of tomato (Solanum
Lycopersicum): Dissertation. Colorado State University Department of Biology Fort Collins, Colorado. 2010.
32. HULSKAMP, M., NIKESH, S.P., GRINI, P., SCHNEITZ, K., ZIMMERMANN, I., LOLLE, S.J., PRUITT, R.E.
The STUD gene is required for male-specific cytokinesis after telophase II of meiosis in Arabidopsis thaliana. In:
Dev. Biol., 1997, vol.187, p.114-124.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
78
33. HWANG, S., OH, B., KNOWLES, B., SOLTER, D., LEE, J. Expression of genes involved in mammalian meiosis during the transition from egg to embryo. In: Mol. Reprod. Dev., 2001, vol.59, p.144-158.
34. JI, Y.F. et al. Acandidate recombination modifier gene for Zea mays L. In: Genetics, 1999, vol.151, p.821-830. 35. KAUL, M.L.H. AND MURTHY, T.G.K. Mutant genes affecting higher plant meiosis. In: Theor. Appl. Genet.,
1985, vol.70, p.449-466. 36. KAUR, J., SEBASTIAN, J., SIDDIQI, I. The Arabidopsis-mei2-likegenes play a role in meiosis and vegetative-
growth in Arabidopsis. In: Plant Cell, 2006, vol.18, p.545-559. 37. KEENEY, S., GIROUX, C.N., KLECKNER, N. Meiosis-specific DNA double-strand breaks are catalyzed by
Spo11, a member of a widely conserved protein family. In: Cell., 1997, vol.88, p.375-384. 38. KEENEY, S. Mechanism and control of meiotic recombination initiation. In: Curr. Top. Dev. Biol., 2001, vol.52, p.1-53. 39. KLIMYUK, V.I., JONES, J.D. AtDMC1, the Arabidopsis homologue of the yeast DMC1 gene: characterization,
transposon-induced allelic variation and meiosis-associated expression. In: Plant J., 1997, vol.11, p.1-14. 40. KOBAYASHI, T., HOTTA, Y., TABATA, S. Isolation and characterization of a yeast gene that is homologous with
a meiosis-specific cDNA from a plant. In: Mol. Gen. Genet., 1993, vol.237, p.225-232. 41. KOBAYASHI, T., KOBAYASHI, E., SATO, S., HOTTA, Y., MIYAJIMA, N., et al. Characterization of cDNAs
induced in meiotic prophase in lily microsporocytes. In: DNA Res 1, 1994, p.15-26 . 42. LIU, Z., AND MAKAROFF, C.A. Arabidopsis separase AESP is essential for embryo development and the release
of cohesin during meiosis. In: Plant Cell., 2006, vol.18, p.1213-1225. 43. MAGNARD, J.L., YANG, M., CHEN, Y.C., LEARY, M., MCCORMICK, S. The Arabidopsis gene tardy asyn-
chronous meiosis is required for the normal pace and synchrony of cell division during male meiosis. In: Plant Phy-siology, 2001, vol.127, p.1157-1166.
44. MARTINEZ-PEREZ, E., SHAW, P., MOORE, G. The Ph1 locus is needed to ensure specific somatic and meiotic centromere association. In: Nature, 2001, vol.411, p.204-207.
45. MATA, J., LYNE, R., BURNS, G., BAHLER, J. The transcriptional program of meiosis and sporulation in fission yeast. In: Nat Genet., 2002, vol.32, p.143-147.
46. MENDES VIEIRA, D., MENDES BONATO, A.B., PAGLIARINI, M.S. & BORGES DO VALLE, C. Abnormal meiotic behavior in Brachiaria brizantha (Poaceae) leading to microspore degeneration, Caryologia: In: Internatio-nal Journal of Cytology, Cytosystematics and Cytogenetics, 2005, vol.58:4, p.396-402.
47. MERCIER, R., ARMSTRONG, S.J., HORLOW, C., JACKSON, N.P., MAKAROFF, C.A., VEZON, D.,
PELLETIER, G., JONES, G.H., FRANKLIN, F.C.H. The meiotic protein SWI1 is required for axial element formation
and recombination initiation in Arabidopsis. In: Development, 2003, vol.130, p.3309-3318.
48. MERCIER, R., VEZON, D., BULLIER, E., MOTAMAYOR, J.C., SELLIER, A., LEFÈVRE, F., PELLETIER, G.,
HORLOW, C. Switch1 (SWI1): a novel protein required for the establishment of sister chromatid cohesion and for
bivalent formation at meiosis. In: Gene Dev., 2001, vol.15, p.1859-1871.
49. MERCIER, R. AND GRELON, M. Meiosis in plants: Ten years of gene discovery. Cytogenet. In: Genome Res.,
2008, vol.120, p.281-290. 50. MOSES, M.J. A new chromosome structure is revealed. In: Chromosoma, 2006, vol.115, p.152-154.
51. MOTAMAYOR, J.C., VEZON, D., BAJON, C., SAUVANET, A., GRANDJEAN, O., MARCHAND, M.,
BECHTOLD, N., PELLETIER, G., HORLOW, C. Switch (swi1), an Arabidopsis thaliana mutant affected in the fe-
male meiotic switch. In: Sex. Plant Reprod., 2000, vol.12, p.209-218.
52. NONOMURA, K., NAKANO, M., EIGUCHI, M., SUZUKI, T., KURATA, N. PAIR2 is essential for homologous
chromosome synapsis in rice meiosis I. In: Cell Science, 2006, vol.119, p.217–225.
53. NONOMURA, K., NAKANO, M., FUKUDA, T., EIGUCHI, M., MIYAO, A., et al. The novel gene homologous-
pairing aberration in rice meiosis1of rice encodes a putative coiled-coil protein required for homologous chromo-
some pairing in meiosis. In: Plant Cell, 2004, vol.16, p.1008-1020.
54. NONOMURA, K.I., MOROHOSHI, A., NAKANO, M., EIGUCHI, M., MIYAO, A., et al. A germ cell specific
gene of the ARGONAUTE family is essential for the progression of premeiotic mitosis and meiosis during sporoge-
nesis in rice. In: Plant Cell, 2007, vol.19, p.2583-2594.
55. PAGE, S.L., HAWLEY, R.S. The genetics and molecular biology of the synaptonemal complex. In: Annual Review
Cell Dev. Biol., 2004, vol.20, p.525-558.
56. PAWLOWSKI, W.P., GOLUBOVSKAYA, I.N., TIMOFEJEVA, L., MEELEY, R.B., SHERIDAN, W.F., CANDE,
W.Z. Coordination of meiotic recombination, pairing and synapsis by PHS1. In: Science, 2004, vol.303, p.89-92.
57. PEARSON, W.R., WOOD, T., ZHANG, Z., MILLER, W. Comparison of DNA sequences with protein sequences.
In: Genomics, 1997, vol.46, p.24- 36.
58. PRIMIG, M., WILLIAMS, R.M., WINZELER, E.A. et al. The core meiotic transcriptome in budding yeasts. In:
Nat. Genet., 2000, vol.26, p.415-423.
59. RABITSCH, K.P., TOTH, A., GALOVA, M. et al. A screen for genes required for meiosis and spore formation
based on whole-genome expression. In: Current Biol., 2001, vol.11, p.1001-1009.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
79
60. ROSS, K.J., FRANSZ, P., ARMSTRONG, S.J., VIZIR, I., MULLIGAN, B., FRANKLIN, F.C., AND JONES, G.H.
Cytological characterization of four meiotic mutants of Arabidopsis isolated from T-DNA-transformed lines. In:
Chromosome Res., 1997, vol.5, p.551-559.
61. SANCHEZ-MORAN, E., MERCIER, R., HIGGINS, J.D., ARMSTRONG, S.J., JONES, G.H., FRANKLIN, F.C. A
strategy to investigate the plant meiotic proteome. In: Cytogenet Genome Res., 2005, vol.109, p.181-189.
62. SANDERS, P.M., BUI, A.Q., WETERINGS, K., MCINTIRE, K.N., HSU, Y.C., LEE, P.Y., TRUONG, M.T.,
BEALS, T.P., and GOLDBERG, R.B. Anther developmental defects in Arabidopsis thaliana male-sterile mutants.
In: Sex. Plant Reprod., 1999, vol.11, p.297-322.
63. SCHIEFTHALER, U., BALASUBRAMANIAN, S., SIEBER, P., CHEVALIER, D., WISMAN, E., SCHNEITZ
Molecular analysis of NOZZLE, a gene involved in pattern formation and early sporogenesis during sex organ deve-
lopment in Arabidopsis thaliana. In: Proc. Natl. Acad. Sci. USA, 1999, vol.96, p.11664-11669.
64. SCHOMMER, C., BEVEN, A., LAWRENSON, T., SHAW, P. AND SABLOWSKI R. AHP2 is required for biva-
lent formation and for segregation of homologous chromosomes in Arabidopsis meiosis. In: Plant Journal, 2003,
vol.36, p.1-11.
65. SCHULTZ, N., HAMRA, F.K., GARBERS, D.L. A multitude of genes expressed solely in meiotic or postmeiotic sper-
matogenic cells offers a myriad of contraceptive targets. In: Proc. Natl. Acad. Sci USA, 2003, vol.100, p.12201-12206.
66. SIDDIQI, I., GANESH, G., GROSSNIKLAUS, U. The DYAD gene is required for progression through female
meiosis in Arabidopsis. In: Development, 2000, vol.127, p.197-207.
67. SPIELMAN, M., PREUSS, D., LI, F.L., BROWNE, W.E., SCOTT, R.J., DICKINSON, H.G. TETRASPORE is
required for male meiotic cytokinesis in Arabidopsis thaliana. In: Development, 1997, vol.124, p.2645-2657.
68. SZOSTAK, J.W., ORR-WEAVER, T.L., ROTHSTEIN, R.J., STAHL, F.W. The double-strand-break repair model
for recombination. In: Cell, 1983, vol.33, p.25-35.
69. TODERAŞ, L. The effect of extreme temperature on Triticale male gametophyte. În: Materialele Congresului Naţi-
onal, Iaşi, România, 1992, p.111-112.
70. VIGNARD, J., SIWIEC, T., CHELYSHEVA, L., VRIELYNCK, N., GONORD, F., et al. The interplay of RecA-
related proteins and the MND1-HOP2 complex during meiosis in Arabidopsis thaliana. In: PLoS Genet., 2007,
vol.3, p.1894-1906.
71. VILLENEUVE, A.M., HILLERS, K.J. Whence meiosis? In: Cell, 2001, vol.106, p.647-650.
72. WANG, Y., MAGNARD, J.L., MCCORMICK, S., YANG, M. Progression through meiosis I and meiosis II in Ara-
bidopsis anthers is regulated by an A-type cyclin predominately expressed in prophase I. In: Plant Physiology, 2004,
vol.136, p.4127-4135.
73. YANG, M., HU, Y., LODHI, M., MCCOMBIE, W.R., MA, H. The Arabidopsis SKP1-LIKE1 gene is essential for
male meiosis and may control homologue separation. In: Proc. Natl. Acad. Sci. USA, 1999, vol.96, p.11416-11421.
74. YANG, X., MAKAROFFCA, M.A.H. The Arabidopsis MALE MEIOCYTE DEATH1 gene encodes aPHD-finger
protein that is required for male meiosis. In: Plant Cell, 2003, vol. 15, p.1281-1295 .
75. YANG, X., TIMOFEJEVA, L., MA, H., MAKAROFF, C.A. The Arabidopsis SKP1 homolog ASK1 controls meio-
tic chromosome remodeling and release of chromatin from the nuclear membrane and nucleolus. In: Cell Science,
2006, vol.119, p.3754-3763.
76. YU, Z.R., GUO, R., GE, Y.H. et al. Gene expression profiles in different stages of mouse spermatogenic cells
during spermatogenesis. In: Biol Reprod., 2003, vol.69, p.37-47.
77. БАБЫНИН, Э.В. Mолекулярный механизм гомологичной рекомбинации в мейозе: происхождение и биоло-
гическое значение. В: Цитология, 2007, том 49, №3.
78. БОГДАНОВ, Ю. Белковые механизмы мейоза. B: Природа, 2008, №3.
79. БОГДАНОВ, Ю.Ф. Сходство доменной организации белков у филогенетически далеких организмов как
основа консерватизма мейоза. B: Онтогенез, 2004, том 35, № 6, с.415-423.
80. МИХАЙЛОВА, Е. Молекулярно-цитогенетический анализ ключевых событий мейоза у ржи Secale cereale L:
Автореферат диcс. на соис. уч. ст. докт. биол. наук. Санкт Петербург 2011.
Prezentat la 20.08.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
80
EVALUAREA REZISTENŢEI UNOR HIBRIZI AVANSAŢI
DE TRITICUM AESTIVUM L. LA AGENŢII PUTREGAIULUI DE RĂDĂCINĂ
ÎN CONDIŢII DE TEREN ŞI DE LABORATOR
Elena SAŞCO
Institutul de Genetică şi Fiziologie a Plantelor al AŞM
Hibrizii reciproci (F5) de Triticum aestivum în condiţii de teren au manifestat rezistenţa avansată a ambilor părinţi
sau la nivelul acestora pentru agenţii de putregai de rădăcină şi de fuzarioza spicului. Părinţii – linia L 101, genotipul
Basarabeanca, şi 2 din hibrizii ce deţin în calitate de genitor citoplasmatic linia L 101 au format clusterul cu manifestare
avansată a maladiilor. Alţi 4 hibrizi, inclusiv 3 cu citoplasma Basarabeanca, au suportat atac diminuat al maladiilor. În
condiţii de temperatură suboptimă de laborator metaboliţii Fusarium oxysporum var. orthoceras au produs atât inhibiţia,
cât şi stimularea creşterii rădăciniţei şi tulpiniţei. Metaboliţii Helminthosporium avenae au fost doar inhibitivi pentru
populaţiile de grâu în studiu. Analiza bifactorială a relaţiilor genotip de grâu x filtrat de cultură al tulpinelor patogene
constată că rolul preponderent aparţine genotipului de grâu pentru creşterea rădăciniţei (47,26%) în cazul F. oxysporum
var. orthoceras şi pentru creşterea tulpiniţei (50,47%) în cazul H. avenae. Cuvinte-cheie: Triticum aestivum L., hibrizi reciproci avansaţi, putregai de rădăcină, fuzarioza spicului, metaboliţi
Fusarium şi Helminthosporium.
EVALUATION OF RESISTANCE ADVANCEDS HYBRIDS TRITICUM AESTIVUM TO THE AGENTS
ROOT ROT IN THE FIELD AND LABORATORY CONDITIONS Reciprocal hybrids (F5) of Triticum aestivum under field conditions manifested advanced resistance both of the
parents or at their level for root rot agents and fusariose ear of wheat. Genitors – L 101 line, genotype Basarabeanca and 2 hybrids, which have genitor cytoplasmic line L 101 have formed cluster of advanced manifestation of diseases. Other 4 hybrids, including 3 in the cytoplasm Basarabeanca have supported diminished attack of diseases. In laboratory conditions temperature sub-optimal metabolites of strains F. oxysporum var. orthoceras have caused inhibition and stimulation of rootlet and stem growth. The metabolites H. avenae were only inhibitive for studied wheat populations. Bifactorial analysis of the relations genotype wheat x filtrate cultural of pathogenic strains detects that preponderent role belongs to genotype of wheat for rootlet growth (47.26%) in case of F. oxysporum var. orthoceras and for the stem growth (50.47%) in case of H. avenae.
Keywords: Triticum aestivum L., reciprocal hybrids advanced, root rot, fusariose ear of wheat, metabolites Fusarium and Helminthosporium.
Introducere
Fungii speciilor Fusarium şi Helminthosporium, agenţi ai putregaiului de rădăcină (PR), prezintă o con-
strângere majoră a producerii de cereale la nivel mondial. Pierderile de recoltă, evaluate la 3-50%, sunt ra-
portate în multe regiuni ale lumii, în special în zonele de producere de cereale din Asia de Vest, Asia de Est,
Africa de Nord, Australia şi Canada. Maladia este deosebit de severă în regiunile de stepă [1,5,7,11]. Pentru
diferite culturi cerealiere şi zone ecologo-geografice în complexul de agenţi patogeni au fost depistate anumite
legităţi ale raportului speciilor Fusaium/ Helminthosporium [11]. Bipolaris sorokiniana, agent al PR complex al
grâului în regiunea Morghan, nord-vestul Iranului, a demonstrat patogenitate superioară tulpinilor F. culmorum,
F. pseudograminearum, F. crookwellence şi F. udum [5]. În condiţiile Republicii Moldova incidenţa fungilor
Fusarium şi Helminthosporium, implicaţi în dezvoltarea PR la grâul de toamnă, constituie 31,9-83,4 şi 16,6-
27,0%, respectiv. Dintre speciile Fusarium, incidenţă mai înaltă a prezentat F. oxysporum var. orthoceras [2].
Cu o penetrare mai profundă a miceliului în ţesutul gazdei este caracterizat fungul H. avenae, ce-l face mai re-
zistent la diverse condiţii, inclusiv la tratament fungicid. Drechslera avenae (Sin. Pyrenophora chaetomioides,
H. avenae), specializată la diferite specii de cereale şi ierburi, în condiţii de câmp a afectat toate genotipurile de
ovăz. Patogenitatea fiind determinată de capacitatea avansată a fungului de a produce metaboliţi secundari [8].
Dificultăţile legate de ameliorarea rezistenţei grâului la putrezirea rădăcinii sunt determinate, în special,
de lipsa genelor eficiente de rezistenţă în genofondul acestei culturi şi de caracterul facultativ, cu specializare
largă, al parazitismului agenţilor cauzali. Cu toate acestea, sunt menţionate diverse soiuri de grâu care mani-
festă rezistenţă la această maladie în anumite regiuni [3,6,12].

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
81
Liniile cu normă de reacţie largă la schimbările factorilor de mediu au avantaje ecologice pentru rezistenţa
la factorii biotici. Screening-ul speciilor Triticum, Aegilops şi Triticale pentru putrezirea radiculară produsă
de Cochliobolus sativus a pus în evidenţă existenţa variabilităţii înalte a rezistenţei plantelor [12]. În Germania,
soiuri cu rezistenţă avansată la putrezirea radiculară au fost obţinute din populaţii hibride cu rezistenţă şi tole-
ranţă medie, fenomen determinat de morfogeneza transgresivă [9]. Forme transgresive valoroase se atestă şi la
hibridarea soiurilor de grâu sensibile la fuzarioză [4]. În legătură cu aceasta, în cadrul proceselor de ameliorare
a păioaselor s-ar putea renunţa la excluderea formelor sensibile la etapele timpurii de selecţie recurentă [10].
Prin colaborare integrată de CIMMYT, Mexic şi Turcia au obţinut progrese substanţiale în dezvoltarea germo-
plasmei rezistente: din 16 surse de grâu de toamnă identificate, 4 (de origine din Turcia) s-au dovedit a fi bine
adaptate [7]. Evaluarea preventivă a liniilor nou-create la utilizarea metaboliţilor sau suspensiilor de spori ale
agenţilor patogeni se efectuează implicând tulpinile patogene larg răspândite în localitatea dată [7,10,11].
Scopul prezentelor cercetări constă în evaluarea rezistenţei/sensibilităţii populaţiilor de hibrizi reciproci
de grâu de toamnă la atacul de putregai de rădăcină şi fuzarioza spicului în condiţii de teren şi la acţiunea
metaboliţilor Fusarium oxysporum var. orthoceras şi Helminthospoium avenae în condiţii de temperatură
suboptimă de laborator.
Material şi metode Au fost cercetate genotipurile de grâu de toamnă: genitorii L 101, Basarabeanca, şi hibrizii reciproci avan-
saţi (F5) – L 101 x Basarabeanca (58, 59 şi 68) şi Basarabeanca x L 101 (60, 61 şi 62) în condiţii de teren şi
de laborator.
Rezistenţa plantelor la agenţii putregaiului de rădăcină în condiţii de câmp a fost apreciată la etapa de
coacere lapte-ceară în baza suprafeţei brunificate sau necrotizate de la baza tulpinii, în scara de 5 trepte:
0 – imune (I); 0,1 – puternic rezistente (RR); 1 – rezistente (R); 2 – sensibile (S); 3 – puternic sensibile (SS).
Fuzarioza spicului a fost evaluată la scara de 6 trepte: 0, 1, 2, 3, 4, 5, în care „0” prezintă plante imune, iar
„5” – plante atacate (>75%).
În condiţii de temperatură suboptimă de laborator (7,0-7,5oC) în calitate de factor selectiv au fost utilizate
filtratele de cultură (FC) a câte 3 tulpini F. oxysporum var. orthoceras şi H. avenae. Sensibilitatea genotipu-
rilor de grâu a fost determinată la tratarea seminţelor cu FC (18 ore). Capacitatea fitotoxică a fost înregistrată
în baza variabilităţii parametrilor cu receptivitate înaltă la patogenii facultativi – lungimea rădăciniţei şi a tul-
piniţei plantulelor de grâu. Indicii cantitativi au fost apreciaţi la 10 zile de creştere în condiţii de frigider.
Analiza datelor a fost efectuată în pachetul de soft STATISTICA 7.
Rezultate şi discuţii
În condiţiile anului 2011 atacul de putregai de rădăcină la populaţiile cercetate a variat în limitele 0,95-
2,15 grade. Genotipul-părinte Basarabeanca a manifestat sensibilitate sporită (SS), gradul de atac (2,15 grade)
fiind veridic distins în raport cu genitorul L 101 şi cei 6 hibrizi. Linia L 101 a prezentat sensibilitate medie (SM).
Hibrizii reciproci au manifestat rezistenţă avansată celui mai sensibil din genitori – soiului Basarabeanca.
Doar unul din hibrizi – L 101 x Basarabeanca–68 a prezentat sensibilitate la nivelul genitorului matern L 101,
fiind totodată veridic distins de cel patern – Basarabeanca (Tab.1).
Tabelul 1
Atacul grâului de toamnă de putregai de rădăcină şi de fuzarioza spicului în condiţii de câmp (grad)
Nr. crt. Varianta Putregai de rădăcină Fuzarioza spicului
1 Linia L 101 1,77±0,11V 0,91±0,29
2 Basarabeanca 2,15±0,12* 1,01±0,35
3 F5 L 101 x Basarabeanca–58 1,00±0,14*V
0,21±004*V
4 F5 L 101 x Basarabeanca–59 1,18±0,13*V 1,27±0,11
5 F5 L 101 x Basarabeanca–68 1,59±0,15 V
1,00±0,15
6 F5 Basarabeanca x L 101–60 1,24±0,12*V 0,19±0,01*
V
7 F5 Basarabeanca x L 101–61 0,95±0,11*V 0,39±0,05*
V
8 F5 Basarabeanca x L 101–62 1,03±0,12*V 0,20±0,02*
V
* – deosebire cu suport statistic de genitorul L 101, la nivelul p<0,05; V – deosebire cu suport statistic de genitorul Basarabeanca, la nivelul p<0,05.
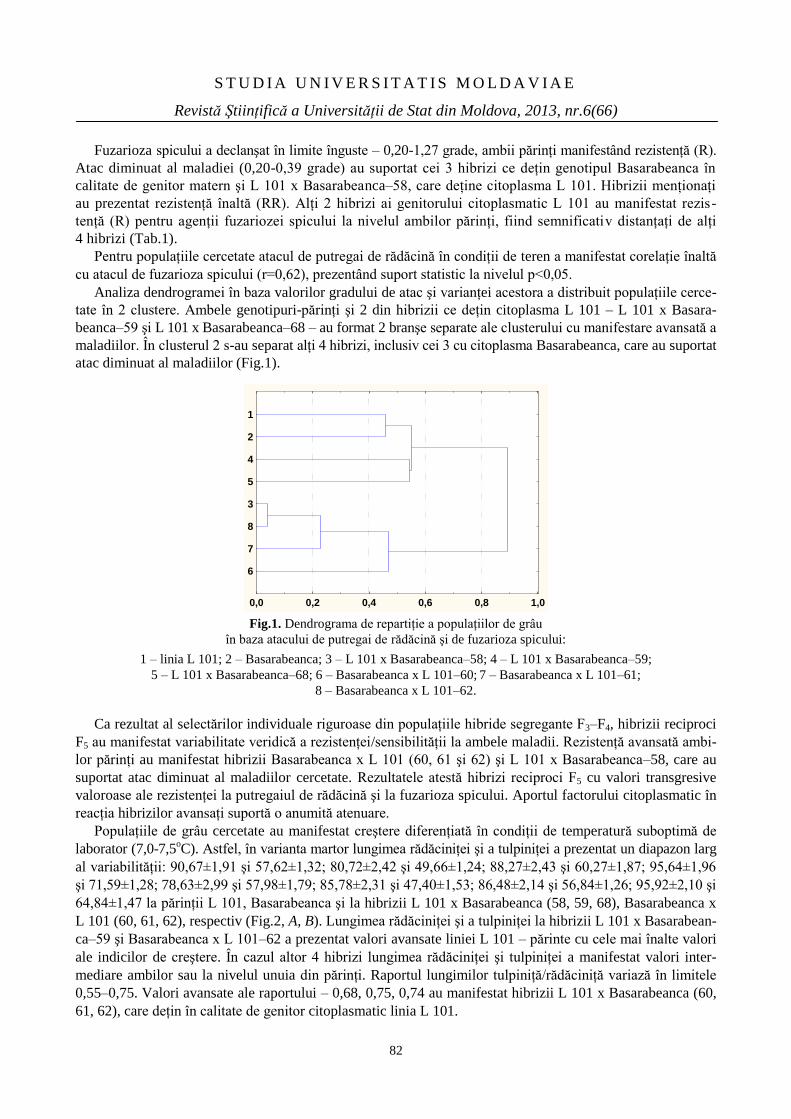
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
82
Fuzarioza spicului a declanşat în limite înguste – 0,20-1,27 grade, ambii părinţi manifestând rezistenţă (R).
Atac diminuat al maladiei (0,20-0,39 grade) au suportat cei 3 hibrizi ce deţin genotipul Basarabeanca în
calitate de genitor matern şi L 101 x Basarabeanca–58, care deţine citoplasma L 101. Hibrizii menţionaţi
au prezentat rezistenţă înaltă (RR). Alţi 2 hibrizi ai genitorului citoplasmatic L 101 au manifestat rezis-
tenţă (R) pentru agenţii fuzariozei spicului la nivelul ambilor părinţi, fiind semnificativ distanţaţi de alţi
4 hibrizi (Tab.1).
Pentru populaţiile cercetate atacul de putregai de rădăcină în condiţii de teren a manifestat corelaţie înaltă
cu atacul de fuzarioza spicului (r=0,62), prezentând suport statistic la nivelul p<0,05.
Analiza dendrogramei în baza valorilor gradului de atac şi varianţei acestora a distribuit populaţiile cerce-
tate în 2 clustere. Ambele genotipuri-părinţi şi 2 din hibrizii ce deţin citoplasma L 101 – L 101 x Basara-
beanca–59 şi L 101 x Basarabeanca–68 – au format 2 branşe separate ale clusterului cu manifestare avansată a
maladiilor. În clusterul 2 s-au separat alţi 4 hibrizi, inclusiv cei 3 cu citoplasma Basarabeanca, care au suportat
atac diminuat al maladiilor (Fig.1).
0,0 0,2 0,4 0,6 0,8 1,0
6
7
8
3
5
4
2
1
Fig.1. Dendrograma de repartiţie a populaţiilor de grâu
în baza atacului de putregai de rădăcină şi de fuzarioza spicului:
1 – linia L 101; 2 – Basarabeanca; 3 – L 101 x Basarabeanca–58; 4 – L 101 x Basarabeanca–59;
5 – L 101 x Basarabeanca–68; 6 – Basarabeanca x L 101–60; 7 – Basarabeanca x L 101–61;
8 – Basarabeanca x L 101–62.
Ca rezultat al selectărilor individuale riguroase din populaţiile hibride segregante F3–F4, hibrizii reciproci
F5 au manifestat variabilitate veridică a rezistenţei/sensibilităţii la ambele maladii. Rezistenţă avansată ambi-
lor părinţi au manifestat hibrizii Basarabeanca x L 101 (60, 61 şi 62) şi L 101 x Basarabeanca–58, care au
suportat atac diminuat al maladiilor cercetate. Rezultatele atestă hibrizi reciproci F5 cu valori transgresive
valoroase ale rezistenţei la putregaiul de rădăcină şi la fuzarioza spicului. Aportul factorului citoplasmatic în
reacţia hibrizilor avansaţi suportă o anumită atenuare.
Populaţiile de grâu cercetate au manifestat creştere diferenţiată în condiţii de temperatură suboptimă de
laborator (7,0-7,5oC). Astfel, în varianta martor lungimea rădăciniţei şi a tulpiniţei a prezentat un diapazon larg
al variabilităţii: 90,67±1,91 şi 57,62±1,32; 80,72±2,42 şi 49,66±1,24; 88,27±2,43 şi 60,27±1,87; 95,64±1,96
şi 71,59±1,28; 78,63±2,99 şi 57,98±1,79; 85,78±2,31 şi 47,40±1,53; 86,48±2,14 şi 56,84±1,26; 95,92±2,10 şi
64,84±1,47 la părinţii L 101, Basarabeanca şi la hibrizii L 101 x Basarabeanca (58, 59, 68), Basarabeanca x
L 101 (60, 61, 62), respectiv (Fig.2, A, B). Lungimea rădăciniţei şi a tulpiniţei la hibrizii L 101 x Basarabean-
ca–59 şi Basarabeanca x L 101–62 a prezentat valori avansate liniei L 101 – părinte cu cele mai înalte valori
ale indicilor de creştere. În cazul altor 4 hibrizi lungimea rădăciniţei şi tulpiniţei a manifestat valori inter-
mediare ambilor sau la nivelul unuia din părinţi. Raportul lungimilor tulpiniţă/rădăciniţă variază în limitele
0,55–0,75. Valori avansate ale raportului – 0,68, 0,75, 0,74 au manifestat hibrizii L 101 x Basarabeanca (60,
61, 62), care deţin în calitate de genitor citoplasmatic linia L 101.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
83
M
M±SE
M±1,96*SE
1 3 5 7 9 11 13 15 17 19 21 2360
65
70
75
80
85
90
95
100
105
M
M±SE
M±1,96*SE
1 3 5 7 9 11 13 15 17 19 21 2340
45
50
55
60
65
70
75
80
A B
Fig.2. Lungimea rădăciniţei (A) şi a tulpiniţei (B) în condiţii de temperatură suboptimă, mm.
Pe orizontală: 1, 2, 3 – L 101; 4, 5, 6 – Basarabeanca; 7, 8, 9 – L 101 x Basarabeanca–58; 10, 11, 12 – L 101 x
Basarabeanca–59; 13, 14, 15 – L 101 x Basarabeanca–68; 16, 17, 18 – Basarabeanca x L 101–60; 19, 20, 21 –
Basarabeanca x L 101–61; 22, 23, 24 – Basarabeanca x L 101–62, respectiv: martor,
FC Fusarium oxysporum var. orthoceras şi FC Helminthosporium avenae.
Metaboliţii a 3 tulpini F. oxysporum var. orthoceras au produs atât inhibiţie, cât şi stimulare a creşterii în
limitele 85,2–105,1 şi 82,8–113,7% din martor, respectiv, pentru rădăciniţă şi tulpiniţă (Fig.2, A, B). Hibridul
L 101 x Basarabeanca–68 a manifestat sensibilitate la FC F. oxysporum var. orthoceras la nivelul părintelui
citoplasmatic – L 101. Hibrizii reciproci L 101 x Basarabeanca–59 şi Basarabeanca x L 101–62 au manifestat
sensibilitate avansată liniei L 101, cu distincţie veridică de martor. La acţiunea inhibitoare a FC F. oxysporum
var. orthoceras tulpiniţa a fost superior afectată în raport cu rădăciniţa. Acţiunea stimulativă a metaboliţilor
Fusarium, avansată pentru tulpiniţă, se manifestă pronunţat la genotipul-părinte Basarabeanca şi la hibrizii
cu citoplasma acestuia – Basarabeanca x L 101 (60 şi 61). Metaboliţii H. avenae au manifestat fitotoxicitate
avansată în raport cu F. oxysporum var. orthoceras pentru toate populaţiile. La acţiunea FC H. avenae atât
genitorii, cât şi hibrizii reciproci au prezentat doar inhibiţie pentru ambii indici. Acţiunea inhibitoare maximă
a fost suportată de rădăciniţa şi tulpiniţa liniei L 101. Acest părinte a manifestat sensibilitate medie la agenţii
PR în condiţii de câmp. Spre deosebire de F. oxysporum var. orthoceras, acţiunea FC H. avenae a fost supe-
rior inhibitoare pentru lungimea rădăciniţei, excepţie prezentând doar linia L 101 şi hibridul L 101 x Basara-
beanca–59. Astfel, la acţiunea metaboliţilor agenţilor PR populaţiile de grâu au manifestat o specificitate
înaltă, atât de genotip, cât şi de organ testat, mai puternic manifestată în cazul FC cu fitotoxicitate avansată –
H. avenae. La administrarea acestora creşte decalajul dintre lungimea rădăciniţei şi cea a tulpiniţei. Feno-
menul fiind mai pronunţat în cazul H. avenae şi, totodată, la hibrizii cu citoplasma Basarabeanca, fenomen
specific genitorului matern (Tab.2).
Tabelul 2
Reacţia plantulelor de grâu la acţiunea filtratelor de cultură (% din martor, media)
Organul testat L 101
Basara-
beanca
L 101 x
Basara-
beanca
L 101 x
Basara-
beanca
L 101 x
Basara-
beanca
Basara-
beanca
x L 101
Basara-
beanca
x L 101
Basara-
beanca
x L 101
Nr.înreg. (58) (59) (68) (60) (61) (62)
Fusarum oxysporum var. orthoceras
Rădăciniţa 93,3* 105,1* 96,5 85,2* 93,5* 105,0* 102,2 89,3*
Tulpiniţa 92,9* 111,8* 104,6 83,8* 90,9* 113,7* 110,0* 82,8*
Helminthosporium avenae
Rădăciniţa 80,1* 82,6* 82,6* 85,1* 87,7* 85,4* 76,7* 80,8*
Tulpiniţa 79,4* 95,2* 89,1* 79,3* 92,1* 96,4 88,2* 85,2*
* – deosebire cu suport statistic de varianta martor la nivelul p<0,05.
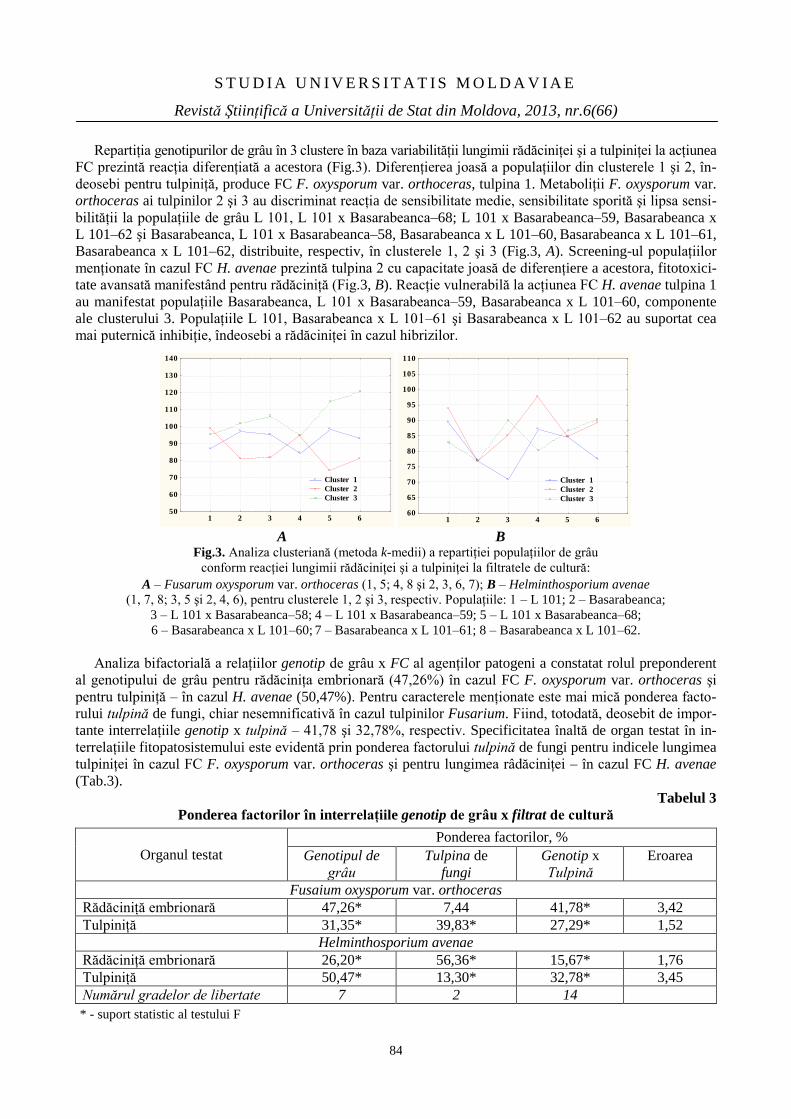
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
84
Repartiţia genotipurilor de grâu în 3 clustere în baza variabilităţii lungimii rădăciniţei şi a tulpiniţei la acţiunea
FC prezintă reacţia diferenţiată a acestora (Fig.3). Diferenţierea joasă a populaţiilor din clusterele 1 şi 2, în-
deosebi pentru tulpiniţă, produce FC F. oxysporum var. orthoceras, tulpina 1. Metaboliţii F. oxysporum var.
orthoceras ai tulpinilor 2 şi 3 au discriminat reacţia de sensibilitate medie, sensibilitate sporită şi lipsa sensi-
bilităţii la populaţiile de grâu L 101, L 101 x Basarabeanca–68; L 101 x Basarabeanca–59, Basarabeanca x
L 101–62 şi Basarabeanca, L 101 x Basarabeanca–58, Basarabeanca x L 101–60, Basarabeanca x L 101–61,
Basarabeanca x L 101–62, distribuite, respectiv, în clusterele 1, 2 şi 3 (Fig.3, A). Screening-ul populaţiilor
menţionate în cazul FC H. avenae prezintă tulpina 2 cu capacitate joasă de diferenţiere a acestora, fitotoxici-
tate avansată manifestând pentru rădăciniţă (Fig.3, B). Reacţie vulnerabilă la acţiunea FC H. avenae tulpina 1
au manifestat populaţiile Basarabeanca, L 101 x Basarabeanca–59, Basarabeanca x L 101–60, componente
ale clusterului 3. Populaţiile L 101, Basarabeanca x L 101–61 şi Basarabeanca x L 101–62 au suportat cea
mai puternică inhibiţie, îndeosebi a rădăciniţei în cazul hibrizilor.
1 2 3 4 5 650
60
70
80
90
100
110
120
130
140
Cluster 1
Cluster 2
Cluster 3
1 2 3 4 5 660
65
70
75
80
85
90
95
100
105
110
Cluster 1
Cluster 2
Cluster 3
A B
Fig.3. Analiza clusteriană (metoda k-medii) a repartiţiei populaţiilor de grâu
conform reacţiei lungimii rădăciniţei şi a tulpiniţei la filtratele de cultură:
A – Fusarum oxysporum var. orthoceras (1, 5; 4, 8 şi 2, 3, 6, 7); B – Helminthosporium avenae
(1, 7, 8; 3, 5 şi 2, 4, 6), pentru clusterele 1, 2 şi 3, respectiv. Populaţiile: 1 – L 101; 2 – Basarabeanca;
3 – L 101 x Basarabeanca–58; 4 – L 101 x Basarabeanca–59; 5 – L 101 x Basarabeanca–68;
6 – Basarabeanca x L 101–60; 7 – Basarabeanca x L 101–61; 8 – Basarabeanca x L 101–62.
Analiza bifactorială a relaţiilor genotip de grâu x FC al agenţilor patogeni a constatat rolul preponderent
al genotipului de grâu pentru rădăciniţa embrionară (47,26%) în cazul FC F. oxysporum var. orthoceras şi
pentru tulpiniţă – în cazul H. avenae (50,47%). Pentru caracterele menţionate este mai mică ponderea facto-
rului tulpină de fungi, chiar nesemnificativă în cazul tulpinilor Fusarium. Fiind, totodată, deosebit de impor-
tante interrelaţiile genotip x tulpină – 41,78 şi 32,78%, respectiv. Specificitatea înaltă de organ testat în in-
terrelaţiile fitopatosistemului este evidentă prin ponderea factorului tulpină de fungi pentru indicele lungimea
tulpiniţei în cazul FC F. oxysporum var. orthoceras şi pentru lungimea râdăciniţei – în cazul FC H. avenae
(Tab.3).
Tabelul 3
Ponderea factorilor în interrelaţiile genotip de grâu x filtrat de cultură
Organul testat
Ponderea factorilor, %
Genotipul de
grâu
Tulpina de
fungi
Genotip x
Tulpină
Eroarea
Fusaium oxysporum var. orthoceras
Rădăciniţă embrionară 47,26* 7,44 41,78* 3,42
Tulpiniţă 31,35* 39,83* 27,29* 1,52
Helminthosporium avenae
Rădăciniţă embrionară 26,20* 56,36* 15,67* 1,76
Tulpiniţă 50,47* 13,30* 32,78* 3,45
Numărul gradelor de libertate 7 2 14
* - suport statistic al testului F

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
85
Concluzii Hibrizii reciproci (F5) au manifestat rezistenţă diferită: mai înaltă ca la ambii/unul din genitori sau la ni-
velul acestora pentru putregaiul de rădăcină şi fuzarioza spicului în condiţii de teren. Între atacul de putregai de rădăcină şi de fuzarioza spicului s-a constatat o corelaţie înaltă semnificativă.
Rezultatele atestă hibrizi reciproci F5 – L 101 x Basarabeanca–58, Basarabeanca x L 101–60, 61 şi 62 cu indici transgresivi valoroşi ai rezistenţei la putregaiul de rădăcină şi la fuzarioza spicului.
Raportul lungimilor tulpiniţă/rădăciniţă în condiţii de temperatură suboptimă de laborator variază în limi-tele 0,55–0,75. Valori avansate ale raportului au manifestat hibrizii L 101 x Basarabeanca, care deţin în cali-tate de genitor citoplasmic linia L 101 – părinte cu cele mai înalte valori ale indicilor de creştere.
Filtratele de cultură ale tulpinilor Fusarium oxysporum var. orthoceras au produs atât inhibiţia, cât şi stimu-larea creşterii rădăciniţei embrionare şi a tulpiniţei. Tulpiniţa a fost superior afectată în raport cu rădăciniţa. Acţiunea stimulativă a metaboliţilor Fusarium, avansată pentru tulpiniţă, s-a manifestat pronunţat la genotipul-părinte Basarabeanca şi la hibrizii cu citoplasma acestuia – Basarabeanca x L 101 (60 şi 61).
Populaţiile-părinţi şi hibrizii reciproci au manifestat sensibilitate mult mai avansată la metaboliţii Helmin-thospoium avenae. Acţiune inhibitoare maximă a fost suportată de rădăciniţa şi tulpiniţa liniei L 101. Filtra-tele menţionate au fost superior inhibitive pentru lungimea rădăciniţei, excepţie prezentând linia L 101 şi hibridul L 101 x Basarabeanca–59.
S-a constatat creşterea decalajului dintre lungimea rădăciniţei şi cea a tulpiniţei. Fenomenul fiind mai pro-nunţat în cazul tulpinilor H. avenae şi, totodată, la hibrizii cu citoplasma Basarabeanca, fenomen specific genitorului citoplasmatic.
Analiza bifactorială a relaţiilor genotip de grâu x FC al agenţilor patogeni a constatat rolul preponderent al genotipului de grâu pentru rădăciniţa embrionară (47,26%) în cazul FC F. oxysporum var. orthoceras şi pentru tulpiniţă (50,47%) în cazul H. avenae.
Este evidentă specificitatea înaltă de organ testat în interrelaţiile fitopatosistemului: ponderea factorului tulpină de fungi pentru indicele lungimea tulpiniţei în cazul FC F. oxysporum var. orthoceras şi pentru lun-gimea râdăciniţei – în cazul FC H. avenae.
Bibliografie:
1. COTUNA, O. şi al. Comportarea unor soiuri de grâu de toamnă la atacul ciupercii Drechslera tritici – repentis (Died.) Shoemaker, în condiţiile climatice din Câmpia Banatului. În: Vegetal, 2011, Anul III, nr.I (VIII), p.58-65.
2. LUPAŞCU, G., SAŞCO, E., GAVZER, S. Componenţa speciilor de fungi care produc boli la Triticum aestivum L. În: Buletinul AŞM. Ştiinţele vieţii, 2008, nr.2(305), p.66-73. ISSN 1857-064X
3. BOVILL, W. et al. Identification of novel QTL for resistance to crown rot in the doubled haploid wheat population ‘W21MMT70’x ‘Mendos’. In: Plant Breeding, 2006, vol.125, no.6, p.538-543.
4. COLLARD, B. et al. Development of molecular markers for crown rot resistance in wheat: mapping of QTL for seedling resistance in a ‘2-49’ x ‘Janz’ population. In: Plant Breeding, 2005, vol.124, p.532-537.
5. HAJIEGHRARI, B. Wheat crown and root rotting fungi in Moghan area, Northwest of Iran. In: African Journal of Biotechnology, 2009, vol.8, no.22, p.6214-6219. ISSN 1684-5315
6. MA, J. et al. Identification and validation of a major QTL conferring crown rot resistance in hexaploid wheat. In: Theoretical and Applied Genetics, 2010, vol.120, no.6, p.1119-1128.
7. NICOL, J. et al. The international Breeding Strategy for the Incorporation of resistance in bread wheat against the soil borne pathogens (Dryland root rot and cyst and lesion cereal nematodes) using conventional and molecular tools. In: BUCK, H. et al. (eds.). Wheat Production in Stressed Environments. Developments in Plant Breeding, 2007, vol.12, p.125-137.
8. PANDEY, S. et al. Sources of inoculum and reappearance of spot blotch of wheat in rice–wheat cropping. In: European Journal of Plant Pathology, 2005, vol.111, no.1, p.47-55.
9. RODERMAN, B., BARTELS, G. Reduction of mycotoxins in cereal grain using the resistance of wheat and triticale varieties against Fusarium. In: 29
th Mycotoxin Workshop. Conference abstracts. Fellbach, Germany, 2007, p.42.
10. ЛУПАШКУ Г.А. Иммуногенетические основы устойчивости культуры тритикале к фузариозу / Автореф. докт. дисc. Москва, 1999. 46 с.
11. ОВСЯНКИНА, А.В. Сортоизучение и подбор сортов зерновых культур, обладающих устойчивостью к корне-вой гнили. К 110-летию академика ВАСХНИЛ и Россельхозакадемии М.С. Дунина. В: Культурные растения для устойчивого сельского хозяйства в XXI веке (иммунитет, селекция, интродукция). Москва, 2011, том IV, часть I, с.96-100.
12. СМУРОВА, С.Г. Новые источники и доноры устойчивости пшеницы к Cochliobolus sativus Drechs.ex Dastur / Автoреф. дисc. канд. биол. наук. Пушкин, Ленинград. обл., 2008. 23 с.
Prezentat la 23.08.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
86
ВЛИЯНИЕ БАКТЕРИАЛЬНЫХ МЕТАБОЛИТОВ НА ХАРАКТЕР
ПРОРАСТАНИЯ КОНИДИЙ ГРИБА АЛЬТЕРНАРИЯ
Аркадий НИКОЛАЕВ, Светлана НИКОЛАЕВА, Виктория ШУБИНА
Институт защиты растений и экологического земледелия АН Молдовы
INFLUENŢA METABOLIŢILOR BACTERIENI ASUPRA CARACTERULUI DE GERMINARE
A CONIDIILOR CIUPERCII ALTERNARIA Culturile B. subtilis cu vârsta de trei zile inhibau germinarea conidiilor de Alternaria mai puternic decât culturile cu
vârsta de două zile. În plus, faţă de inhibare efectul antifungal s-a manifestat în formarea umflăturilor veziculoase în locul hifelor germinative.
Cuvinte-cheie: Bacillussubtilis, Alternaria, efect antifungal, germinarea conidiilor, umflături veziculoase.
EFFECT OF BACTERIAL METABOLITES ON THE CHARACTER OF GERMINATION CONIDIA OF
FUNGUS ALTERNARIA Three day liquid cultures of B. subtilis have inhibited germination of conidia stronger than 2-day ones. In addition to
suppression of conidia germination the antifungal effect manifested also in formation of vesicular swellings instead of filamentous germs.
Keywords: Bacillus subtilis, Alternaria, antifungal effect, conidia germination, vesicular swelling.
Защита любой сельскохозяйственной культуры от вредителей и болезней предполагает комплекс мероприятий, включающих высокий уровень агротехники (севооборот, система обработки почвы, обеспеченность питательными веществами), устойчивые сорта, грамотное использование пестицидов химической или биологической природы.
В последние годы все больше внимания уделяется экологическому земледелию, получению эколо-гически чистой продукции, хотя доля и того, и другого остается невысокой (в развитых странах не превышает 10%) [10]. Органическое земледелие активно развивается в странах ЕС, США, Японии, Китае. Продукция органического земледелия пользуется большим спросом.
В настоящее время в мире возрастает научный и практический интерес к разработке и производ-ственному использованию защитных биопрепаратов как в качестве самостоятельных средств, так и в системах интегрированной защиты растений. В мировом производстве биологических средств защи-ты растений биопрепараты для контроля вредителей растений составляют 10%, для контроля возбу-дителей болезней – 4,6%, для контроля сорняков – 1,3% Исследования биоагентов, биопрепаратов и разработка технологий их применения проводятся в США, странах ЕС и других развитых и разви-вающихся странах [8]. Выход на рынки США и Канады большого количества биопестицидов объяс-няется упрощенной системой их регистрации в этих странах.
Большое внимание уделяется биометоду в России, Беларуси, Украине. Этому способствуют пра-вительственные решения Российской Федерации, возможность наработки биопрепаратов на заводах Беларуси, наличие мощных биолабораторий в Украине. Биологической защите растений посвяща-лись международные научно-практические конференции в Краснодаре (2004, 2008, 2010 гг.), Киеве (Совещание ВПРС МОББ, 2009), Кишиневе (2010), Беларуси (2011), где были представлены материа-лы научных исследований ученых разных стран.
Основное количество биопестицидов производится на базе бактерий, в меньшей степени – на базе грибов и вирусов. Из бактерий основными продуцентами биопрепаратов являются споровые бактерии и псевдомонады, из грибов – грибы рода триходерма.
В США из биофунгицидов наиболее востребованы препараты на основе Вacillus subtilis и В. pumilis. По данным NASS, ретроспективной динамики и по другим косвенным показателям, В. subtilis обра-батывают почти 40% сладкого перца, более 20% томатов, около 10% баклажанов, 2% апельсиновых деревьев, 15% танжеринов, 1,3% яблонь, 25% виноградников. В. pumilis широко используют для об-работки кочаннoго салата и шпината, 5% яблонь и 2% персиков.
В настоящее время в России на рынке представлены около 20 активных агентов биопестицидов инсектицидного (Bacillus thuringiensis var. kurstaki, B.th. var. tenebriosis, B.th. var. thuringiensis) и фун-гицидного (Bacillus subtilis, Trichoderma harzianum и бактерии p. Pseudomonas) действия.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
87
На российском рынке преобладают препараты с фунгицидной активностью (почти 80%). Наиболее востребованными активными агентами являются бактериальные микроорганизмы [3].
В Молдове на базе псевдомонад зарегистрированы биопрепараты Ризоплан (планриз) и Паурин, а также препараты на базе грибов Trichoderma lignorum и T. harzianum.
Обычно заражение растений начинается с прорастания конидий, попавших на них. В связи с этим предотвращение прорастания конидий с помощью фунгицидов является одним из основных элемен-тов технологии защиты растений. Поэтому обоснованный выбор фунгицидов следует начинать с оценки их действия на прорастание конидий. Это и составляло цель наших исследований.
Задачей исследований являлись: оценка фунгицидного действия споровых бактерий на конидии возбудителя альтернариоза томатов; характер их прорастания в различных концентрациях жидких культур; определение минимальных эффективных концентраций биопрепарата.
Материалы и методы исследования В ходе исследований использовали четыре культуры бактерий B. subtilis, ингибирующих рост гри-
ба Альтернария в опытах in vitro и отличающихся культурально-морфологическими признаками. Культуры нарабатывали на оригинальной жидкой питательной среде в течение 2-3-х суток. Тем-
пература инкубации в течение суток колебалась в пределах 24-320 С.
Титр 3-суточных бактериальных культур колебался в пределах (1,3-3,1)×1010
КОЕ/мл. Конидии гриба Альтернария, полученные по методике AllenS.J. и др. [1], проращивали в 2-х и
3-суточных культурах споровых бактерий в термостате (без подсветки) при 280С. Использовали ис-
ходные культуры и их разведения 1:10; 1:20; 1:100. Для лучшей смачиваемости конидий разведения готовили на 0,01% растворе Твина 80.
Параллельно конидии Альтернарии проращивали в растворе химического препарата OxideWP, ре-комендованного для защиты культуры томата от альтернариоза. В качестве исходной брали концен-трацию, рекомендованную для производственного применения.
Контролем служили вода и питательная среда, на которых культивировали микроорга-низмы. Питательную среду брали в тех же раз-ведениях, что и культуральные жидкости.
Характер прорастания конидий учитывали через 2, 3, 4, 5 и 24 часа экспозиции.
Результаты и их обсуждение В воде (контроль 2) уже через 2 часа экспо-
зиции конидии начали прорастать нормальным ростком, через 3 часа проросло более 50% ко-нидий, через 4 часа нормально проросли все конидии (табл.).
По сравнению с водой, питательная среда во всех использованных концентрациях сти-мулировала прорастание конидий.
Раствор химического эталона Oxide WP во
всех концентрациях, включая исходную (ре-
комендованную для практического примене-
ния), через сутки экспозиции терял фунгицид-
ное действие на конидии. В течение первых 5
часов экспозиции ни в одной из концентраций
конидии не прорастали. В суспензиях 2-суточных бактериальных
культур (исходная, 10%, 5%) всех исследуе-мых бактерий после 4-х часов экспозиции не наблюдалось нормального прорастания кони-дий, а в 1% концентрации культуры Я кони-дии вообще не проросли нормально (рис.1).
Рис.1. Действие антифунгальных метаболитов B. Subtilis на
прорастание конидий возбудителя альтернариоза томатов. (А.Н. Николаев, В.Э. Шубина, С.И. Николаева, 2013 [9]) Слева вверху – контроль (вода), прорастание нормальным
ростком; справа вверху – прорастание пузырем в растворе
метаболитов B. subtilis; внизу – более наглядный
фрагмент прорастания пузырями.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
88
В культуре 10-98 наблюдалось прорастание нормальным ростком только единичных конидий, в культурах А и 2К отмечено только прорастание малыми пузырями. Через 5 часов экспозиции в 1% концентрациях у трех культур (2К, 10-98, Я) отмечено прорастание нормальным ростком или боль-шим пузырем единичных конидий. Такая же картина сохранялась и через 24 часа экспозиции, когда прорастание малым или большим пузырем отмечено и для концентраций 5%,10%.
В исходной концентрации прорастания конидий не отмечено ни у одной из исследуемых культур. Фунгицидное действие трехсуточных культур бактерий более выражено, чем у 2-суточных, что хо-рошо отражено в таблице.
Таблица
Характер прорастания конидий Альтернарии в культуральной жидкости культур Bacillus subtilis
Культура
бактерий
Концен-
трация
жидкой
культуры
Возраст бактериальной культуры и время
экспозиции в часах
2-суточная 3-суточная
2 3 4 5 24 2 3 4 5 24
А
исходная - - - - - - - - - -
10% - - - - - - - - - -
5% - - - - - - - - - -
1% - - -МП -МП -МП - - -МП -МП -МП
2К
исходная - - - - - - - - - -
10% - - - - -БП - - - - -
5% - - - -БП -БП - - - - -
1% - - -МП + -БП + -БП - - -МП -БП -БП
10-98
исходная - - - - - - - - - -
10% - - - - - - - - - -
5% - - - - -МП - - - - -МП
1% - - + - + -БП + -БП - - -БП -БП -БП
Я
исходная - - - - - - - - - -
10% - - - - -МП - - - - -
5% - - - -МП -МП - - - - -БП
1% - - - + -БП + -БП - - -МП -БП -БП
Oxide WP
исходная - - - - ++
10% - - - - ++
5% - - - - ++
1% - - - - +++
Котроль1
(пит.
среда)
исходная ++ ++ ++ ++ ++++
10% ++ ++ ++ ++ ++++
5% ++ ++ ++ ++ ++++
1% ++ ++ ++ ++ ++++
Котроль2
(вода)
- + - + ++ ++ +++
Условные обозначения: - — прорастания конидий нет;
-МП — прорастание конидий малым пузырьком;
-БП — прорастание конидий большим пузырем;
+ - — прорастание нормальным ростком единичных конидий;
+-БП — прорастание нормальным ростком или большими пузырями единичных конидий;
+ — более 50% конидий проросло нормальным ростком;
++ — все конидии проросли нормальным ростком;
+++ — все конидии проросли нормальным ростком, проростки длинные;
+++ — обильный воздушный мицелий.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
89
Формирование пузыревидных вздутий вместо гифовых ростков под влиянием метаболитов B. subtilis
имеет место при прорастании конидий не только у Альтернарии, но и у других грибов.
Так, On-Uma Ruangwong и др. [5] отмечали такое явление у Colletotrichum gloesporioides (рис.2).,
Коломиец Э.И. и др. [7] – у Peniciillium expansum и Botrytis cinerea, Ivan Petatan-Sagahon и др. [2] –
для Stenocarpella maydis (рис.3).
Рис.2. Действие антибиотических веществ B. Subtilis на конидии C. gloeosporioides. (по On-Uma Ruangwong и др. [5])
c – нормальное прорастание, d – непроросшие конидии, е – шаровидное видоизменение ростковой трубочки
под влиянием антифунгальных метаболитов.
Рис.3. Влияние термостабильного фильтрата Bacillus subtilis 160 на прорастание конидий Stenocarpella maydis. (по Ivan Petatan-Sagahon и др. [2])
а) нормальное прорастание конидий, в) прорастание под влиянием метаболитов Bacillus.
Стрелками показано образование пузыревидных вздутий, напоминающих хламидоспоры
Mara Favilla и др. [4] сообщают об аналогичных нарушениях у Aspergillus под воздействием био-
фунгицида фузапирона. Jošiĉ Dragana и др. [3] наблюдали этот феномен у Alternaria tenuissima под
воздействием антибиотических веществ Pseudomonas (рис.4).
Рис.4. Влияние фунгицида DACOFLO 0,2% (A) и изолятов Pseudomonas Q16 (B) и
B25 (C) в концентрации 105 на прорастание конидий A. tenuissima
(по Dragana Jošiĉ и др. [3])

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
90
Это указывает на наличие специфического механизма действия антибиотических веществ B. subtilis
и некоторых других бактерий на характер роста ростковых гиф. Это, по-видимому, должно отразить-
ся на способности ростковых гиф проникать в клетки растений хозяев и вызывать их заражение.
Выводы
Исследованные нами жидкие культуры B. subtilis 2-х и 3-суточного возраста обнаруживают высо-
кое фунгицидное действие на конидии возбудителя альтернариоза томатов, которое сохранялось в
той или иной мере и в 1% разведении.
Трехсуточные культуры более активны, чем 2-суточные. Большая фунгицидная активность обна-
ружена у 3-суточных культур А и 2К, которая сохранялась при 5% концентрациях в течение всего
периода наблюдений (24 часа).
В зависимости от концентрации жидкой культуры и от штамма бактерий наблюдалась задержка
прорастания конидий от 3 до 24 часов.
Под влиянием антифунгальных метаболитов исследованные культуры B. subtilis кроме задержки
прорастания конидий вызывали и изменения характера их прорастания: вместо нормального ростка
гиф образуются пузыри.
Библиография:
1. ALLEN, S.J., BROWN, J.F., KOCHMAN, J.K. Production of inoculum and field assessment of Alternaria helianthi
on sunflower. In: Plant disease, 1983, v.67, p.665-668.
2. IVAN PETATAN-SAGAHON и др. Isolation of bacteria with antifungal activity against the phytopathogenic fungi
Stenocarpella maydis and Stenocarpella macrospora. In: Molecular Sciences, 2011, v.12, p.5522-5537.
3. JOŠIĈ DRAGANA, KATARINA PROTOLIPAC, MIRA STAROVIĈ, SAŠA STOJANOVIĈ, SNEŢANA
PAVLOVIĈ, MILADINOVIĈ, M. AND SVETLANA RADOVIĈ. Phenazines produsing Pseudomonas isolates de-
crease Alternaria tenuissima growth, pathogenicity and disease incidence on cardoon. In: Arch. Biol. Sci, Belgrade,
2012, 64(4), p.1495-1503.
4. MARA FAVILLA, MICHELANGELO PASCALE, ALESSANDRA RICELLI, ANTONIO EVIDENTE,
CARMINE AMALFITANO AND CLAUDIO ALTOMARE. Inhibition of Species of the Aspergillus Section Nigri
and Ochratoxin A Production in Grapes by Fusapyrone. In: Applied and Environmental Microbiology, Apr. 2008,
v.74, no.7, p.2248-2253.
5. ON-UMA RUANGWONG, CHI-L CHANG, SENGHOR AMADOU LAMINE AND WEN-JINN LIANG. Identi-
fication of antifungal compound produced by Bacillus subtilisLB5 with ability to control anthracnose disease caused
by Colletotrichumgloeosporioides. In: African Journal of Microbiology Research, 30 April, 2012, vol. 6(16),
p.3732-3736.
6. БИЗЮКОВА, О.В. Обзор мирового рынка микробиопрепаратов. В: Защита и карантин растений, 2012, №3,
с.9-12.
7. КОЛОМИЕЦ, Э.И., КИЛЬЧЕВСКАЯ, О.В., КУПЦОВ, В.Н., РОМАНОВСКАЯ, Т.В., СВИРИДОВ А.В.
Научные и практические основы создания биопрепарата для защиты сахарной свеклы от кагатной гнили.
В: Информационный бюллетень ВПРС МОББ. С.-Петербург, 2007, №38, с.145-147.
8. МОНАСТЫРСКИЙ, О.А. Современные направления в создании и практическом применении защитных
биопрепаратов и биотехнологий в растениеводстве. В: Биологическая защита растений – основа стабили-
зации агроэкосистем. Вып.6. Краснодар, 2010, с.335-342.
9. НИКОЛАЕВ, А.Н., ШУБИНА, В.Э., НИКОЛАЕВА, С.И. Действие Bacillus subtilis на прорастание конидий
возбудителя альтернариоза томатов. В: Защита растений в современных технологиях возделывания сель-
скохозяйственных культур: Материалы международной научно-практической конференции (п. Краснообск,
24-6 июля 2013 г.). Новосибирск, 2013, с.256-259.
10. ФОКИН, А.В. Биологическая защита растений – процесс циклический. В: Защита и карантин растений,
2010, №3, с.25.
Prezentat la 19.09.2013

Seria “Ştiinţ e reale şi ale naturii”
Biologie ISSN 1814-3237
91
ДЕЙСТВИЕ ЭКСТРАКТОВ СЕМЯН ВИНОГРАДА НА ФИТОПАТОГЕНЫ
В УСЛОВИЯХ IN VITRO
Аркадий НИКОЛАЕВ, Светлана НИКОЛАЕВА, Виктория ШУБИНА
Институт защиты растений и экологического земледелия АН Молдовы
ACŢIUNEA EXTRACTELOR DIN SEMINŢE DE STRUGURI ASUPRA FITOPATOGENILOR
ÎN CONDIŢII IN VITRO Extracte din seminţe de struguri prin diferite metode au fost testate privitor la activitatea lor fungicidică şi fungista-
tică contra fitopatogenilor în condiţii in vitro. A fost demonstrată activitatea fungistatică a extractelor în cazul introdu-cerii lor în mediul de cultivare.
Cuvinte-cheie: extracte, seminţe de struguri, acţiune fungicidică, acţiune fungistatică.
EFFECT OF EXSTRACTS OF GRAPE SEEDS ON PHYTOPATHOGENS IN VITRO Extracts of grape seeds by different procedures were tested on their fungicidal and fungistatical activity under in
vitro condition. It was demonstrated their fungistatical activity when introduced into cultural medium. Keywords: extracts of grape seeds, antifungal and fungistatical effect.
Молдова – республика виноградарства и виноделия. С одного гектара виноградников после пере-работки винограда на выжимки приходится примерно 3 тонны, из которых 0,7-1,2 тонны составляют семена. Биологически активные вещества, выделенные из семян винограда, находят применение в медицине, косметологии, кондитерской промышленности [3]. Вполне объясним интерес к экстрактам семян и со стороны представителей других специальностей. Так, по данным Настас Т. и др. [4], био-логически активные вещества, экстрагированные из семян винограда, обладают хорошо выраженными инсектицидными, антифидантными и гербицидными свойствами.
В течение ряда лет (2006, 2011 и 2012 гг.) мы выясняли наличие фунгицидного эффекта экстрактов виноградных семян. Нам были предоставлены препараты Эноксил (экстракт этиловым спиртом), Халотанил (экстракт 15% метиловым спиртом), Энотанин (экстракт пропиленгликолем).
Методика исследований Экстракты виноградных семян испытывали в исходном виде и в разведениях, в качестве контроля
брали соответствующие растворители в таком же разведении. В процессе исследований один из предложенных спиртовых экстрактов проверяли на фитотоксич-
ность по отношению к проросткам и побегам томатов сорта Ранний 83. Препарат Халотанил в разве-дениях 1:100, 1:500 и 1:1000 (два последних разведения рекомендованы для испытания) проверяли на фитотоксичность по отношению к проросткам и побегам томатов сорта Ранний 83, остро реагирую-щим на токсин гриба Альтернария. Для каждого варианта, включая воду, брали по 50 семян томатов. Семена на одни сутки замачивали в соответствующих растворах, после чего проращивали во влаж-ных камерах. Учитывали энергию прорастания семян и состояние проростков.
Побеги томата (по 5 штук на вариант) помещали в воду и в тестируемые растворы. В течение недели наблюдали за состоянием побегов.
На примере препарата Энотанин изучали влияние экстракта семян на прорастание конидий Аль-тернарии.
Конидии выращивали на влажной фильтровальной бумаге по методике Allen et al. [1]. Для опытов использовали предметные стекла с лункой, помещенные в большую чашку Петри диаметром 16,5 см. В лунки предметных стекол заливали препарат (или растворитель) в соответствующих разведениях, а затем осторожно, легким прикосновением скальпеля, соскабливали конидии патогена. Все предмет-ные стекла с растворами разной концентрации находились в одной чашке Петри, то есть в равных условиях проведения опыта (рис.1).
Для наблюдения за прорастанием конидий в первые 5 часов эксперимента предметные стекла про-сматривали под микроскопом ежечасно, а в период наиболее вероятного начала прорастания конидий (спустя 2 часа после начала эксперимента) – каждые 30 минут. Затем предметные стекла просматри-вали через 24 и 48 часов.
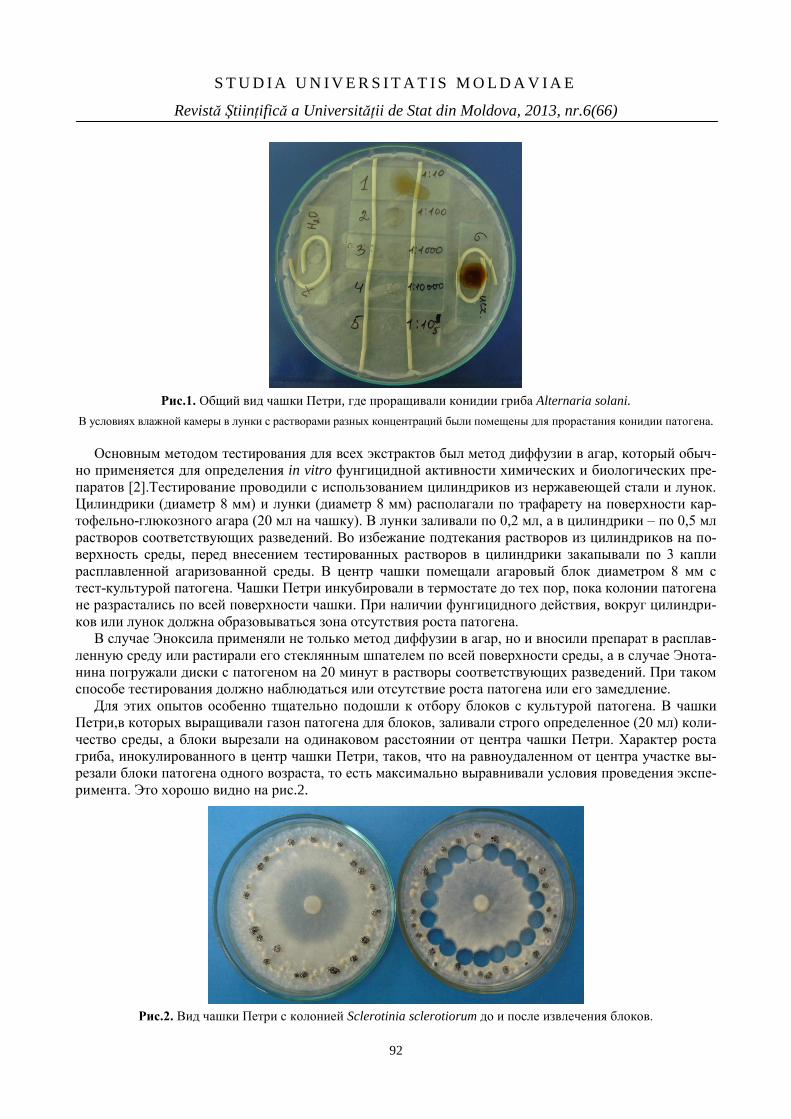
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
92
Рис.1. Общий вид чашки Петри, где проращивали конидии гриба Alternaria solani.
В условиях влажной камеры в лунки с растворами разных концентраций были помещены для прорастания конидии патогена.
Основным методом тестирования для всех экстрактов был метод диффузии в агар, который обыч-
но применяется для определения in vitro фунгицидной активности химических и биологических пре-
паратов [2].Тестирование проводили с использованием цилиндриков из нержавеющей стали и лунок.
Цилиндрики (диаметр 8 мм) и лунки (диаметр 8 мм) располагали по трафарету на поверхности кар-
тофельно-глюкозного агара (20 мл на чашку). В лунки заливали по 0,2 мл, а в цилиндрики – по 0,5 мл
растворов соответствующих разведений. Во избежание подтекания растворов из цилиндриков на по-
верхность среды, перед внесением тестированных растворов в цилиндрики закапывали по 3 капли
расплавленной агаризованной среды. В центр чашки помещали агаровый блок диаметром 8 мм с
тест-культурой патогена. Чашки Петри инкубировали в термостате до тех пор, пока колонии патогена
не разрастались по всей поверхности чашки. При наличии фунгицидного действия, вокруг цилиндри-
ков или лунок должна образовываться зона отсутствия роста патогена.
В случае Эноксила применяли не только метод диффузии в агар, но и вносили препарат в расплав-
ленную среду или растирали его стеклянным шпателем по всей поверхности среды, а в случае Энота-
нина погружали диски с патогеном на 20 минут в растворы соответствующих разведений. При таком
способе тестирования должно наблюдаться или отсутствие роста патогена или его замедление.
Для этих опытов особенно тщательно подошли к отбору блоков с культурой патогена. В чашки
Петри,в которых выращивали газон патогена для блоков, заливали строго определенное (20 мл) коли-
чество среды, а блоки вырезали на одинаковом расстоянии от центра чашки Петри. Характер роста
гриба, инокулированного в центр чашки Петри, таков, что на равноудаленном от центра участке вы-
резали блоки патогена одного возраста, то есть максимально выравнивали условия проведения экспе-
римента. Это хорошо видно на рис.2.
Рис.2. Вид чашки Петри с колонией Sclerotinia sclerotiorum до и после извлечения блоков.

Seria “Ştiinţ e reale şi ale naturii”
Biologie ISSN 1814-3237
93
В опытах использовали исходный раствор и разведения: 1:10; 1:100; 1:1000; 1:10000; 1:100000.
Большие разведения препарата были взяты с целью проследить возможный стимулирующий эффект
низких концентраций препарата.
В качестве тест-объектов использовали следующие фитопатогены:
Sclerotinia sclerotiorum (из огурца) – возбудитель белой гнили;
Botrytis cinerea (из ягод винограда) – возбудитель серой гнили;
Rhizoctonia solani (из сеянцев томата) – один из возбудителей корневых гнилей;
Alternaria solani (из листьев томата) – возбудитель альтернариоза;
Fusarium sp. (из корней земляники и корнеплода сахарной свеклы) – возбудитель фузариозной
гнили.
Выбор указанных патогенов обусловлен тем, что они поражают широкий круг экономически зна-
чимых растений, выращиваемых в открытом и закрытом грунте. Кроме того, они причиняют серьез-
ные потери сельхозпродукции во время хранения.
В разные годы исследований использовали не менее трех фитопатогенов, часто отдавая предпочте-
ние S. sclerotiorum и Rh. Solani как быстрорастущим на питательных средах и дающим ровный газон
на поверхности среды.
Повторность опытов 3-х – 5-кратная.
Результаты учетов обрабатывали статистически с использованием пакета программ Microsoft Office.
Результаты исследований
При фитотоксической оценке не обнаружено различий ни по энергии прорастания семян, ни по
состоянию проростков между опытными и контрольными вариантами.
На побегах томатов в течение недели не выявлено никаких признаков токсикоза, более того, отме-
чено образование корней на побегах. То есть, Халотанил в разведениях 1:100, 1:500 и 1:1000 нефито-
токсичен.
В опыте по изучению действия Энотанина на прорастание конидий Альтернарии получены сле-
дующие результаты, отраженные в таблице 1.
Таблица 1
Прорастание конидий Альтернарии в растворах Энотанина и растворителя
№
п/п Варианты
Время экспозиции
1 час 2 часа 3 часа 4 часа 5 часов 24 часа 48 часов
Э Р Э Р Э Р Э Р Э Р Э Р Э Р
1 Исходный
раствор - - - - - - - - - - - - - -
2 1:10 - - - - - - - - - - - - *)
- - *)
3 1:100 - - - - - ± - + + + + + + +
4 1:1000 - - - - ± ± + + + + + + + +
5 1:10000 - - - - ± ± + + + + + + + +
6 1:100000 - — - — ± — + — + — + — + —
7 Вода - - - + + + +
Условные обозначения: Э – Энотанин; Р – растворитель; - – конидии не проросли; + – массовое прораста-
ние конидий, ростковая трубка превышает длину конидий; ± – единичные конидии наклюнулись, ростковые
трубки не превышают длину конидий; — - вариант отсутствовал; - *)
– массовое прорастание конидий, но рост-
ковая трубка не превышала ширины конидий.
В первые 2 часа эксперимента ни в одном из вариантов, включая контроль, проросших конидий не
обнаружено, что вполне логично, так как для прорастания конидий в благоприятных температурных
условиях требуется 2-3 часа. Спустя 3 часа в воде конидии не проросли, а в вариантах с большим раз-
ведением и растворителя, и Энотанина появились ростковые трубки. То есть проявился некоторый
стимулирующий эффект по сравнению с контролем, причем в растворителе стимуляция отмечена в
разведении 1:100, а в растворе Энотанина такого же разведения прорастание конидий не отмечено.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
94
Однако стимулирующий эффект был непродолжительным. После 4-часовой выдержки конидии нор-
мально проросли в воде и больших разведениях и Энотанина, и растворителя. В разведении 1:100
раствор Энотанина тормозил прорастание конидий, а в таком же разведении растворителя конидии
проросли. После 5-часовой выдержки в разведениях 1:100 – 1: 100000 конидии проросли и в воде, и в
растворах Энотанина, и в растворителе. После 48-часовой выдержки конидии не прорастали в исход-
ном растворе Энотанина и растворителе, а в разведении 1:10 не прорастали в растворе Энотанина, а
образовали небольшие ростковые трубки в 10-кратноразбавленном пропиленгликоле (90% пророс-
ших конидий, но их ростковые трубки не превышали ширину конидий или равнялись ей).
Таким образом, можно отметить, что в течение длительного времени (более 4 часов) Энотанин
сдерживал прорастание конидий только в исходном препарате и при 10-кратном разведении.
Раствор Энотанина влиял на смачиваемость конидий, которая в воде была хуже, чем в растворе
Энотанина. В каплях раствора Энотанина (всех разведений) конидии распределялись по всей поверх-
ности раствора, а в воде они скапливались по периметру капли. Отмечено также, что цепочки конидий
сохранялись во всех вариантах кроме исходного раствора. В исходном растворе не было обнаружено
ни одной цепочки конидий: все конидии располагались отдельно друг от друга или кучками; в разве-
дении 1:10 этого явления уже не отмечено (рис.3 и 4).
Рис.3. Внешний вид конидий в цепочках
с ростковыми трубками в воде.
Рис.4. Конидии без цепочек в исходном
растворе Энотанина.
Использование метода диффузии в агар не показало четкого фунгицидного или фунгистатического
эффекта растворов Энотанина. На рис.5 представлен результат действия Энотанина на культуру
Fusarium sp. (из земляники). Только в позициях 1 и 2 (в первой несколько больше) прослеживалось
некоторое торможение роста патогена, но оно было очень кратковременным.
Рис.5. Действие растворов Энотанина на культуру Fusarium sp.
Слева – вид чашки сверху; справа – со дна чашки.
Условные обозначения: 1 – разведение 1:10; 2 – разведение 1:100; 3 – разведение 1:1000;
4 – разведение 1:10000; 5 – разведение 1:100000; 6 – вода.

Seria “Ştiinţ e reale şi ale naturii”
Biologie ISSN 1814-3237
95
На рис.6 показан результат действия растворов Энотанина на патоген Sclerotinia sclerotiorum. И в
данном случае нет стерильной зоны вокруг лунок , то есть мы не отмечаем фунгицидного действия
Энотанина на рост гриба.
Рис.6. Действие растворов Энотанина на культуру Sclerotinia sclerotiorum.
Условные обозначения: 1 – разведение 1:10; 2 – разведение 1:100; 3 – разведение 1:1000;
4 – разведение 1:10000; 5 – разведение 1:100000; 6 – вода.
Логичным является отсутствие фунгицидного действия растворителя пропиленгликоля в разных
разведениях (в лунках по 0,2 мл раствора), тест-культура – склеротиния.
Таким образом, метод диффузии в агар не показал выраженного фунгицидного действия Энотанина
и в исходном виде, и в разведениях.
Для сравнения приведем результат испытания действия Энотанина и бактерий-антагонистов на
гриб Fusarium sp. (рис.7). Просматривается наличие стерильных зон в позициях 2, 3, 5 и 6 (блоки
бактерий-антагонистов с выраженным антифунгальным действием) и отсутствие их в позициях 1 и 4.
Колония патогена приобрела вытянутую форму, максимально приближенную к месту внесения
Энотанина.
Рис.7. Сравнительное действие Энотанина и бактерий-антагонистовна Fusarium sp. (Вид сверху чашки (слева) и с подложки колоний (справа).
Условные обозначения: 1 – исходный раствор Энотанина; 4 – Энотанин в разведении 1 : 10;
2,3,5 и 6 – блоки разных культур бактерий-антагонистов.
При непосредственном контакте патогена с растворами Энотанина или растворителя (замачивание
блоков с культурой склеротинии в течение 20 минут в разных концентрациях Энотанина) не выявило
какого-либо действия на патоген (рис.8 а,б).
«Лысинка» непосредственно вокруг блока патогена характерна для роста патогена на агаризован-
ной среде (рис.2).
Только в исходном растворе Эноксила (без разведения) в отношении S. sclerotiorum при использо-
вании метода диффузии в агар нами был отмечен очень незначительный, но стойкий фунгистатиче-
ский эффект, сохранявшийся около месяца. В разведении 1:1 этот эффект был едва заметен. В отно-
шении B. Cinerea и R. solani не обнаружено ни фунгицидного, ни фунгистатического действия.
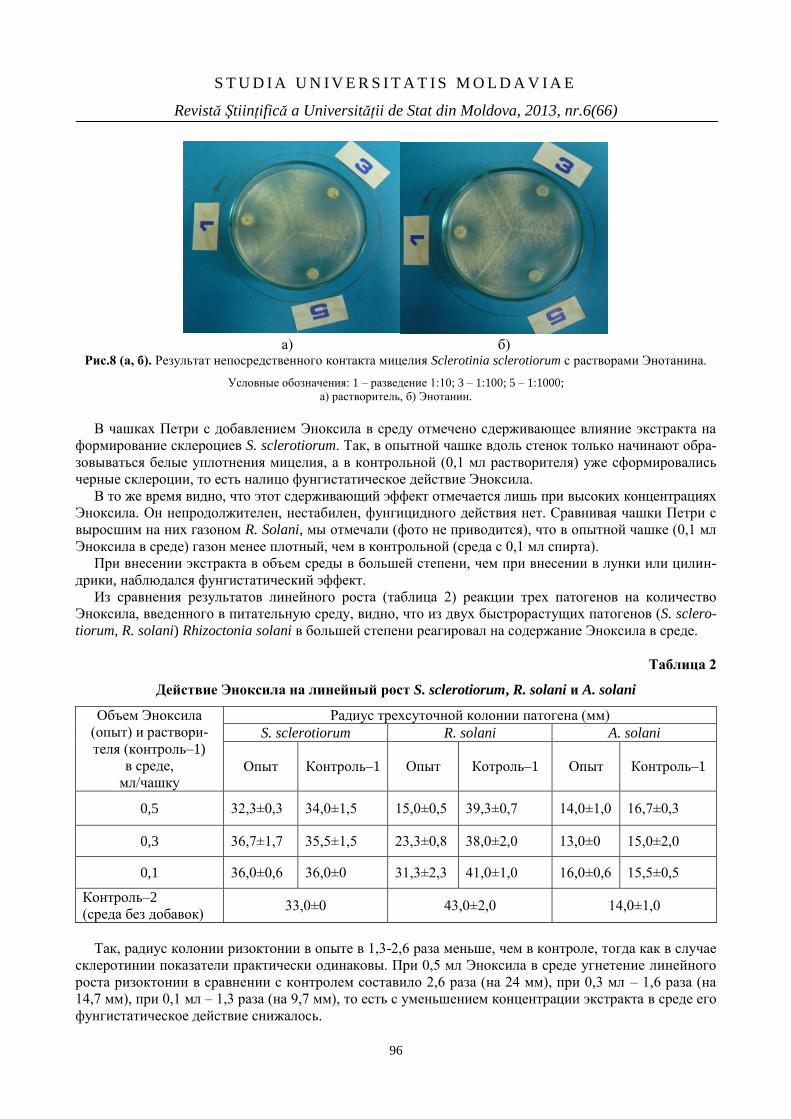
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
96
а) б) Рис.8 (а, б). Результат непосредственного контакта мицелия Sclerotinia sclerotiorum с растворами Энотанина.
Условные обозначения: 1 – разведение 1:10; 3 – 1:100; 5 – 1:1000;
а) растворитель, б) Энотанин.
В чашках Петри с добавлением Эноксила в среду отмечено сдерживающее влияние экстракта на
формирование склероциев S. sclerotiorum. Так, в опытной чашке вдоль стенок только начинают обра-
зовываться белые уплотнения мицелия, а в контрольной (0,1 мл растворителя) уже сформировались
черные склероции, то есть налицо фунгистатическое действие Эноксила.
В то же время видно, что этот сдерживающий эффект отмечается лишь при высоких концентрациях
Эноксила. Он непродолжителен, нестабилен, фунгицидного действия нет. Сравнивая чашки Петри с
выросшим на них газоном R. Solani, мы отмечали (фото не приводится), что в опытной чашке (0,1 мл
Эноксила в среде) газон менее плотный, чем в контрольной (среда с 0,1 мл спирта).
При внесении экстракта в объем среды в большей степени, чем при внесении в лунки или цилин-
дрики, наблюдался фунгистатический эффект.
Из сравнения результатов линейного роста (таблица 2) реакции трех патогенов на количество
Эноксила, введенного в питательную среду, видно, что из двух быстрорастущих патогенов (S. sclero-
tiorum, R. solani) Rhizoctonia solani в большей степени реагировал на содержание Эноксила в среде.
Таблица 2
Действие Эноксила на линейный рост S. sclerotiorum, R. solani и A. solani
Объем Эноксила
(опыт) и раствори-
теля (контроль–1)
в среде,
мл/чашку
Радиус трехсуточной колонии патогена (мм)
S. sclerotiorum R. solani A. solani
Опыт Контроль–1 Опыт Котроль–1 Опыт Контроль–1
0,5 32,3±0,3 34,0±1,5 15,0±0,5 39,3±0,7 14,0±1,0 16,7±0,3
0,3 36,7±1,7 35,5±1,5 23,3±0,8 38,0±2,0 13,0±0 15,0±2,0
0,1 36,0±0,6 36,0±0 31,3±2,3 41,0±1,0 16,0±0,6 15,5±0,5
Контроль–2
(среда без добавок) 33,0±0 43,0±2,0 14,0±1,0
Так, радиус колонии ризоктонии в опыте в 1,3-2,6 раза меньше, чем в контроле, тогда как в случае
склеротинии показатели практически одинаковы. При 0,5 мл Эноксила в среде угнетение линейного
роста ризоктонии в сравнении с контролем составило 2,6 раза (на 24 мм), при 0,3 мл – 1,6 раза (на
14,7 мм), при 0,1 мл – 1,3 раза (на 9,7 мм), то есть с уменьшением концентрации экстракта в среде его
фунгистатическое действие снижалось.

Seria “Ştiinţ e reale şi ale naturii”
Biologie ISSN 1814-3237
97
Выводы
1. Препарат Халотанил в рекомендуемых разведениях 1:100 и 1:500 нефитотоксичен по отноше-
нию к проросткам и побегам томата сорта Ранний 83.
2. Исходный раствор Энотанина и растворитель в разведениях 1:10 сдерживали прорастание кони-
дий Альтернарии более 2-х суток. При больших разведениях конидии прорастали после двухчасовой
экспозиции.
3. Метод диффузии в агар при тестировании спиртовых экстрактов виноградных семян не обнару-
живал образования зон отсутствия роста тест-культур. Для тестирования экстрактов более подходя-
щим является способ введения экстрактов в питательную среду.
4. Результаты тестирования зависят от тест-культуры и объема экстракта в питательной среде.
Исследуемые экстракты сдерживали рост патогенов по типу фунгистатического действия. Фунгицид-
ный эффект ни по одному из экстрактов не обнаружен.
Библиография:
1. ALLEN, S.J., BROWN, J.F., KOCHMAN, J.K. Production of inoculum and field assessment of Alternaria helianthi
on sunflower. In: Plant disease, 1983, v.67, p. 665-668.
2. ЕГОРОВ, Н.Е. Основы учения об антибиотиках. Москва: Высшая школа, 1969. 479 с.
3. ЗУЕВА, Т.А. Разработка малоотходной технологии переработки семян винограда и получение на их основе
лекарственных и косметических средств: Автореф. канд. дисс. Пятигорск, 2004.
4. НАСТАС, Т., ЕЛИСОВЕЦКАЯ, Д., ГЛАДКАЯ, А., ОДОБЕСКУ, В., ХАРГЕЛ, П. Экстракт из семян Vitisvinifera
в защите растений. В: Биологическая защита растений – основа стабилизации экосистем. Материалы докла-
дов Международной научно-практической конференции, ВНИИБЗР, Краснодар, 2008, вып.5, с.363-365.
Prezentat la 07.10.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
98
PROCEDEE BIOTEHNOLOGICE DE PRODUCERE A PREPARATELOR VIRALE
PENTRU COMBATEREA DEFOLIATORULUI OMIDA PĂROASĂ A DUDULUI
(Hyphantria cunea Drury)
Aurelia STÎNGACI
Institutul Protecţia Plantelor şi Agriculturii Ecologice al AŞM
În multe ţări au fost puse la punct preparate virale care se aplică în practică pentru reducerea numărului insectelor
nocive. Articolul conţine rezultatele aplicării preparatelor virale şi a procedeelor biotehnologice de producere a prepa-
ratului viral Virin-ABB-3 pentru protejarea culturilor agricole, ornamentale şi forestiere împotriva Hyphantria cunea
Drury în condiţii de laborator şi de câmp. Preparatul este bazat pe virusurile poliedrozei (VPN) şi granulozei (VG), cu
acţiune cumulativă şi sinergistă. Un element important în procesul tehnologic al preparatelor biologice este dezvoltarea
unor tehnici pentru creşterea în masă a larvelor de dăunători în serie continuă, care oferă materialul biologic viral pe tot
parcursul anului.
Cuvinte-cheie: Hyphaniria cunea Drury, insecticide virale, baculovirusuri, Virin-ABB-3, VG, VPN.
BIOTEHNOLOGICAL PRODUCTION PROCESSES VIRAL PPREPARATION FOR CONTROL
OF THE FALL WEBWORM MOTH (Hyphantria cunea Drury)
In many countries there were established viral preparations applied in practice in order to reduce the number of pests.
Report contains the results of application viral preparation and biotechnological processes for the production of biological
preparation Virin-ABB-3 for protection of agricultural, ornamental and forest crops of Hyphantria cunea Drury in labora-
tory and field conditions. The preparation is based on viruses of nuclear polyhedrosesis and granuloses with cumulative
and synergetic action. One important element in the technological process of biological preparations is to develop
techniques for increasing the mass of larvae of pests in continuous series, which gives the viral biological material all
round year.
Keywords: Hyphantria cunea Drury, viral insecticides, baculovirus, Virin-ABB-3, VG, VPN.
Introducere Necesitatea combaterii organismelor dăunătoare este determinată de faptul că culturile agricole sunt ata-
cate de o gamă imensă de organisme dăunătoare. Pierderile de recoltă constituie 25-30%, iar uneori depăşesc 40-50% sau organismele dăunătoare totalmente compromit culturile agricole. La nivel global, pierderile re-
coltei constituie 50 tril.$, iar în Republica Moldova depăşesc 2 mild. lei [9]. Omida păroasa a dudului (H. cunea), care a pătruns în Europa din America, a devenit în scurt timp un
dăunător periculos al culturilor agricole, decorative şi forestiere. În prezent, formarea arealului secundar este departe de a fi complet. Fitofagul se dezvoltă treptat şi ocupă noi teritorii. De exemplu, el a apărut recent în ţările din Asia Centrală, spre exemplu în 2003 în Iran, iar în prezent acesta a însuşit toată partea nordică a ţării; în anul 2005 a fost depistat în Noua Zeelanda. Astăzi el continuă să se extindă şi în alte regiuni nordice
ale Europei 2,4,6 . Atacul, uneori catastrofal, al H. cunea, a dus la cercetări multidirecţionale în echipă, cu prioritate pentru a
cunoaşte biologia insectei, patologia omizilor, pupelor şi modul de întrebuinţare a biopreparatelor de comba-
tere a lor. În scopul reducerii impactului invaziilor organismelor dăunătoare se elaborează tehnologii de combatere
şi minimalizare a randamentului culturilor agricole, care necesită prezentarea unei game largi de mijloace biologice alternative celor chimice, asigurând indici biologici sporiţi. Aceasta necesită elaborarea metodelor
de selecţie şi ameliorare a eficacităţii biologice, modificând indicii dezvoltării individuale atât a plantelor-gazde, cât şi capacităţile agenţilor utilizaţi pentru combaterea organismelor dăunătoare [1,5].
Din toate preparatele ecologice utilizate în scopul reglării densităţii organismelor nedorite cele mai efi-ciente sunt preparatele virale. Ele acţionează selectiv şi reduc densitatea organismelor nedorite până sub pra-
gul dăunării economice. Preparatele se transmit din generaţie în generaţie şi reglează densitatea organismelor timp de mai mulţi ani de zile, fiind rentabile din punct de vedere economic, deoarece pot declanşa epidemii
virale, care se menţin o perioadă îndelungată 8,7 .

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
99
Pentru obţinerea şi procesarea produselor ecologice este necesară organizarea şi producerea largă a mijloa-
celor biologice de protecţie a plantelor. Printre numeroasele mijloace microbiologice de protecţie a plantelor
baculovirusurile ocupă un loc deosebit, fiind utilizate pentru combaterea dăunătorilor. Rezultate interesante
se obţin la aplicarea biopreparatelor pe bază de virusuri ai insectelor dăunătoare, care provoacă epizootii pe
arealuri mari cu unele legităţi de manifestare a efectului de postacţiune [4,7].
Dat fiind faptul că extinderea arealului lui H. cunea în Republica Moldova a produs pagube materiale în-
semnate, diferite aspecte ale biologiei şi combaterii dăunătorului au constituit subiecte ale cercetărilor noastre
desfăşurate între anii 2004 şi 2013, cercetări ale căror rezultate le prezentăm în cele ce urmează.
Material şi metode
Stocul iniţial de adulţi a fost obţinut din larve H. cunea care au fost colectate din diferite regiuni ale munici-
piului Chişinău şi din diferite localităţi ale Moldovei; acestea au fost aduse în laborator şi crescute pe hrană
naturală (frunze de arţar). Selecţia adulţilor folosiţi în experimentele ulterioare s-a făcut pe baza unor caracte-
ristici morfofiziologice, cum ar fi mărimea şi aspectul normal. În vederea elucidării acestui aspect, am com-
parat variaţia dimensională a adulţilor obţinuţi din larve crescute pe hrană naturală (frunze de arţar) cu cea a
adulţilor obţinuţi din larve hrănite cu mediu, precum şi apariţia diferitelor anomalii la adulţii din ambele va-
riante. Măsurătorile privind lungimea corpului şi amplitudinea aripilor au fost efectuate după moartea adulţi-
lor. De asemenea, au fost stabiliţi parametrii de bază privind prolificitatea şi fertilitatea femelelor crescute în
condiţii controlate, care trebuie să fie mai mici cu maximum 20% decât ale femelelor din câmp. Acumularea
biomasei baculovirale s-a efectuat cu folosirea populaţiei naturale de insecte în viviere speciale. În condiţii
de laborator le-am repartizat pe buchete de plante. Observaţiile efectuate zilnic au urmărit momentul începerii/
finalizării depunerii pontei, locul şi modul de depunere a pontei, numărul de ponte depuse.
S-a experimentat biopreparatul Virin-ABB-3 realizat de către IPPAE al AŞM. Larvele bolnave au fost
determinate după simptomele respective, apoi cu ajutorul microscopului. Infectarea larvelor s-a efectuat cu
suspensii virale cu titrul de 106 SPVC la un individ. Observaţiile s-au efectuat începând cu ziua a treia după
infecţie. Eficienţa preparatului viral s-a determinat după formula Abbot, care prevede mortalitatea naturală a
insectelor.
Testarea suşelor identificate şi recombinate de VPN şi VG ale insectelor de H. cunea s-a efectuat pe larve
specifice ale insectelor de vârsta a doua, crescute pe medii selective de cultură. Pentru aceasta s-a aplicat
metoda diluţiilor succesive de la 10 până la 1000 particule virale pentru o larvă. Larvele au fost hrănite cu
mediu infectat, ulterior fiind menţinute la temperatura de 26-28ºC. Doza letală s-a determinat până la 200 de
ore din momentul infectării. Timpul letal s-a determinat la infectarea larvelor cu doză sporită de particule
virale (250 poliedre la o larvă). Testarea în condiţii de laborator şi în câmpul de experienţă s-a efectuat în
4 repetiţii rendomizate respectiv, în conformitate cu cerinţele generale de acest gen [10,11].
Prepararea incluziunilor baculovirotice. Larvele moarte din cauza infecţiei virale au fost macerate şi sus-
pendate în soluţie de SDS de 0,1% care conţine câte 50 mg/l de mortalitate. După filtrare poliedrele s-au se-
dimentat prin centrifugare la 5000 tur/min, iar granulele – la 12000 tur/min. După 3-5 cicluri de spălare în
această soluţie, incluziunile virotice au fost resuspendate într-o soluţie de 0,6% de NaCl, care conţine meteorat.
Pregătirea incluziunilor baculovirale a fost efectuată conform documentelor pentru preparatele virale,
prevăzute de Regulamentul pentru pregătirea loturilor de preparate sus-numite. În calitate de suport au fost
folosite resturi vegetale, şlam, melasă, făină de grâu. S-a efectuat forma vastă a preparatelor baculovirale.
Rezultate şi discuţii Rezultatele cercetărilor noastre sunt rezumate şi sintetizate în tabele şi figuri.
Cercetările efectuate au scos în evidenţă că infestarea cea mai puternică (100%) a fost înregistrată la arbo-
rii ornamentali – dud Morus nigra f. şi arţar Acer negundo, apoi la o serie de pomi fructiferi – măr (66,1%),
nuc (55,4%), cireş (49,5%), prun (39,4%), gutui (33,7%), păr (30,7%) etc. (Tab.1).
Pentru a trece la extinderea combaterii virologice în protecţia plantelor decorative şi pădurilor, este necesar
să fie elaborate tehnici de creştere în masă a larvelor de dăunători în serii continue, care să permită obţinerea
de material biologic virozat pe tot timpul anului. Creşterile pe hrană naturală permit obţinerea de material
biologic numai în cursul sezonului de vegetaţie şi numai în perioadele când condiţiile climatice sunt favora-
bile dezvoltării larvelor. În general, aceste creşteri se realizează pe perioade scurte, care coincid cu existenţa
în natură a larvelor şi a unei hrane accesibile (frunze tinere).
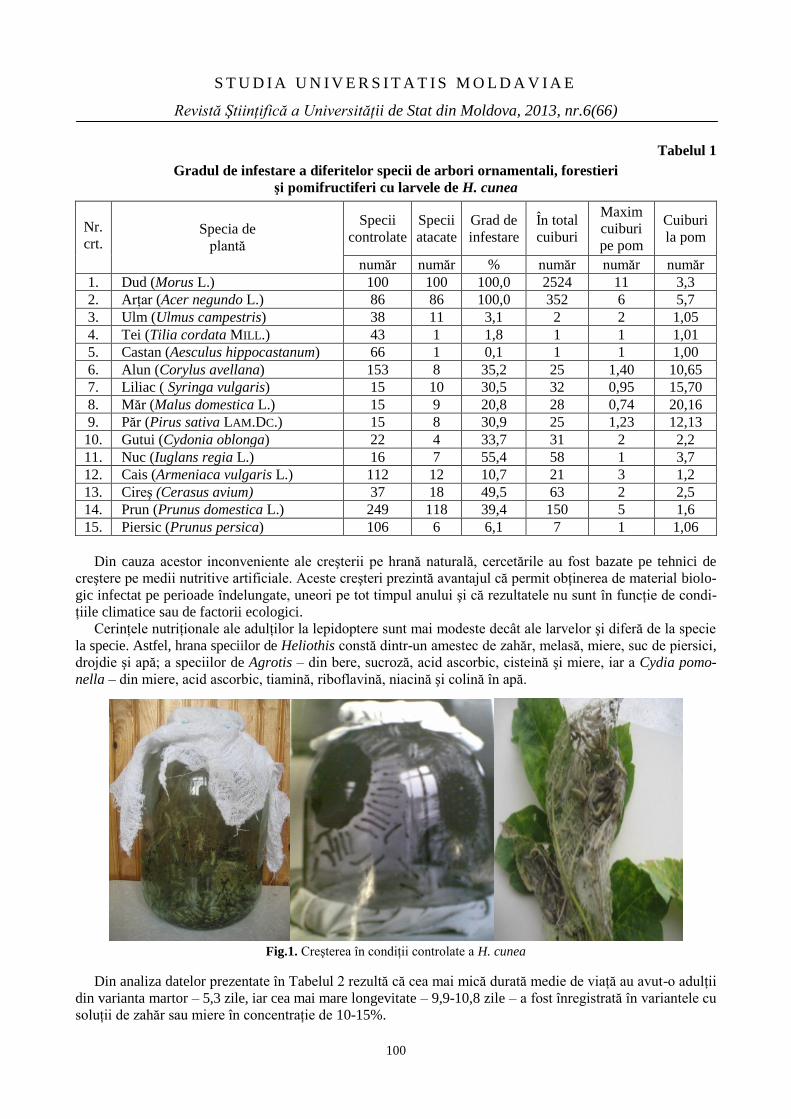
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
100
Tabelul 1
Gradul de infestare a diferitelor specii de arbori ornamentali, forestieri
şi pomifructiferi cu larvele de H. cunea
Nr.
crt. Specia de
plantă
Specii
controlate
Specii
atacate
Grad de
infestare
În total
cuiburi
Maxim
cuiburi
pe pom
Cuiburi
la pom
număr număr % număr număr număr
1. Dud (Morus L.) 100 100 100,0 2524 11 3,3
2. Arţar (Acer negundo L.) 86 86 100,0 352 6 5,7
3. Ulm (Ulmus campestris) 38 11 3,1 2 2 1,05
4. Tei (Tilia cordata MILL.) 43 1 1,8 1 1 1,01
5. Castan (Aesculus hippocastanum) 66 1 0,1 1 1 1,00
6. Alun (Corylus avellana) 153 8 35,2 25 1,40 10,65
7. Liliac ( Syringa vulgaris) 15 10 30,5 32 0,95 15,70
8. Măr (Malus domestica L.) 15 9 20,8 28 0,74 20,16
9. Păr (Pirus sativa LAM.DC.) 15 8 30,9 25 1,23 12,13
10. Gutui (Cydonia oblonga) 22 4 33,7 31 2 2,2
11. Nuc (Iuglans regia L.) 16 7 55,4 58 1 3,7
12. Cais (Armeniaca vulgaris L.) 112 12 10,7 21 3 1,2
13. Cireş (Cerasus avium) 37 18 49,5 63 2 2,5
14. Prun (Prunus domestica L.) 249 118 39,4 150 5 1,6
15. Piersic (Prunus persica) 106 6 6,1 7 1 1,06
Din cauza acestor inconveniente ale creşterii pe hrană naturală, cercetările au fost bazate pe tehnici de
creştere pe medii nutritive artificiale. Aceste creşteri prezintă avantajul că permit obţinerea de material biolo-
gic infectat pe perioade îndelungate, uneori pe tot timpul anului şi că rezultatele nu sunt în funcţie de condi-
ţiile climatice sau de factorii ecologici.
Cerinţele nutriţionale ale adulţilor la lepidoptere sunt mai modeste decât ale larvelor şi diferă de la specie
la specie. Astfel, hrana speciilor de Heliothis constă dintr-un amestec de zahăr, melasă, miere, suc de piersici,
drojdie şi apă; a speciilor de Agrotis – din bere, sucroză, acid ascorbic, cisteină şi miere, iar a Cydia pomo-
nella – din miere, acid ascorbic, tiamină, riboflavină, niacină şi colină în apă.
Fig.1. Creşterea în condiţii controlate a H. cunea
Din analiza datelor prezentate în Tabelul 2 rezultă că cea mai mică durată medie de viaţă au avut-o adulţii
din varianta martor – 5,3 zile, iar cea mai mare longevitate – 9,9-10,8 zile – a fost înregistrată în variantele cu
soluţii de zahăr sau miere în concentraţie de 10-15%.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
101
Tabelul 2
Influenţa hranei asupra longevităţii adulţilor de H. cunea crescuţi în condiţii controlate
Varianta (soluţii nutritive)
Durata de supravieţuire,
zile
Minimă-maximă Media
Martor 2-7 5,3
5% zahăr în apă 3-9 6,4
10% zahăr în apă 4-12 9,9
15% zahăr în apă 3-14 10,5
5% miere în apă 4-8 6,7
10% miere în apă 3-13 10,2
15% miere în apă 5-14 10,8
Miere (5%), zahăr (5%),
acid ascorbic (1%) 5-13 9,3
Dezvoltarea oricărui lepidopter este strâns legată de calitatea hranei administrate larvelor. Boguleanu şi
colab. (2005) ajung la concluzia că valorile ponderale şi dimensionale ale omizii păroase a dudului variază
mult în funcţie de planta-gazdă: dimensiunea corpului la femele variază între 14,2 mm, când larvele sunt hră-
nite cu dud, şi 8,32 mm, când acestea sunt hrănite cu tei, iar la masculi variaţia este cuprinsă între 11,36 mm
(în cazul hrănirii cu dud) şi 7,98 mm (în cazul hrănirii cu tei). Corelând aceste date cu procentul de adulţi
anormali (4,8 faţă de 40,3 în variantele menţionate) autorii consideră că, deşi H. cunea este o insectă polifagă,
dezvoltarea normală se asigură pe plantele-gazdă preferate. Beratlief şi colab. (2005), experimentând creşte-
rea omizii păroase a dudului pe câteva medii artificiale, ajung la concluzia că proporţia de adulţi anormali în
cazul hrănirii larvelor cu medii sintetice variază între 6,3% şi 14,5%, în timp ce la hrănirea cu frunze de dud
procentul de adulţi anormali este de 6,8% [3].
Reieşind din rezultatele cercetărilor efectuate (Tab.3), se poate constata că în varianta creşterii larvelor pe
hrană naturală lungimea corpului la femele este în medie de 14,3 mm, iar anvergura aripilor atinge valori
medii de 34,57 mm. La masculi lungimea corpului are valori medii de 12,8 mm şi anvergura medie a aripilor
de 33,67 mm. În varianta creşterii larvelor pe mediu artificial adulţii au mărimi medii ceva mai mici: feme-
lele au lungimea medie a corpului de 12 mm şi anvergura aripilor de 29,23 mm, iar masculii 11,2 mm media
lungimii corpului şi 27,7 mm anvergura aripilor. În ce priveşte procentul de adulţi anormali în varianta creş-
terii pe mediu artificial, acesta a avut o valoare de 7,33% în timp ce în cazul creşterilor pe hrană naturală a
avut valoarea de 5,67 %.
Tabelul 3
Caracteristicile adulţilor de H. cunea obţinuţi în creşterile pe diferite substraturi
Varianta
Repetiţia
Lungimea corpului,
mm
Anvergura aripilor,
mm
Adulţi cu anomalii
%
♀ ♂
0+
♂
VI – adulţi obţinuţi
prin creşterea larvelor
pe hrană naturală
R1 13,3 12,3 34,7 29,7 6
R2 13,7 11,7 35,3 28,7 4
R3 13,3 12,0 33,7 29,3 7
MEDIA 13,43 12,0 34,57 29,23 5,67
V2 – adulţi obţinuţi
prin creşterea larvelor
pe mediu artificial
R1 12,7 11,3 33,7 27,7 8
R2 13,0 11,0 34,0 26,7 5
R3 12,7 11,3 33,3 28,7 9
MEDIA 12,8 11,2 33,67 27,7 7,33
Un număr de 10 ponte, depuse de femelele obţinute prin creşterea larvelor pe medii artificiale în laborator,
au fost prelevate aleator din cuştile de creştere ale adulţilor. Pontele care au reprezentat repetiţiile în cazul
cercetărilor din această fază au fost numărate, introduse în vase de incubaţie şi supuse observaţiilor zilnice.
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
102
Cercetările, în cazul creşterilor pe mediu artificial, au relevat următoarele (Tab.4). Femela depune una –
maximum două ponte numărând între 408 şi 1514 ouă, cu o medie de 630,7 ouă/femelă. Ovipoziţia a avut o
perioadă cuprinsă între două şi şase zile, cu o medie de 3,2 zile.
Suportul pentru ovipoziţie preferat de femelele de H. cunea colectate din câmp este reprezentat de frunzele
introduse în acest scop în cuştile de creştere. În ce priveşte femelele de H. cunea obţinute prin creşterile în
generaţii succesive în condiţii de laborator, se poate spune că acestea nu au o preferinţă anume referitor la
suportul de depunere a pontelor.
Pentru stabilirea duratei incubaţiei şi viabilităţii ouălor la populaţia de H. cunea colectată din câmp, cerce-
tările efectuate în condiţiile IPPAE al ASM s-au materializat cu următoarele rezultate. Imediat după ecloziune
larva neonată se hrăneşte parţial cu corionul oului. Deşi ecloziunea unei singure ponte poate continua timp
de 3 zile, cea mai mare parte a larvelor neonale apar în câteva ore. Procentul de ecloziune a ouălor a fost de
80,3%, iar durata incubaţiei de 4 până la 8 zile (Tab.4).
Tabelul 4
Viabilitatea pontelor depuse de femele de H. cunea din câmp
Repetiţia Numărul de ouă/pontă Intervalul de ecloziune,
zile
Ecloziune,
%
1 544 5-7 73,90
2 398 5-7 80,90
3 346 6-7 86,41
4 789 6-8 69,71
5 518 5-7 77,61
6 503 6-7 84,49
7 415 4-6 73,01
8 1034 7-8 85,30
9 420 5-6 79,76
10 312 6-7 92,30
MEDIA 471 - 80,34
Cercetările privind factorul de nutriţie, în special calitatea hranei în stimularea îmbolnăvirii cu viruşi, au
început cu mult timp, referindu-se la viermele de mătasă (Bombyx mory). Astfel, observaţiile efectuate cu
preparatul Virin-ABB-3 scot în evidenţă că schimbarea plantei-gazdă de către omizile H. cunea, în timpul
dezvoltării lor, poate duce la apariţia bolii poliedrice. Omizile defoliatorului s-au hrănit în condiţii de labora-
tor cu frunze de diferite specii (dud, arţar, salcie, plop, tei, măr, păr, nuc, cais, prun, cireş şi vişin). Experi-
mentările au fost efectuate în două variante diferite, şi anume: cu frunze tratate cu poliedre virale şi cu frunze
netratate. Rezultatele acestor experimentări conduc la concluzia că rezistenţa cea mai ridicată la infecţia
virală au dovedit-o omizile hrănite cu frunze de dud. Rezultă deci că natura hranei influenţează direct starea
fiziologică a omizilor şi influenţează sensibilitatea lor faţă de virusul poliedrozei nucleare.
Experienţele de combatere pe scară largă a altor omizi defoliatoare cu ajutorul biopreparatului Virin-ABB-3
cu omizi contaminate (larve) de H. cunea pe diferite specii de plante cu aceeaşi concentraţie şi acelaşi număr
de omizi, precum şi eficacitatea biologică a devenit o necesitate pentru determinarea procentului de mortali-
tate. Rezultatele experimentelor sunt prezentate în Tabelul 5. În ceea ce priveşte mortalitatea omizilor pe VG
2011 la a 15-a zi a fost de 78-82%, iar pe VPN de 74-80%. Dar, în opţiuni au fost folosite ambele virusuri –
VG şi VPN (1:1), caz în care mortalitatea a crescut de la 88% până la 92%. Eficienţa biologică a suşelor era
la nivelul de 75-92%.
Materialul a fost utilizat pentru pregătirea preparatului Virin-ABB-3. Suşele reînnoite de VPN şi VG din
a.2011 au fost folosite pentru obţinerea materialului viral şi în experimentele de schimbare a tehnologiilor de
obţinere a preparatelor virale pe baza acestor suşe la prima şi a doua generaţie de H. cunea în anul 2012. Cerce-
tările subliniază faza critică a H. cunea şi vor fi foarte utile pentru gestionarea tratamentelor cu baculovirusuri.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
103
Tabelul 5
Gradul de infectare a H. cunea cu baculovirus nou. Hrană naturală frunze de dud
Variantele Repetiţiile Nr.
de larve
Vârsta
larvelor
Concentraţia
poliedre/ml
Ziua mortalităţii
7 15
Nr.
de larve %
Nr. de
larve %
VG 2012 I 50 II-III 105 24 48,0 39 78,0
VG 2012
VPN 2012
II
I
50
50
II-III
II-III
105
105
25
26
50,0
52,0
41
37
82,0
74,0
VPN 2012 II 50 II-III 105 27 54,0 40 80,0
VPN±VG
(1:1) I 50 II-III 10
5 28 56,0 46 92,0
VPN±VG
(1:1) II 50 II-III 10
5 28 56,0 44 88,0
CONTROL I 50 II-III 105 0 - 1 4,0
CONTROL II 50 II-III 105 0 - 2 6,6
Rezultatele obţinute, prezentate în Figura 2, pun în evidenţă că larvele sunt cu atât mai sensibile la infec-
ţie, cu cât stadiul expus este mai precoce. Acest aspect, descris de numeroşi autori, are o importanţă deose-
bită când se pune problema utilizării virusului pentru limitarea densităţii populaţiilor de insecte dăunătoare.
Dacă larvele sunt infectate la vârste mici, se înregistrează procente ridicate de mortalitate (97,3% la prima
vârstă şi 94% la vârsta a doua), în timp ce la vârsta a cincea mortalitatea este foarte redusă (7,5%), apropiată
de valoarea mortalităţii înregistrate în cazul larvelor care au fost hrănite cu hrană netratată cu baculovirusuri.
În ceea ce priveşte aspectul urmărit de noi, în vederea obţinerii de biomasă virală, infectarea la vârste prea
mici sau prea mari nu corespunde scopului propus, cele mai bune rezultate din punct de vedere cantitativ
fiind obţinute prin infectarea larvelor la vârsta a treia.
Fig.2. Mortalitatea produsă de la infecţia cu virusul granulozei.
Concluzii
Mijloacele microbiologice contemporane, inclusiv preparatele baculovirale, constituite din agenţi biolo-
gici naturali, se caracterizează prin indicatori mai mici ai calităţii, ceea ce necesită ameliorarea permanentă
a acestora. Pentru obţinerea biomasei baculovirale eficiente au fost propuse procedee eficiente de sporire a
indicilor de activitate, care reprezintă o pârghie eficientă pe calea elaborării insecticidelor virale. Urmărirea
corelaţiei dintre densitatea larvelor, procentul de ecloziune şi de obţinere a adulţilor (randamentul de creştere)
în două sisteme de creştere (individual şi în grup) a reliefat faptul că ambele sisteme au avantaje şi dezavan-
taje. Creşterea în sistemul individual asigură procente mai mari de obţinere a adulţilor (peste 80%, compa-
rativ cu creşterea în grup, în care procentul a fost de 46-48%), cu menţinerea larvelor în vasele de creştere
97,3
8
94
5
75,3
414,7
5 7,3 30
20
40
60
80
100
mo
rtalita
tea,%
L1 L2 L3 L4 L5
V1 V2 V3 V4 V5
Vîrsta larvelor,Varianta
Mortalitatea medie produsă de virusul Granuloze % Mortalitatea înregistrată în lotul martor %

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
104
până la obţinerea adulţilor (în sistemul de creştere în grup fiind necesar transferul lor repetat, pe măsura creş-
terii în dimensiuni, ceea ce presupune mai mult timp şi manopere suplimentare). Comparând cerinţele trofice
ale adulţilor de H. cunea cu cele ale altor specii de lepidoptere, s-a constatat că aceştia nu sunt pretenţioşi,
o soluţie de zahăr sau de miere în concentraţie de 10-15% fiind suficientă pentru menţinerea viabilităţii şi
fertilităţii în parametri normali. Păstrarea activităţii biologice a baculovirusurilor şi asigurarea eficacităţii
ecologice şi economice a preparatelor constituite în baza lor necesită aplicarea cunoştinţelor profunde privind
crearea condiţiilor tehnologice optimale pentru utilizarea acestora în combaterea insectelor dăunătoare, acţiu-
nea sinergetică dintre agenţii preparatului baculoviral şi suşele naturale de virusuri, precum şi aplicarea for-
melor preparative eficiente ale preparatelor elaborate.
Bibliografie:
1. BECKAGE, N.E., DREZEN, J.M. Polydnaviruses as Endocrine Regulators. In: Parasitoid Viruses. London, UK:
Academic Press., 2012, p.163-168.
2. BOGULEANU, G., NICA, F., PETRESCU, E., BERATLIEF, C. Influenţa factorului trofic asupra dezvoltării
omisei păroase a dudului (Hyphantria cunea Drury). În: Analele ICDPP, România, 2005, vol.11, p.167-175.
3. BERATLIEF, C., BOGULEANU, G., IONESCU, C., NICA, F., PETRESCU, E. Activitatea sezonieră a adulţilor
de Hyphantria cunea Drury (Lepidoptera: Arctiidae). În: Analele ICDPP, Romania, 2005, vol.12, p.217-227.
4. CIUHRII, M., ARMENESCU–CIUHRII, E. Virusul – spaimă şi speranţa omenirii. Bucureşti: Mirabilis, 2008,
p.248.
5. CHANDLER, D., GREAVES, J., PRINCE, G., TATCHELL, M., BAILEY, A. Biopesticides: Pest Management
and Regulation. CABI, 2010, p.256.
6. KEAN, J.M., KUMARASINGHE, L.B. Predicting the seasonal physiology of fall webworm (Hyphantria cunea) in
New Zealand. In: New Zealand Plant Protec, 2007, vol.60, p.279-285.
7. KOUL, O., CUPERUS, G.W. Ecologically-Based Integrated Pest Management. CABI, 2007, p.480.
8. HALBERG, N., ALROE, H.F., KNUDSEN, M.T., KRISTENSEN, E.S. Global Development of Organic Agriculture:
Challenges and Prospects. CABI, 2006, p.384.
9. VOLOŞCIUC, L. Biotehnologia producerii şi aplicării preparatelor baculovirale în protecţia plantelor. Chişinău:
Mediul ambiant, 2009, p.262.
10. ГАР, К.А. Методы испытания токсичности и эффективности инсектицидов. Москва, 1963, с.283.
11. ДОСПЕХОВ, Б.А. Методика полевого опыта. Москва: Колос, 1979, с.416.
Prezentat la 24.12.2012

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
105
ЭФФЕКТИВНОСТЬ ФЕРОМОНА ФРУКТОВОЙ ПОЛОСАТОЙ МОЛИ
(Anarsia lineatella Z.)
AIDAROSS NASSER NASR NAJI, Diana CRISTMAN*
Университет Адена, Йемен *Институт защиты растений и экологического земледелия АН Молдовы
EFICIENŢA FEROMONULUI MOLIEI VĂRGATE A FRUCTELOR (Anarsia lineatella Z.)
Molia vărgată a fructelor (Anarsia lineatella Z.) în livezile de mere din Moldova se dezvoltă în două-trei generaţii.
Procentul de lăstari şi fructe atacate de dăunător poate atinge 30-60%. Pentru monitorizarea populaţiei şi optimizarea
perioadei de tratamente poate fi utilizat feromonul sexual sintetic.
Cuvinte-cheie: feromon, capcană, masculi, monitoring, populaţie.
EFFICIENCY OF PHEROMONES FRUIT MOTH STIPED (Anarsia lineatella Z.) Fruit striped moth (Anarsia lineatella Z.) in apple orchards of Moldova developed in two-three generations. Percen-
tage of occupied shoots and fruits may reach 30-60%. To monitoring the population, the timing of treatments using
synthetic sex feromons.
Keywords: pheromone, trap, male, monitoring, population.
Введение
Фруктовая полосатая моль (Anarsia lineatella Z.,Gelehiidae) – маленькая узкокрылая ночная бабоч-
ка с размахом крыльев 11-14 мм. Передние крылья темно-серые, с прерывающимися продольными
черными черточками, задние – серые, с длинной серебристой бахромой. Гусеница красновато-бурая
или коричневая, с черными головкой, грудным щитком и грудными ножками. Длина взрослой гусе-
ницы – 8-12 мм (фото 1,2). Зимуют очень маленькие гусеницы (длиной около 1-1,5 мм) в коре тонких
веток, преимущественно в развилках. Весной они пробуждаются, внедряются сначала в почки, а за-
тем в молодые зеленые побеги, выгрызая их сердцевину (фото 3), в результате чего побеги увядают
и засыхают. Позднéе гусеницы переползают на другие веточки и, нанося им такое же повреждение,
вызывают их гибель. За время своего развития одна гусеница может повредить 4-5 побегов. Закончив
питание, гусеницы прогрызают отверстие в поврежденных побегах, выходят наружу и прячутся в
щелях коры, сухих листьях, под комочками земли или в почве на глубине 2-10 см и в других местах,
где плетут себе паутинные коконы, внутри которых окукливаются. В конце мая – июне из куколок
выходят бабочки, которые летают в сумерках. Как и листовертки, фруктовая полосатая моль летит
ночью на яркий свет. Поэтому сигналом для борьбы с этим вредителем до недавнего времени было
обнаружение бабочек в светоловушках. Бабочки откладывают яйца на молодые ветки и побеги у
основания почек, а также на листья и плоды. Отрождающиеся гусеницы вгрызаются в почки, побеги
и плоды и протачивают в них ходы. В июле они снова окукливаются, а в конце июля – августе начи-
нают летать бабочки второй генерации, которые и откладывают яйца на побеги. Отрождающиеся
гусеницы до осени питаются в коре побегов и остаются там зимовать. Бабочки имеют две генерации.
В конце мая-июне – лѐт первой генерации, лѐт бабочек второго поколения длится с июня по август.
В последние годы установлено, что неоплодотворенные самки выделяют феромон, привлекающий
самцов, который идентифицирован как 5,9-диметилгептадекан с оптимальной дозой 1мг/испаритель.
Вылет самцов у данного вида опережает вылет самок на 5-7 дней. Самцы способны реагировать
на феромон самки и спариваться уже в первый день после выхода из куколки [1,2], что является
хорошей предрасположенностью для мониторинга развития популяций вредителя и проведения
феромонных методов регулирования численности вредителя за счѐт «массового отлова самцов» на
феромоно-клеевые ловушки, их «стерилизации» в феромонных ловушках, обработанных стери-
лизующими веществами, или «дезориентации» за счѐт насыщения воздуха феромонами высокой
концентрации.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
106
Фруктовая полосатая моль имеет широкое распространение, встречается во всех южных и централь-
ных районах садоводства, где на больших площадях выращивают персик, сливу и абрикос: в южных
районах Европы, в Украине, Молдове, в Крыму, на Кавказе и в Средней Азии, а также в восточном
Средиземноморье и Северной Америке. Фруктовая полосатая моль повреждает все косточковые пло-
довые культуры, лох узколистный, яблоню и хурму. На поврежденных побегах и плодах косточковых
пород образуются наплывы, из которых вытекает камедь с экскрементами гусениц. Поврежденные
плоды преждевременно опадают и загнивают. Величина урожая и его качество резко снижаются.
В целях профилактики проводят вырезание и сжигание поврежденных гусеницами фруктовой
моли и восточной плодожорки увядающих побегов; сбор и вывоз из сада падалицы с последующим
ее уничтожением; рыхление почвы под кронами деревьев; накладывание ловчих поясов на стволы и
скелетные ветви для отлова гусениц вредителей, уходящих на окукливание в почву [3].
Из мер борьбы с фруктовой полосатой молью могут быть эффективными опрыскивания золоном,
карбофосом, хлорофосом и другими более современными инсектицидами, во время распускания
почек и в более поздние сроки. Первое опрыскивание обычно проводится весной, до распускания
почек, второе – сразу после цветения, в фазе «осыпания лепестков» персика и абрикоса или в фазе
«розового бутона» у яблони. Затем – по мере необходимости [3].
В связи с тем, что персиковые сады Центральной почвенно-климатической зоны Молдовы помимо
фруктовой полосатой моли (Anarsia lineatella Z.) заселяются и восточной плодожоркой (Grapholita
molesta Busck), продолжительность развития которой, в период вегетации растений персика, и характер
повреждений побегов и плодов практически трудно отличимы от повреждений гусеницами фрукто-
вой полосатой моли, мы на одном и том же опытном участке в 4 гектара, равномерно (из расчѐта 10
ловушек/га) разместили феромонные ловушки обоих видов для изучения возможности регулирования
плотности популяций вредителей за счѐт массового отлова самцов.
Материалы и методы исследования
Испытания синтетического полового феромона фруктовой полосатой моли Anarsia lineatella Z -
5,9-диметилгептадекан, синтезированного в Институте защиты растений АНМ, проводились нами в
персиковом саду АОО «Агробрио», с. Бачой, Яловенского района (Центральная почвенно-климати-
ческая зона Молдовы). Феромонно–клеевые ловушки типа «Дельта», оснащѐнные феромоном фрук-
товой полосатой моли в дозе 1 мг на каучуковом испарителе, вывешивали на высоте 1,5-2 м через
каждые 50 метров. В саду площадью до 10 га вывешивали одну ловушку на 1-2 га, а площадью свыше
10 га – одну ловушку на 5 га. Осмотр ловушек и выборку отловленных бабочек проводили через 2-3
дня. Феромонные капсулы заменяли через 30 дней, а клеевые вкладыши – по мере загрязнения, но
не реже чем один раз в 12-14 дней. По окончании периода вегетации и после уборки урожая ловушки
удаляли из сада и уничтожали.
Для определения плотности популяции вредителя осмотр ловушек и учѐты отловленных бабочек
проводили через 3-5 дней. На основании отлова первых бабочек определяли сроки начала лѐта, а на
основании числа отловленных бабочек за 2-3 дня – начало массового лѐта и его завершение. Изуче-
ние динамики отлова позволяет определять продолжительность лѐта по поколениям, продолжитель-
ность массового лѐта и число поколений за вегетационный период развития растений.
В целях определения биологической эффективности метода массового отлова самцов фруктовой
полосатой моли и восточной плодожорки, феромонные ловушки вывешивали после отлова первых
бабочек на сигнальные ловушки. В случае присутствия на участке вредителей обоих видов, феромон-
ные ловушки каждого вида размещали на соседних рядах (30 х 30 м между ловушками), на высоте 2-
2,5 м. Осмотр, выборку и учѐт отловленных бабочек проводили один раз в неделю. Замену клеевых
вкладышей проводили по мере загрязнения, но не реже чем один раз в 14 дней. Феромонные испари-
тели (диспенсеры) заменяли через каждые 30 дней. Комплекты феромонных материалов, после завер-
шения вегетационного периода, из сада удаляли.
Результаты исследований
Использование нами феромонных ловушек для изучения сезонной динамики лѐта бабочек пока-
зало, что в условиях Центральной зоны Молдовы лѐт бабочек фруктовой полосатой моли в 2012 году
длился непрерывно 161 день, с 27 апреля по 18 сентября, с тремя пиками (во второй половине июня, в
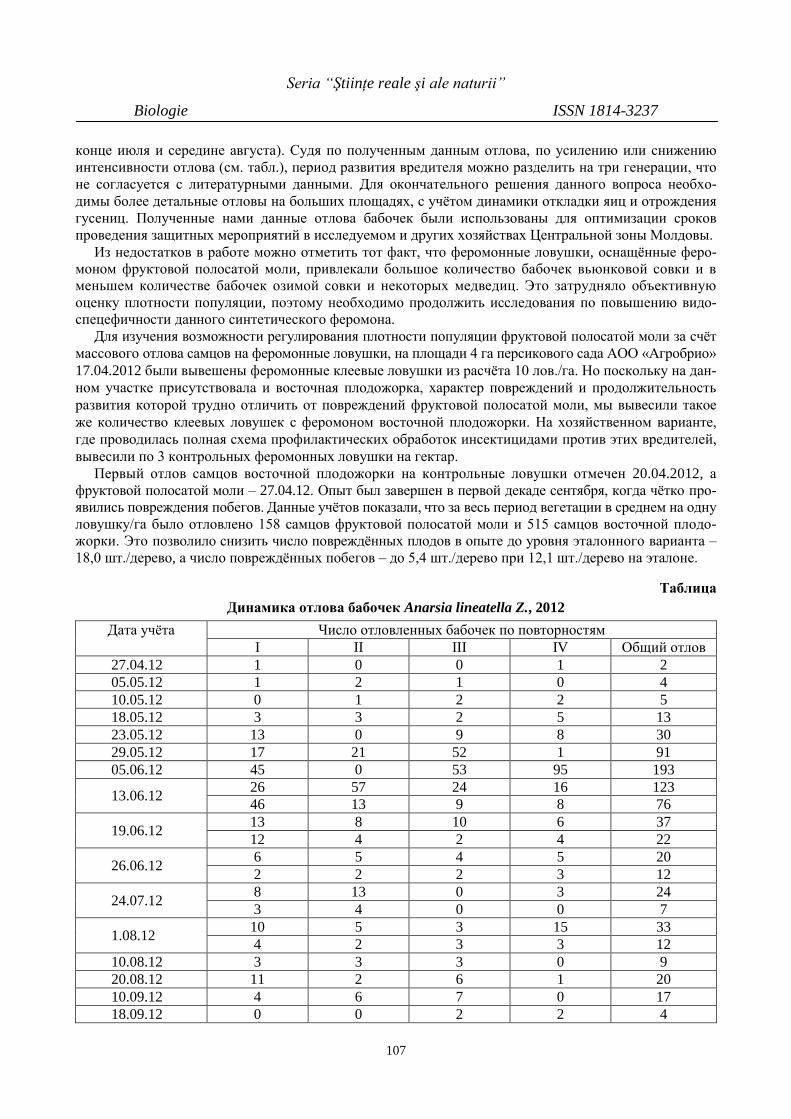
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
107
конце июля и середине августа). Судя по полученным данным отлова, по усилению или снижению
интенсивности отлова (см. табл.), период развития вредителя можно разделить на три генерации, что
не согласуется с литературными данными. Для окончательного решения данного вопроса необхо-
димы более детальные отловы на больших площадях, с учѐтом динамики откладки яиц и отрождения
гусениц. Полученные нами данные отлова бабочек были использованы для оптимизации сроков
проведения защитных мероприятий в исследуемом и других хозяйствах Центральной зоны Молдовы.
Из недостатков в работе можно отметить тот факт, что феромонные ловушки, оснащѐнные феро-
моном фруктовой полосатой моли, привлекали большое количество бабочек вьюнковой совки и в
меньшем количестве бабочек озимой совки и некоторых медведиц. Это затрудняло объективную
оценку плотности популяции, поэтому необходимо продолжить исследования по повышению видо-
спецефичности данного синтетического феромона.
Для изучения возможности регулирования плотности популяции фруктовой полосатой моли за счѐт
массового отлова самцов на феромонные ловушки, на площади 4 га персикового сада АОО «Агробрио»
17.04.2012 были вывешены феромонные клеевые ловушки из расчѐта 10 лов./га. Но поскольку на дан-
ном участке присутствовала и восточная плодожорка, характер повреждений и продолжительность
развития которой трудно отличить от повреждений фруктовой полосатой моли, мы вывесили такое
же количество клеевых ловушек с феромоном восточной плодожорки. На хозяйственном варианте,
где проводилась полная схема профилактических обработок инсектицидами против этих вредителей,
вывесили по 3 контрольных феромонных ловушки на гектар.
Первый отлов самцов восточной плодожорки на контрольные ловушки отмечен 20.04.2012, а
фруктовой полосатой моли – 27.04.12. Опыт был завершен в первой декаде сентября, когда чѐтко про-
явились повреждения побегов. Данные учѐтов показали, что за весь период вегетации в среднем на одну
ловушку/га было отловлено 158 самцов фруктовой полосатой моли и 515 самцов восточной плодо-
жорки. Это позволило снизить число повреждѐнных плодов в опыте до уровня эталонного варианта –
18,0 шт./дерево, а число повреждѐнных побегов – до 5,4 шт./дерево при 12,1 шт./дерево на эталоне.
Таблица
Динамика отлова бабочек Anarsia lineatella Z., 2012
Дата учѐта Число отловленных бабочек по повторностям
I II III IV Общий отлов
27.04.12 1 0 0 1 2
05.05.12 1 2 1 0 4
10.05.12 0 1 2 2 5
18.05.12 3 3 2 5 13
23.05.12 13 0 9 8 30
29.05.12 17 21 52 1 91
05.06.12 45 0 53 95 193
13.06.12 26 57 24 16 123
46 13 9 8 76
19.06.12 13 8 10 6 37
12 4 2 4 22
26.06.12 6 5 4 5 20
2 2 2 3 12
24.07.12 8 13 0 3 24
3 4 0 0 7
1.08.12 10 5 3 15 33
4 2 3 3 12
10.08.12 3 3 3 0 9
20.08.12 11 2 6 1 20
10.09.12 4 6 7 0 17
18.09.12 0 0 2 2 4

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
108
1 2 3
Фото. Фруктовая полосатая моль: 1 – бабочка; 2 – гусеницы; 3 – повреждѐнный побег персика. Источник: danaida.ru
Выводы:
1. Синтетический половой феромон Anarsia lineatella Z., Gelehiidae – 5,9-диметилгептадекан при
оптимальной дозе 1мг/испаритель в сочетании с клеевой ловушкой позволяет точно определять про-
должительность лѐта бабочек вредителя за весь период вегетации растений персика и получать объек-
тивные данные о плотности популяции по поколениям в обследуемых насаждениях.
2. По итогам испытаний 2012 года предложена методика применения феромона фруктовой поло-
сатой моли Anarsia lineatella Z, позволяющая точно определять сезонную динамику развития и плот-
ность популяций вредителя в насаждениях. Рекомендуется оптимальное размещение феромонных
ловушек в кроне деревьев, на высоте 1,5-2 м, через каждые 50 метров. В садах площадью до 10 га
следует вывешивать одну ловушку на 1-2 га, а площадью свыше 10 га – одну ловушку на 5 га.
3. Необходимо продолжить исследования по повышению видоспецефичности феромона фрукто-
вой полосатой моли в целях повышения его биологической эффективности и избегания привлечения
бабочек других видов.
4. Исходя из предварительных данных 2012 года, плотность популяции фруктовой полосатой
моли в персиковом саду можно регулировать посредством массового отлова самцов на феромонно-
клеевые ловушки. В случае присутствия в саду и восточной плодожорки, необходимо вывешивать и
соответствующее количество феромонно-клеевых ловушек и для массового отлова самцов восточной
плодожорки. Для более точного определения периода развития исследуемого вредителя, данный
эксперимент следует повторять на бóльшей площади.
Библиография:
1. БИЧИНА, Т.И. Борьба с молью-малюткой и другими минирующими молями в садах. Кишинев: Изд-во ЦК
КП Молдавии, 1970. 46 с.
2. ВОЙНЯК, В., ХАЙДАРЛЫ, Ю., МУСЛЕХ, М., СЫРБУ, В. Эффективность массового отлова самцов в
регулировании численности вредителей яблони. Материалы Международной научно-практической кон-
ференции «Биологическая защита растений – основа стабильности агроэкосистем», 23-25 сентября 2008.
Краснодар, 2008, вып. 5, с.328-330.
3. ЧЕРКЕЗОВА, С.Р. Экологизация систем защиты яблони от доминирующих вредителей / Автореф. дисс. …
канд. биол. наук. Краснодар, 2011. 24 с.
Prezentat la 10.10.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
109
МИНИРУЮЩИЕ МОЛИ В ЯБЛОНЕВЫХ САДАХ МОЛДОВЫ
AIDAROSS NASSER NASR NAJI
Университет Адена, Йемен
MOLILE MINIERE ÎN LIVEZILE DE MERE DIN MOLDOVA
Molile miniere (Cemiostoma scitella Zell.), (Lithocolletis corylifoliella Hw.), (Lithocolletis pyrifoliella Grsm.) în
livezile de mere din Moldova se dezvoltă în trei generaţii. Procentul foliilor populate de ele poate atinge 80%. Pentru
monitorizarea populaţiilor şi optimizarea perioadei de tratamente au fost utilizaţi feromonii sexuali sintetici.
Cuvinte-cheie: moli miniere, generaţii, feromoni sexuali, capcane, monitoring, tratamente.
MINED MOTH IN APPLE ORCHARDS OF MOLDOVA
Mined moth (Cemiostoma scitella Zell.), (Lithocolletis corylifoliella Hw.), (Lithocolletis pyrifoliella Grsm.) in apple
orchards of Moldova developed in three generations. Percentage of occupied leaves can reach 80%. For population
monitoring and optimizing the timing of treatments using synthetic sex pheromones.
Keywords: mined moth, generations, sex pheromones, traps, monitoring, treatments.
Введение
В яблоневых садах Молдовы, в связи со значительным уменьшением объема использования хими-
ческих инсектицидов или нерегулярным их применением в последние годы, широкое распространение
получили минирующие моли, которые по данным РСТЗР практически встречаются во всех насажде-
ниях республики c процентом заселѐнных листьев от 3,1% до 10,9% [10]. Наиболее вредоносными из
них являются кружковая боярышниковая (Cemiostoma scitella Zell.), верхнесторонняя (Lithocolletis
corylifoliella Hw.) и нижнесторонняя (Lithocolletis pyrifoliella Grsm.) моли, гусеницы которых, внедря-
ясь в паренхиму листовой пластинки, выгрызают ходы (мины), не повреждая эпидермис [1,2,5,6,9].
При плотности, в среднем, 8-10 мин на лист происходит преждевременный листопад, отставание
побегов в росте, уменьшение массы плодов и ухудшение их качества. Вредоносность минирующих
молей часто носит очаговый характер и зависит от количества мин на листе, сильнее проявляясь в
верхнем ярусе крон. При массовом размножении минѐры способны вызвать осыпание свыше 80%
листьев, которое часто сопровождается потерей урожая и осенним цветением деревьев. Дефолиация
более 30% листьев уже приводит к ослаблению растений, ухудшению качества плодов и их опадению.
А более сильная и повторяющаяся несколько лет подряд дефолиация может вызвать частичное или
полное усыхание деревьев [2,5,6,7,9].
При организации и проведении защитных мероприятий против минирующих молей необходимо
иметь в виду, что все три вида вредителей могут встречаться в саду одновременно, в разных соотно-
шениях, либо с преобладанием одного из них [2,3,5,6,7,8,10]. В связи с тем, что фенологические сроки
и биология развития указанных видов минѐров имеет много общих черт и сроков, меры борьбы с
каждым видом и сроки их проведения могут совпадать и под обработку могут попадать все три вида
одновременно. Кроме того, проводимые ранневесенние обработки против поливольтинных видов
листовѐрток могут совпадать со сроками проведения обработок против молей минѐров. В дальней-
шем, после опадения завязи, «минѐры» могут попадать под обработки, проводимые против яблоневой
плодожорки. В таком случае специальных обработок можно избежать.
Мерами борьбы с минирующими молями на плодовых культурах являются следующие. Агротех-
нические: зяблевая вспашка и перекопка приствольных полос для уничтожения зимующих куколок,
сбор и уничтожение осенью или ранней весной опавших листьев, очистка коры. Химические: обра-
ботки плодовых культур инсектицидами в начальный период массового образования мин (диаметр не
более 0,5 см) сразу после цветения яблони и повторно через 10-12 дней. Деревья необходимо опры-
скивать до цветения инсектицидами: цианокс, золон, карбофос, метатион, метафос, нексион, фосфа-
мид, хлорофос, децис, суми-альфа инсегар, калипсо, актара, димилин, матч, римон, номолт и др.
Отмечается, что большинство препаратов эффективны лишь при борьбе с молодыми гусеницами.
В целом, система защиты яблони от минирующих молей основана на биологических особенностях

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
110
доминирующего вида, имеющего в данный период основное хозяйственное значение. Для каждого
из видов рекомендованы от 1 до 3 обработок в различные сроки, что ведет к увеличению кратности
обработок.
Во избежание развития устойчивости вредителей необходимо соблюдать правила чередования
препаратов из различных групп соединений. Кроме того, многообразие действующих схем усложняет
проведение защитных мероприятий, направленных на снижение вредоносности группы минирующих
молей. Однако многочисленные публикации свидетельствуют о невысокой эффективности защитных
мероприятий против этих вредителей в связи с недостаточной изученностью вопросов биологии, эко-
логии и вредоносности минеров, сложной идентификацией преимагинальных стадий при визуальных
учетах, скрытостью местообитания гусениц и достаточно высокой их миграционной активностью
[5,6,7]. В целях организации эффективного применения средств защиты в борьбе с минирующими
молями на яблоне и других плодовых культурах, приводим некоторые сведения по биологии и эко-
логии основных наиболее опасных видов минирующих молей, обитающих в садах Молдовы и других
регионах с близкими климатическими условиями.
Результаты исследований и их обсуждение
Кружковая моль-минер, боярышниковая кружковая моль (Cemiostoma scitella Zell.), встречается
повсеместно: в Европе (кроме севера), Малой Азии, Иране, Сев. Африке, Китае, Ближнем Востоке.
Вредитель розоцветных культур (повреждает яблоню, грушу, айву, вишню, сливу, персик, рябину, боярышник). Бабочка с размахом крыльев 6-7 мм. Яйцо размером 0,28 мм, светло-серое, овальное,
сверху вогнутое. Гусеница до 5 мм длиной, серо-зеленая, с коричневой головкой. Куколка светло-коричневая в плотном белом ромбовидном коконе. Зимуют куколки в коконах среди опавшей листвы,
под отставшей корой, в трещинах коры. Вылет первых бабочек в различных регионах отмечается в начале-середине апреля [1,2,5,6,7,10], в Молдове – в третьей декаде апреля [1,2,3,7] и совпадает с
фазой развития яблони «выдвижение соцветий», а начало массового лѐта – с фазой «розового бутона». Массовый лѐт бабочек длится до завершения цветения. Продолжительность лѐта бабочек по поколе-
ниям может достигать 34-37 суток. Вылетевшие самки весной откладывают яйца на нижнюю сторону листьев, далеко от их краев, и очень редко на верхнюю. Средняя плодовитость самки – 55-60 яиц,
максимальная 80. Эмбриональное развитие длится 9-10 суток при температуре 23-24°С, при более низких температурах эмбриональное развитие более продолжительное. Отродившиеся гусеницы не
выходят на поверхность листа, а проникают в его ткани. Внедряясь в паренхиму листовой пластинки,
они выгрызают ходы (мины), не повреждая эпидермис. Мина округлая, со средним диаметром около 1 см, с концентрическими кругами темно-коричневых экскрементов, находящихся с верхней стороны
листа. Развитие гусеницы перезимовавшего поколения длится на протяжении 20-30 суток, а летних поколений – 15-18 суток. Окукливание весенних и летних поколений часто происходит на листьях,
гусеницы последнего поколения переходят на окукливание под отставшую кору, в трещины коры, под опавшие листья. В период зимовки, от различных причин, могут погибнуть от 33 до 53% куколок.
Кружковая моль относится к поливольтинным видам и за вегетационный период, в зависимости от региона, развивается в 1-4-х поколениях (в Якутии – 1, на юге Казахстане – 2-3, на юге Украины – 3-4,
в Молдове – 3 поколения). Первое поколение вредителя развивается со второй половины апреля до конца июня, второе – с июня до конца июля, третье – с июля до конца августа, четвертое – с августа
до ухода на зимовку. В зонах с 3-4-мя поколениями в отдельные годы последние поколения разви-ваются лишь частично. Вылет первых бабочек в южных районах Украины и на Кавказе отмечается в
начале-середине апреля [2,5,6], в Молдове – в третей декаде апреля [2,3,7] и, как правило, совпадает с периодом «выдвижения соцветий». Начало массового лѐта бабочек совпадает с фазой «розового
бутона». Массовый лѐт бабочек длится до завершения цветения, после чего на листьях появляются первые мины. Продолжительность лѐта бабочек по поколениям зависит от погодных условий и может
длиться от 24 до 37 суток. На одном листе может быть отложено до 7-18 и более яиц. Плодовитость
бабочки 40-80 яиц. Эмбриональное развитие длится 9-10 дней при температуре 23-24°С. Гусеницы не выходят на поверхность листа, а внедряются в его ткани, прогрызая хорион у основания яйца. Разви-
тие гусениц длится от 15 до 30 дней и проходит в мине. Окукливаются гусеницы первых поколений с нижней стороны листьев, зимующего поколения – под корой или под листьями. Развитие куколки
продолжается 9-16 дней. Самки выходят из куколок готовыми к спариванию, с уже созревшими яйцами

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
111
и не нуждаются в дополнительном питании. Неоплодотворенные самки выделяют феромон, привле-кающий самцов, идентифицированный как 5,9-диметилгептадекан [3]. Выход самцов из куколок
опережает выход самок на 5-7 дней, что является весьма важным фактором для успешного использо-
вания феромонных ловушек в целях мониторинга их развития, распространения и оптимизации сроков проведения защитных мероприятий, а также для регулирования плотности популяций путѐм массового
отлова самцов на феромонные ловушки или их дезориентации. Самцы способны реагировать на фе-ромон и спариваться уже в первый день после выхода из куколки [3,8]. Использование феромонных
ловушек для изучения сезонной динамики лѐта показало, что в условиях 2007 года лѐт бабочек имел место 161 день, с 27 апреля по 18 сентября, с тремя пиками (во второй половине июня, в конце июля
и середине августа). Первая генерация длилась 40 дней, вторая – 38 и третья – 53 дня [3]. Для Красно-дарского края эти сроки были следующими: начало лѐта бабочек первого поколения – 24.04.10, мас-
совое отрождение гусениц – 06.05.10; начало лѐта бабочек второго поколения – 12.06.10, отрождение гусениц – 22.06.10; лѐт бабочек 3-го поколения – 21.07.10, отрождение гусениц – 27.07.10; лѐт бабочек
4-го поколения – 24.08.10, отрождение гусениц – 02.09.10 [6]. Многолетние наблюдения за развитием, размножением и распространением кружковой моли и за еѐ вредоносностью в различных регионах
России [1,5,6] позволили разработать порог экономической вредоносности (ЭПВ), который состав-ляет для видов весенней группы – 50 гусениц на 100 учетных розеток, в фенофазу яблони «розовый
бутон», для видов летней группы – 3 гусеницы на 1 лист, или 70% заселенных розеток в период «осыпания избыточной завязи», что соответствует 50 гусеницам на 100 листьев, в период роста и
развития плодов (III декада июня – I декада июля). Период наибольшей вредоносности видов весен-ней группы наблюдается в сроки формирования урожая – с фенофазы «розовый бутон» до опадения
избыточной завязи. Второй – в период роста и развития плодов при максимальной плотности гусениц
комплекса видов, в конце июня–июле. Второе поколение вредителя обычно дает больше особей, чем первое. При плотности до 10 мин в среднем на лист происходит преждевременный листопад, что
может существенно снижать урожай. При большом количестве мин происходит отставание побегов в росте и уменьшение массы плодов. Вредоносность часто носит очаговый характер, зависит от коли-
чества мин на листе и сильнее выражена в верхнем ярусе крон.
Верхнесторонняя плодовая минирующая моль (Lithocolletis corylifoliella Hw.), как и кружковая
моль, имеет широкий ареал распространения: встречается в странах Малой Азии и Западной Европы,
на всей европейской части России, в Закавказье, Средней Азии, в Молдове, в Украине (более много-
численна в Крыму). Повреждает яблоню, грушу, айву, черешню, вишню, боярышник, мушмулу,
рябину, сливу, иргу, кизильник. Бабочка в размахе крыльев – 8-10 мм. Яйцо размером 0,3 мм, полу-
прозрачное, с зеленоватым оттенком, округлое. Гусеница до III возраста безногая с прогнатической
головой, IV–V возрастов – цилиндрическая, с развитыми ножками, длиной 5-6 мм. Куколка 3-4 мм,
первоначально кремовато-желтая, но к концу развития становится темно-коричневой. Зимуют гусе-
ницы последнего возраста в минах на опавших листьях. Вылет бабочек весной начинается перед цве-
тением яблони (в Крыму – во второй-третьей декадах апреля). Неоплодотворѐнные самки выделяют
феромон, привлекающий самцов, E-4, Z-7-ридекадиенилацетат. Бабочки живут от 3 до 12 дней и за
это время самки поодиночке откладывают на верхнюю сторону листа вблизи жилок или на жилки от
14 до 63 яиц. Период эмбрионального развития длится 7-10 дней. Отродившаяся гусеница не выходит
на поверхность листа, а внедряется в его ткань, под оболочку яйца. Гусеница до III-го возраста пита-
ется, как сокоед, разрушая и поглощая содержимое палисадной ткани, IV–V возрастов – углубляет
мину, выгрызая как палисадную, так и губчатую паренхиму листа. Мина верхнесторонняя, складча-
тая, расположена всегда вдоль центральной или боковых жилок листа. Цвет мины серовато-желтый,
иногда серебристый. Продолжительность развития гусеницы в летний период – от 24 до 42 дней.
Гусеница окукливается в мине. Продолжительность развития куколки весной – от 16 до 36 дней
(март–апрель), летних поколений – от 5 до 17 дней. В Крыму развивается три поколения в году.
Развитие одного поколения завершается за 36-65 дней. По данным отлова на феромонные ловушки и
визуальным учѐтам [3,4,5,6,7] в Молдове и в Краснодарском крае отмечены следующие сроки разви-
тия верхнесторонней моли : в Молове – лѐт бабочек начался 26.04.07 и длился до 13.09.07 с тремя
пиками, т.е. было три наслаивающихся одно на другое поколения (1-ое длилось 31 день, 2-ое – 67 и
3-ее – 48 дней). В Краснодаре: лѐт 1-ое – 18.04.10, отрождение гусениц – 05.05.10; лѐт 2-ое – 16.06.10,

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
112
гусеницы – 22.06.10; лѐт 3-е – 21.07.10, гусеницы – 30.07.10, окукливание – 15.08.10. Полученные
данные свидетельствуют, что для повышения эффективности обработок и сокращения их числа за
сезон необходимо постоянное наблюдение за продолжительностью развития вредителя, что в большой
степени зависит от погодных особенностей года.
Нижнесторонняя плодовая минирующая моль (Lithocolletis pyrifoliell F.) – многоядный вреди-
тель розоцветных плодовых пород деревьев и кустарников. Ареал распространения тот же, что и у
двух предыдущих молей минѐров. Гусеницы минируют листья культурных и дикорастущих семечко-
вых розоцветных растений, особенно яблони и айвы, реже груши, сливы, вишни, боярышника, муш-
мулы, ирги, рябины. Вредоносность вида определяется комплексом условий: плотностью популяции,
сортом кормового растения, районом и условиями произрастания культур. На ранних сортах яблони
плотность 2-е мины на лист уже стимулирует преждевременное созревание и опадение яблок, а при
плотности 4 мины/лист уменьшается размер плодов. Бабочка с размахом крыльев 7-9 мм. Яйцо раз-
мером 0,3 мм, полупрозрачное, с зеленоватым оттенком, округлое. Гусеница имеет две формы. Тело
сокоядной гусеницы дорсовентрально сплющенное, председатель клиновидный, прогнатичный, нож-
ки отсутствуют. Гусеница четвертого и пятого поколений цилиндрическая, имеет 8 пар ножек и у нее
развит прядильный аппарат. Длина гусеницы первого возраста 0,4-0,6 мм, пятого возраста – 5,5-6 мм.
Куколка длиной 4-4,5 мм, темно-коричневая, с острым клювовидный выростом. Зимуют куколки в
минах опавших листьев. Вылет бабочек начинается при среднесуточной температуре 11°С, что совпа-
дает с фенофазой «выдвижения соцветий» у зимних сортов яблони. Массовый лет отмечается в фено-
фазе «розовый бутон» и продолжается во время цветения яблони. Кладку яиц самка начинает через
2-3 часа после спаривания. Яйца откладывает по одному на нижнюю сторону листа. Плодовитость –
90 яиц. Эмбриональное развитие продолжается 6-10 суток. Отродившаяся гусеница через оболочку
яйца и эпидермис проникает в паренхиму листа, где и питается 21-28 суток, из которых в форме со-
коядной гусеницы – 11-13 суток. Окукливаются гусеницы в минах. Стадия куколки длится 8-13 суток.
В лесостепи Украины развивается два поколения, в степных районах – три. Зимуют куколки послед-
него поколения. До третьего возраста гусеницы питаются соком, высасывая его из клеток миниро-
ванного листа. Гусеница первого возраста прогрызает в листке тонкую мину длиной до 1-1,2 см, во
втором возрасте расширяет и удлиняет ее до 2 см, в третьем – до 2,5-3 см. Гусеницы старших возрас-
тов выгрызают ткани, оставляя эпидермис. В Беларуси вредитель развивается в 2-х поколениях, на
юге Украины, в Молдове, Северном Кавказе и в Закавказье – в 3-х поколениях. Лѐт бабочек начинает-
ся в конце апреля и длится до октября, поколения накладываются друг на друга [2,3,4,5,6,7]. Бабочки
живут более 2 недель, активны в вечерние часы. Их лѐт начинается с 18 часов и прекращается с на-
ступлением темноты. Они перелетают с ветвей на листья, иногда могут скапливаться на стволах.
Оптимальная температура для откладки яиц, при которой скорость откладки максимальна, 26°С.
Неоплодотворѐнные самки выделяют феромон для привлечения самцов – Е-10-додеценилацетат.
Использование феромонных ловушек для фенологических наблюдений показало, что в Молдове в
условиях 2007 года лѐт бабочек начался 26.04.07 и длился непрерывно до 23.09.07 с тремя пиками
(лѐт 1-го поколения длился 37 дней, 2-го – 56 и 3-го – 44 дня). Первые мины появлялись на 6-9 день
массового лѐта [3]. Оплодотворенные самки начинают кладку яиц в следующую за копуляцией ночь.
Для развития яиц необходима сумма эффективных температур 172°C при нижнем пороге развития
9°С. Соотношение полов зависит от температуры и численности популяции.
Выводы:
1. Среди листовых вредителей яблони наиболее опасными являются кружковая боярышниковая
(Cemiostoma scitella Zell.), верхнесторонняя (Lithocolletis corylifoliella Hw.) и нижнесторонняя (Litho-
colletis pyrifoliella Grsm.) моли.
2. Синтетические половые феромоны можно успешно использовать в целях мониторинга развития
молей минѐров на яблоне.
Библиография:
1. АНТЮХОВА, О.В. Биоэкологические особенности минирующих молей и защита от них декоративных
растений в Приднестровье / Автореферат дисс. … канд. биол. наук. Санкт-Петербург, 2010. 32 с.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
113
2. БИЧИНА, Т.И. Борьба с молью-малюткой и другими минирующими молями в садах. Кишинев: Изд-во ЦК
КП Молдавии, 1970. 46 с.
3. ВОЙНЯК, В., ХАЙДАРЛЫ, Ю., МУСЛЕХ, М., СЫРБУ, В. Эффективность массового отлова самцов в
регулировании численности вредителей яблони: Материалы Международной научно-практической конфе-
ренции «Биологическая защита растений – основа стабильности агроэкосистем», 23-25 сентября 2008.
Краснодар, 2008, вып.5, с.328-330.
4. Временные методические указания по выявлению и учету численности вредных и полезных организмов,
болезней с/х культур и сорняков. Кишинев, 1988 с.54-58.
5. КРЮКОВА, А.В. Минирующие моли – вредители яблони на Северо-Западе России и биоэкологическое обо-
снование мер борьбы с ними / Автореферат дисс. … канд. биол. наук. Санкт-Петербург, 2004. 28 с.
6. ЧЕРКЕЗОВА, С.Р. Экологизация систем защиты яблони от доминирующих вредителей / Автореф. дисс. …
канд. биол. наук. Краснодар, 2011. 24 с.
7. BOUBĂTRÂN, I. Combaterea bolilor şi dăunătorilor în plantaţiile pomicole. Chişinău, 2002. 52 p.
8. OLOI, I. Datele raportului 2004. Chişinău (needitate).
9. PERJU, T. Entomologia agricolă – componenţă a protecţiei integrate a agroecosistemelor. Bucureşti: Ceres, 1995,
vol.2, p.231-234.
10. Pronosticul răspândirii principalilor dăunători şi boli a culturilor agricole pentru anul 2010. Chişinău, 2009. 74 p.
Prezentat la 08.10.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
114
ПОЛУЧЕНИЕ ИНОКУЛЮМА ALTERNARIA КУЛЬТИВИРОВАНИЕМ ПАТОГЕНА
НА БУМАЖНЫХ ФИЛЬТРАХ
Виктория ШУБИНА
Институт защиты растений и экологического земледелия АН Молдовы
OBŢINEREA INOCULULUI DE ALTERNARIA PRIN CULTIVAREA PATOGENULUI
PE FILTRE DE HÂRTIE Pentru obţinerea inoculului, fitopatogenul a fost cultivat pe filtre de hârtie înmuiate în mediu de nutriţie lichid. Mediul
de nutriţie a fost pregătit pe bază de decoct de tomate, ardei, varză sau cartof-morcov, la care s-a adăugat 0,2% zaharoză. Incubaţia a durat 7 zile, la temperatura de 24,5°C, o fotoperiodă de 12 ore.
Cuvinte-cheie: Alternaria, sporulaţie, concentrat de inocul.
PRODUCTION OF INOCULUM OF ALTERNARIA WAS ACHIEVED BY CULTURING THE
PATHOGEN ON PAPER FILTERS When obtaining conidia the moistened paper filters, wetted with liquid nutritive media, prepared on the base of one
of the following cultures: tomato, paper, cabbage or potato and carrot liquor have been used. The liquid contained 0,2% sucrose. Time and temperature of incubation were 7 days, 24°C under a 12-hours photoperiod.
Keywords: Alternaria, sporulation, inoculum concentration.
Альтернариозы, вызываемые грибами рода Alternaria, встречаются повсеместно. Растениями-хозяевами гриба Alternaria являются разнообразные сельскохозяйственные культуры: томаты, карто-фель, баклажаны, перец, морковь, разные виды капусты, табак, цитрусы, множество декоративных растений и сорняки [7]. Поражает Alternаria вегетативную часть растений и плоды (в период вегетации и хранения).
Фитопатологи и селекционеры в своей работе используют искусственный инфекционный фон для тестирования химических и биологических препаратов, определения вирулентности гриба, отбора устойчивых сортов сельскохозяйственных культур.
В естественной среде не всегда имеется достаточное количество инфицированного материала, и оценка может быть недостоверной вследствие неравномерного распределения инфекции на обследуе-мой площади, поэтому на практике чаще всего создаются искусственные инфекционные фоны.
При создании инфекционного фона для болезней, вызываемых грибными патогенами, поражаю-щими вегетативную часть растений, обычно рекомендуется инокуляция растений суспензией спор гриба. Отмечено, что грибы рода Alternaria относительно хорошо растут на искусственных питатель-ных средах, однако не всегда удается добиться хорошего спороношения. На отсутствие споруляции или слабую споруляцию при культивировании изолятов Alternaria in vitro указывают многие авторы [1, 5, 6, 10], в связи с чем много работ посвящено стимуляции конидиеобразования гриба Alternaria.
Для получении инокулюма предлагаются различные методы стимулирования споруляции гриба: воздействие светом различных ламп (ультрафиолетовое облучение, флуоресцентное освещение, лампы белого накаливания), изменение фотопериода, разрушение и удаление воздушного мицелия микро-биологическими инструментами, выращивание на фильтровальной бумаге, выращивание на средах специального состава и различные комбинации этих методов. При использовании этих методик спо-руляция сильно варьирует, что препятствует получению стабильного количества инокулюма.
Важнейшими факторами споруляции гриба являются условия питания, температура, влажность и световые условия.
Японскими учеными Ohmori Kaori & Nakajima Mitsuo [9] отмечалось, что эффективность споруля-ции Alternaria kikuchiana Tanaka зависела не только от среды и температуры, но и от удаленности флуоресцентных ламп от мицелия. При высокой температуре и увеличении удаленности мицелия от ламп споруляция гриба уменьшалась, но увеличивался рост воздушного мицелия и, наоборот, при низкой температуре и близком расположении к лампам споруляция гриба увеличивалась. Спорообра-зование гриба было обильным и при стимулировании мицелия ультрафиолетовыми лампами при низ-кой температуре и небольшой удаленности от ламп.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
115
Masangkay R.F. et al. [8] отмечали влияние состава среды и освещенности на споруляцию. Спору-
ляция гриба при близком расположении к ультрафиолетовым лампам при температуре 28°С заметно
увеличивалась на среде V-8, но существенно уменьшалась на среде ½ КГА (картофельно глюкозный
агар). В темноте происходило обратное: на среде ½ КГА было получено самое большое количество
конидий, на среде V-8 число конидий существенно снизилось.
В работе Carvalho D.D.C. et al. приводится сравнительная характеристика методологических под-
ходов к стимулированию конидиеобразования изолятов альтернарии, выделенных с цитрусов. При
сравнении действия ламп с различными характеристиками наряду с мицелиальным стрессом, связан-
ным с механическим повреждением и холодовым шоком при 12 и 24 часах освещенности, проводи-
лось инокулирование стерильной ткани цитрусовых. Первоначально гриб выращивали на среде КГА
в течение 5 дней при температуре 25°С с 12-часовым фотопериодом, затем блоками гриба инфициро-
вали кусочки ткани и через 4 дня наблюдали рост мицелия. Эта методика удобна для проверки фито-
токсичности изолятов гриба [5].
Методика, разработанная Shahin E.A. & J.F. Shepard [11], является одной из успешных попыток
получения обильного конидиеобразования. Эта методика двухфазная. Сначала гриб выращивали на
первичной среде, каковой являлся КГА (лучшей для мицелиального роста гриба) [2, 5, 6, 8, 11, 12].
Затем использовали S-среду с добавлением карбоната кальция для получения обильного спороноше-
ния. Работая ранее по этой методике, мы внесли в нее некоторые коррективы, подходившие для наших
условий [3]. Это было обусловлено тем, что на среде КГА при температуре 25°С в темноте [10] нам
не удавалось получить колонии без воздушного мицелия. В наших опытах колонии росли медленно и
образовывали пышный воздушный мицелий без конидий. При этом ни в темноте, ни при естественном
фотопериоде (при освещении как верхней, так и нижней стороны колоний) конидии не образовывались
ни при 25°С, ни при 19°С, вплоть до высыхания среды в чашках Петри. Колонию гриба разрезали
стерильным скальпелем на прямоугольные блоки, которые переносили на чашки Петри со средой,
содержащей СаСО3 [3,11]. Спороношение получали уже после 2-х суток культивирования.
В практике получения конидий на искусственных питательных средах не всегда, используя тот
или иной метод, удается получить стабильное количество необходимого инокулюмного материала.
Одним из методов получения стабильного количества инокулюма может являться использование
бумажных фильтров [4, 6, 10].
Методика использования бумажных фильтров также является двухфазной. Первичная стадия на-
работки мицелия может быть различна. В случае Dhingra O.D. & Sinclair J. [6], наработка первичного
инокулюма идет во встряхиваемой культуре до тех пор, пока не сформируется достаточный мицелий.
Мицелий собирается путем центрифугирования, измельчается в блендере и повторно центрифугиру-
ется. Осадок ресуспендируют и распределяют на сухие бумажные фильтры в чашки Петри. Никакого
питания на бумажные фильтры не добавляют.
При изучении влияния состава питательных сред на рост гриба Alternaria (гриб выращивали в ста-
ционарной и глубинной культуре, при определении биомассы гриба использовали бумажные фильтры),
мы обратили внимание на массовое спорообразование гриба как на самом мицелии, так и на внутрен-
ней и наружной сторонах фильтра [2]. Поэтому наше внимание привлекла методика получения ино-
кулюма альтернарии культивированием патогена на бумажных фильтрах [4]. Методика была раз-
работана для Alternaria helianthi – серьезного патогена подсолнечника. Культуру гриба выращивали
на среде КГА в чашках Петри, а затем переносили растиранием агаризованного блока с мицелием
гриба на бумажные 3-слойные фильтры, смоченные экстрактом семян подсолнечника и 0,2% сахаро-
зы (для получения повторного спороношения). Чашки Петри с фильтрами стерилизовали при 0,5 атм
15 минут. После стерилизации фильтры инокулировали агаризованными блоками с мицелием гриба
Alternaria helianthi. Инокулированные фильтры инкубировали при температуре 23-25°С при 12-часо-
вом фотопериоде. Многочисленные конидии получали в течение 7 дней и после полного высыхания
фильтров хранили их в чашках Петри при комнатной температуре. Жизнеспособность конидий сохра-
нялась более 12 месяцев. Титр конидий не определяли. Спороношение оценивалось как обильное или
слабое, Allen et al. [4].
Цель нашего исследования – адаптировать методику Allen et al. [4], разработанную для Alternaria
helianthi, для альтернарии с пасленовых культур.
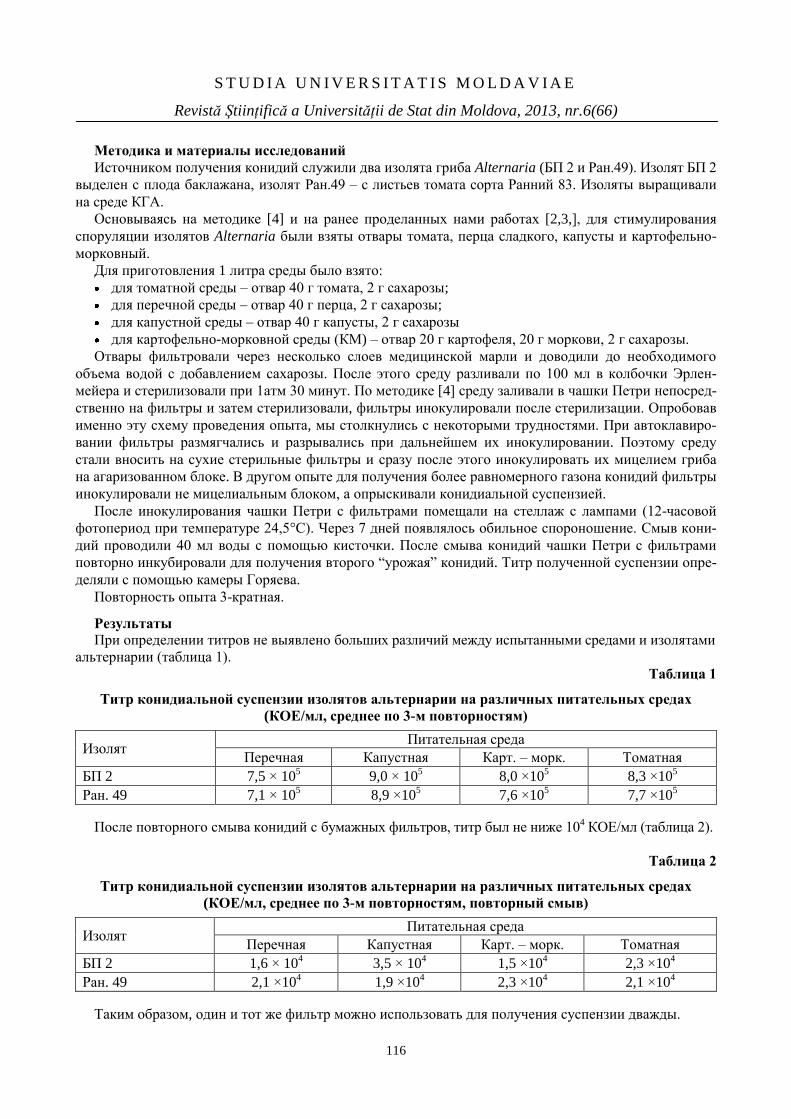
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
116
Методика и материалы исследований
Источником получения конидий служили два изолята гриба Alternaria (БП 2 и Ран.49). Изолят БП 2
выделен с плода баклажана, изолят Ран.49 – с листьев томата сорта Ранний 83. Изоляты выращивали
на среде КГА.
Основываясь на методике [4] и на ранее проделанных нами работах [2,3,], для стимулирования
споруляции изолятов Alternaria были взяты отвары томата, перца сладкого, капусты и картофельно-
морковный.
Для приготовления 1 литра среды было взято:
для томатной среды – отвар 40 г томата, 2 г сахарозы;
для перечной среды – отвар 40 г перца, 2 г сахарозы;
для капустной среды – отвар 40 г капусты, 2 г сахарозы
для картофельно-морковной среды (КМ) – отвар 20 г картофеля, 20 г моркови, 2 г сахарозы.
Отвары фильтровали через несколько слоев медицинской марли и доводили до необходимого
объема водой с добавлением сахарозы. После этого среду разливали по 100 мл в колбочки Эрлен-
мейера и стерилизовали при 1атм 30 минут. По методике [4] среду заливали в чашки Петри непосред-
ственно на фильтры и затем стерилизовали, фильтры инокулировали после стерилизации. Опробовав
именно эту схему проведения опыта, мы столкнулись с некоторыми трудностями. При автоклавиро-
вании фильтры размягчались и разрывались при дальнейшем их инокулировании. Поэтому среду
стали вносить на сухие стерильные фильтры и сразу после этого инокулировать их мицелием гриба
на агаризованном блоке. В другом опыте для получения более равномерного газона конидий фильтры
инокулировали не мицелиальным блоком, а опрыскивали конидиальной суспензией.
После инокулирования чашки Петри с фильтрами помещали на стеллаж с лампами (12-часовой
фотопериод при температуре 24,5°С). Через 7 дней появлялось обильное спороношение. Смыв кони-
дий проводили 40 мл воды с помощью кисточки. После смыва конидий чашки Петри с фильтрами
повторно инкубировали для получения второго “урожая” конидий. Титр полученной суспензии опре-
деляли с помощью камеры Горяева.
Повторность опыта 3-кратная.
Результаты
При определении титров не выявлено больших различий между испытанными средами и изолятами
альтернарии (таблица 1).
Таблица 1
Титр конидиальной суспензии изолятов альтернарии на различных питательных средах
(КОЕ/мл, среднее по 3-м повторностям)
Изолят Питательная среда
Перечная Капустная Карт. – морк. Томатная
БП 2 7,5 × 105
9,0 × 105
8,0 ×105
8,3 ×105
Ран. 49 7,1 × 105
8,9 ×105
7,6 ×105
7,7 ×105
После повторного смыва конидий с бумажных фильтров, титр был не ниже 104 КОЕ/мл (таблица 2).
Таблица 2
Титр конидиальной суспензии изолятов альтернарии на различных питательных средах
(КОЕ/мл, среднее по 3-м повторностям, повторный смыв)
Изолят Питательная среда
Перечная Капустная Карт. – морк. Томатная
БП 2 1,6 × 104 3,5 × 10
4 1,5 ×10
4 2,3 ×10
4
Ран. 49 2,1 ×104
1,9 ×104
2,3 ×104
2,1 ×104
Таким образом, один и тот же фильтр можно использовать для получения суспензии дважды.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
117
Титр конидий равномерного конидиального газона был на уровне 105 КОЕ/мл: на капустной
среде 5,1×105
спор/мл; томатной среде 3,4×105
спор/мл; КМ среде 2,7×105
спор/мл; перечной среде
2,3×105 спор/мл (использовали только изолят Ран.49, рис.1).
Рис.1. Чашка Петри с конидиями альтернарии
(изолят Ран.49, фильтр пропитан жидкой картофельно-морковной средой).
Фильтры с конидиями до использования могут храниться в лаборатории без потери жизнеспособ-
ности в течение довольно продолжительного времени. Нами установлено, что конидии жизнеспо-
собны в течение полугода. Наблюдения по сохранению жизнеспособности конидий продолжаются.
Выводы
1. Подобраны питательные среды для продуцирования конидий на бумажных фильтрах изолятов
Alternaria, выделенных с пасленовых культур.
2. При использовании одной и той же инфицированной чашки Петри, можно получить повторный
сбор конидий для получения суспензии.
Библиография:
1. ДЕМИДОВ, Е.С., САДЫКИНА, Е.И., САЙЧУК, А.И. Методы селекции томата на устойчивость к альтер-
нариозу. Тирасполь, 2006. 99 с.
2. НИКОЛАЕВА, С.И., НИКОЛАЕВ, А.Н., ШУБИНА, В.Э., ВОЛОЩУК, Л.Ф. Влияние состава питательной
среды на рост грибов рода Alternaria. В: Studia Universitatis. Seria Ştiinţe reale şi ale naturii, 2011, №1(41),
р.117-123.
3. ШУБИНА, В.Э., НИКОЛАЕВА, С.И., НИКОЛАЕВ, А.Н. Метод инициации конидиеобразования Alternaria
solani (Ell. & Mart.) Jones & Grout на искусственной питательной среде. В: Материалы докладов Между-
народного симпозиума «Защита растений – достижения и перспективы». Кишинев, 2009, с.209-210.
4. ALLEN, S.J., BROWN, J.F., KOCHMAN, J.K. Production of inoculum and field assessment of Alternaria helianthi
on sunflower. In: Plant disease, 1983, №39, p.665-668.
5. CARVALHO, D.D.C., ALVES, E., BATISTA, T.R.S., CAMARGOS, R.B., LOPES, E.A.G.L. Comparison of
methodologies for conidia production by Alternaria alternate from citrus. In: Brazillian Journal of Microbiology,
2008, №39, p.792-798.
6. DHINGRA, O.D., SINCLAR James. Basic plant pathology methods. 1995. 434 p.
7. FRANKLIN, L. Alternaria diseases. http://anrcatalog.ucdavis.edu
8. MASANGKAY, R.F., PAULITZ, T.C., HALLETT, S.G., WATSON, A.K. Biocontrol science and technology,
2000, vol.10, №4, p.385-397.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
118
9. OHMORI, K., NAKAJIMA, M. Effect of light on sporulation of Alternaria kikuchiana Tanaka. In: The Phytopatho-
logical Society of Japan, 1970, vol.36, №1, p.11-16.
10. RODRIGUES, T.T.M.S., MAFIA, L.A., DHINGRA, O.D., MIZUBUTI, E.S.G. In vitro production of conidia of
Alternaria solani. In: Tropical plant pathology, 2010, vol.35, №4, p.203-212.
11. SHAHIN, E.A., SHEPARD, J.F. An efficient technique for inducing profuse sporulation of Alternaria species. In:
Phytopathology, 1979, №69, p.618-620.
12. SINGH, U.P., SINGH, S.K., SARMA, B.K. Time-dependent sporulation, conidial size and germ tube formation in
Alternaria tenissiana (Kunza ex Pers.) Wiltshire on different media. In: J. Phytopathology, 2000, №148, p.413-416.
Prezentat la 25.09.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
119
ИЗОЗИМЫ ЭСТЕРАЗЫ ИНБРЕДНЫХ ЛИНИЙ И ГИБРИДОВ КУКУРУЗЫ
Андрей БАБИЦКИЙ
Славянский университет, г. Кишинев, Молдова
A fost elaborată o metodă nouă, înalt sensibilă, de depistare a esterazei pe gel prin combinarea indigo şi coloranţilor tetrazolii. Identificarea alelelor E4 esterazei din rădăcinile plantulelor liniilor inbrede s-a efectuat utilizând PAA-gel stan-dard. A fost depistată o diferenţă semnificativă în spectrele esterazei la inbrezi şi hibrizi. Ca punct de referinţă a fost luată izoforma esterazei E8. S-a constatat că E4, formă descrisă anterior, ce conţine o serie de linii pe gel, este artefact al interacţiunii dintre hidrixamatul ciclic şi ramurile lizinei din E4 esterază. A fost stabilit că E4 esteraza este monomer. Este dată modalitatea de descompunere a esterazei E8 în monomeri. Ea este heterodimer. Specificitatea substratului şi link dimerului prin intermediul grupurilor de sulfhidril poate identifica E8 esteraza ca acetilcolinesterazei.
Cuvinte-cheie: esterază, E4, E8 izoenzime, acetilcolinesteraze, metoda înalt sensibilă, depistarea esterazei, linii inbrede şi hibride ale porumbului.
ESTERASE ISOZYMES OF MAIZE INBRED LINES AND HYBRIDS The separation of esterase isozymes from seedling of maize was carried out using standard disc PAGE technique.
Significant differences were observed in esterase pattern in maize inbred lines and their hybrids. The esterase E8 enzyme, whose structural locus was demonstrated to be elsewhere in the genome, was referred as the mark for identification esterase isozymes on the gel. New highly sensitive method to detect the esterase by means of combination of indigo and tetrazolium dyes is developed. It has be found that the E4 esterase described before as being composed in series lines on the gel is really the artifact of interaction between cyclic hydroxamate molecules with lysine groups of this isozyme. Additive patterns of inheritance E4 esterase isozymes in hybrid produced from parental lines have been shown that the esterase E4 isozymes are the monomers. There has found the new way to split E8 esterase into monomers and it is prown as being a heterodimer. The substrate specificity and the dimer link by means of the sulphydryl groups can identify E8 esterase as acetylcholinesterase.
Keywords: esterase, E4, E8 isozymes, acetylholinesterase, highly sensitive method to detect esterase, maize inbred lines and hybrids.
Введение
Для изучения молекулярно-генетических процессов жизнедеятельности, идентификации геноти-
пов возделываемых растений, среди которых заметное место занимает и кукуруза, уже общепринято
применять энзимы. При этом большое внимание уделяется эстеразе, используемой при изучении ин-
бредных линий и гибридов кукурузы, для чего создана номенклатура ее изозимов [9, 13, 17-19] и про-
ведено их хромосомное картирование. Однако для использования этого банка данных необходим
ключ для узнавания отдельных изозимов на цельном электрофоретическом спектре, в соответствии с
общепринятой номенклатурой их обозначения. Но в большинстве научных публикаций указаны толь-
ко части того геля, на котором был проведен электрофорез, чтобы выделить ту изоформу, о которой
идет речь, а общая картина остается неизвестной. В дополнение к этому необходимо учитывать, что
все изозимы эстеразы кукурузы были открыты, идентифицированы и генетически прокартированы, в
основном, на американском континенте и на американских инбредных линиях кукурузы.
Совершенно очевидно, что, не владея способом узнавания изоформ эстеразы в общем спектре,
любое проведение экспериментов и изложение полученных данных почти лишено научной информа-
тивности, поскольку как для самого автора, так и для остальных исследователей остается не ясным, о
какой из изоформ из суммарного спектра эстеразы идет речь в публикации. Этими недочетами стра-
дают все публикации отечественных исследователей и ряда европейских, поскольку у нас до сих пор
не все инбредные линии изучены и прокартированы по энзиму эстеразы. К служебным же результа-
там исследований американских ученых доступа нет, несмотря на то, что номенклатура эстераз куку-
рузы начала создаваться более 45 лет тому назад. Поэтому вполне естественно, что пока в статьях
российских и других авторов предложенная американскими учеными номенклатура изоферментов
эстеразы в полной мере не использована [6, 7, 10, 11, 22].
Попытки уклоняться от научных дискуссий на общепринятом языке путем введения своей номен-
клатуры, вопреки уже существующей, вряд ли способствуют научному прогрессу. Здесь, как и вообще

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
120
в науке, необходимо вести научный диалог по общепринятым обозначениям, введенным первооткры-
вателями этих изоформ эстераз, и не осуществлять подмену предмета исследования и обсуждения
своими мнениями. Кроме этих факторов, существенное значение имеет то, что при создании различ-
ными авторами номенклатуры изоферментов эстеразы при электрофорезе использованы различные
гелевые носители: целлюлозный, агаровый, крахмальный, полиакриламидный с различными буфер-
ными системами, что не позволяет сравнивать результаты различных исследователей. Имеется обзор
литературы по проведенным вплоть до 1986 года исследованиям по изозимам эстеразы кукурузы [1],
где также показано, что изозимный спектр существенно зависит от применяемого электрофоретиче-
ского буфера. Эта номенклатура изозимов эстеразы дополнена новой, обнаруженной нами мембранно-
связанной изоформой эстеразы [12]. Номенклатуру эстераз начал создавать Д.Шварц, разделив на крахмальном геле в боратной буферной
системе эстеразы эндосперма кукурузы на стадии молочной спелости и обозначив их в порядке опи-сания как Е1, Е2 и Е3 изозимы. Однако в боратной буферной системе подвижность белков в значитель-ной степени определяется размером ковалентно связанного с ними углеводного фрагмента, и ввиду значительных погрешностей методики эти изоформы были оставлены без внимания. Настоящее раз-витие номенклатура получила после применения в качестве носителя при электрофорезе полиакрил-амидного геля в других буферных системах, но порядок их наименования был продолжен и появилась изоформа Е4, впервые описанная Харрисом [15], который на основании еѐ подвижности к аноду идентифицировал 4 аллели: наиболее медленную – С, и наиболее подвижную – F аллель, так что между их подвижностями располагаются другие при соотношении C<D<E<F. При этом было показано, что Е4 изоформа является мономером и локализована на третьей хромосоме [16].
Цель данной публикации – показать на общей картине результата электрофореза на геле расположе-ние изозимов, узнать их и, соответственно этому, изложить собственные экспериментальные данные: показать расположение Е4 эстеразы на общем виде геля по отношению к стартовой линии и к повсе-местно присутствующей изоформе Е8 эстеразы, являющейся димером и локализованной на коротком плече хромосомы 3 [20], описать характерные свойства Е4 эстеразы, идентифицировать ряд молдав-ских инбредных линий кукурузы по аллелям изофермента Е4 эстеразы. Показать на геле расположение открытой нами мембранной эстеразы [12]. Для стандартизации условий электрофореза нами применена стандартная методика прерывыстого (диск) электрофореза в щелочной буферной системе [14].
Материалы и методы исследования Семена различных инбредных линий и гибридов кукурузы проращивались в растильнях между
двумя слоями фильтровальной бумаги, увлажненной слабым 0,2 г/л раствором NaCl в термостате при 22
0С. Из одного проростка на 6-7 день проращивания нами отделялся корень, обсушивался снаружи
фильтровальной бумагой, взвешивался и измельчался при 0 С в ступке над капроновой мелкоячеис-той тканью с равным по весу объемом раствора 20% сахарозы на 0,063 M Трис HCl формирующем буфере с рН 6,9. Для связывания присутствующего в тканях кукурузы алкилирующего NH2 и SH группы белков, циклического гидроксамата DIMBOA [16], перед растиранием корня в ступку добав-ляли 0,1 г уравновешенного с дистиллированной водой слабоосновного анионита АН-15 в хлоридной форме. Гомогенат отжимали через капроновую ткань и супернатант центрифугировали до просвет-ления. В ячейку геля вносили 0,1 мл надосадочного раствора. В связи с тем, что каждая дорожка на электрофорезе представляет один генотип, а чистота исследуемого материала не была гарантирована, то между отдельными зимограммами существует некоторое расхождение вследствие генетического полиморфизма исследуемого генетического материала.
Электрофорез проводили в ступенчатой буферной системе с трис-глициновым буфером в 7,5% полиакриламидном пластинчатом геле толщиной 1,5 мм. Инкубация гелей на ферментативную актив-ность проводилась в 0,1 М фосфатном буфере с рН 8,3. В качестве субстрата мы использовали вначале
индоксилацетат, -нафтил и -нафтилацетаты или смесь последних двух. Суммарное содержание суб-страта составляло 1 мг/мл. Для одновременного азосочетания образующихся нафтолов в реакционную среду вносили соль прочного синего ББ в количестве 0,5 мг/мл инкубационного раствора.
Результаты и их обсуждение
Публикация по Е4 эстеразе [15, 16] настолько лаконична, что совершенно отсутствует методика
извлечения фермента, условия его электрофореза и окрашивания. Поэтому нами были начаты поиски

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
121
соответствующих методов исследований, позволяющих визуально на общем спектре увидеть эти
изоформы. После серии экспериментов с использованием боратной, вероналовой и трис-глициновой
буферных систем для выделения эстеразы и еѐ электрофореза, нам удалось установить, что наиболее
подходящим буфером для извлечения Е4 эстеразы, чтобы она на электрофоретическом геле давала
то изображение, которое представил в своих публикациях Харрис [15, 16], является трис-хлоридный
буфер. А для самого электрофореза необходимо использовать классический прерывистый (диск)
электрофорез по Дэвису [14].
В таких условиях на геле можно видеть одну аллель Е4 эстеразы, представленную серией полос в
виде гармошки, начиная от самой медленной и наиболее интенсивной, и последующих, градуально
снижающихся по интенсивности, по направлению их электрофоретического движения к аноду (рис.1).
Причем, число видимых линий в серии варьирует от трех до пяти при использовании гелей с суб-
стратом -нафтилацетатом в сочетании с солью прочного синего ББ. При использовании одного ин-
доксилацетата чувствительности методики недостаточно, чтобы видеть все эти серии полос. Наи-
большее число полос видно при использовании смеси индоксилацетата и нитросинего тетразолия.
Анализируя условия, необходимые для того, чтобы видеть то изображение Е4, которое представил
Харрис в своих публикациях, нам стало ясно, что серия полос – это артефакт, неизбежно создаваемый
плохими условиями выделения изоферментов из растительной ткани. Как известно, в клетках кукурузы
находится глюкозид циклического гидроксамата DIMBOA, который гидролизуется при измельчении
растительной ткани с освобождением агликона, алкилирующего свободные сульфгидрильные и амино-
группы белков и ферментов. Поэтому при извлечении фермента эстеразы в буфере без защитной
добавки полиамидного порошка, поливинилпирролидона или цистеина, он алкилируется по свободной
аминогруппе, боковым лизиновым группам, а также по свободным сульфгидрильным группам.
Рис.1. Участок геля с изображением аллелей Е4 эстеразы из корней кукурузы в порядке увеличения их подвижностей к аноду при электрофорезе по Дэвису при рН 8,9: C<D<E<F ( артефактные серии изоформ по
Харрису [15,16] как результат алкилирования лизиновых остатков); C’,D’,E’, F’ – нативные аллели Е4 эстеразы.
Одна молекула DIMBOA способна алкилировать две близлежащие аминогруппы лизиновых остат-
ков эстеразы. В результате молекула эстеразы дискретно уменьшает свой положительный заряд на 2
единицы, что эквивалентно соответствующему увеличению еѐ отрицательного заряда, и как результат–
дискретному возрастанию еѐ подвижности к аноду, с одновременным уменьшением ее ферментативной
активности. Такая алкилированная форма Е4 эстеразы будет дискретно отличаться как по подвижности,
так и по активности фермента от исходной еѐ нативной формы и создавать артефакт наличия мно-
жественности существования этого изозима [3]. Поскольку Е4 – один из самых малоподвижных изо-
ферментов в щелочной буферной системе, то он имеет наименьший отрицательный заряд и, следова-
тельно, наиболее обогащен аминогруппами и более сильно алкилируется. Появление при электрофо-
резе серии полос c градуально уменьшающейся интенсивностью по направлению электрофоретичес-
кого движения алкилированного по лизиновым остаткам белка хорошо доказано в исследованиях на
лизинбогатых гистонах [8]. Отсюда, и количество линий в каждой серии Е4 эстеразы также отражает
количество алкилированных лизиновых остатков Е4 эстеразы, при которых эстераза еще проявляет
свои ферментативные свойства.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
122
Для доказательства этого мы измельчили корни кукурузы в трис-хлоридном буфере с добавлением анионита АН-15, представляющего высокомолекулярный полимер с боковыми аминогруппами. В результате на электрофореграммах эстеразы корней кукурузы Е4 эстераза лишилась серии дискрет-ных полос с убывающей по направлению к аноду ферментативной активностью и на геле появилась одна линия более высокой интенсивности. Эти аллели на рис.1 нами правильно обозначены как C’, D’, E’, F’. Вместе с тем то, что аллели Е4 эстеразы можно видеть в виде серии полос, помогло нам идентифицировать этот изофермент на общей картине энзимограммы на геле. Таким образом, появилась вторая опорная точка, в дополнение к известному положению Е8 эстеразы.
Пользуясь выявленными нами условиями выделения и электрофореза эстеразы, мы изучили спект-ры инбредных линий кукурузы для идентификации их по аллелям Е4 эстеразы (рис. 2). В результате установлено, что наименее подвижная аллель Е4 эстеразы – аллель С, присутствует у инбредной линии Од-140. Аллель D присутствует у целого ряда инбредных линий: П-346, МК-159, МК-109; аллель Е не обнаружена, аллель F присутствует у линий МК-302, МБ-201, ВИР-44, ВИР-38, П-502.
Исходя из теоретических положений учения о химических связях, мы разработали способ допол-нительного разделения изоферментов эстеразы по субстратной специфичности на топоизомеры. Квантово-механическое рассмотрение сложноэфирной связи показывает, что между карбонильным и эфирным кислородами возможен резонанс валентных связей, и эти атомы расположены копланарно. В этой плоскости возникает цис- и транс-положение заместителей вдоль эфирной связи кислорода и углерода боковых заместителей кислотной и спиртовой составляющих молекулы сложного эфира. Эти цис- и транс-энантиомеры молекул сложных эфиров разлагаются соответствующими топоизо-мерными ферментами эстераз, которые также можно идентифицировать изомерными ингибиторами.
Например, в нервной клетке вырабатываются два топоизомера по эфирной связи ацетилхолина: цис-форма и транс-форма, первая из которых имеет сродство к никотиновым рецепторам (блокируе-мым тубокурарином) синаптических мембран скелетных мышц, в то время как вторая (транс-форма) разлагается мускариновыми рецепторами мышц вегетативной системы, где располагаются, соответ-ственно, топоизомерные к этим формам ацетилхолина ферменты ацетилхолинэстеразы, состоящие из димеров. В растительной ткани также находится фермент ацетилхолинэстераза, которая на геле после электрофореза до сих пор не идентифицирована. Были веские основания полагать, что Е8 эстераза, присутствующая во всех тканях и являющаяся димером, может принадлежать к одной из топоизоме-ров ацетилхолинэстеразы растений. Поэтому Е8 эстераза была подвергнута внимательному изучению.
Рис.2. Аллели Е4 эстеразы из корней инбредных линий кукурузы:
1. Од-140; 2.П-346; 3. Мк-159; 4. МК-109; 5. МК-302; 6. МБ-201; 7. ВИР-44; 8. ВИР-38; 9. П-502.
Слева греческими буквами обозначены топоизомерные формы эстераз.
Для идентификации топоизомеров мы использовали смесь из - и - нафтилацетатов в качестве субстратов для инкубации гелей. Для этого вносили уравновешенную по количеству мест азосочета-
ний смесь из них в пропорции 1:2, поскольку -нафтол имеет 2 места сочетания с азокомпонентой, а
-нафтол – только одно.
В качестве копулирующей азокомпоненты вносили соль прочного синего ББ. При этом зоны изофер-
ментов на геле, имеющие наибольшее сродство к -нафтилацетату, окрашивались в черный цвет, а
зоны изоферментов, имеющие большее сродство к -нафтилацетату, окрашивались в красный цвет. Используя этот метод, мы смогли разделить изоферменты эстеразы по сродству к различным субстратам

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
123
на - и -изоформы. Так найдено, что Е4 эстераза является -топоизомером эстеразы кукурузы, в то
время как вездесущая изоформа Е8 является -изоформой и может быть ацетилхолинэстеразой. Кроме Е8 свойства ацетилхолинэстеразы проявляет не идентифицированный нами еще один непостоянно и слабо проявляющийся на геле изоферемент, предшествующий по подвижности Е8 эстеразе. Это дает до-полнительный способ идентификации изоформ и аллелей на общем электрофоретическом спектре на геле.
Далее нами было изучено наследование Е4 эстеразы различными гибридами кукурузы. При этом было показано, что Е4 эстераза у гибридов представлена суммой серийных линий, и гибридной формы с промежуточной подвижностью не образуется, что подтверждает сведения Харриса о том, что Е4 эстераза является мономером. При этом у гибридов функционируют обе аллели Е4 эстеразы, так что содержа-ние ферментов каждой аллели в них в 2 раза меньше, чем у инбредных линий. Поэтому серии линий в аллелях представлены меньшим числом, т. к. остальные линии находятся за пределами обнаружения ферментативной активности с субстратом нафтилацетатом (рис. 3). Таким образом, фермент Е4 не образует гибридных димеров и поэтому гетерозигота представляет собой просто механическую смесь двух альтернативных молекулярных форм.
Рис. 3. Наследование Е4 эстеразы в корнях гибридов кукурузы исходя из данных
родительских инбредных линий. 1. Од-140; 2. Од-140х МК 302; 3. МК-302; 4. П-346; 5. П-346хП- 502;
6. П-502; 7. П-502хМБ-201; 8. МБ-201; 9. П-302х МБ-201.
При изучении различных ингибиторов метаболизма в процессе прорастания проростков кукурузы нами найдено, что ингибитор синтеза матричной РНК актиномицин Д не изменяет на геле спектра эстеразы из колеоптилей и корней проростков кукурузы. Таким же свойством обладает хлорамфени-кол. Это показывает, что в процессе прорастания ферменты эстеразы синтезируются на уже предсу-ществующих в рибосомах матрицах РНК, поскольку при проращивании семян в растворе актиноми-цина Д в концентрации 1 мг/мл спектр эстеразы корней и колеоптилей не отличается от такового при проращивании семян в воде. На спектр не повлияли такие соединения, как квартазин, примулин, N-диметиламид янтарной кислоты.
Значительное усиление окраски Е4 эстеразы на геле показал ремантадин, что позволяет считать его фактором, удлиняющим время пребывания матричной РНК на рибосомах. Ингибитор сульфгидриль-ных групп N–этилмалеинимид привел к разделению одиночной полосы Е8 эстеразы на две (рис. 4).
Рис.4. Изоферменты эстеразы колеоптиля: 1,2 – щитка; 3,4 – корня; 5,6 – проростка инбредной
линии ВИР-44. Показано расположение мембранной формы эстеразы Ем. Дорожка 7 представляет действие ремантадина, дорожка 8 – это действие N-этилмалеинимида.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
124
Это доказывает, что Е8 эстераза является гетеродимером. Для четкой демонстрации открытой нами мембранной формы эстеразы на этом геле, множественное возникновение серийных артефактных полос Е4 эстеразы предотвращено введеним в среду измельчения растительной ткани слабого анио-нита АН-15, намного большее количество свободных аминогрупп которого связали DIMBOA и защи-тили аминогруппы Е4 эстеразы от алкилирования. В этих условиях Е4 эстераза представлена одной нативной формой и еѐ серийные линии не накладываются на полосы других изоферментов. Мембран-
ная форма эстеразы Ем является типичной формой -эстеразы и присутствует в колеоптиле и щитке, но отсутствует в корнях. Наибольшая активность Ем эстеразы проявляется в щитке. Изоформа Ем солюбилизирована нами путем введения в среду измельчения растительной ткани детергентов диги-тонина, холата, дезоксихолата натрия, додецилсульфата натрия. Наибольшую сохранность всех изофер-ментов проявляет дигитонин, однако при его применении на геле появляется слабый затеняющий фон. Холат и дезоксихолат натрия несколько снижают активность полос и также не свободны от зате-няющего фона. Совершенно прозрачный гель с сохранением высокой активности Ем дает применение детергента додецилсульфата натрия, однако при этом снижаются активности иных изоферментов, таких как Е4 и Е8.
Чувствительность обнаружения изоферментов в значительной степени определяется методикой обнаружения изоферментов на геле. Нами найдено, что с одним индоксилацетатом чувствительность определения изоферментов на геле ниже, чем с использованием нафтилацетата совместно с азоком-понентой. Но и этот метод является все же недостаточно чувствительным для обнаружения фермен-тов, находящихся в эндосперме кукурузы. В результате теоретического рассмотрения механизмов действия различных субстратов в ферментативных реакциях нами предложена новая субстратная смесь, состоящая из 1 мг/мл индоксилацетата и 0,1 мг/мл тетранитротетразолия синего в 0,05 М фос-фатном буфере с рН 8,2. Эта смесь обладает чувствительностью обнаружения эстеразы в 5-10 раза бóльшей, чем используемая до сих пор смесь из нафтилацетата и азокомпоненты. В новом методе соль тетразолия выступает в качестве окислителя белого индиго, появляющегося в результате действия эстеразы на индоксилацетат. Восстанавливаясь, тетразолий образует интенсивно окрашенный форма-зан, а индоксил, окисляясь, образует в свою очередь интенсивно окрашенное индиго. Отсюда резуль-тирующая окраска является их суммарным действием [2, 5]. В этой энзиматической реакции обе исход-ные компоненты смеси дают интенсивно окрашенные синим цветом нерастворимые осадки коллоид-ного индиго и мелкокристаллического формазана.
Интенсивность окраски формазана, образующегося при восстановлении тетразолия, намного пре-вышает интенсивность окраски азокрасителей, образующихся в результате азосочетания нафтола с азокомпонентой. Максимальная чувствительность этой смеси достигается при отсутствии кислорода в инкубационной среде, куда гель помещается для окраски. Удобно использовать ванночку с раство-ром субстрата, выдержанным 2-3 часа в эксикаторе над поглотительным раствором из 10% смеси сульфита и карбоната натрия. Эффективность удаления кислорода возрастает, если в поглотительный раствор внести еще и гидрохинон. При использовании этой методики выявления изоферментов эсте-разы на геле и при измельчении растительной ткани в присутствии анионита АН-15, нам удалось по-лучить значительно более интенсивный спектр эстеразы на геле (рис. 5).
Рис. 5. Эстераза из первого листа проростков инбредных линий и гибрида Пионер: 1,2,3 – инкубация гелей в
среде азосочетания; 4,5,6 – инкубация в индоксилацетате совместно с тетранитрометразолием синим;
1,4 – линия П-346; 2,5 – гибрид П-346хП-502; 3,6 – линия П-502.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
125
Из рисунка 5 видно, что интенсивность окраски изоферментов намного выше в среде индоксилаце-
тата с тетранитротетразолием синим. Преимущество нашей инкубационной среды наиболее сильно
видно при электрофорезе белков из эндосперма прорастающих семян, где многократно увеличивается
количество полос и их интенсивность.
Высокая подвижность Е8 в этих условиях свидетельствует о том, что этот изофермент является кис-
лым белком с большим количеством карбоксильных остатков. Значительно меньшее содержание боковых
аминогрупп лизиновых остатков делает его устойчивым к инактивирующему ферментативную актив-
ность действию присутствующего в тканях кукурузы циклического гидроксамата ДИМБОА путем алки-
лирования им свободных аминогрупп белков и ферментов. При этом на геле это видно как по значи-
тельному уменьшению интенсивности окраски линий изоферментов, так и по их некоторой размытости,
вызванной вариацией подвижности вследствие случайного изменения молекулярного веса изофермента
и различной степени алкилирования его аминогрупп. Однако из-за малого содержания аминогрупп Е8
эстераза почти не инактивируется, мало размывается и на геле видна резко и ярко.
Димерность Е8 эстеразы и способ ее разделения на мономеры описаны в более ранней публика-
ции [4]. Способ состоял из процедуры выращивания семян кукурузы в среде алкилятора свободных
сульфгидрильных групп белков и последующего электрофореза. Так, при проращивании семян куку-
рузы в течение 6 дней в растворе, содержащем 1 мг/мл N-этилмалеинимида, алкилирующего белки и
ферменты по свободным SH группам, и последующем выделении фермента эстеразы и его электро-
форезе в щелочной буферной системе, найдено раздвоение одиночной полосы Е8 эстеразы на две.
Однако при введении этого ингибитора в среду измельчения, на геле полоса Е8 не раздваивалась. Это
свидетельствует о том, что после окончания трансляции на рибосомах, мономеры Е8 эстеразы неко-
торое время еще существуют со свободными SH-группами, когда они доступны для алкилирования, а
затем димеризуются в последующем процессинге в цитоплазме путем образования между мономера-
ми дисульфидной связи. Этот способ позволяет разделить Е8 эстеразу на мономеры. При этом ока-
залось, что Е8 эстераза является гетеродимером.
Выводы Описан изофермент эстеразы Е4 из корней проростков кукурузы. Показана локализация его на
общем виде электрофоретически разделенных эстераз. Доказано, что серии полос Е4 эстеразы, пред-
ставленные в сообщении Харриса, являются артефактами и результатом алкилирования лизиновых
остатков Е4 эстеразы циклическим гидроксаматом DIMBOA. Предложен способ идентификации
топоизомеров эстеразы на общем геле. Все эстеразы методом специфического окрашивания разделе-
ны на - и -топоизомеры. Инбредные линии кукурузы идентифицированы по аллелям Е4 эстеразы.
Методом электрофореза гибридов из этих линий показано, что Е4 эстераза является мономером и в
гибридах представлена механической смесью аллельных изозимов. Описана мембранно-связанная
эстераза Ем, присутствующая в колеоптилях и щитках проростков кукурузы, продемонстрировано еѐ
расположение на геле. Приведен метод выделения этой эстеразы. Найден способ разделения димер-
ной Е8 эстеразы на мономеры и показано, что Е8 эстераза является гетеродимером, связанным ди-
сульфидной связью. В соответствии с таким строением и по субстратной специфичности Е8 эстераза
идентифицирована как ацетилхолинэстераза. Найдена более чувствительная субстратная смесь, со-
стоящая из индоксилацетата и тетранитросинего тетразолия, для обнаружения изозимов эстераз на
геле. Чувствительность ее более чем в 5 раз превышает чувствительность применявшейся до сих пор
среды для инкубации гелей со способом выявления полос путем азосочетания.
Литература:
1. БАБИЦКИЙ, А.Ф., КАНИЩЕВА, Е.Ф. Особенности электрофореза изоферментов эстеразы кукурузы. В:
Селекционно–генетические методы повышения урожайности, качества и устойчивости полевых культур.
Труды Кишиневского СХИ. Кишинев, 1988, с.56-61.
2. БАБИЦКИЙ, А.Ф., ТОМА, З.Г. Высокочувствительный метод выявления зон эстеразной активности с индо-
ксилацетатом и солью тетразолия на электрофореграммах белков эндосперма инбредных линий и гибрида
кукурузы. В: Analele Ştiinţifice ale Universităţii de Stat din Moldova. Seria: Ştiinţe chimico-biologice. Chişinău,
2006, p.313-316.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
126
3. БАБИЦКИЙ, А.Ф., ТОМА, З.Г. Е4 эстераза инбредных линий и гибридов кукурузы. В: Plant agrobiodiversity.
Chişinău, 2006, с.102-112.
4. БАБИЦКИЙ, А.Ф. Разделение на мономеры Е8 эстеразы кукурузы. В: Biotehnologii avansate – realizării
perspective al III-lea Simpozion naţional cu participare internaţională. TEZE, 24-25 оctombrie 2013. Chişinău,
2013, р.31.
5. БАБИЦКИЙ, А.Ф., ТОМА, З.Г. Высокочувствительный метод выявления эстеразы при электрофорезе белков
эндосперма кукурузы. В: Украинская научно-исследовательская станция карантина растений ИЗР НААН.
Информационный бюллетень № 44: Фитосанитарная безопасность и контроль сельскохозяйственной про-
дукции. Бояны, 2013, с.20-24.
6. ПЕТРОВА, Н.Л., СУХОРЖЕВСКАЯ, Т.Б., ВОЙНИКОВ, В.К. Изменения изоферментов эстеразы у про-
растающих семян кукурузы. В: Оперативные информационные материалы СИФИБР. Иркутск, 1983, с.25–28.
7. ПЕТРОВА, Н.Л., СУХОРЖЕВСКАЯ, Т.Б., ВОЙНИКОВ, В.К. Выявление изоферментов эстеразы у пророст-
ков кукурузы методом диск-электрофореза в полиакриламидном геле. В: Оперативные информационные
материалы СИФИБР. Иркутск, 1983, с.28-32.
8. РОЗОВ, С.М., БЕРДНИКОВ, А.В. Определение числа остатков лизина, положительного заряда и молеку-
лярной длины гистонов Н1 и Н5 методом неполного сукцинилирования. В: Биохимия, 1982, т.47, № 8,
с.1378-1385.
9. СИМИНЕЛ, В.Д., БАБИЦКИЙ, А.Ф. Е1 эстераза инбредных линий и гибридов кукурузы. В: IV съезд
генетиков и селекционеров Молдавии. Кишинев, 1986, с. 99.
10. ТУРБИН, Н.В., ПАЛИЛОВА, А.Н., ПИКО, В.Н. Электрофоретическое изучение набора ферментов в пыль-
никах кукурузы с цитоплазматической мужской стерильностью. В: Генетика, 1976, том 12, № 8, с.14-21.
11. ШЕЛПАКОВА, О.Л. О возможности использования изоферментов при изучении апомиксиса у растений. В:
Генетические основы селекции. Ред. Д. В. Петров. Новосибирск, 1982, с. 81–85.
12. BABITSKY, A., TOMA, Z. Esteraza membranică la porumb. În: Rezumatele lucrărilor simpozionului national de
fiziologia plantelor. Bucuresti, 1995, nr.13, р. 5.
13. BRAUN, A.H., ALLARD, D. Inheritance of isozyme differences among the inbred parents of reciprocal recurrent
selection population of maize. In: Crop Sci., 1969, vol.9, no1, p.72–75.
14. DAVIS, B.I. Disc electroforesis. II. Method and application to human serum proteins. In: Ann. N. Y. Acad. Sci.,
1964, vol.121, p.404-428.
15. HARRIS, J.W. The E4 esterase. In: Maize Genet. Coop. New Letter, 1966, vol. 40, p.53–56.
16. HARRIS, J.W. Location of the E4 esterase locus on chromosome 3. In: Maize Genet. Coop. New Letter, 1968,
vol.42, p.72-74.
17. HENDRICH–SORBINO, E., CORDEIRO, A.R. Codominant isozyme alleles of markers of genetic diversity correlated
with heterosis in maize. In: Theoret. App. Genet., 1975, vol. 46, no4, p.197–199.
18. HUNTER, R.B., KONNENBERG, L.W. Isozyme characterization of corn inbreds and its relationship to single cross
hybrid performance. In: Canad. J. Genet. Cytol., 1971, vol.13, no4, p.649–655.
19. KLEASE, R.A., PHILIPS, R.L. Electrophoretric mutants as useful markers for chromosome abberations. In:
Genetics, 1972, vol.72, no3, p.537-540.
20. NEWTON, K., GOODMAN, M., STUBER, C. Localization of esterase E 8 on the short arm of chromosome 3. In:
Maize Genet. Coop. New Letter, 1982, vol. 56, p.154–155.
21. PEREZ, F.J., NIEMEYER, N.N. Reaction of DIMBOA with amines. In: Phytochemistry, 1989, vol.28, no7, p.1831-
1834.
22. SCANDALIOS, I.G. Tissue specific isozyme variations in maize. In: Journ. Hered., 1964, vol.55, no4, p.281-285.
Prezentat la 16.09.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
127
ОТ ЧЕГО ЗАВИСИТ СОДЕРЖАНИЕ ФОСФОРА В СЕМЕНАХ ПШЕНИЦЫ
Андрей БАБИЦКИЙ
Славянский университет, г. Кишинев, Молдова
FACTORI CE DETERMINĂ CONŢINUTUL DE FOSFOR ÎN SEMINŢELE DE GRÂU
În condiţii de câmp a fost studiată acţiunea nutriţiei minerale la trei regimuri de umiditate a solului asupra conţinu-
tului de fosfor în seminţele de grâu şi capacităţii lor de rodnicie. Acumularea excesivă a fosforului în seminţele cu capa-
cităţi diminuate de rodnicie are loc în condiţiile de deficit hidric în sol. În condiţii optime de hidratare a solului se for-
mează seminţe cu conţinut mediu de fosfor şi cu capacităţi excelente de rodnicie. În condiţiile surplusului de umiditate a
solului seminţele posedă cel mai redus conţinut de fosfor. Rezultatele obţinute interpretează procesul de descompunere
a ţesutului vegetativ în faza finală a ontogenezei şi transportul hidrolizatelor substanţelor organice în seminţe.
Cuvinte-cheie: grâu, nutriţie minerală, umiditate a solului, conţinut de fosfor în seminţe, capacitate de rodnicie a
seminţelor.
WHAT DETERMINES THE PHOSPHORUS CONTENT IN WHEAT SEEDS
The phosphorus content and yielding capacity of the wheat seed produced during field experiments under the influence
of 13 variants of mineral nutrition upon 3 modes of soil moisture have been studied. The greatest accumulation of
phosphorus in seeds with their less yielding capacity occurs upon water deficit soil. The intermediate phosphorus content
in seeds with the higher yielding capacity of the seeds are formed at the optimum soil moisture. The lowest phosphorus
accumulated in waterlogged soil. Between the phosphorus content in the seeds and soil phosphorus level is no direct
relationship. These data are interpreted as the result of the decay of vegetative tissues in the final stages of ontogeny and
transfer the products of hydrolysis of organic compounds into the wheat seeds.
Keywords: wheat, mineral nutrition, soil moisture, seed phosphorus, seed yielding capacity.
Введение
Одним из важнейших показателей, определяющих как алиментарное качество зерна пшеницы, так
и посевные и урожайные качества (УК) ее семян, является содержание в них фосфора. Более 80%
всего фосфора семян находится в виде фитиновой кислоты [15].
Продукты питания из зерна пшеницы с высоким содержанием фитина у людей с низкой активно-
стью фермента фосфатазы в слюне вызывают отложение камней на зубах, и этим людям необходимо
диетическое питание продуктами из низкофитиновой пшеницы. Младенцам также противопоказано
одностороннее питание пшеничной манной кашей из-за того, что фитиновая кислота блокирует вса-
сывание кальция в пищеварительной системе [15] и это создает дефицит кальция в организме и может
создать рецедив рахита. Напротив, людям с онкологическими заболеваниями в области толстой кишки
необходима диета с продуктами питания из высокофитиновой пшеницы, поскольку фитин проявляет
протекторное действие вследствие его антиоксидантной активности.
Cведения о содержании фосфора в семенах имеют важное значение и для семеноводов, поскольку
среди них широко распространено мнение [11,16-18], что семена с высоким содержанием фосфора и
белка обладают высокими УК. Поэтому в сельском хозяйстве во всех научно-исследовательских
институтах и на опытных станциях по заказу агрономов и селекционеров проводятся десятки тысяч
анализов образцов семян пшеницы, чаще всего под номерным шифром без указания условий репро-
дукции и сорта, на содержание в них белка и фосфора. Более того, эти образцы, как правило, пред-
ставляют собой произвольно отобранную порцию семян из валовой партии, поскольку агрономы
исходят из того, что содержание фосфора в семенах, главным образом, определяется генотипом сорта
и в некоторой степени варьирует в зависимости от условий их репродукции. Как правило, такой фак-
тор, как фракционный размер семян, представленных на анализ, агрономами не учитывается. Однако
по нашим данным это существенный фактор, что и демонстрируют нижеприведенные данные.
Результат анализа каждого образца представляется при этом одной цифрой и в таком виде выдается
агрономам. Далее эти данные в неосмысленном виде, как полученные ими лично, публикуются в таб-
личной форме и такими таблицами в течение многих десятилетий переполняются страницы журнала
«Агрохимия». Таким образом, обессмысливается напряженный труд квалифицированных биохимиков.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
128
Хотя результат этого анализа только тогда представляет ценность и достоверность, когда он непо-
средственно исходит от первоисточника с указанием всех факторов, влияющих на содержание фос-
фора в функционально-графическом виде.
Имея один показатель, график построить невозможно, ибо через одну точку можно провести бес-
численное число линий с различной степенью наклона и кривизны. Поэтому традиционно представ-
ленные в литературе данные в табличной форме бессмысленны, и в них почти невозможно отделить
главные факторы, определяющие содержание фосфора в семенах, от второстепенных.
Материалы и методы исследования
Опыты по выявлению влияния различных доз внесения минеральных удобрений и их сочетаний и режимов влажности почвы на взаимосвязь между урожаем зерна и содержанием в нем белка были про-
ведены на яровой твердой пшенице Харьковская 46 в полевых условиях Одесской области. Норма высева 5 млн. семян/га, глубина заделки 4 см, ширина междурядьев 15 см. Опыт состоял из 13 вариантов раз-
личных сочетаний минеральных удобрений, обозначенных номерами от 1 до 13 в следующем порядке: 1 - без удобрения; 2 - N30P30K30; 3 - N60P60K60; 4 - N90P90K90; 5 - N0P60K60; 6 - N30P60K60;
7 - N90P60K60; 8 - N60P60K0; 9 - N60P60K30; 10 - N60P60K90; 11 - N60P0K60; 12 - N60P30K60;
13 - N60P90K60. Удобрения вносились поздней осенью под зяблевую вспашку после парового пред-шественника. Выращивание растений производилось при трех режимах влажности почвы: 1 - без по-
лива (дефицит влаги); 2 - 75% от полной почвенной влагоемкости (ППВ) (оптимум почвенной влаги); 3 - 90% ППВ (избыток влаги). Итого было 39 вариантов сочетаний уровней минерального питания и
режимов влажности почвы. Каждый вариант засевался в 6-кратной повторности при рандомизирован-ном размещении вариантов в пределах однородного уровня влажности почвы. Итого было засеяно
234 делянки по 18 квадратных метров каждая. Необходимый режим влажности почвы обеспечивали поливом водопроводной водой с помощью передвижной дождевальной установки, навешенной на
трактор ДТ - 54. Уборку урожая осуществляли комбайном Сидмайстер. Очищенное зерно взвешивали для учета урожая по вариантам. Зерно с каждого варианта фракцио-
нировали на зерновых решетах и обозначали серединой интервала между ширинами ячеек проходного и задерживающего решет. Так, фракция 2,6 мм получалась при проходе зерна через ячейки решета
2,75 мм и задержке на сите с шириной ячеек 2,5 мм. Полученные образцы семян высушивали до посто-янного веса при 105°С в термостате в течение 4 часов. Затем зерна измельчали на мельнице “Пируэт”
и на аналитических весах брали навеску муки для определения фосфора. Чаще всего содержание фосфора в растительной ткани определяется путем сухого сжигания образ-
цов в тиглях в муфельной печи. При этом происходит значительная потеря анализируемого материала
за счет вылетающих при этом дымовых частиц и отсюда погрешности анализа превышают 10-15% и содержание фосфора дается в виде миллиграммов P2O5 на общий вес золы или в процентах на сухой
вес биологического образца. Для увеличения точности анализа применен метод минерализации в серной кислоте с катализато-
ром – перекисью водорода. При этом фосфор определяли фотометрически, с использованием диффе-ренциальной колориметрии на ФЭК - 56 по методу Таусски и Шора [1, 2]. Результаты представлены в
классических биохимических единицах измерения количества фосфора в биологическом материале – в микромолях неорганического фосфата на один грамм сухого веса биологического материала.
Однако при выяснении взаимосвязи содержания фосфора с урожаем и УК эти данные приведены к общепринятой агрохимиками единице измерения – в процентах P2O5 на сухой вес. Окончательные
данные представлены на рисунках в графическом виде с помощью компьютера по программам Excel и Paint.
Результаты и их обсуждение
Во-первых, найдено, что размер фракции семян пшеницы, представленных на анализ, существенно
влияет на полученные результаты содержания фосфора в них. Таким образом, размер семян можно обозначить как геометрический или конфигурационный фактор их изменчивости по содержанию
фосфора. Из факторов внешней среды наиболее существенным является уровень влажности почвы,
при котором выращивались материнские растения, на которых были репродуцированы эти семена. Влияние уровней и доз минеральных удобрений не имеет четкой функциональной зависимости и
также в сильной степени модифицируется условиями влажности почвы.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
129
В результате проведенных анализов получены три семейства графиков (рис. 1), определяемых размером семян, уровнями и дозами внесенных удобрений и режимом влажности почвы. При этом все графики имеют седловидную форму такого вида, что более мелкие и более крупные фракции семян показывают бóльшее содержание фосфора, чем средняя наиболее массовая фракция семян в интервале 2,5- 2,75 мм их ширины. Следовательно, агрономы должны четко понимать, что представ-лять в лабораторию на биохимический анализ не откалиброванные по размеру семена пшеницы не имеет смысла, поскольку различия по условиям выращивания перекрываются различиями фракцион-ного состава, так что различия, вызванные в минеральным питанием окажутся меньше, чем различия, вызванные величиной крупности семян. Что касается уровня влажности почвы, то самый высокий уровень фосфора наблюдается в партии семян, выращенных без дополнительного полива при естест-венном увлажнении за счет атмосферных осадков.
Ранее такой же характер функциональной зависимости нами был найден при анализе семян пше-ницы на содержание в них белка [7,8] и дана схожая рекомендация по предоставлению образцов зерна на анализ по содержанию белка. Учитывая, что основным белком семян пшеницы является глиадин [8] и основным фосфорсодержащим компонентом является фитин [15], можно считать, что глиадин и фитин образуют единый глиадино-фитиновый комплекс в эндосперме зерна.
Семена, репродуцированные при 75% ППВ почвы, на графике зависимости содержания фосфора от размера семян также показывают характерную впадину на уровне фракции средних семян, но се-мейство этих кривых несколько опущено вниз по оси ординат. Семена, полученные при избыточном увлажнении почвы 90% ППВ, имеют пониженное содержание фосфата при наименее выраженной седловине в районе средних размеров семян.
Из рис.1 явствует, что наибольшее влияние на содержание фосфора в семенах наиболее массовой
фракции 2,5-2,75 мм оказывает не минеральное питание, а режим влажности почвы. Наиболее четко вышеописанные закономерности видны на графике содержания среднего значения фосфора в преде-
лах каждой влажности семян. При этом наибольшие различия между каждыми режимами влажности видны на фракции крупных семян. Это является дальнейшим подтверждением того факта, что круп-
ные семена в колосе пшеницы поглощают питательные вещества не только из стержня колоса, но и
перекачивают их из фракции средних семян [3, 4]. Уровень минерального питания имеет слабое влияние на содержание фосфора в семенах и перекрывается факторами размера семян и на графике
наблюдается как некий разброс кривых вокруг их усредненных данных, представленных с правой стороны и внизу рис.1.
Повышенные величины содержания белка и фосфора в семенах, полученных в условиях дефицита
влаги, показывают, что главным фактором, определяющим эти показатели, является отток питатель-
ных веществ из листьев в семена при дегидратации листьев и стебля пшеницы. Дефицит влаги уси-
ливает процесс дегидратации вегетативной ткани растения и вызывает апоптоз клеток и тканей с
переносом в подвижном виде по флоэмной сосудистой системе продуктов распада биологического
материала к репродуктивным органам и поглощение их семенами. При этом сосуды ксилемы закупо-
риваются, и это усиливает состояние дегидратации листьев и стебля, которое необходимо для за-
ключительной стадии налива семян и их обезвоживания. Важность онтогенетически управляемого отмирания некоторых клеток и тканей и использования
продуктов их распада как источника энергетического и информационного материала для формиро-
вания органов размножения растений, начиная с процессов гаметогенеза и до заключительных ста-
дий формирования зародышей и семян, обстоятельно описана в многочисленных исследованиях С.Колесникова [12,13]. На заключительной стадии жизни растения при формировании репродуктив-
ных органов эти процессы охватывают все органы и ткани вегетативной системы и имеют главенст-вующее значение [12,13].
Условия высокой влажности почвы или дожди в конце онтогенеза на стадии созревания зерна пре-
пятствуют процессу заключительной стадии дегидратации вегетативной ткани и ее ксилемные сосу-
ды продолжают функционировать, тормозится процесс распада ее клеток и тканей и отток в колос
веществ, состоящих из продуктов распада, значительно снижается, что вызывает пониженное содер-
жание в семенах азота и фосфора. Таким образом, для получения высококачественного зерна пшеницы
на конечных стадиях роста растений, в заключительный момент формирования семян, должны про-
ходить согласованные процессы распада вегетативной ткани [19,20-23], ее дегидратации и переноса
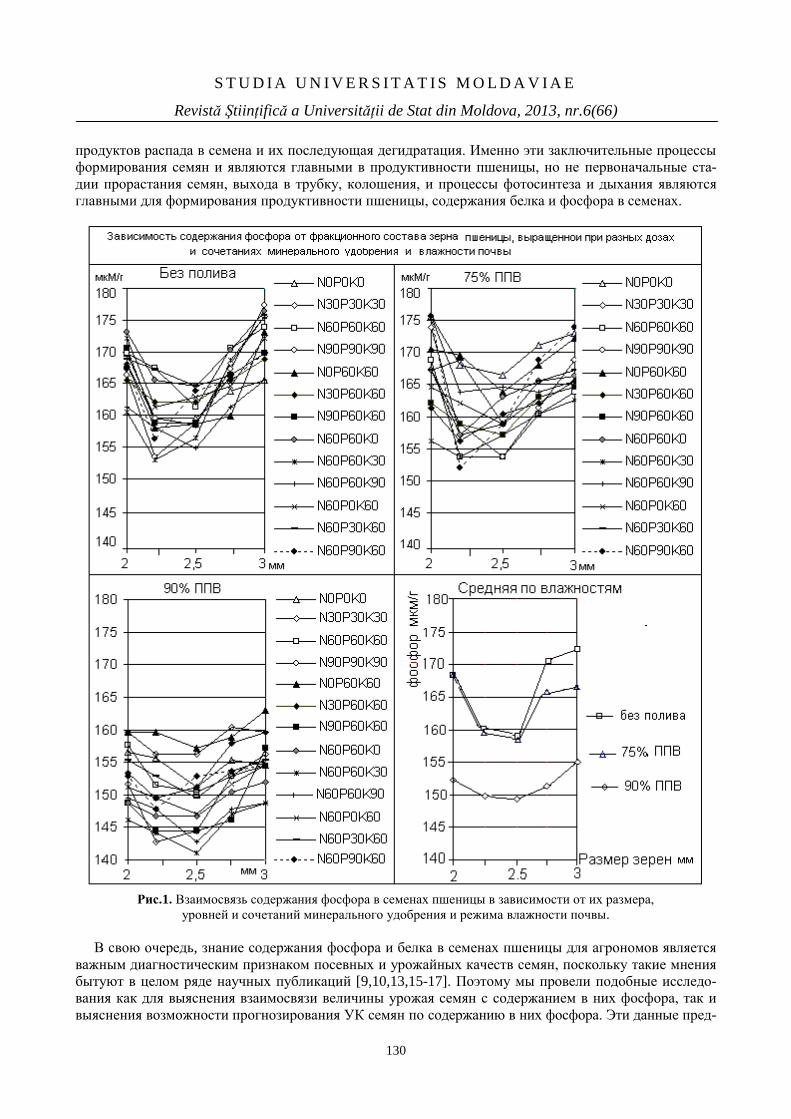
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
130
продуктов распада в семена и их последующая дегидратация. Именно эти заключительные процессы
формирования семян и являются главными в продуктивности пшеницы, но не первоначальные ста-
дии прорастания семян, выхода в трубку, колошения, и процессы фотосинтеза и дыхания являются
главными для формирования продуктивности пшеницы, содержания белка и фосфора в семенах.
Рис.1. Взаимосвязь содержания фосфора в семенах пшеницы в зависимости от их размера,
уровней и сочетаний минерального удобрения и режима влажности почвы.
В свою очередь, знание содержания фосфора и белка в семенах пшеницы для агрономов является
важным диагностическим признаком посевных и урожайных качеств семян, поскольку такие мнения
бытуют в целом ряде научных публикаций [9,10,13,15-17]. Поэтому мы провели подобные исследо-
вания как для выяснения взаимосвязи величины урожая семян с содержанием в них фосфора, так и
выяснения возможности прогнозирования УК семян по содержанию в них фосфора. Эти данные пред-

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
131
ставлены на рис.2, в котором содержание фосфора дано в Р2О2% на сухое вещество, как это принято
среди агрономов и селекционеров, чтобы данные рис.2 можно было сравнить с данными других
исследователей при оценке достоверности прогнозирования УК семян по содержанию в них фосфора.
Из левой части рис.2 видно, что непосредственной связи как между урожаем зерна в период воз-
действия различных доз и уровней минерального питания и режимов влажности почвы и содержа-
нием в нем фосфора не наблюдается, однако видна некоторая тенденция к уменьшению содержания
фосфора при повышении урожая. Однако это можно объяснить скорее тем, что самая высокая уро-
жайность достигается за счет усиления влажности почвы, и это является косвенным влиянием режима
влажности почвы на фосфор в зерне. Представленная в правой части рис.2 взаимосвязь между содер-
жанием фосфора в семенах, взятых из зерна, полученного, как описано выше, и определенных во
втором году их УК при выращивании без удобрений и при естественной влажности почвы, также
не показывает непосредственной связи. Однако точки на графике группируются согласно уровням
влажности почвы. Это свидетельствует, что неурожай не определяет содержание фосфора в зерне, а
некие другие факторы вегетации растений. Отсюда следует главный вывод, что прогнозировать УК
семян по содержанию в них фосфора невозможно.
Это расхождение двух процессов – формирование биомассы семян и содержание в них фосфора,
можно разрешить только признанием наличия иного фактора, влияющего на содержание фосфора в
семенах, которым является отток питательных веществ из вегетативной ткани в процессе гибели
клеток при апоптозе, активации гидролитических и протеолитических ферментов [20-23] при водном
дефиците в вегетативной ткани, последующей ее дегидратации на заключительных стадиях онтоге-
неза растений при формировании семян, косвенно связанной с величиной биомассы урожая, которая
определяется интегральной длительностью роста и развития растения. Стресс растений пшеницы,
вызванный водным дефицитом, индуцирует активацию многих генов, среди которых находятся и
гены, кодирующие эндопротеазы [22,23].
Рис.2. Взаимосвязь между урожаем семян и содержанием в них фосфора и между содержанием фосфора
в семенах и их урожайными качествами.
В агрономической литературе нередко можно встретить сведения о прямо пропорциональной
ответной отзывчивости накопления фосфора в семенах на внесенную в почву дозу фосфора. При
проведении данного опыта в этом направлении нами найдено, что такой прямой пропорциональности
не существует (рис.3). Слабое увеличение содержания фосфора в семенах в ответ на повышенное
фосфорное удобрение почвы наблюдается только при ее оптимальной и избыточной влажности.
В условиях дефицита почвенной влаги, воздействие фосфорного удобрения на содержание фосфора
в семенах в сильной степени сопряжено с действием элементов питания азота и калия. Наибольшее
количество фосфора в семенах накапливается при дефиците азота в почве при оптимальном и избы-
точном увлажнении, в несколько меньшей степени влияет дефицит калия в почве. Можно предполо-
жить, что это вызвано компенсаторной реакцией растения на дефицит этих элементов путем уси-
ления фосфорного обмена и соответственного заключительного переноса этого фосфора в семена.
Ранее отсутствие непосредственной связи между уровнем почвенного фосфора и количеством на-
копленного в семенах было показано в работах [10,11,17].

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
132
Рис.3. Влияние уровней и сочетаний минеральных удобрений
на накопление фосфора в семенах пшеницы.
Это дополнительно свидетельствует о том, что начальные этапы роста и развития пшеницы не
имеют непосредственной связи с накоплением запасных и минеральных веществ в семенах пшеницы,
а главным является процесс дегидратации вегетативной ткани и перенос из нее в семена продуктов
распада органического вещества и минеральных соединений. Широко распространенное мнение, что
семена с повышенным содержанием фосфора обладают высокими УК, в нашем исследовании не
подтвердилось, наоборот, доказано, что такие семена обладают пониженными УК. Это согласуется с
данными, что семена, репродуцированные при дефиците влаги в почве, имеют более высокую массу
1000 семян [9], обладают высоким содержанием белка и пониженными УК [5,6].
Выводы
Результаты данного исследования показывают, что из агроэкологических факторов репродукции
семян наибольшее значение имеет уровень влажности почвы. Семена, выращенные в условиях дефи-
цита почвенной влаги, имеют самое высокое содержание фосфора и пониженные урожайные качества.
Содержание фосфора в семенах не может служить критерием для прогнозирования их урожайных
качеств.
Минеральные удобрения имеют косвенное влияние на содержание фосфора и вовсе не влияют на
урожайные качества полученных семян. Между уровнем фосфора в почве и накоплением его в семе-
нах имеется слабая положительная связь при оптимальных и избыточных условиях влажности почвы.
Наилучшими урожайными качествами обладают семена, репродуцированные при оптимальной
влажности почвы.
Полученные результаты интерпретируются исходя из придания важности процессам распада веге-
тативной ткани на заключительной стадии жизни растения, переноса продуктов гидролиза органи-
ческих веществ вегетативной ткани в семена, что и является главным процессом, определяющим
накопления веществ в семенах пшеницы и ее продуктивность.
Литература:
1. БАБИЦКИЙ, А. Фотофосфорилирование на хлоропластах кукурузы. В: Труды Всесоюзного Института
растениеводства. Ленинград. 1969. Том 40. Приложение: Методы комплексного изучения фотосинтеза,
с.142-154.
2. БАБИЦКИЙ, А. Измерение фосфора методом дифференциальной колориметрии для анализа различий про-
цесса фотофосфорилирования у инбредных линий и гибридов кукурузы. В: Труды ВНИИООБ, Овощные и
бахчевые культуры. Астрахань, 1970, вып.1, с.159-178.
3. БАБИЦКИЙ, А. и др. Квазидискретные особенности накопления биомассы зерен в колосе пшеницы. В:
Известия Волгоградского государственного педагогического университета. Серия: Естественные и физико-
математические науки. Волгоград, 2004, №4(9), с.23-29.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
133
4. БАБИЦКИЙ, А. и др. Дискретные взаимодействия зерен в колосе пшеницы. В: Fiziologia şi biochimia plante-
lor de cultură (Aspecte Ecologice). Chişinău, 2004, p.18-21.
5. БАБИЦКИЙ, А. и др. Повышение урожайных качеств семян пшеницы. В: Аграрная наука. Москва, 2006,
№ 9, с.5-7.
6. БАБИЦКИЙ, А. Режим влажности почвы при выращивании пшеницы модифицирует продуктивные качества
ее семян. В: Культурные растения для устойчивого сельского хозяйства в 21 веке (Иммунитет, селекция,
интродукция). Москва, 2011, том 4, часть 1, с.373-380.
7. БАБИЦКИЙ, А. Главный фактор, определяющий содержание белка и его качество в зерне твердой пшеницы.
В: Studia Universitatis. Universitatea de Stat din Moldova. Seria: Ştiinţe reale şi ale naturii. Chişinău, 2011, nr.1
(41), p.92-96.
8. БАБИЦКИЙ, А. Белок зерна пшеницы – от чего зависит и чем является глиадин. В: Рациональное использо-
вание ресурсного потенциала регионов России и сопредельных государств. Сборник научных статей / Ред.
А.А. Афонин. Брянск: Курсив, 2011, с.23-30.
9. БАБИЦКИЙ, А. От чего зависит масса 1000 семян пшеницы. В: Impactul realizărilor ştiinţifice asupra pro-
ducţiei şi calităţii cerealelor spicoase: Materialele conferenţei internaţionale. Republica Moldova, Bălţi, 21 iunie
2013. Chişinău, 2013, p.15-21.
10. ВОЛЛЕЙД, Л. Фосфорный обмен в органах озимой пшеницы в зависимости от уровня азотно-фосфорного
питания. В: Сельскохозяйственная биология, 1973, том 7, № 4, с.548-553.
11. ЗАДОНЦЕВ, А. и др. К вопросу об использовании физиолого-биохимических показателей для прогнозиро-
вания урожайных свойств семян. В: Сельскохозяйственная биология, 1972, том 7, № 5, с.653-658.
12. КОЛЕСНИКОВ, С. О роли процессов неполного распада живого в размножении растений, гетерозиса и
других проявлениях жизнедеятельности организмов. В: Биология оплодотворения и гетерозис культурных
растении. Кишинѐв: Штиинца, 1965, вып. 3, с.123-201.
13. КОЛЕСНИКОВ, С. Итоги цитоэмбриологического изучения возделываемых растений в Молдавии. В:
Известия АН Молдавской ССР. Серия биологических и химических наук, 1967, № 11, с.44-50.
14. СЕЧНЯК, Л. и др. Экология семян пшеницы. Москва: Колос, 1981. 349 с.
15. СОБОЛЕВ, А. Образование и накопление фитина в семенах. В: Физиология растений, том 11, № 1, с.106-111.
16. СТРОНА, И. и др. Посевные и урожайные свойства семян пшеницы Харьковская 46 на разных фонах. В:
Селекция и семеноводство, 1974, № 3, с.50-52.
17. СТРОНА, И. К вопросу о разнокачественности семян и методах ее оценки. В: Труды Украинского НИИ
растениеводства, селекции и генетики им. В.Я. Юрьева. Харьков, 1962, том 7, с.69-76.
18. СТРОНА, И. Общее семеноведение полевых культур. Москва: Колос, 1966. 464 с.
19. PLAUT, Z. e.a. Transport of dry matter into developing wheat kernels and its contribution to grain yield under post –
anthesis water deficit and evaluated temperature. In: Field Crop Res., 2004, vol. 86, p.185-198.
20. SIMONOVA – STOILOVA, L. e.a. Proteolitic activity in wheat leaves during drought stress and recovery. In: Gen.
Appl. Plant Physiology, 2006. Special Issue. Vol. 32. Nо.1-2, p.91-100.
21. DEMIREVSKA, K. e.a. Drought – induced leaf protein alterations in sensitive and tolerant wheat varieties. In: Gen.
Appl. Plant Physiology, 2008, Special Issue. Vol.34. Nо.1-2, p.79-102.
22. ZAGRANSKA, B. e.a. Endoproteinase activities in wheat leaves upon water deficit. In: Acta Biochim. Polon.,
1996. Vol. 43. Nо.3, p.515-520.
23. WINIEWSKI, K. e.a. Genotype-dependent proteolytic response of spring wheat to water deficiency. In: J. Exp.
Bot., 2001. Vol. 52. Nо.360, p.1455-1463.
Prezentat la 29.03.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
134
CONSIDERAŢII CONCEPTUALE ŞI METODE GEOGRAFICE
DE ANALIZĂ A FENOMENULUI TURISTIC
Vitalie SOCHIRCĂ, Corneliu IAŢU*, Alexandru-Iulian CIOBANU
*
Universitatea de Stat din Moldova *Departamentul de Geografie, Universitatea „Al.I. Cuza” din Iaşi, România
Turismul reprezintă o activitate frecventă a omului modern, cu implicaţii pe multiple planuri (natural, economic, so-
cial, cultural, politic etc.). Numeroşi cercetători şi-au pus problema conceptualizării fenomenului turistic şi găsirii unor
parametri de măsurare a gradului de dezvoltare turistică. În relaţia dintre activităţile turistice şi mediul înconjurător, în
timp şi spaţiu, se produc modificări asupra componentelor mediului prin intermediul infrastructurii şi circulaţiei turistice.
Cuvinte-cheie: potenţial turistic, resurse turistice, agrement, produs turistic, piaţă turistică, amenajare turistică.
CONCEPTUAL CONSIDERATIONS AND GEOGRAPHIC ANALYSIS METHODS
OF THE TOURISM PHENOMENON
Tourism is a frequent activity of modern man, with implications on multiple levels (natural, economic, social, cultural,
political etc.). Many researchers have raised the issue of conceptualization of the touristic phenomenon and finding the
right parameters for measuring the level of tourism development. In the relationship between tourism activities and
environment, in time and space, changes occur in the environmental components through infrastructure and tourist traffic.
Keywords: tourism potential, tourism resources, recreation, tourism product, tourism market, tourism planning.
Introducere
Turismul este considerat astăzi, prin conţinutul, amploarea fenomenului, formele de manifestare şi rolul
său, un domeniu distinct de activitate, o componentă de primă importanţă a vieţii economice şi sociale. Re-
ceptiv la prefacerile civilizaţiei contemporane, fenomenul turistic evoluează sub impactul acestora, dinamica
sa integrându-se procesului general de dezvoltare. Mulţi specialişti consideră turismul un fenomen specific
epocii contemporane, dar primele încercări de definire şi caracterizare a acestuia datează încă din secolul al
XIX-lea.
Rezultate şi discuţii
1. Turismul – o abordare sintetică
Marea diversitate de interpretări date conceptului de turism poate fi pusă pe seama faptului că acest termen
încearcă să definească o multitudine de aspecte economice şi psihosociale, dar şi datorită faptului că turismul
reprezintă domeniul de studiu al mai multor discipline ştiinţifice (geografia, economia, sociologia, psihologia,
antropologia etc.).
William F. Theobald afirma că, din punct de vedere etimologic, cuvântul tour derivă din limba latină
(tornare) şi din limba greacă (τορνος), având semnificaţia de cerc – mişcarea în jurul unui punct central sau
o axă [15]. Preluat în limba engleză, cuvântul tour a căpătat semnificaţia acţiunii de a se mişca în cerc. În
consecinţă, un tour reprezintă o călătorie dus-întors şi cel care întreprinde o astfel de călătorie poartă numele
de tourist*.
Preluat în limba franceză, termenul tourisme a fost utilizat pentru prima dată în lucrarea Voyage d’un
français en Angleterre a lui G. Simond, fiind introdus cu rapiditate în aproape toate limbile lumii, extin-
zându-se, ulterior, asupra unei game variate de activităţi economice desfăşurate cu scopul destinderii şi al
* În Franţa, încă din anul 1623, o ordonanţă regală reglementa statutul staţiunilor balneare. După anul 1672, germanii, polonezii, dane-
zii, olandezii şi englezii aveau la îndemână un ghid fidel, unde erau oferite rudimente de limbă, descrieri ale drumurilor care condu-
ceau spre oraşele sau locurile cele mai faimoase, în mod deosebit Parisul. Erau propuse două circuite: micul tur (Parisul şi sud-vestul
Franţei) şi marele tur (sudul, sud-estul Franţei şi Burgundia). De asemenea, „marele tur” desemna călătoria iniţiatică realizată de
către tinerii aristocraţi, însoţiţi de perceptori. Acest voiaj marca trecerea de la perioada studiilor secundare la maturitate, necesară
unei eventuale cariere diplomatice şi avea ca scop contactul cu locuri evocatoare ale originilor civilizaţiei europene: Roma, Atena,
Constantinopol (durata călătoriei era de la 6 luni până la 2 ani, fiecare dintre etape presupunând o staţionare de cel puţin câteva luni).
În Marea Britanie, pe la 1800, termenul turist era utilizat pentru a desemna pe cei care făceau marele tur, iar pe la 1811 termenul
turism desemna teoria şi practica acestor „călătorii de plăcere” (în epocă, motivaţia principală era agrementul) (cf. [11, p.20-22])

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
135
evadării din cotidian, în condiţiile creşterii continue a standardului de viaţă şi ale accentuării stresului la care
organismul uman este supus în contextul civilizaţiei industriale [7, p.9].
O definiţie a turismului a fost elaborată de către Walter Hunziker: „Turismul este ansamblul de relaţii şi
fenomene care rezultă din deplasarea şi sejurul persoanelor în afara domiciliului lor, atâta timp cât sejurul şi
deplasarea nu sunt motivate printr-o stabilire permanentă şi o activitate lucrativă oarecare” [5].
Alţi specialişti de talie mondială care şi-au adus contribuţia la clarificarea conceptului de turism sunt
K.Krapf, J.Krippendorf, R.Baretje, F.W. Ogilvie, A.J. Norval.
Dictionnaire Touristique International (1969) [17] defineşte turismul ca fiind „ansamblul de măsuri puse
în aplicare pentru organizarea şi desfăşurarea unor călătorii de agrement sau în alte scopuri, realizate fie prin
intermediul unor organizaţii, societăţi sau agenţii specializate, fie pe cont propriu, pe o durată limitată de timp,
precum şi industria care concură la satisfacerea nevoilor turiştilor”. În Dicţionarul Turistic Internaţional
(1980) [18] se precizează că „turismul se distinge de călătorie prin aceea că implică pentru persoana în cauză,
pe de o parte, alegerea deliberată a ţintei, pe de alta, preocuparea exclusivă pentru satisfacerea plăcerii sale”.
Piere George a încercat să impună ideea că omul s-a născut turist (o abordare inacceptabilă pentru econo-
mişti, adepţi ai teoriei conform căreia omul devine turist). În accepţiunea sa, turismul reprezintă „activitatea
legată de recreere, care desemnează deplasările sezoniere ale populaţiei urbane spre regiuni favorizate de
resurse naturale şi care răspund prin dotări aşteptărilor turiştilor” [3].
Iniţial, conceptul de turism era utilizat pentru a desemna o călătorie de plăcere, de recreere, de comuni-
care interculturală sau în scopul îngrijirii sănătăţii. O definiţie a conceptului de turist a fost formulată şi de
către Organizaţia Mondială a Turismului*, în 1978: „… orice persoană care se află în afara reşedinţei sale
curente pentru o durată de cel puţin 24 de ore (sau o noapte) şi pentru maximum patru luni din motivele
următoare: agrement (vacanţe şi week-end-uri), sănătate (termalism, talasoterapie etc.), misiuni sau reuniuni
de orice fel (congrese, seminarii, pelerinaje, manifestări sportive etc.), călătorii de afaceri, deplasări profe-
sionale, călătorii şcolare etc.” [4, p.6-7]. În anul 1993, UNWTO a adoptat o nouă definiţie pentru termenul
turist, care „încearcă să ţină cont de practicile turistice efective şi de necesitatea observaţiei, cunoaşterii feno-
menului turistic” [11, p.15-16]. Conform acestei definiţii, turiştii sunt persoanele care „călătoresc sau locuiesc
în locuri din afara zonei lor de reşedinţă permanentă pentru o durată de minimum douăzeci şi patru (24) de
ore, dar nu mai lungă de un an consecutiv, în scop de recreere, afaceri sau altele nelegate de exercitarea
unei activităţi remunerate în localitatea vizitată” [4, p.8].
Pascal Cuvelier definea turismul ca fiind „un ansamblu de practici asociate abandonării temporare a
reşedinţei, având drept obiectiv destinderea sau motivaţii cu caracter sociocultural” [2]. Spre deosebire de
definiţia formulată de UNWTO, această definiţie exclude călătoriile de afaceri.
În definirea conceptului de turism intervin trei componente majore, care argumentează caracterul interdis-
ciplinar al turismului: locul turistic, timpul consumat şi activităţile specifice [9, p.57-70]. Din perspectivă
geografică, termenul „turism” presupune interrelaţionarea a patru elemente esenţiale: turiştii, populaţia locală,
fluxurile şi teritoriile astfel marcate [8].
O altă definiţie a turismului a fost formulată de către Douglas Pearce: turismul reprezintă „un ansamblu
de relaţii şi fenomene rezultate în urma călătoriilor şi sejurului temporar al persoanelor, în special pentru a
se destinde sau recrea” [13]. Această definiţie delimitează net două componente esenţiale ale turismului:
călătoria şi sejurul, care se deosebesc prin durata deplasării. De asemenea, definiţia surprinde interrelaţiona-
rea care se creează ca urmare a acestor deplasări, altfel spus structurile spaţiale, formele culturale, schimburile
(sociale, culturale, economice).
Marc Boyer definea turismul ca fiind „ansamblul fenomenelor generate de călătorie şi de şederea tempo-
rară a oamenilor în afara propriului domiciliu, în timp ce aceste deplasări satisfac, în timpul liber, nevoile
* Organizaţia Mondială a Turismului (UNWTO) este o organizaţie internaţională guvernamentală fondată în anul 1970, activă din
anul 1975 şi integrată în Organizaţia Naţiunilor Unite în anul 1977. UNWTO are sediul la Madrid, România devenind membru în
anul 1974, iar Republica Moldova – în anul 2002. UNWTO s-a format pe structura Uniunii Internaţionale a Organismelor Oficiale de
Turism (fondată în anul 1925) şi are ca principale atribuţii colectarea şi relaţionarea informaţiilor statistice privind turismul interna-
ţional. UNWTO reprezintă organismele turistice din sectorul public, din numeroase ţări din întreaga lume, şi publică statistici cu
privire la fenomenul turistic la nivel global, regional sau naţional. Promovând dezvoltarea de fond, durabilă şi universal accesibilă a
turismului, UNWTO încurajează punerea în aplicare a Codului de etică globală pentru Turism, cu scopul de a asigura maximizarea
efectelor pozitive ale turismului în plan economic, social şi cultural şi minimizarea impactului negativ al turismului asupra mediului
(cf. http://www2.unwto.org/, accesat la 20.11.2013).

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
136
culturale ale civilizaţiei industriale” [1]. Această definiţie pune accentul pe nevoia culturală, ceea ce pre-
supune o construcţie intelectuală progresivă sau o consecinţă a educaţiei. Astfel, turismul este definit ca un
fenomen legat de mijloacele tehnice ale civilizaţiei industriale, precum şi ale civilizaţiei care îi succede
(civilizaţia digitală, post-industrială).
Turismul reprezintă deplasarea temporară, pe termen scurt, a persoanelor în afara locului de reşedinţă sau
lucru, ceea ce poate implica necesitatea unei înnoptări într-o structură de primire turistică şi, eventual, rezer-
varea biletului de călătorie. „Turismul este o activitate umană, fondată pe deplasarea în spaţiu a indivizilor.
În aceasta constă diferenţa faţă de recreere (loisir în franceză) care nu necesită neapărat deplasarea. «A fi
turist» şi «a practica turismul» este echivalent cu abandonarea temporară a spaţiului cotidian de viaţă în
favoarea altui loc, construit pentru şi de către turişti, consacrat exclusiv odihnei. Turismul întreţine astfel
raporturi privilegiate cu spaţiul (…)” [11, p.12].
În definirea conceptului de turism, Philippe Duhamel insistă asupra turismului ca rezultat al urbanizării:
„Turismul este în mod fundamental un proiect urban. El este susţinut de către societăţile urbane, care proiec-
tează asupra spaţiului reprezentările şi modul lor de viaţă. Aceasta induce transformări de tip urban, cum ar
fi o sociabilitate specific urbană, o organizare a spaţiului marcată de simboluri urbane, concentraţii spaţiale
demne de un oraş. Pe de altă parte, turismul este un proiect de societate care îşi însuşeşte spaţii şi le transformă
în funcţie de normele şi valorile proprii şi care transcend categoriile spaţiale admise” [16].
De asemenea, Stéphane Nahrath şi Mathis Stock insistă asupra caracterului urban al turismului, precum şi
asupra consecinţelor intensificării prin turism a urbanizării: „Turismul, invenţie a mediului urban, cristalizează
valorile şi practicile urbane, precum şi formele arhitecturale, şi transferă această urbanitate în toate locurile
incluse în turism, chiar şi în cele care sunt considerate a priori cele mai îndepărtate de modelul urban, cum
sunt staţiunile sau satele turistice, de exemplu la munte sau pe litoral. Într-adevăr, aceste zone devin urbani-
zate, ca urmare a acestui transfer de urbanitate şi de centralitate, şi văd apariţia problemelor legate de planifi-
carea urbană, de trafic şi de redistribuirea capitalului economic, de întâlnirea unor populaţii eterogene, de
gestionarea urbană, de poluare, de violenţă etc. În plus, facilităţile urbane sunt o condiţie sine qua non pentru
ca turismul să se dezvolte acolo” [12, p.7-14].
În literatura de specialitate în limba rusă turismul este definit ca „activitate de călătorie în timpul liber, o
formă de odihnă activă, în diverse scopuri” [19]. Se accentuează, de asemenea, faptul că „turiştii nu desfă-
şoară activităţi remunerate în cadrul călătoriei şi sejurului” [20].
Actualmente, într-un context economic şi social complex şi diversificat, conceptul de turism desemnează,
de asemenea, ansamblul activităţilor economice la care apelează turiştii în timpul sejurului la locul de desti-
naţie (transport, hoteluri, restaurante, baruri, locuri de agrement etc.).
„Turismul este o componentă activă a vieţii social-economice contemporane, care marchează profund
regiunile favorizate de un potenţial natural şi antropic de excepţie, prin atracţia exercitată asupra maselor
largi, în contextul creşterii nivelului de trai şi necesităţii atenuării influenţelor nocive ale mediului urban-
industrial modern” [11, p.15].
2. Concepte de bază în turism
Principalele componente ale turismului sub raport economic sunt: resursele turistice, resursele de muncă
implicate, infrastructura turistică, mijloacele şi echipamentele de producţie, serviciile turistice, bunurile des-
tinate consumului turistic şi structurile organizaţionale turistice.
„Materia primă a turismului este spaţiul şi, fără a subestima alţi factori, condiţiile geografice deţin un rol
de prim-plan, esenţial adesea în atracţia unui anumit loc turistic. Cadrul natural, patrimoniul cultural şi isto-
ric, potenţialul tehnic şi economic sunt tot atâtea criterii geografice care intervin în localizarea turistică, revi-
zuirea lor fiind obligatorie în orice demers geografic. (…) Noţiunea materie primă turistică se confundă, cel
mai adesea, cu potenţialul turistic, resursele turistice sau cu oferta turistică” [11, p.42]. De fapt, există dife-
renţe între resursele turistice potenţiale, amenajabile, amenajate şi resursele utilizate (valorificate), în funcţie
de condiţiile naturale, social-economice, precum şi de posibilităţile tehnice ale societăţii [11, p.44].
Potenţialul turistic al unui teritoriu reprezintă totalitatea elementelor naturale sau antropice care pot con-
stitui obiectul atracţiei turistice. „Potenţialul turistic este o sumă a unor condiţii obiective, naturale sau so-
ciale, dar şi subiective, ce ţin de motivaţii şi necesităţi, aflate într-o continuă dinamică” [11, p.44]. Poten-
ţialul turistic are un caracter dinamic: poate căpăta noi valenţe, se poate extinde, îmbogăţi cu noi surse de
atracţie (generate fie de evoluţia tehnicii şi civilizaţiei, fie de schimbări ce pot interveni în mediul natural)

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
137
sau, dimpotrivă, poate fi diminuat, modificat sau chiar distrus, adesea ca urmare a intervenţiei factorilor
antropici [14, p.6]. „Oricât de bogat ar fi potenţialul turistic, valorificarea sa depinde, în ultimă instanţă, de
distribuţia spaţială a resurselor financiare şi a centrelor de putere care dictează ce zone, regiuni sau centre
merită să fie sprijinite în valorificarea acestuia” [11, p.52].
Resursele turistice reprezintă totalitatea componentelor, proceselor şi fenomenelor naturale şi antropice
ale unui teritoriu, care pot fi valorificate în turism şi care constituie premisele dezvoltării acestuia. Aceste
resurse presupun existenţa unor arii cu atracţii turistice şi depind cu precădere de posibilităţile tehnice ale
societăţii [11, p.42].
Din punct de vedere geografic, noţiunea de ofertă turistică face referire la potenţialul turistic amenajat,
transformat în produs turistic. Acest concept face distincţia dintre aspectele turistice legate de cei care deser-
vesc domeniul şi cele care se referă la consumatorii acestora [11, p.43]. Din perspectivă economică, oferta
turistică reprezintă totalitatea produselor şi serviciilor puse la dispoziţia turiştilor. Din perspectiva agenţiilor
de turism, oferta turistică reprezintă un pachet de servicii care este propus unui anumit client, de regulă, la un
preţ competitiv. În concluzie, oferta turistică reprezintă ansamblul elementelor care motivează deplasarea în
scop turistic: resursele turistice naturale şi antropice, echipamentele turistice, bunurile şi serviciile destinate
consumului turistic, infrastructura turistică, forţa de muncă implicată, condiţiile de comercializare (preţuri,
facilităţi etc.). Pentru a efectua o deplasare în scop turistic, persoanele apelează adesea la serviciile unor agenţii (transpor-
tatori, tur-operatori, lanţuri hoteliere, agenţii de voiaj, societăţi de alimentaţie publică şi de animaţie ş.a.). Astfel încât, pentru a răspunde exigenţelor turiştilor, este necesară o infrastructură complexă, diversificată, alcătuită din instituţii publice şi societăţi private, care să organizeze în mod eficient piaţa turistică. „Piaţa turistică poate fi definită ca fiind ansamblul tranzacţiilor (actelor de vânzare-cumpărare) al căror obiect îl constituie produsele turistice, privite în conexiune cu relaţiile pe care le generează şi cu spaţiul geografic în care se desfăşoară. (…) Piaţa turistică reprezintă sfera economică de interferenţă a ofertei turistice, materiali-zată prin producţia turistică, cu cererea turistică, materializată prin consumul turistic” [10, p.21].
Produsele turistice oferite pentru consumul turiştilor pe parcursul unui sejur pot fi definite ca „un ansamblu de amenajări, imagini şi simboluri supuse unei valorificări specifice printr-un marketing care urmăreşte atât scopuri materiale (rentabilitatea înaltă a investiţiilor), cât şi spirituale (asigurarea unei ambianţe capabile să asigure turistului maximum de confort)” [11, p.116].
Materia primă turistică reprezintă totalitatea resurselor turistice naturale şi antropice ce constituie baza
ofertei turistice potenţiale a unui teritoriu considerat destinaţie turistică. Aceasta este constituită din resursele
turistice (elementele de bază ale potenţialului turistic), obiectivele turistice (componente ale patrimoniului
natural sau antropic) şi atracţia turistică (latura afectivă a diferitelor elemente ale potenţialului turistic, ceea
ce se impune atenţiei, latura estetică şi cognitivă). Aceste aspecte stau la baza dezvoltării turismului, deter-
minând mărimea, intensitatea şi diversitatea fluxurilor turistice [14, p.5].
Amenajarea turistică este un proces dinamic şi complex de organizare a spaţiului turistic (proiectare a
unor noi dotări turistice, recondiţionare a dotărilor existente, redimensionare a acestora în funcţie de cererea
turistică) şi de rentabilizare economică a activităţii turistice, luând în considerare relaţiile dintre mediul ambiant
şi comunităţile umane, precum şi factorii care influenţează aceste relaţii.
„Relieful constituie suportul esenţial în amenajările turistice, fiind şi un element de bază în crearea deco-
rului specific fiecărui sit turistic. (…) Relieful prezintă avantajul că atracţiile pe care le generează nu necesită
amenajări costisitoare şi, de regulă, nu sunt periclitate de frecvenţa turistică excesivă, acestea fiind doar admi-
rate, contemplate şi nu efectiv consumate” [11, p.55].
Ameliorarea şi diversificarea mijloacelor de transport influenţează amenajarea spaţiilor turistice auxiliare:
campinguri, parcări, gări, aeroporturi, porturi ş.a. (acestea trebuie să-şi adapteze structurile şi serviciile în
funcţie de exigenţele turistice).
Staţiunea turistică este o localitate şi/sau areal care prezintă un ansamblu de elemente de atracţie
turistică şi care este dotată cu mijloace de cazare, alimentaţie şi agrement, precum şi cu alte facilităţi turistice
puse în serviciul turiştilor pentru o anumită perioadă de timp*.
* De exemplu, staţiunea balneară este localitatea (şi arealul) situată pe ţărmul mării (sau, în unele cazuri, al unui lac), ce dispune de
substanţe minerale eficiente la nivel terapeutic, instalaţii specifice pentru băi curative şi tratament şi are o organizare care permite
acordarea asistenţei medicale specifice.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
138
În literatura de specialitate, staţiunea turistică este definită ca fiind un spaţiu caracterizat de predominanţa
activităţilor turistice şi de agrement (monofuncţionalitate) – creatoare ex nihilo a acestui loc, de caracterul
sezonier al respectivelor activităţi, de o capacitate de cazare a populaţiei non-permanente rezultante, precum
şi de prezenţa unei populaţii permanente. De asemenea, o staţiune turistică trebuie să dispună de o resursă
turistică (litoral, munte, oraş, sat etc.) şi să fie accesibilă (să fie deservită de o reţea de transport rutier, fero-
viar, aerian, maritim). Staţiunea turistică ar trebui să includă structuri de cazare (hoteluri, locuri de campare
etc.), magazine, restaurante şi facilităţi turistice (de exemplu, pârtii de schi în staţiunile de sporturi de iarnă)
[6, p.193-204].
Principalele aspecte care individualizează o staţiune turistică sunt: existenţa unei infrastructuri generale,
precum şi a uneia specific turistice; dotări, utilităţi, instalaţii, echipamente, facilităţi; servicii şi produse spe-
cific turistice; ospitalitate; accesibilitate; calitatea serviciilor; posibilitatea practicării unor activităţi turistice;
amenajări turistice.
Structurile turistice de primire* sunt reprezentate de orice construcţie sau amenajare care furnizează
permanent sau sezonier serviciul de cazare şi alte servicii specifice pentru turişti (servirea mesei, agrementul,
tratamentul balnear, transportul destinat turiştilor etc.).
Infrastructura turistică este reprezentată de: instalaţiile, dotările şi echipamentele specifice activităţii
de agrement (pârtii de schi, patinoare, piscine, plaje amenajate, porturi turistice etc.); transporturile turistice
(linii de transport montan pe cablu, trenuleţe, vase de croazieră etc.); unităţile de cazare şi alimentaţie destinate
turiştilor; reţeaua unităţilor de agrement (parcuri de distracţie, terenuri de sport etc.) şi de tratament (centre
balneare şi balneoclimaterice).
3. Metodologia evaluării potenţialului turistic
a. Obiectivele evaluării potenţialului turistic
Evaluarea potenţialului turistic al unui teritoriu are ca obiective: cunoaşterea distribuţiei spaţiale a resur-
selor turistice într-un anumit areal; construirea unui instrument de lucru pentru analişti sau pentru orice grup
de acţiune locală; determinarea teritoriului/localităţii în care ar putea fi derulat, în perspectivă, un eventual
proiect de dezvoltare turistică (public sau privat); identificarea unor strategii prin care să se realizeze coope-
rare şi dialog între populaţie, actorii locali şi experţi; formarea unor mentalităţi deschise către turism şi con-
ştientizarea rolului turismului în calitate de motor de dezvoltare locală pe termen mediu şi lung.
b. Evaluarea şi diagnosticarea Etapa de evaluare şi diagnosticare a situaţiei turistice existente presupune: examinarea ofertei teritoriului,
a cererii turistice, a concurenţei teritoriale şi a tendinţelor pieţei, precum şi identificarea unor indicatori eco-
nomici (cifra de afaceri din turism, taxele obţinute de administraţie din activităţile turistice, numărul de locuri
de muncă din turism etc.). Realizarea acestor aspecte presupune colectarea de informaţii şi date în teren, pre-
lucrarea lor, dar şi utilizarea unor informaţii interne şi externe acelui teritoriu.
Confruntarea rezultatelor oferite de analiza situaţiei turistice permite identificarea punctelor tari şi a celor
slabe ale unui teritoriu, precum şi determinarea avantajelor şi riscurilor/ameninţărilor. Definirea elementelor
de unicitate sau de diferenţiere ale teritoriului respectiv oferă posibilitatea construirii unei „strategii turistice
de succes”.
c. Analiza ofertei turistice
Factorii care influenţează oferta turistică sunt: naturali; antropici; socioeconomici; culturali; social-politici;
infrastructura generală (de transport şi telecomunicaţii, tehnico-edilitară, sanitară etc.); serviciile disponibile
în acel teritoriu; infrastructura specific turistică (structurile de primire cu funcţii de cazare, posibilităţile de
servire a mesei, posibilităţile de a desfăşura activităţi sportive şi de petrecere a timpului liber, posibilităţile de
a organiza seminarii şi conferinţe, precum şi bazele de cură şi tratament).
Principalele elemente ale ofertei turistice sunt: populaţia locală, organizaţiile turistice locale, comerţul
turistic, gradul de calificare în turism şi cooperarea între diferiţii actori implicaţi în gestionarea activităţilor
turistice.
* Exemple de structuri turistice de primire: vilă, bungalou, pensiune, căsuţă, camping, hotel, motel, tabără etc. Sunt excluse: structurile
de cazare folosite în exclusivitate de către proprietari/chiriaşi mai mult de un an, locuinţele secundare, căminele studenţeşti, interna-
tele, unităţile spitaliceşti (cu excepţia celor unde se desfăşoară activităţi turistice), vagoanele-dormitor, adăposturile şi refugiile mon-
tane, barăcile şi dormitoarele pentru muncitori, căminele de bătrâni, casele de copii.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
139
În analiza ofertei turistice, aspectele legate de populaţia locală se referă la gradul de informare a acesteia,
exigenţele, modul în care aceasta poate contribui la dezvoltarea sectorului turistic, contribuţia ei la amenajarea
peisajelor, perpetuarea unor valori etnoculturale, prezenţa unor lideri de opinie, dinamica demografică etc. Organizaţiile turistice locale sunt reprezentate de: birouri de informare turistică, salvamont, angajaţi din
cadrul administraţiei cu probleme de turism, organizaţii non-profit, asociaţii, agenţii de turism detailiste sau tur-operatoare. Examinarea acestor elemente vizează: analiza activităţilor turistice desfăşurate, a competenţe-lor persoanelor implicate, a bugetului alocat, a disponibilităţii de cooperare, precum şi existenţa unor activităţi de promovare turistică (în curs de desfăşurare sau în perspectivă).
Noţiunea de comerţ turistic face referire la: politica de ofertă-preţ, canalele de distribuţie a produselor turis-tice, gradul de promovare a turismului, instrumentele de promovare turistică, strategiile de comercializare etc.
În analiza ofertei turistice, aspectele legate de formarea în turism vizează: calificări, traininguri, gradul de calificare a angajaţilor din turism, programe de formare turistică etc.
Cooperarea în turism presupune: sprijin din partea autorităţilor publice şi private, proiecte comune în curs de iniţiere sau în desfăşurare, parteneriat potenţial pentru realizarea de proiecte, cooperarea între operatori turistici locali, elaborarea unor strategii comune, reprezentarea lor în faţa factorilor de decizie publici etc.
d. Cererea turistică Cererea turistică este un concept economic ce se referă la cantitatea de produse turistice (transport, cazare,
alimentaţie, tratament, divertisment, vizitarea unor obiective turistice, suveniruri, produse ale agenţiilor de turism, utilizarea instalaţiilor de agrement etc.) pe care turiştii, existenţi sau potenţiali, sunt dispuşi să le con-sume la un anumit preţ, la un moment dat şi în anumite împrejurări
*.
Noţiunea de cerere turistică se mai referă şi la numărul de turişti, cu caracteristicile acestora (număr, vârstă,
sex, ocupaţie, venituri, buget, preferinţe, experienţă turistică, mijloc de deplasare ales etc.). În acest sens, se
poate diferenţia noţiunea de cerere efectivă (numărul de persoane care participă efectiv la activităţile turistice)
de noţiunea de cerere potenţială (numărul de persoane care au dorinţa de a călători, însă nu au posibilitatea,
din anumite cauze). Atunci când factorul restrictiv dispare, cererea potenţială poate deveni cerere efectivă.
e. Modele de analiză spaţială a fenomenului turistic
Modelele turistice propun o bază teoretică şi conceptuală pentru studiul dinamicii spaţiale a turismului.
Majoritatea modelelor de analiză a fenomenului turistic se fundamentează pe structura origine-traseu-destinaţie:
modelele de fluxuri turistice (au în centru călătoria sau itinerariul), modelele de tipul origine-destinaţie (pun
accentul pe relaţia dintre spaţiul de emisie şi spaţiul de recepţie), modelele structurale şi modelele evolutive.
f. Metode de analiză a fenomenului turistic
Analiza cantitativă (studiu de statistică) furnizează informaţii referitoare la: câţi turişti frecventează teri-
toriul în cauză; câte/ce produse turistice au consumat; la câte/ce servicii au apelat; cât timp/unde au înnoptat;
cât au cheltuit (în medie, maximum, minimum) etc. Indicatorii cantitativi utilizaţi în realizarea analizelor turistice sunt: numărul de sosiri şi de nopţi petrecute
(înnoptări) de către turişti în localitatea respectivă; distribuţia înnoptărilor turistice în cadrul diferitelor struc-turi de cazare (motel, pensiune, camping, gazdă, hotel, tabără etc.); durata medie de şedere (raportul dintre numărul de înnoptări şi numărul de sosiri); rata de ocupare a structurilor turistice; bugetul alocat de către turişti; analiza evoluţiei indicatorilor pe mai multe luni/ani; distribuţia indicatorilor de-a lungul unui an, pentru a identifica intensitatea sezonieră a activităţilor turistice.
Analiza calitativă (studiu de teren) furnizează date referitoare la: ce tip de clienţi frecventează acel teritoriu/ localitate/structură turistică; provenienţa acestora; motivaţiile şi aşteptările lor; ce le-a plăcut/displăcut mai mult etc.
Indicatorii calitativi utilizaţi în realizarea analizelor turistice sunt: durata, scopul şi tipul călătoriei; mijlo-
cul de transport utilizat; tipul de cazare ales; activităţile realizate de către turişti; motivaţiile, aşteptările, gra-
dul de satisfacţie a turiştilor; reputaţia şi atractivitatea zonei; motivaţia alegerii acelui teritoriu; frecventarea
acelui loc (în trecut sau existenţa unor proiecte în perspectivă); date statistice calitative generale asupra turiş-
tilor (vârstă, sex, ocupaţie, educaţie, provenienţă geografică, venituri) etc.
g. Metode de colectare a informaţiilor şi a datelor statistice în turism
În ceea ce priveşte cererea turistică, metodele de colectare a informaţiilor şi a datelor statistice sunt: ches-
tionarea directă a turiştilor (oral sau în scris); chestionarea telefonică; prelucrarea formularelor de înregistrare
* Există o regulă economică: dacă preţul este mare, cererea pentru acel produs este mai mică, şi viceversa (cu anumite excepţii).

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
140
a turiştilor la hoteluri; identificarea originii turiştilor după numărul de înmatriculare al maşinilor; organizarea
de întâlniri, conferinţe, mese rotunde; realizarea de interviuri cu actori politici, publici sau privaţi; achiziţio-
narea de date de la organele de statistică ş.a.
Concluzii Turismul, ca activitate umană în continuă evoluţie, s-a format şi dezvoltat în interacţiune cu mediul natu-
ral, care a exercitat permanent o influenţă directă în dezvoltarea societăţii, în ansamblul său. Ca rezultat al acţiunii modelatoare asupra mediului ambiant a factorilor naturali, pe de o parte, şi a celor antropici, pe de altă parte, potenţialul turistic al unui teritoriu reprezintă o manifestare spaţial-funcţională a capacităţii habita-tului respectiv de a susţine activităţi specifice turismului.
Fenomen social-economic extrem de complex, ce implică multe variabile, turismul se dezvoltă sub o multitudine de forme şi tipuri. Delimitarea clară a conceptului de turism, a conţinutului fiecărei forme de turism prezintă importanţă pentru identificarea, pe de o parte, a comportamentului turiştilor în materie de consum şi cheltuieli şi, pe de altă parte, a responsabilităţilor şi obligaţiilor agenţiilor turistice (tur-operatori şi agenţii de voiaj) şi/sau prestatorilor de servicii (societăţi de transport, hoteluri, restaurante etc.).
Bibliografie:
1. BOYER, M. Le tourisme en France (vade-mecum). Editions EMS, 2003. ISBN (10): 2912647908,
ISBN (13): 978-2912647900
2. CUVELIER, P. Anciennes et nouvelles formes de tourisme: Une approche socio-économique. Paris: L Harmattan,
1998. ISBN (10): 2738469930, ISBN (13): 978-2738469939
3. GEORGE, P. Dictionnaire de la géographie. Paris: PUF, ediţia a doua, 1974. ISBN (10): 2130454925,
ISBN (13): 9782130454922
4. HOERNER, J-M. Géographie de l'industrie touristique. Paris: Ellipses, 1997. ISBN (10): 2729847871,
ISBN (13): 978-2729847876
5. HUNZIKER, W., KENS, A.G. Le Tourisme Social: caractères et problèmes [Social tourism: its nature and problems].
Publication de la Commission Scientifique de l'Alliance Internationale de Tourisme (în franceză). Geneva: Imprim.
Federative SA., 1951. OCLC 255533141. OL 21122881M
6. KNAFOU, R., BRUSTON, M., DEPREST, F., GAY, J-C., DUHAMEL, P., SACAREAU, I. Une approche géogra-
phique du tourisme, în L'Espace géographique. Paris: Belin- Reclus, 3/1997 (no.26). p.193-204. ISSN: 0046-2497,
ISSN (online): 1776-2936
7. LOZATO-GIOTART, J-P. Geographie du tourisme. Paris: Masson, 1991, apud: MUNTELE, I., IAŢU, C.
Geografia turismului: concepte, metode şi forme de manifestare spaţio-temporală. Iaşi: Sedcom Libris, 2006.
ISBN (10): 973-670-209-x, ISBN (13): 978-973-670-209-9
8. MARIÉ, M. Les terres et les mots. Une traverseé des sciences sociales. Paris: Méridiens-Klincksieck, 1989.
ISBN (10): 2865632482, ISBN (13): 978-2865632480
9. MITCHELL, L.S., MURPHY, P.E. Geography and tourism, în Annals of Tourism Research. Vol.18, Elsevier, 1991,
p.57-70. ISSN: 0160-7383
10. MORARIU, D., WEISZ, J. Economia turismului. Timişoara: Eurostampa, 2006. ISBN: 973-687-471-0
11. MUNTELE, I., IAŢU, C. Geografia turismului: concepte, metode şi forme de manifestare spaţio-temporală. Iaşi:
Sedcom Libris, 2006. ISBN (10): 973-670-209-x, ISBN (13): 978-973-670-209-9
12. NAHRATH, S., STOCK, M. Urbanité et tourisme: une relation à repenser, în Espaces et sociétés. Paris: Éditions
érès, 3/2012 (nr. 151), pp. 7-14. ISSN: 0014-0481, ISSN (online): 1961-8700, DOI: 10.3917/esp.151.0007. Cf.
http://www.espacesetsocietes.msh-paris.fr/blog/2013/02/11/editorial-urbanite-et-tourisme-une-relation-a-repenser/,
[accesat la 20.11.2013].
13. PEARCE, D. Geographie du tourisme. Paris: Nathan, 1993. ISBN (10): 2091902446, ISBN (13): 978-2091902449
14. SOCHIRCĂ, V., BEJAN, I. Resurse turistice. Chişinău: CEP USM, 2011. ISBN 978-9975-71-083-1
15. THEOBALD, F.W. Global Tourism: The Next Decade. Oxford: Butterworth-Heinemann, 1994.
ISBN (10): 0750615680, ISBN (13): 978-0750615686
16. VIOLIER, P., DUHAMEL, P. Tourisme et littoral, un enjeu du monde. Paris: Belin, 2009. ISBN (10): 2701153654,
ISBN (13): 978-2701153650
17. Dictionnaire Touristique International. Académie Internationale du Tourisme, Ed. Monte Carlo, ediţia a 3-a revizuită.
OCLC Number: 54123998
18. Dicţionar Turistic Internaţional. NEGUŢ, S. Bucureşti, Editura Sport-Turism, 1980.
19. Современная энциклопедия. 2000. Cf. http://dic.academic.ru/searchall.php?SWord=туризм [accesat la 21.11.2013].
20. http://www.triadatur.ru/termin.php [accesat la 21.11.2013].
Prezentat la 02.12.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
141
ASPECTE PRIVIND REPARTIŢIA DEPUNERILOR DE IONI MINERALI ÎN
ECOSISTEMELE FORESTIERE DIN REPUBLICA MOLDOVA ŞI DIN EUROPA
Valeriu BRAŞOVEANU
Institutul de Ecologie şi Geografie al AŞM
Este efectuată o analiză comparativă a depunerilor atmosferice sub coronamentul pădurilor din Republica Moldova
cu cele determinate în suprafeţele experimentale de monitoring forestier intensiv (nivelul II) din Europa. Monitorizarea
ionilor de S-SO42-
, N-NO3- şi N-NH4
+ a fost realizată în baza metodologiei recomandate de programul ICP Forests. S-a
constatat că în ecosistemele forestiere din nordul Republicii Moldova depunerile atmosferice ale ionilor monitorizaţi
sunt mai reduse faţă de depunerile înregistrate în ecosistemele forestiere din centrul ţării, acestea din urmă ameninţând
mai grav starea de sănătate a arborilor şi funcţionalitatea ecosistemelor forestiere. Depunerile ionilor minerali (S-SO42-
,
N-NO3-, N-NH4
+) înregistrate sub coronamentul arboretelor studiate în Republica Moldova sunt comparabile cu depu-
nerile înregistrate în regiunile din centrul Europei şi în cele central-estice ale ei.
Cuvinte-cheie: poluare atmosferică, monitoringul depunerilor, ecosistem forestier, ioni minerali.
ASPECTS OF REPARTITION THE ION MINERAL DEPOSITION
IN THE FOREST ECOSYSTEMS IN MOLDOVA AND EUROPE
It performed a comparative analysis of throughfall atmospheric deposition in the Republic of Moldova to that deter-
mined in the intensive (level II) forest monitoring plots in Europe. Monitoring ions S-SO42-
, N-NO3- şi N-NH4
+ was
carried out according to the methodology recommended by the International Co-operative Programme on Assessment
and Monitoring of Air Pollution Effects on Forests (ICP Forests). It was found that the atmospheric ions deposition mo-
nitored in forest ecosystems in northern the country are reduced than deposits recorded in forest ecosystems in the
center of the country, the latter being subject to increased risk of threat to the health of the trees and functionality forest
ecosystems. Throughfall deposits of mineral ions (S-SO42-
, N-NO3-, N-NH4
+) recorded in forests studied in Republic of
Moldova are comparable to deposits recorded in the central and central-eastern European regions.
Keywords: atmospheric pollution, deposition monitoring, forest ecosystem, mineral ions.
Introducere Depunerile atmosferice de sulf şi azot, care formează compuşi acidifianţi şi azot anorganic ca nutrienţi şi ca-
tioni de bază, constituie un factor major ce influenţează şi/sau afectează starea pădurilor din Europa. Astfel, acestea cuprind toate procesele ce conduc la depunerea unei anumite cantităţi de element conţinut în aerosoli,
ape de precipitaţii sau compuşi gazoşi pe o suprafaţă receptoare, care este, în cazul ecosistemului forestier, par-tea aeriană a arborilor (ramuri, frunze sau ace), vegetaţia ierboasă şi solul [8].
Poluarea atmosferică, ca factor de stres, în principal antropic, se caracterizează printr-o dinamică mult mai intensă decât procesele naturale adaptive ale pădurilor, deoarece arborii, ca organisme vii, prezintă o ca-
pacitate lentă de adaptare la schimbările condiţiilor de mediu [7]. În studiul de doctorat Draaijers (1993) [2] face o caracteristică a principalelor tipuri de poluanţi atmosferici transportaţi la distanţă, care sunt consideraţi
ca potenţială cauză ce contribuie la intensificarea degradării stării ecosistemelor forestiere. Astfel, poluanţii atmosferici care influenţează direct sau indirect declinul pădurilor sunt: (1) în stare gazoasă – compuşi ai sul-
fului (SO2, SO3), compuşi ai azotului (NO, NO2, HNO3, NH4NO3, HNO2, NH3), fotooxidanţi, hidrocarburi, clorofluorocarburi (freonii), amestecuri de poluanţi; (2) în stare lichidă – ploi şi ceaţă acide conţinând NO3
-,
SO42-
, NH4+, oxidanţi (H2O2), hidrocarburi; (3) în stare solidă (particule) – compuşi acizi ai sulfului şi ai azo-
tului conţinând NO3-, SO4
2-, NH4
+, metale grele, hidrocarburi [7].
Pentru supravegherea pe scară largă a efectelor poluării aerului asupra pădurii, în anul 1985, în temeiul
Convenţiei privind poluarea transfrontalieră a aerului atmosferic la distanţe mari (Geneva, 1979), la nivel european a fost lansat un program complex de cercetare (ICP Forests – International Cooperative Programme
on Assessment and Monitoring of Air Pollution Effects on Forests) cu două niveluri diferite ale intensităţii de monitorizare, care derulează pe baza unor metode unitare de investigare, elaborate în comun de specialişti
din toate ţările europene [11]. Prima reţea (nivelul I) se bazează pe aproximativ 6000 de parcele de observare pe o grilă sistematică transnaţională de 16x16 km în întreaga Europă. Nivelul de supraveghere intensivă cu-
prinde aproximativ 700 de parcele de nivelul II în ecosisteme forestiere din Europa [9].

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
142
Pornind de la experienţa unor ţări europene de monitorizare a depunerilor atmosferice de ioni minerali în eco-sistemele forestiere şi de la faptul că până în prezent în Republica Moldova astfel de studii nu au fost efectuate [1], scopul cercetărilor noastre este de a evalua cantitativ şi calitativ depunerile atmosferice pentru a preveni impactul ecologic şi pentru a contribui la ameliorarea stării ecosistemelor forestiere din Republica Moldova.
Material şi metode În calitate de obiecte de studiu au servit ecosistemele forestiere din Republica Moldova în care sunt am-
plasate suprafeţele experimentale (SE) incluse în reţeaua europeană sistematică (16x16 km) de monitoring forestier. Aceasta a fost elaborată la nivelul întregului fond forestier european, ţării noastre revenindu-i sarci-na de a înregistra date în 10 suprafeţe experimentale (Fig.1).
Amplasarea colectoarelor de precipitaţii în suprafeţele experimentale pentru măsurarea şi eşantionarea lunară a acestora în vederea analizelor de laborator s-a realizat conform metodologiei recomandate de Programul ICP Forests [11]. Au fost instalate câte 5-8 colectoare de tip deschis (captator cu pungă din polietilenă) pentru precipi-taţii, un colector având suprafaţa de colectare de 571 cm
2. Colectoarele de precipitaţii de sub coronament au fost
amplasate în interiorul suprafeţei experimentale, la o distanţă de 7-10 m între ele, în formă de zig-zag, la înălţimea de 1,2 m de la sol. În scopul de a surprinde variabilitatea precipitaţiilor căzute sub coronament, amplasarea a fost realizată pe curba de nivel – în cazul terenurilor înclinate şi pe direcţia E-V – în cazul terenurilor plane [11].
Colectoarele de precipitaţii au fost improvizate şi confecţionate respectându-se anumite cerinţe [3,11]. Pentru perioada vegetativă a anului, marginea inferioară a colectorului este conectată la un rezervor (peturi de sub apă minerală/plată cu volumul de 3-3,5 l), care este îngropat în substratul de sol în scopul de a limita influenţa razelor solare şi de a minimaliza procesul de evaporare ş.a. Conexiunea dintre sac şi rezervor are un diametru mic, la fel pentru a minimaliza procesul de evaporare a probei colectate.
Fig.1. Amplasarea suprafeţelor experimentale din Republica Moldova.
După recepţionarea probelor, în condiţii de laborator a fost efectuată filtrarea probelor şi analiza chimică
propriu-zisă. Indicii chimici ai apei au fost determinaţi în cadrul laboratoarelor Calitatea Mediului şi Ecobioin-
dicaţie şi Radioecologie de la Intitutul de Ecologie şi Geografie, prin metodele recomandate de EMEP (1995).
Astfel, ionul NO3- a fost determinat prin metoda spectrofotometrică cu reactivul Griess; NH4
+ – prin metoda spec-
trofotometrică, cu reactivul Nessler; SO42-
– prin metoda spectrofotometrică, cu soluţie de BaCl2 [10].

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
143
Concentraţiile de sulf şi azot din ionii analizaţi în precipitaţiile atmosferice s-au calculat în baza raportului
masei atomice a elementului la masa atomică a ionului, pentru ionii de SO42-
, NO3- şi NH4
+. Pentru a calcula
fluxul de ioni pentru o anumită perioadă, s-a folosit formula:
F = ∑ Pi Ci/100,
unde:
F – fluxul cantitativ al ionului analizat, în kg/ha/perioadă;
Pi – precipitaţiile, în mm, corespunzătoare perioadei i;
Ci – concentraţia unui element sau compus, exprimată în mg/l, corespunzătoare perioadei i.
Rezultate şi discuţii
Studiul fluxurilor ionilor minerali atmosferici efectuat în 10 ecosisteme forestiere din Republicii Moldova,
în care sunt amplasate suprafeţele experimentale ale reţelei europene sistematice (16x16 km) de monitoring fo-
restier, denotă unele legităţi şi tendinţe de acumulare, în dependenţă de zona de amplasare şi tipul de poluant.
Rezultatele analizelor chimice demonstrează că în ecosistemele forestiere din Republica Moldova, pentru anul
2012, depunerile de S-SO42-
, conform scării de evaluare a depunerilor de sub coronament (,,throughfall”) la
nivel european [6,7,11], se atribuie categoriilor de depuneri mari (5,7-8 kg/ha/an) şi depuneri foarte mari (8-32
kg/ha/an) (Tab.2). Fluxurile ionilor de sulfat (S-SO42-
) determinate sub coronamentul pădurilor studiate variază
de la 7,57 kg/ha/an (SE 807) până la 12,50 kg/ha/an (SE 1117) (Tab.1). Astfel înregistrându-se un coeficient de
variaţie (CV) mic (13,7%), ceea ce demonstrează o distribuţie, practic omogenă, a depunerilor de S-SO42-
pe te-
ritoriul Republicii Moldova. Pentru majoritatea SE studiate (SE 304, 405, 206, 612, 210 şi 1315) valorile depu-
nerilor de S-SO42-
nu înregistrează diferenţe statistic demonstrate (DMS – 1,56). Valoarea minimă de 7,57
kgS/ha/an (SE 807) diferă semnificativ de valorile determinate pentru restul SE studiate, cu excepţia valorilor
din SE 1315 (8,38 kgS/ha/an); la fel, valoarea maximă a depunerilor atmosferice de S-SO42-
– 12,5 kgS/ha/an
(SE 1117) diferă semnificativ faţă de depunerile înregistrate în restul SE studiate.
Depunerile totale de S-SO42-
, înregistrate sub coronamentul pădurilor de foioase din Republica Moldova,
sunt determinate, în mare parte, de fluxurile transfrontaliere de S-SO42-
. Conform rapoartelor EMEP [4], în cva-
dratele EMEP (50x50 km), unde sunt amplasate SE 1315, 807 şi 1214 – ecosisteme în care s-au înregistrat de-
puneri mai reduse, comparativ cu restul SE studiate, cota fluxului transfrontalier constituie 80-90%. În cvadra-
tele unde sunt amplasate SE ce înregistrează depuneri sporite de S-SO42-
, cota fluxului transfrontalier fiind de
peste 90%. Astfel, pe lângă impactul transfrontalier SE cad şi sub influenţa surselor de poluare locală. Acestea,
în dependenţă de poziţia fizico-geografică şi de factorii biotici şi abiotici, manifestă un impact negativ indivi-
dual pentru fiecare ecosistem studiat, care şi determină micile diferenţe dintre depunerile înregistrate.
Depunerile totale ale ionilor de nitrat (N-NO3-) înregistrate sub coronamentul pădurilor studiate au indicat
valori cuprinse între 1,53 şi 5,2 kgN/ha/an (Tab.1), fluxuri ce reprezintă o repartizare neuniformă, comparativ
cu depunerile de S-SO42-
. Tendinţele cele mai scăzute privind depunerile de N-NO3- s-au înregistrat în ecosiste-
mele forestiere amplasate în zona de nord a republicii (SE 304, 405, 206, 807 şi 612), ale căror variaţii nu au di-
ferenţe semnificative (DMS – 1,15), cu excepţia depunerilor din SE 807 (1,5 kgN/ha/an), care este semnificativ
mai mică comparativ cu restul SE studiate, excepţie făcând SE 405. Conform scării de evaluare a depunerilor
de N-NO3- la nivel european, depunerile de N-NO3
- înregistrate sub coronamentele studiate s-au încadrat, în
majoritatea cazurilor, în categoria depunerilor reduse (1,8-3,2 kg/ha/an). Depuneri foarte reduse (0-1,8 kg/ha/an)
sunt specifice pentru SE 807 (1,53 kg/ha/an), depuneri mijlocii (3,2-4,5 kg/ha/an) – pentru SE 1214 şi 1315
(4,2-3,6 kg/ha/an), iar depuneri mari (4,5-6,3 kg/ha/an) – pentru SE 1117 (5,2 kg/ha/an).
Tabelul 1
Depunerile anuale de ioni minerali înregistrate sub coronament (,,throughfall”) în ecosistemele
forestiere studiate din Republica Moldova, 2012
Suprafaţa
experimen-
tală (SE)
Întreprinde-
rea de Stat
pentru
Silvicultură
Ocolul
Silvic
Compoziţia
dendrologică
S-SO4
(kg/ha/an) N-NO3
(kg/ha/an) N-NH4
(kg/ha/an)
SE 304 Edineţ Briceni 7ST2STR1MO 10,42 2,83 6,36
SE 405 Edineţ Edineţ 10FR 10,18 2,27 6,15

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
144
SE 206 Edineţ Otaci 7GO1FR2TE 10,11 2,79 6,31
SE 807 Glodeni Călineşti 10ST+CI 7,57 1,53 4,04
SE 612 Şoldăneşti Olişcani 10ST+FR 10,85 2,81 6,35
SE 210 Călăraş Vărzăreşti 7FR2ST1ULC 10,76 3,17 6,77
SE 1214 Hânceşti Buţeni 10ST+FR 9,19 4,16 9,15
SE 1213 Hânceşti Mereşeni 6GO4FR 10,38 2,77 8,99
SE 1117 Tighina Hârbovăţ 10STP+SC 12,50 5,20 9,10
SE 1315 Răzeni Zloţi 10SC 8,38 3,60 6,56
Diferenţa minimal semnificativă (DMS) 1,56 1,15 1,85
Amplitudinea Minimală 7,6 1,5 4,0
Maximală 12,5 5,2 9,2
Media aritmetică 10,0 3,1 7,0
Deviaţia standartd 1,4 1,0 1,6
Coeficientul de variaţie 13,7 32,7 23,4
Notă: ST – Stejar penduculat (Quercus robur), STR – Stejar roşu (Quercus rubra), STP – Stejar pufos (Quercus
pubescens), GO – Gorun (Quercus petraea), FR – Frasin comun (Fraxinus excelsior), MO – Molid (Picea abies),
CI –Cireş (Cerasusavium), TE – Tei argintiu (Tilia tomentosa), SC – Salcâm (Robinia pseudacacia), ULC – Ulm de
câmp (Ulmus campestris).
Suprafeţele experimentale 1117 şi 1214, amplasate în zone cu impact sporit al emisiilor de la transportul
auto (pentru SE 1214 – emisiile de la fluxul sporit al unităţilor de transport de pe traseul auto Chişinău –
Cahul şi din or. Chişinău şi Hânceşti, iar pentru SE 1117 – emisiile de la unităţile de transport de pe traseul
auto Chişinău – Tighina – Tiraspol şi din oraşele respective), înregistrează valori maxime ale depunerilor de
N-NO3- (5,2 şi, respectiv, 4,16 kgN/ha/an).
Tabelul 2
Scala europeană pentru aprecierea intensităţii depunerilor de ioni minerali sub cotonament [6,9]
Intensitatea depunerilor
(ICP Forests, 2010; 2012)
S-SO4
(kg/ha/an)
N-NO3
(kg/ha/an)
N-NH4
(kg/ha/an)
Depuneri foarte reduse 0-3,3 0-1,8 0-1,6
Depuneri reduse 3,3-4,2 1,8-3,2 1,6-3,3
Depuneri mijlocii 4,2-5,7 3,2-4,5 3,3-5,1
Depuneri mari 5,7-8 4,5-6,3 5,1-7,5
Depuneri foarte mari 8-32 6,3-23,5 7,5-22,4
Fluxul ionilor de N-NH4+ de sub coronament, în 6 cazuri (SE 304, 405, 206, 612, 201 şi 1315), se încadrează
în categoria depunerilor mari (5,1-7,5 kg/ha/an), în trei cazuri (SE 1214, 1213 şi 1117) în categoria depunerilor
foarte mari (7,5-22,4 kg/ha/an) şi într-un caz, SE 807, a depunerilor mijlocii (3,3-5,1 kg/ha/an) (Tab.1 şi 2). Valo-
rile cele mai mari s-au înregistrat în SE 1214, 1117 şi 1213, constituind 9,15 kgN/ha/an, 9,10 kgN/ha/an şi, res-
pectiv, 8,99 kgN/ha/an, valori statistic semnificativ mai mari (DMS = 1,85) decât în restul SE, indicând un aport
mai sporit al influenţei emisiilor locale prin sporirea depunerilor uscate alături de depunerile transfrontaliere. În
restul ecosistemelor forestiere studiate, depunerile anuale ale N-NH4+ sub coronament au fost, practic, aceleaşi, cu
excepţia SE 807, care a avut depuneri semnificativ mai reduse decât în toate ecosistemele forestiere studiate.
Comparând fluxurile ionilor de N-NH4+ cu cel de N-NO3
-, determinate sub coronament, constatăm că în
majoritatea arboretelor studiate (SE 304, 405, 206, 807, 612, 210 şi 1214) fluxurile de N-NH4+ sunt, în
medie, de 2,5 ori mai ridicate decât cele de N-NO3-. Rezultatele obţinute demonstrează clar dominanţa ionu-
lui de N-NH4+ faţă de ionul de N-NO3
- în depunerile atmosferice, rezultate înregistrate şi în cercetările efec-
tuate în peste 500 de SE europene în cadrul Programului ICP Forests [9]. În funcţie de poziţia fizico-geografică a ecosistemelor forestiere studiate, constatăm că în ecosistemele
din nordul republicii (SE 304, 405, 206, 807, 612) s-au înregistrat depuneri mai reduse faţă de ecosistemele din centrul republicii (SE 210, 1214, 1213, 1117, 1315) (Tab.1). O diferenţă semnificativă se înregistrează

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
145
pentru depunerile de azot, unde pentru NO3- fluxul mediu înregistrat în ecosistemele din centrul republicii
este cu circa 21% mai sporit faţă de fluxul înregistrat în zona de nord. Pentru NH4+ se înregistrează aproxi-
mativ acelaşi tablou, cu 16% şi, în sumă, riscul poluării ecosistemelor forestiere cu azot anorganic total (NO3
- + NH4
+) este cu 18% mai sporit pentru ecosistemele din zona de centru faţă de cele din zona de nord.
Deci, observăm că pentru toţi ionii analizaţi suprafaţa experimentală 807, amplasată în zona est-nordică a repu-blicii (Întreprinderea de Stat pentru Silvicultură – Glodeni, Ocolul Silvic – Călineşti), înregistrează cele mai reduse depuneri totale de ioni atmosferici. Factorul principal, care a influenţat aceste depuneri, este deficitul de precipitaţii anuale înregistrate în această zonă (298 mm). Astfel, cu cantităţi reduse de precipitaţii au fost antrenate, respectiv, cantităţi reduse de depuneri atmosferice umede, depuneri cărora le revine o cotă importantă în depunerile totale.
Analiza comparativă a rezultatelor obţinute în anul 2012 în ecosistemele forestiere din ţara noastră, cu cele determinate în alte regiuni din Europa, raportate în anii 2010-2012, pentru suprafeţele europene de mo-nitoring intensiv (nivelul II), conform ,,Technical Report of ICP Forests” [5,6,9], s-a realizat prin completa-rea hărţilor de distribuţie a depunerilor sub coronament cu valori originale pentru Republica Moldova, calcu-late în baza modelelor prezentate de Fischer et al. (2010) şi Lorenz et al. (2012).
Depunerile anuale ale ionilor de S-SO42-
, înregistrate sub coronamentul pădurilor din Republica Moldova, au fost mari şi foarte mari şi sunt comparabile cu cele înregistrate în Europa Centrală: Polonia, Germania, Republica Cehă, Austria şi România (Fig.2). Conform rapoartelor Programului ICP Forests [6,9], în suprafe-ţele europene de monitoring intensiv (nivelul II), depunerile medii anuale ale ionilor de S-SO4
2-, pentru
ultimii 5 ani, au constituit: 16,3% – depuneri foarte reduse, 9,3% – depuneri reduse, 18,6% – depuneri mij-locii, 29,3% – depuneri mari şi 26,5% – depuneri foarte mari. Astfel, depunerile înregistrate în ecosistemele forestiere din Republica Moldova se încadrează în ultimele două categorii, care sunt cele mai specifice ţărilor europene, depuneri (mari şi foarte mari) ce pot spori riscul degradării pădurilor de foioase prin distrugerea clorofilei şi diminuarea procesului de fotosinteză.
Depunerile reduse de N-NO3- înregistrate sub coronamentul arboretelor studiate în Republica Moldova
sunt comparabile cu depunerile înregistrate în România şi în alte regiuni din centrul şi central-estice ale Europei – Slovacia, Austria, Polonia, Lituania (Fig.3). Pentru ultimii 5 ani aceste depuneri în pădurile din Europa au constituit: 12,6% – depuneri foarte reduse, 9,7% – depuneri reduse, 11,6% – depuneri mijlocii, 24,2% – depuneri mari şi 41,9% – depuneri foarte mari. Deci, majoritatea ecosistemelor studiate din Repu-blica Moldova au înregistrat depuneri reduse, categorie căreia îi revine cea mai mică cotă la nivel european, fapt ce exclude riscul provocării unor efecte de poluare şi degradare ulterioară a ecosistemelor forestiere.
A B
Fig.2. (A) Repartiţia depunerilor de S-SO42-
sub coronament (,,throughfall”) în suprafeţele de monitoring intensiv din Europa, după Fischer et al., 2010 şi Lorenz et al., 2012. (B) Valorile depunerilor
de S-SO42-
sub coronament în suprafeţele experimentale din Republica Moldova completate de noi constituie date originale pentru anul 2012, kg/ha/an.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
146
A B
Fig.3. (A) Repartiţia depunerilor de N-NO3- sub coronament (“throughfall”) în suprafeţele de monitoring intensiv din Europa,
după Fischer et al., 2010 şi Lorenz et al., 2012. (B) Valorile depunerilor de N-NO3- sub coronament în suprafeţele
experimentale din Republica Moldova completate de noi constituie date originale pentru anul 2012, kg/ha/an.
Fluxul mediu anual al ionilor de N-NH4
+, înregistrat în ecosistemele forestiere studiate din Republica
Moldova, s-a încadrat în categoriile depuneri mari şi foarte mari, valori caracteristice depunerilor din Europa Centrală şi Central–Estică: Germania, Republica Cehă, Austria, Polonia, Lituania, Slovacia şi România (Fig.4). La nivel european, depunerile medii anuale de N-NH4
+ sub coronamentul arboretelor au avut urmă-
toarea pondere: 13,0% – depuneri foarte reduse, 13,0% – depuneri reduse, 17,7% – depuneri mijlocii, 24,2% – depuneri mari şi 32,1% – depuneri foarte mari [5,6,9]. Ca şi în cazul depunerilor de S-SO4
2-, depunerile
ionilor de N-NH4+, înregistrate în ecosistemele forestiere studiate, se încadrează în categoriile cele mai repre-
zentative pentru ecosistemele forestiere din Europa – depuneri mari şi foarte mari.
A B
Fig.4. (A) Repartiţia depunerilor de N-NH4+ sub coronament (,,throughfall”) în suprafeţele de monitoring intensiv din Europa,
după Fischer et al., 2010 şi Lorenz et al., 2012. (B) Valorile depunerilor de N-NH4+ sub coronament în suprafeţele
experimentale din Republica Moldova completate de noi constituie date originale pentru anul 2012, kg/ha/an.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
147
Concluzii În ecosistemele forestiere din nordul republicii (SE 304, 405, 206, 807, 612) s-au înregistrat depuneri mai
reduse faţă de ecosistemele din centrul republicii (SE 210, 1214, 1117, 1315). Astfel, cele din urmă sunt supuse
unui risc mai sporit de ameninţare a stării de sănătate a arborilor şi a funcţionalităţii ecosistemelor forestiere.
Depunerile ionilor minerali (SO42-
, NO3-, NH4
+), înregistrate sub coronamentul arboretelor studiate din
Republica Moldova sunt comparabile cu depunerile înregistrate în România şi în alte regiuni din centrul
Europei şi în cele central–estice ale ei (Slovacia, Republica Cehă, Austria, Polonia, Lituania).
Bibliografie:
1. BOAGHIE, D. Monitoring ecologic şi forestier. Chişinău, 2004. 186 p.
2. DRAAIJERS, G. The variability of atmospheric deposition to forests: the effects of canopy structure and forest
edges. Ph.D. Thesis, University of Utrecht, Netherlands, 1993, 200 p.
3. EMEP, Co-operative Programme for Monitoring and Evaluation of the Long-Range Transmission of Air Pollutants
in Europe. Manual for sampling and chemical analysis. EMEP/CCC-Report 1/95, Norwegian In. for Air Res.,
Kyeller, Norway, 1995, 176 p.
4. EMEP Status Report 1/2013. Transboundary air pollution by main pollutants (S, N, O3) and PM in 2011. The
Republic of Moldova. Joint MSC-W & CCC & CEIP Report, 2013, 24 p. [Accesat 06.09.2013] Disponibil:
http://www.emep.int/mscw/index_mscw.html
5. FISCHER, R., LORENZ, M., KÖHL, M. et al. Forest Condition in Europe, 2010. Technical Report of ICP Forests.
Work Report of the Institute for World Forestry 2010/1. ICP Forests, Hamburg, 2010. 175 p
6. FISCHER, R., WALDNER, P., CARNICER, J. et al. The Condition of Forests in Europe, 2012. Executive Report. ICP
Forests, Hamburg, 2012, 24 p. [Accesat 03.09.2013] Disponibil: [http://www.icp-forests.org/RepEx.htm]
7. IACOBAN, C. Evaluarea poluării atmosferice în ecosisteme forestiere din România. Bucureşti: Editura Silvică,
2009. 243 p.
8. IMPENS, R. Les pluies acides. Une menace pour nos forets. An. Gembloux, 1984. p.13-17.
9. LORENZ, M., BECHER, G. Forest Condition in Europe, 2012. Technical Report of ICP Forests. Work Report of
the Thünen Institute for World Forestry 2012/1. Hamburg, 2012. 167 p.
10. SANDU, M., LOZAN, R., TĂRÎŢĂ, A. Metode şi instrucţiuni privind controlul calităţii apelor. Chişinău: ,,Ericon”
SRL, 2010. 173 p.
11. UN/ECE, ICP Forests. Sampling and Analysis of Deposition. Part XIV. In: Manual on methods and criteria for
harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests. Hamburg, 2010,
66 p. [Accesat: 26.03.2013] Disponibil: [http://www.icp-forests.org/Manual.htm]
Prezentat la 01.11.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
148
PARTICULARITĂŢI ŞI TENDINŢE ALE UNOR VALORI CLIMATICE DIN CADRUL
REZERVAŢIEI ŞTIINŢIFICE ,,PĂDUREA DOMNEASCĂ”
Iulian MAMAI
Universitatea de Stat din Moldova
Existenţa şi modul de repartizare structurală a ecosistemelor se află într-o strânsă corelaţie cu factorii climaterici.
Acţiunea factorilor climaterici determină repartiţia diferenţiată a plantelor în dependenţă de preferinţele ecologice ale
fiecărei specii. Toate procesele biofizice şi biochimice ale plantelor şi cele de creştere şi dezvoltare sunt influenţate de
factorul climateric. Astfel, distribuţia vegetaţiei din cadrul unei lunci se face în dependenţă de limitele de toleranţă a
fiecărei specii şi se află într-o dependenţă faţă de variaţiile condiţiilor climaterice existente.
Cuvinte-cheie: elemente climatice, rezervaţie naturală, precipitaţii, temperatură.
PECULIARITIES AND TRENDS OF CLIMATE VALUES
OF THE NATURAL RESERVE “PĂDUREA DOMNEASCĂ”
The existence and way of structural distribution of the ecosystem are in a tight correlation with climatic factors. The
action of climatic factors determines the differentiated distribution of the plants according to the ecological preferences
of each species. All biophysical and biochemical processes of plants and the growth and development are influenced by
climatic factor. Thus, the distribution of vegetation within a meadow is made depending on the tolerance limits of each
species and is in a dependency upon the existing climatic conditions changes.
Keywords: climate elements, natural reserve, precipitation, temperature.
Introducere
Analiza factorilor de mediu care au un impact major asupra menţinerii echilibrului ecologic din această
zonă ar putea constitui un mesaj clar cu privire la modul şi măsurile necesare de aplicat în procesul de gestio-
nare a resurselor naturale din cadrul rezervaţiei, totodată ar reprezenta şi un indicator-cheie cu privire la elu-
cidarea modului de acţiune în timp a climatului.
Cunoaşterea factorilor climaterici care ar putea fi consideraţi în zona respectivă ca factori limitativi sau a
căror acţiune impune o anumită tendinţă de dezvoltare a ecosistemului dat ar permite în viitor diminuarea
acţiunii negative a acestor factori prin implementarea unor măsuri de redresare sau minimalizarea efectului
indus de acţiunea factorilor respectivi.
Acţiunea factorilor climaterici determină repartiţia diferenţiată a plantelor în dependenţă de preferinţele
ecologice ale fiecărei specii, determinând pe parcursul timpului adaptări morfologice care au ca rezultantă
evoluţia în timp a biocenozei asupra căreia acţionează.
Material şi metode
Pentru a avea o viziune cât mai amplă asupra particularităţilor şi tendinţelor indicilor climaterici care au
acţionat în zona dată de-a lungul timpului a fost analizată evoluţia indicilor respectivi pentru o perioadă de
cca 112 ani.
Vom utiliza două baze de date cu privire la analiza factorilor climaterici care ar prezenta interes în con-
textul temei noastre de cercetare. Prima bază de date este informaţia oferită de Serviciul Hidrometeorologic
de Stat cu referire la cantitatea de precipitaţii lunare şi la temperatura medie lunară a aerului pentru perioada
1958-2011, înregistrată în cadrul Staţiei Meteorologice din oraşul Făleşti (cea mai apropiată staţie faţă de zona
de studiu)*. Cea de a doua bază de date cu privire la cantitatea de precipitaţii lunare şi la temperatura medie
lunară a aerului pentru zona de studiu a fost preluată din baza de date climatice cu rezoluţia de 0,5ox0,5
o CRU2
(Mitchell şi Jones, 2005) pentru perioada 1901-2009 1 .
În acest context au fost trasate o serie de obiective:
* Informaţia privind cantitatea de precipitaţii lunare şi temperatura medie lunară a aerului pentru perioada 1958-2011,
înregistrate în cadrul Staţiei Meteorologice din oraşul Făleşti, a fost preluată de la Serviciul Hidrometeorologic de Stat
al Republicii Moldova.
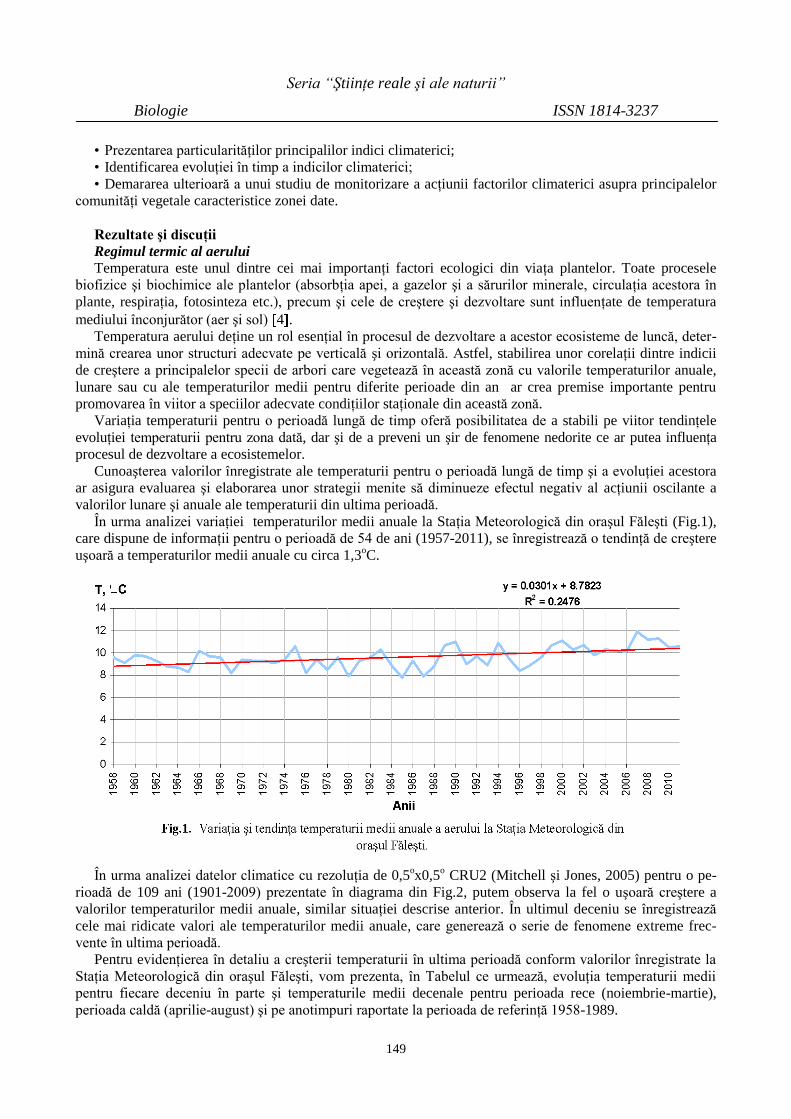
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
149
• Prezentarea particularităţilor principalilor indici climaterici;
• Identificarea evoluţiei în timp a indicilor climaterici;
• Demararea ulterioară a unui studiu de monitorizare a acţiunii factorilor climaterici asupra principalelor
comunităţi vegetale caracteristice zonei date.
Rezultate şi discuţii
Regimul termic al aerului
Temperatura este unul dintre cei mai importanţi factori ecologici din viaţa plantelor. Toate procesele
biofizice şi biochimice ale plantelor (absorbţia apei, a gazelor şi a sărurilor minerale, circulaţia acestora în
plante, respiraţia, fotosinteza etc.), precum şi cele de creştere şi dezvoltare sunt influenţate de temperatura
mediului înconjurător (aer şi sol) 4 .
Temperatura aerului deţine un rol esenţial în procesul de dezvoltare a acestor ecosisteme de luncă, deter-
mină crearea unor structuri adecvate pe verticală şi orizontală. Astfel, stabilirea unor corelaţii dintre indicii
de creştere a principalelor specii de arbori care vegetează în această zonă cu valorile temperaturilor anuale,
lunare sau cu ale temperaturilor medii pentru diferite perioade din an ar crea premise importante pentru
promovarea în viitor a speciilor adecvate condiţiilor staţionale din această zonă.
Variaţia temperaturii pentru o perioadă lungă de timp oferă posibilitatea de a stabili pe viitor tendinţele
evoluţiei temperaturii pentru zona dată, dar şi de a preveni un şir de fenomene nedorite ce ar putea influenţa
procesul de dezvoltare a ecosistemelor.
Cunoaşterea valorilor înregistrate ale temperaturii pentru o perioadă lungă de timp şi a evoluţiei acestora
ar asigura evaluarea şi elaborarea unor strategii menite să diminueze efectul negativ al acţiunii oscilante a
valorilor lunare şi anuale ale temperaturii din ultima perioadă.
În urma analizei variaţiei temperaturilor medii anuale la Staţia Meteorologică din oraşul Făleşti (Fig.1),
care dispune de informaţii pentru o perioadă de 54 de ani (1957-2011), se înregistrează o tendinţă de creştere
uşoară a temperaturilor medii anuale cu circa 1,3oC.
În urma analizei datelor climatice cu rezoluţia de 0,5ox0,5
o CRU2 (Mitchell şi Jones, 2005) pentru o pe-
rioadă de 109 ani (1901-2009) prezentate în diagrama din Fig.2, putem observa la fel o uşoară creştere a
valorilor temperaturilor medii anuale, similar situaţiei descrise anterior. În ultimul deceniu se înregistrează
cele mai ridicate valori ale temperaturilor medii anuale, care generează o serie de fenomene extreme frec-
vente în ultima perioadă.
Pentru evidenţierea în detaliu a creşterii temperaturii în ultima perioadă conform valorilor înregistrate la
Staţia Meteorologică din oraşul Făleşti, vom prezenta, în Tabelul ce urmează, evoluţia temperaturii medii
pentru fiecare deceniu în parte şi temperaturile medii decenale pentru perioada rece (noiembrie-martie),
perioada caldă (aprilie-august) şi pe anotimpuri raportate la perioada de referinţă 1958-1989.
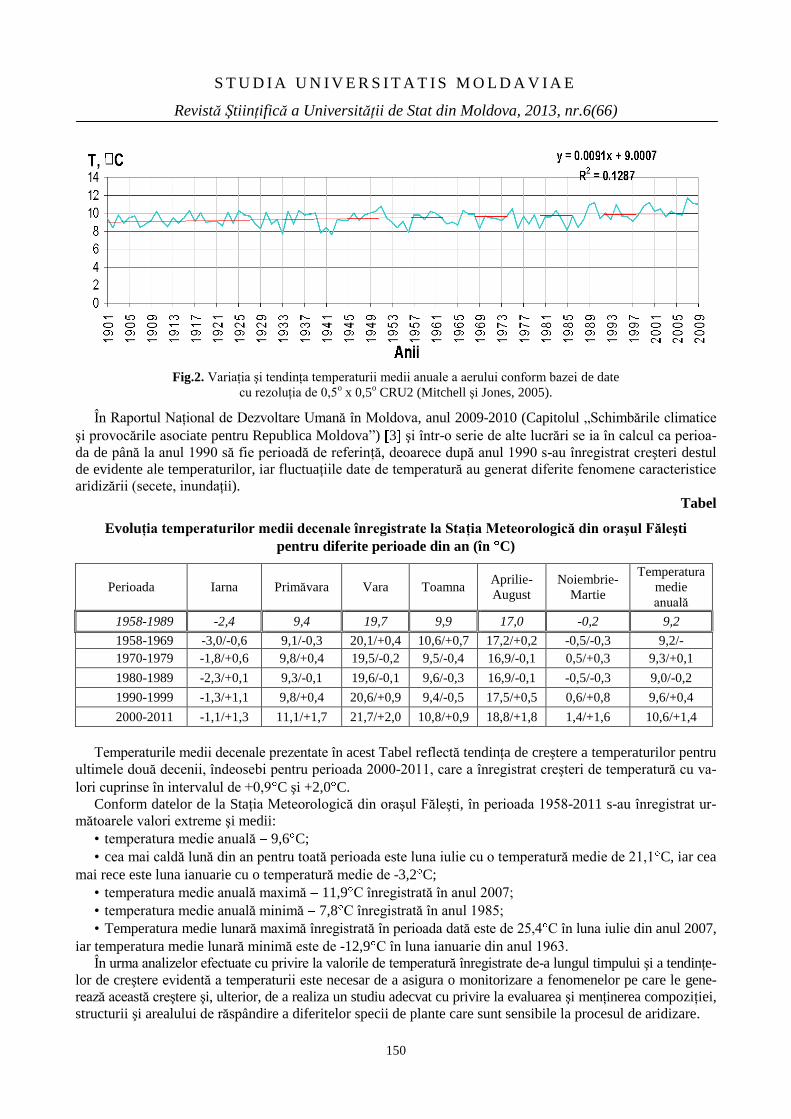
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
150
Fig.2. Variaţia şi tendinţa temperaturii medii anuale a aerului conform bazei de date cu rezoluţia de 0,5
o x 0,5
o CRU2 (Mitchell şi Jones, 2005).
În Raportul Naţional de Dezvoltare Umană în Moldova, anul 2009-2010 (Capitolul „Schimbările climatice
şi provocările asociate pentru Republica Moldova”) 3 şi într-o serie de alte lucrări se ia în calcul ca perioa-
da de până la anul 1990 să fie perioadă de referinţă, deoarece după anul 1990 s-au înregistrat creşteri destul
de evidente ale temperaturilor, iar fluctuaţiile date de temperatură au generat diferite fenomene caracteristice
aridizării (secete, inundaţii).
Tabel
Evoluţia temperaturilor medii decenale înregistrate la Staţia Meteorologică din oraşul Făleşti
pentru diferite perioade din an (în C)
Perioada Iarna Primăvara Vara Toamna Aprilie-
August
Noiembrie-
Martie
Temperatura
medie
anuală
1958-1989 -2,4 9,4 19,7 9,9 17,0 -0,2 9,2
1958-1969 -3,0/-0,6 9,1/-0,3 20,1/+0,4 10,6/+0,7 17,2/+0,2 -0,5/-0,3 9,2/-
1970-1979 -1,8/+0,6 9,8/+0,4 19,5/-0,2 9,5/-0,4 16,9/-0,1 0,5/+0,3 9,3/+0,1
1980-1989 -2,3/+0,1 9,3/-0,1 19,6/-0,1 9,6/-0,3 16,9/-0,1 -0,5/-0,3 9,0/-0,2
1990-1999 -1,3/+1,1 9,8/+0,4 20,6/+0,9 9,4/-0,5 17,5/+0,5 0,6/+0,8 9,6/+0,4
2000-2011 -1,1/+1,3 11,1/+1,7 21,7/+2,0 10,8/+0,9 18,8/+1,8 1,4/+1,6 10,6/+1,4
Temperaturile medii decenale prezentate în acest Tabel reflectă tendinţa de creştere a temperaturilor pentru
ultimele două decenii, îndeosebi pentru perioada 2000-2011, care a înregistrat creşteri de temperatură cu va-
lori cuprinse în intervalul de +0,9 C şi +2,0 C.
Conform datelor de la Staţia Meteorologică din oraşul Făleşti, în perioada 1958-2011 s-au înregistrat ur-
mătoarele valori extreme şi medii:
• temperatura medie anuală 9,6 C;
• cea mai caldă lună din an pentru toată perioada este luna iulie cu o temperatură medie de 21,1 C, iar cea
mai rece este luna ianuarie cu o temperatură medie de -3,2 C;
• temperatura medie anuală maximă 11,9 C înregistrată în anul 2007;
• temperatura medie anuală minimă 7,8 C înregistrată în anul 1985;
• Temperatura medie lunară maximă înregistrată în perioada dată este de 25,4 C în luna iulie din anul 2007,
iar temperatura medie lunară minimă este de -12,9 C în luna ianuarie din anul 1963.
În urma analizelor efectuate cu privire la valorile de temperatură înregistrate de-a lungul timpului şi a tendinţe-
lor de creştere evidentă a temperaturii este necesar de a asigura o monitorizare a fenomenelor pe care le gene-
rează această creştere şi, ulterior, de a realiza un studiu adecvat cu privire la evaluarea şi menţinerea compoziţiei,
structurii şi arealului de răspândire a diferitelor specii de plante care sunt sensibile la procesul de aridizare.
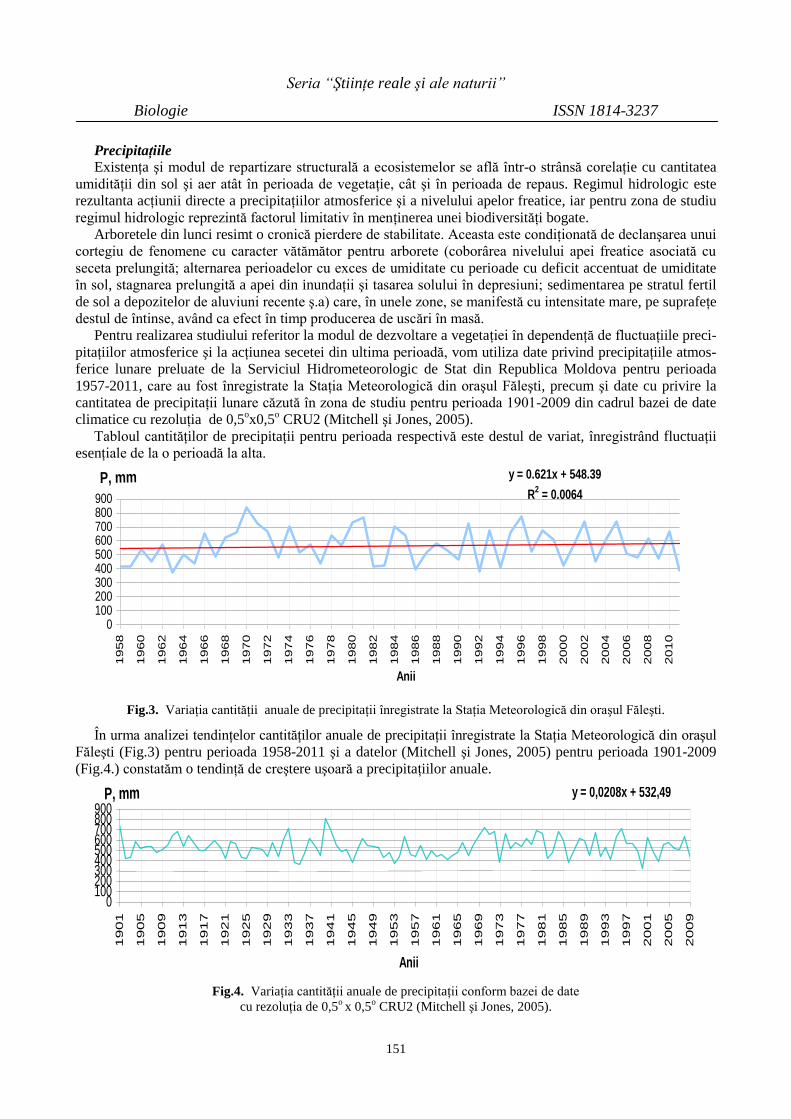
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
151
Precipitaţiile
Existenţa şi modul de repartizare structurală a ecosistemelor se află într-o strânsă corelaţie cu cantitatea
umidităţii din sol şi aer atât în perioada de vegetaţie, cât şi în perioada de repaus. Regimul hidrologic este
rezultanta acţiunii directe a precipitaţiilor atmosferice şi a nivelului apelor freatice, iar pentru zona de studiu
regimul hidrologic reprezintă factorul limitativ în menţinerea unei biodiversităţi bogate.
Arboretele din lunci resimt o cronică pierdere de stabilitate. Aceasta este condiţionată de declanşarea unui
cortegiu de fenomene cu caracter vătămător pentru arborete (coborârea nivelului apei freatice asociată cu
seceta prelungită; alternarea perioadelor cu exces de umiditate cu perioade cu deficit accentuat de umiditate
în sol, stagnarea prelungită a apei din inundaţii şi tasarea solului în depresiuni; sedimentarea pe stratul fertil
de sol a depozitelor de aluviuni recente ş.a) care, în unele zone, se manifestă cu intensitate mare, pe suprafeţe
destul de întinse, având ca efect în timp producerea de uscări în masă.
Pentru realizarea studiului referitor la modul de dezvoltare a vegetaţiei în dependenţă de fluctuaţiile preci-
pitaţiilor atmosferice şi la acţiunea secetei din ultima perioadă, vom utiliza date privind precipitaţiile atmos-
ferice lunare preluate de la Serviciul Hidrometeorologic de Stat din Republica Moldova pentru perioada
1957-2011, care au fost înregistrate la Staţia Meteorologică din oraşul Făleşti, precum şi date cu privire la
cantitatea de precipitaţii lunare căzută în zona de studiu pentru perioada 1901-2009 din cadrul bazei de date
climatice cu rezoluţia de 0,5ox0,5
o CRU2 (Mitchell şi Jones, 2005).
Tabloul cantităţilor de precipitaţii pentru perioada respectivă este destul de variat, înregistrând fluctuaţii
esenţiale de la o perioadă la alta.
y = 0.621x + 548.39
R2 = 0.0064
0100200300400500600700800900
1958
1960
1962
1964
1966
1968
1970
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
Anii
P, mm
Fig.3. Variaţia cantităţii anuale de precipitaţii înregistrate la Staţia Meteorologică din oraşul Făleşti.
În urma analizei tendinţelor cantităţilor anuale de precipitaţii înregistrate la Staţia Meteorologică din oraşul
Făleşti (Fig.3) pentru perioada 1958-2011 şi a datelor (Mitchell şi Jones, 2005) pentru perioada 1901-2009
(Fig.4.) constatăm o tendinţă de creştere uşoară a precipitaţiilor anuale.
y = 0,0208x + 532,49
0100200300400500600700800900
1901
1905
1909
1913
1917
1921
1925
1929
1933
1937
1941
1945
1949
1953
1957
1961
1965
1969
1973
1977
1981
1985
1989
1993
1997
2001
2005
2009
Anii
P, mm
Fig.4. Variaţia cantităţii anuale de precipitaţii conform bazei de date
cu rezoluţia de 0,5o x 0,5
o CRU2 (Mitchell şi Jones, 2005).
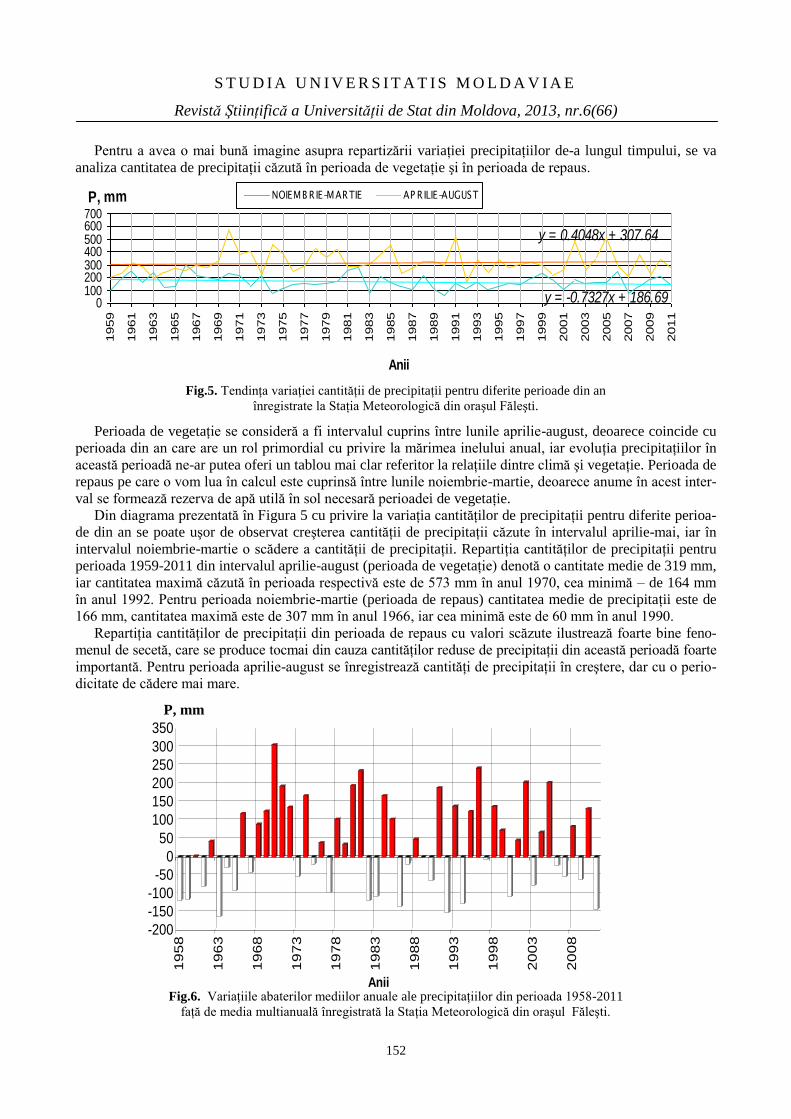
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
152
Pentru a avea o mai bună imagine asupra repartizării variaţiei precipitaţiilor de-a lungul timpului, se va
analiza cantitatea de precipitaţii căzută în perioada de vegetaţie şi în perioada de repaus.
y = -0.7327x + 186.69
y = 0.4048x + 307.64
0100200300400500600700
1959
1961
1963
1965
1967
1969
1971
1973
1975
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
2003
2005
2007
2009
2011
Anii
P, mm NOIEMB R IE-MARTIE AP R ILIE-AUGUST
Fig.5. Tendinţa variaţiei cantităţii de precipitaţii pentru diferite perioade din an
înregistrate la Staţia Meteorologică din oraşul Făleşti.
Perioada de vegetaţie se consideră a fi intervalul cuprins între lunile aprilie-august, deoarece coincide cu
perioada din an care are un rol primordial cu privire la mărimea inelului anual, iar evoluţia precipitaţiilor în
această perioadă ne-ar putea oferi un tablou mai clar referitor la relaţiile dintre climă şi vegetaţie. Perioada de
repaus pe care o vom lua în calcul este cuprinsă între lunile noiembrie-martie, deoarece anume în acest inter-
val se formează rezerva de apă utilă în sol necesară perioadei de vegetaţie.
Din diagrama prezentată în Figura 5 cu privire la variaţia cantităţilor de precipitaţii pentru diferite perioa-
de din an se poate uşor de observat creşterea cantităţii de precipitaţii căzute în intervalul aprilie-mai, iar în
intervalul noiembrie-martie o scădere a cantităţii de precipitaţii. Repartiţia cantităţilor de precipitaţii pentru
perioada 1959-2011 din intervalul aprilie-august (perioada de vegetaţie) denotă o cantitate medie de 319 mm,
iar cantitatea maximă căzută în perioada respectivă este de 573 mm în anul 1970, cea minimă – de 164 mm
în anul 1992. Pentru perioada noiembrie-martie (perioada de repaus) cantitatea medie de precipitaţii este de
166 mm, cantitatea maximă este de 307 mm în anul 1966, iar cea minimă este de 60 mm în anul 1990.
Repartiţia cantităţilor de precipitaţii din perioada de repaus cu valori scăzute ilustrează foarte bine feno-
menul de secetă, care se produce tocmai din cauza cantităţilor reduse de precipitaţii din această perioadă foarte
importantă. Pentru perioada aprilie-august se înregistrează cantităţi de precipitaţii în creştere, dar cu o perio-
dicitate de cădere mai mare.
P, mm
-200
-150
-100
-50
0
50
100
150
200
250
300
350
19
58
19
63
19
68
19
73
19
78
19
83
19
88
19
93
19
98
20
03
20
08
Anii
Fig.6. Variaţiile abaterilor mediilor anuale ale precipitaţiilor din perioada 1958 - 2011
faţă de media multianuală înregistrată la Stația Meteorologică din oraşul Făleşti.
Fig.6. Variaţiile abaterilor mediilor anuale ale precipitaţiilor din perioada 1958-2011
faţă de media multianuală înregistrată la Staţia Meteorologică din oraşul Făleşti.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
153
Prezentarea grafică a variaţiilor abaterilor mediilor anuale ale precipitaţiilor faţă de media multianuală (Fig.6),
care este considerată cantitate optimă de precipitaţii, evidenţiază anii sau perioadele care au înregistrat căderi
de precipitaţii în exces sau insuficienţă faţă de media multianuală calculată pentru perioada 1958-2011.
Anii cu precipitaţii în exces faţă de cele medii sunt anii ploioşi, care s-au înregistat în perioada 1968-1972,
1978-1982; în rest, perioadele cu exces şi cu insuficienţă de precipitaţii au înregistrat oscilaţii cu o periodi-
citate în sens pozitiv sau negativ de 1-2 ani.
Anii secetoşi sunt cei în care precipitaţiile sunt deficitare, respectiv cantitatea totală este sub valoarea
medie multianuală considerată normală, iar distribuţia ploilor pe parcursul sezonului de vegetaţie este neco-
respunzătoare în raport cu cerinţele speciilor forestiere 2 .
Concluzii
1. Pentru zona de luncă un rol important revine factorilor climaterici, deoarece ei determină dezvoltarea
vegetaţiei, iar oscilaţiile sau intensitatea exagerată cu care acţionează aceşti factori pot duce la un dezechilibru
ecologic sau la unele dereglări ale potenţialului productiv al acestei zone; astfel, aceşti factori climaterici pot
deveni destabilizatori şi limitativi.
2. Condiţiile climaterice reprezintă unul dintre cei mai importanţi factori care pot determina existenţa şi
distribuţia vegetaţiei atât pe verticală, cât şi pe orizontală. Toate procesele biofizice şi biochimice ale plante-
lor şi cele de creştere şi dezvoltare sunt influenţate de factorul climateric. Astfel, distribuţia vegetaţiei din
cadrul unei lunci se face în dependenţă de limitele de toleranţă ale fiecărei specii şi se află într-o dependenţă
faţă de variaţiile condiţiilor climaterice existente.
3. Din analiza abaterilor valorilor medii lunare faţă de valorile medii multianuale lunare ale temperaturi-
lor şi precipitaţiilor rezultă că vremea a înregistrat în următoarele două decenii unele devieri la prima vedere
minore. În realitate însă, abaterile înregistrate determină prezenţa unor fenomene climaterice extreme, desta-
bilizând astfel funcţionalitatea normală a vegetaţiei din teritoriul studiat.
4. Cunoaşterea, monitorizarea şi aplicarea unor metodologii de management complex bazat pe studii şi
observaţii cu privire la reacţia plantelor la diferite variaţii ale indicilor climaterici ar crea premise esenţiale în
asigurarea dezvoltării comunităţilor vegetale de luncă viabile, reducând la minimum vulnerabilitatea acestor
ecosisteme faţă de tendinţele climei.
Bibliografie:
1. Baza de date climatice cu rezoluţia de 0,5o x 0,5
o CRU2 (Mitchell şi Jones, 2005).
2. POTOP, V., CONSTANTINOV, T. Manifestarea fenomenelor de uscăciune şi secetă în Republica Moldova.
Chişinău, 2010, p.28.
3. Raportul Naţional de Dezvoltare Umană în Moldova anul 2009-2010, p.42-45.
4. IVAN, V., NISTOR, S., ROŞU, C., ANASTASIU, C. Reconstrucţia ecologică a terenurilor din lunci cu referire
specială la lunca Prutului. În: Revista pădurilor, 2010, nr.1, p.32.
Prezentat la 05.11.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
154
FLORA SINANTROPĂ NECULTIVATĂ A REPUBLICII MOLDOVA
Mihai MÂRZA, Andrei NEGRU
* , Iulian MAMAI
Universitatea de Stat din Moldova *Grădina Botanică (Institut) a AŞM
Flora sinantropă necultivată include 618 specii din 292 genuri şi 63 familii. În calitate de taxoni noi pentru flora
republicii au fost înregistrate şi descrise 32 specii (inclusiv 4 specii noi pentru flora bazinului de nord-vest al Mării
Negre). Componenţa taxonomică a florei sinantrope necultivate a Republicii Moldova este dominată de reprezentaţii
familiilor: Asteraceae, Brassicaceae, Fabaceae, Lamiaceae, Apiaceae, Caryophyllaceae, Scrophulariaceae, Boraginaceae,
care constituie 66,18%.
În baza analizei structurii biotopurilor compoziţiei floristice şi fitocenotice specifice biotopurilor florei sinantrope
necultivate au fost evidenţiate şi descrise 3 grupe de complexe ecologo-floristice (Edaphofodioerodophyton, Conspis-
sophyton, Sciadophyton), 9 ecofitoni (Ruderophyton, Aggeratiophyton, Erodoalluviophyton, Agrophyton, Viaphyton,
Pascuaphyton, Derelictophyton, Confragophyton, Runcatiodrymophyton) şi 4 ecobiogrupe.
Cuvinte-cheie: floră, floră necultivată, floră sinantropă, agrofitocenoză, complexe floristice, adventive.
FALLOW SYNANTHROPE FLORA OF REPUBLIC MOLDOVA
Fallow synanthrope flora of Republic Moldova includes 618 species from 292 genres and 63 families. As a new taxa
for flora of Republic Moldova has been registered and described 32 species (including 4 new species for flora of North -
West Black Sea basin). Taxonomy component of the fallow synanthrope flora is dominated by representatives of families:
Asteraceae, Brassicaceae, Fabaceae, Lamiaceae, Apiaceae, Caryophyllaceae, Scrophulariaceae, Boroginaceae - 66,18%
wich they belong to.
Based on the structure analysis of biotopes,floristic and phytocenotic composition specific for biotopes of fallow
synanthrope flora were outlined and described 3 groups of ecologo-floristic complexes (Edaphodioerodophyton,
Conshissophyton, Sciadophyton) 9 ecophyton (Ruderophyton, Aggeratiophyton, Erodoalluviophyton, Agrophyton,
Viaphyton, Pascuaphyton, Derelietophyton, Confragophyton, Runcatiodrymophyton) and 4 ecobiogroups.
Keywords: flora, fallow, synanthrope, agrophytocenose, floristic complex, adventive.
Introducere
În ultimii 300 de ani, mai cu seamă în a doua jumătate a secolului XX, activitatea umană a provocat
schimbări neprevăzute în structura florei şi vegetaţiei. Pe suprafeţe vaste vegetaţia spontană a fost substituită
de grupări vegetale parţial sau total formate din specii cultivate şi necultivate. Răspândirea excesivă a unui
şir de specii sinantrope necultivate exercită asupra florei sălbatice o acţiune negativă considerabilă sau chiar
schimbă traiectoria dezvoltării ei în unele regiuni ale Terrei.
Problema speciilor sinantrope necultivate, inclusiv a celor invazive, a fost şi este pusă în discuţie la diverse
forumuri internaţionale, precum Conferinţa ONU, la care a fost abordată problema speciilor alohtone, precum
şi la un şir de forumuri şi seminare specializate consacrate fitoinvaziei [1,2,7,19,20]. Concluzia principală
formulată la aceste forumuri este că problema în cauză poate fi soluţionată numai la nivel internaţional, întru-
cât expansiunea speciilor sinantrope necultivate nu are frontiere, iar activităţile pe plan local, de regulă, nu se
finalizează cu rezultate pozitive.
În scopul elaborării atât a metodelor de prevenire a răspândirii speciilor dăunătoare, cât şi a metodelor de
combatere a acestui grup de plante a apărut necesitatea acută de a obţine informaţii despre particularităţile
biologice şi ecologice ale acestor specii. Studierea florei sinantrope necultivate, care se dezvoltă în concor-
danţă cu condiţiile pedoclimatice şi în virtutea interacţiunii complexe a diferiţilor factori, poate să ne ofere
sugestii valoroase cu privire la schimbarea climei ce are loc sub influenţa presingului antropic în regiuni luate
separat.
Materiale referitoare la caracterizarea stării actuale a florei studiate şi la prognoza schimbărilor posibile în
viitor sunt necesare şi pentru înţelegerea corectă a istoriei dezvoltării florei regionale, pentru aprecierea situaţiei
ecologice, precum şi pentru elaborarea concepţiei de utilizare raţională şi de ocrotire a resurselor vegetale.
În ultimii 180 de ani s-au efectuat cercetări de mare valoare privind nu doar flora, dar şi formaţiile natura-
le vegetale, cu includerea în componenţa lor a elementului sinantrop necultivat. Lucrările publicate conţin, în

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
155
fond, date fragmentare despre pătrunderea în acest teritoriu a unor plante adventive şi rolul lor în modificarea
structurii şi funcţionalităţii covorului vegetal. Speciile de plante-buruiene descrise până în prezent cuprind
numai cca 30% din numărul total de specii sinantrope necultivate şi nu oglindesc pe deplin situaţia actuală
reală.
Material şi metode
În calitate de obiect de studiu au servit speciile de plante şi comunităţile vegetale sinantrope necultivate ce
habitează în toate tipurile de ecosisteme terestre din Republica Moldova. Suportul metodologic şi teoretico-
ştiinţific reuneşte îndrumările ce ţin de organizarea şi realizarea cercetărilor floristice şi geobotanice expuse
în lucrările fundamentale consacrate plantelor sinantropice necultivate din bazinul de nord-vest al Mării
Negre [3,4,6,8,18,21-27,29,47,48,52,53,55].
Rezultate şi discuţii
Apariţia şi formarea florei sinantrope necultivate
Problema ce ţine de acţiunea omului asupra covorului vegetal în procesul de gospodarire a preocupat sa-
vanţii din diferite ţări din timpurile cele mai vechi. Dar numai nu demult au început cu o anumită fundamen-
talitate să fie studiate plantele sinantrope necultivate din punct de vedere fitogeografic, sistematic şi biologic.
Studii importante privnd plantele sinantrope cultivate şi necultivate în epoca neolitică pe teritoriul republicii
au efectuat Н.Кузминова [33] şi З.Янушевич [56-58]. I.Morariu [9] subliniază pregnant importanţa deosebită a migraţiilor de popoare pentru expansiunea plan-
telor sinantrope cultivate şi necultivate. Autorul presupune că încă din holocen s-au colonizat multe dintre plantele antropofile. Un factor esenţial, care a contribuit la importarea plantelor sinantrope cultivate şi necul-
tivate, au fost năvălirile şi migraţiunile nomazilor (goţii, hunii, slavii, bulgarii, ungurii, tătarii, turcii, grecii). Din epoca migraţiei popoarelor estice se prea poate că au apărut o serie de specii vest-pontice şi irano-turanice,
ca: Artemisia annua, Ceratocarpus arenarius, Euphorbia steposa, Kochia latifolia, K. prostrata, Xanthium strumarium etc.
De năvălirile turcilor (sec.XV-XVI) este legată, probabil, apariţia unor specii sudice, ca: Cardaria draba, Centaurea iberica, Cynanchum acutum, Helitropium europaeum, Kochia scoparia, Medicago sativa, Salsola
australis, Tribulus terrestris etc. Începând cu sec. XVI, landşafturile agricole devin mai mult sau mai puţin stabile, iar la începutul sec. XVII s-a declanşat o restructurare mai radicală a zonei de stepă.
În sec. XVII-XIX formarea biotopurilor tipice xerofile în partea de sud a contribuit la migrarea spre nord
a unui şir de specii sinantrope necultivate, în special, mediteraniene. Apar în Europa şi un şir de specii adven-tive americane, care s-au extins rapid datorită reţelelor de comunicaţii, schimburilor de mărfuri, deplasării
trupelor în timpul războaielor etc., iar în sec. XX pătrunderea speciilor adventive s-a intensificat şi mai mult. Analiza căilor actuale de pătrundere a plantelor sinantrope necultivate adventive pe teritoriul R. Moldova
ne arată că cele mai multe specii importate în ultimii 30 de ani apar la început în zona de stepă şi de litoral a Mării Negre. Un rol semnificativ în importul acestor plante adventive îl joacă Dunărea. Д.Дубына şi В.Про-
топопова [30,31] identifică în cursul inferior al acestui fluviu 11 specii noi pentru flora Basarabiei şi 3 specii noi pentru flora fostei URSS.
Principalele etape de formare şi dezvoltare a florei sinantrope necultivate a Republicii Moldova, procedeele de importare, căile şi etapele de răspândire, direcţiile şi viteza de migraţie sunt descrise detaliat într-o serie
de lucrări [5,10-17,34,35,37-46,51,54]. Analiza modalităţilor de răspândire a plantelor sinantrope necultivate ne-a dat posibilitatea de a evidenţia
particularitatea lor destul de pronunţată ca plante polihore. Un rol destul de important în extinderea arealelor lor secundare îl joacă adaptările lor de răspândire naturală, care efectiv se îmbină cu cele antropohore. Ca mărturisire serveşte, pe de o parte, modificarea compoziţiei învelişului vegetal până la deplina dispariţie a unor specii autohtone, iar, pe de altă parte – expansiunea şi invadarea biotopurilor antropizate de către spe-ciile alohtone. Deci, este evident faptul că flora sinantropă necultivată constituie nu doar unul dintre cele mai tinere elemente ale florei republicii, dar şi un element cu influenţă vădită asupra structurii şi funcţionării lumii vegetale regionale.
În procesul de formare a arealelor celor mai multe specii vast răspândite pot fi schiţate trei etape: formarea
coloniilor difuze, îndeosebi pe calea speirohoriei, apoi migrarea agestohoră în direcţii radiale, concentrarea arealului pe calea interacţiunii complexe (antropohore şi naturale), răspândirea sub acţiunea dirijată a presin-
gului antropic şi, în sfârşit, extinderea rapidă a arealului într-o direcţie bine determinată.
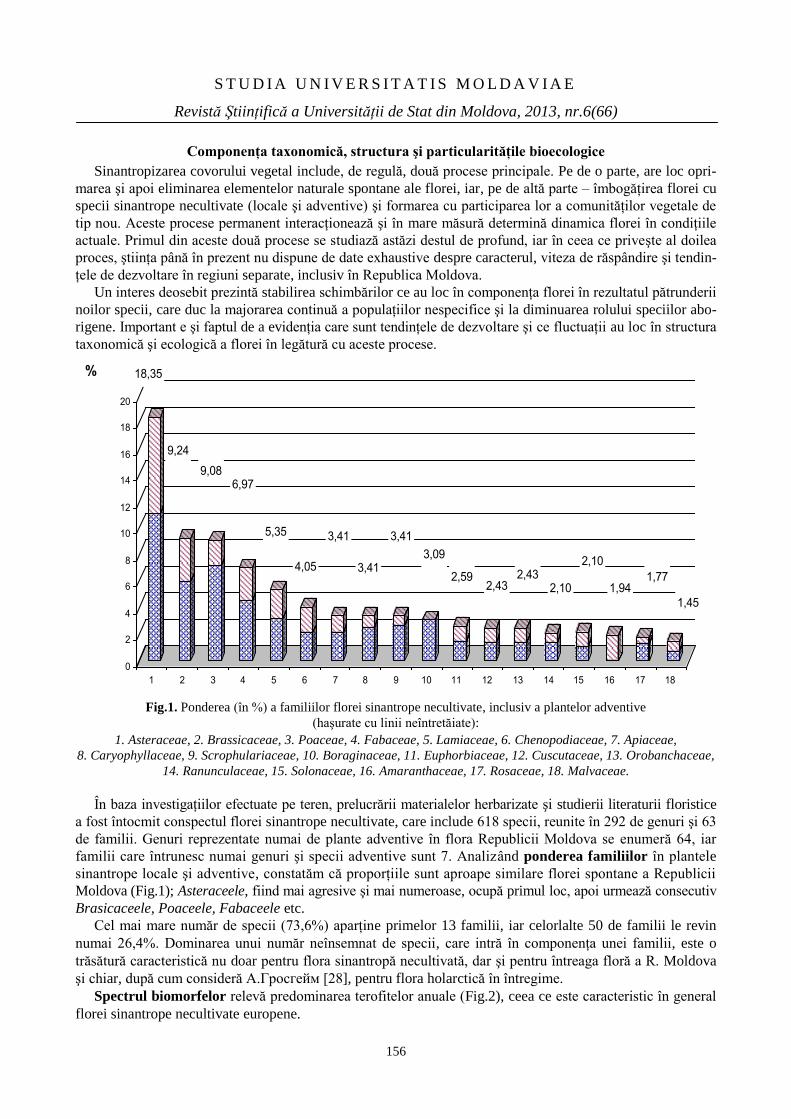
S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
156
Componenţa taxonomică, structura şi particularităţile bioecologice
Sinantropizarea covorului vegetal include, de regulă, două procese principale. Pe de o parte, are loc opri-
marea şi apoi eliminarea elementelor naturale spontane ale florei, iar, pe de altă parte – îmbogăţirea florei cu
specii sinantrope necultivate (locale şi adventive) şi formarea cu participarea lor a comunităţilor vegetale de
tip nou. Aceste procese permanent interacţionează şi în mare măsură determină dinamica florei în condiţiile
actuale. Primul din aceste două procese se studiază astăzi destul de profund, iar în ceea ce priveşte al doilea
proces, ştiinţa până în prezent nu dispune de date exhaustive despre caracterul, viteza de răspândire şi tendin-
ţele de dezvoltare în regiuni separate, inclusiv în Republica Moldova.
Un interes deosebit prezintă stabilirea schimbărilor ce au loc în componenţa florei în rezultatul pătrunderii
noilor specii, care duc la majorarea continuă a populaţiilor nespecifice şi la diminuarea rolului speciilor abo-
rigene. Important e şi faptul de a evidenţia care sunt tendinţele de dezvoltare şi ce fluctuaţii au loc în structura
taxonomică şi ecologică a florei în legătură cu aceste procese.
18,35
9,24
9,086,97
5,35
4,05
3,41
3,41
3,41
3,09
2,592,43
2,432,10
2,10
1,941,77
1,45
0
2
4
6
8
10
12
14
16
18
20
%
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Fig.1. Ponderea (în %) a familiilor florei sinantrope necultivate, inclusiv a plantelor adventive
(haşurate cu linii neîntretăiate):
1. Asteraceae, 2. Brassicaceae, 3. Poaceae, 4. Fabaceae, 5. Lamiaceae, 6. Chenopodiaceae, 7. Apiaceae,
8. Caryophyllaceae, 9. Scrophulariaceae, 10. Boraginaceae, 11. Euphorbiaceae, 12. Cuscutaceae, 13. Orobanchaceae,
14. Ranunculaceae, 15. Solonaceae, 16. Amaranthaceae, 17. Rosaceae, 18. Malvaceae.
În baza investigaţiilor efectuate pe teren, prelucrării materialelor herbarizate şi studierii literaturii floristice
a fost întocmit conspectul florei sinantrope necultivate, care include 618 specii, reunite în 292 de genuri şi 63
de familii. Genuri reprezentate numai de plante adventive în flora Republicii Moldova se enumeră 64, iar
familii care întrunesc numai genuri şi specii adventive sunt 7. Analizând ponderea familiilor în plantele
sinantrope locale şi adventive, constatăm că proporţiile sunt aproape similare florei spontane a Republicii
Moldova (Fig.1); Asteraceele, fiind mai agresive şi mai numeroase, ocupă primul loc, apoi urmează consecutiv
Brasicaceele, Poaceele, Fabaceele etc.
Cel mai mare număr de specii (73,6%) aparţine primelor 13 familii, iar celorlalte 50 de familii le revin
numai 26,4%. Dominarea unui număr neînsemnat de specii, care intră în componenţa unei familii, este o
trăsătură caracteristică nu doar pentru flora sinantropă necultivată, dar şi pentru întreaga floră a R. Moldova
şi chiar, după cum consideră A.Гросгейм [28], pentru flora holarctică în întregime.
Spectrul biomorfelor relevă predominarea terofitelor anuale (Fig.2), ceea ce este caracteristic în general
florei sinantrope necultivate europene.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
157
A 33,29%
C 35,82%
B 29,43%
D 1,46%
54,7
9,0
24,7
3,9 1,5 1,8 0,6 0,8 0,3 2,7
0
10
20
30
40
50
60%
1 2 3 4 5 6 7 8 9 10
1. Terofite anuale – 54,7;
2. Terofite bianuale – 9,0;
3. Hemicriptofite – 24,7;
4. Geofite – 3,9;
5. Microfanerofite – 1,5;
6. Camefite – 1,8;
7. Megafanerofite – 0,6;
8. Helohidatofite – 0,8
9. Nanofanerofite – 0,3
10. Nedeterminate – 2,7.
Fig.2. Spectrul biomorfelor.
Gruparea speciilor pe elemente fitogeografice în prezentare numerică şi procentuală (Tab.1) indică parti-
ciparea diferitelor categorii de elemente la alcătuirea florei sinantrope necultivate a republicii.
Tabelul 1
Ponderea elementelor fitogeografice în cormoflora sinantropă necultivată a R.Moldova
Elementele nordice şi central-eurasiatice constituie 33,29%, iar cele sudice participă în proporţie de 29,43%
(Fig.3). Aceasta se explică prin poziţia teritoriului R. Moldova la contactul a trei zone biogeografice şi prin
încadrarea ei în subregiunea Eurosiberiană, regiunea Central-Europeană. Este destul de înalt procentul ele-
mentelor polihore (35,82%) – particularitate specifică în general florii sinantrope necultivate.
Fig.3. Ponderea elementelor fitogeografice:
A – nordice; B – sudice; C – polihore; D – elemente nedeterminate.
El.
flor.
Dintre care: Nr. În total El. flor. Dintre care: Nr. În total
Eua. Eua.
Eua.(Medit.)
Eua.(cont.)
113
76
50
239
Pont.-Pan.-
Medit.
Pont.-Medit.
Pont.-Pan.
Pont.-Balc.
Pont.-Pan.-Medit
Medit.
Pont.-Pan.-Balc.
6
17
4
3
1
1
32
Medit. 36 36 Balc. Balc.
Balc.-Anat.
3
1
4
Cosm. 30 30 Alt.-Medit. 3 3
Eur. Eur.
Eur.-Medit.
Eur.(cont.)
18
11
7
36
Carp.-Balc. Carp.-Pan.
Carp.-Balc.-Cauc.
1
1
2
Circ. Circ.
Circ.(Bor.)
Circ.(Bor.)
Balc.
5
7
1
13
Adv. 191 191
Euc. Euc.
Euc.(Medit.)
Euc.(Balc.)
5
17
1
23
Nedetermi-
nate
9 9

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
158
Pentru o ilustrare cât mai vădită a realităţii în ceea ce priveşte specificul plantelor sinantrope necultivate din R. Moldova au fost întocmite (pe lângă tabelele spectrelor biomorfelor şi a elementelor fitogeografice) şi
spectrele indicilor ecologici: umiditate, temperatură şi reacţie a solului (Fig.4-6).
1. Xerofite – 8,1%, U1 - U1.5
2. Xeromezofite – 40,9%, U2 - U2.5
3. Mezofite – 28,3%, U3 - U3.5
4. Mezohidrofite–ultrahidrofite – 5,6%, U4 - U6
5. Amfitolerante – 3,6%,U0
6. Nedeterminate – 13,5%
Fig.4. Spectrul indicilor de umiditate (U).
0
10
20
30
40
50
1 2 3 4
1. Mezoterme – 46,0%, T3 - T3,5
2. Moderat termofile – 28,2%, T4 - T4,5
3. Amfitolerante – 9,0%, T0
4. Nedetrminate – 16,8%
Fig.5. Spectrul indicilor de temperatură (T).
0
10
20
30
40
50
60
1 2 3 4 5
1. Acidofile – 0,6%, R1 - R2
2. Slab acidoneutrofile – 50,6%, R3 - R4.5
3. Neutrobazifile – 1,6%, R5
4. Eurionice – 33,1%, R0
5. Nedeterminate – 14,1%
Fig.6. Spectrul indicilor de reacţie a solului (R).
Analiza indicilor ecologici pune în evidenţă ponderea înaltă de participare a xeromezofitelor (40,9%) şi a mezofitelor (28,3%), a speciilor mezoterme (46,0%) şi a celor moderat termofile (28,2%), a plantelor slab acidoneutrofile (50,6%) şi a celor eurionice (33,1%), ceea ce reflectă condiţiile edafoclimatice ale teritoriului inclus în studiu.
În felul acesta, rezultatele obţinute denotă originea şi evoluţia florei sinantrope necultivate a regiunii de nord-vest a Mării Negre în general, precum şi a florei R. Moldova în special.
Apartenenţa ecosistemică a speciilor sinantrope În dependenţă de ecosistemul sau habitatul preferat, plantele sinantrope necultivate se grupează în urmă-
toarele categorii şi subcategorii:
A. Specifice ecosistemelor cerealiere de păioase. Particularităţile biologice ale cerealelor păioase şi
tehnologia cultivării lor condiţionează apariţia şi existenţa unui şir de specii şi asociaţii de plante sinantrope
necultivate, care au devenit specifice pentru aceste tipuri de biotop. Primăvara, aspectul caracteristic în cultu-
rile de cereale este format atât de speciile sinantrope necultivate care germinează toamna, iernează sub formă
de plantule, cât şi de speciile care germinează primăvara timpuriu şi înfloresc tot primăvara. Majoritatea sunt
plante efemere de talie scundă şi nu concurează între ele decât în mică măsură cu plantele de cultură. Aspectul
de vară, caracteristic începutului lunii iunie, este determinat de speciile care germinează toamnă sau primă-
vara timpuriu şi dispun de un ritm rapid de creştere-dezvoltare în prima jumătate a lunii iunie, maturizându-se
până la recoltarea cerealelor păioase sau în timpul recoltării, diminuând evident roada.
0
10
20
30
40
50
1 2 3 4 5 6

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
159
Cercetările întreprinse arată că la îmburuienarea cerealelor păioase participă cca 194 de specii (Tab.2),
dintre care 113 sunt anuale, 33 perene, 14 anuale-hibernante, 12 anuale-bianuale, 9 – bianuale. Ca plante
sinantrope necultivate invazive pot fi considerate cca 19 specii.
B. Specifice ecosistemelor de culturi prăşitoare. Tehnologia prelucrării şi particularităţile biologice ale
culturilor de prăşitoare condiţionează existenţa unui număr de specii şi asociaţii sinantrope necultivate mai
mic decât în culturile de păioase. Pe aceste suprafeţe se dezvoltă plante sinantrope necultivate, care iernează
sub formă de plantule şi cele care germinează primăvara târziu. Din această cauză ele sunt întodeauna mai
puternic dezvoltate decât în cerealele păioase.
După efectuarea ultimelor lucrări agrotehnice în culturile de prăşitoare apar plantele sinantrope necultivate
termofile: Amaranthus hybridus, A. retroflexus, Chenopodium album, Echinocloa crusgali, Hibiscus trionum,
Xanthium californicum etc.
Datorită tehnologiilor similare aplicate în culturile de prăşitoare, speciile sinantrope necultivate sunt cam
aceleaşi, cu excepţia culturilor de floarea-soarelui, sfeclă de zahăr, cartof şi chiar de tutun, unde, pe lângă spe-
ciile sinantrope necultivate comune pentru toate prăşitoarele, se mai dezvoltă şi unele plante specifice. În con-
tinuare prezentăm separat lista speciilor sinantrope necultivate, care însoţesc principalele culturi de prăşitoare.
a) Culturi de porumb. Dintre speciile care însoţesc culturile de porumb numai 12 sunt mai răspândite:
Amaranthus hybridus, A. retroflexus, Chenopodium album, Echinocholoa crusgali, Setaria glauca, S. viridus,
Cirsium arvense, Elytrigia repens, Polygonum hydropiper, Sinapis arvensis, Convolvulus arvensis, Xanthium
californicum.
În total au fost evidenţiate 138 de specii sinantrope necultivate (Tab.2). Dominante sunt plantele anuale
(87 de specii).
b) Culturi de floarea-soarelui. Din cele 143 de specii, care însoţesc culturile de floarea-soarelui, cca 16
sunt invazive (Tab.2). Predomină plantele sinantrope necultivate anuale şi bienale. Pe unele suprafeţe pagube
foarte mari produce culturii de floarea-soarelui specia invazivă Cyclachaena xanthifolia.
c) Culturi de sfeclă de zahăr. Din toate culturile prăşitoare sfecla de zahăr impune cele mai grele probleme
de combatere a plantelor sinantrope necultivate.
În urma cercetărilor efectuate s-a constatat că numărul de specii de plante sinantrope necultivate partici-
pante de la an la an, în urma folosirii erbicidelor, rămâne aproape neschimbat, dar pentru majoritatea specii-
lor s-a mărit densitatea plantelor. S-au înregistrat şi unele diminuări ale speciilor: Raphanus raphanistrum,
Centaurea cyanus, Consolida regalis, Capsella bursa-pastoris. Flora sinantropă necultivată din aceste plan-
taţii este reprezentată printr-un număr de 163 de specii (Tab.2). Predomină terofitele anuale (103 sp.), apoi
urmează hemicriptofitele (15 sp.) şi terofitele anuale–bianuale (13 sp.). Ca specii invazive pot fi considerate
14 specii.
d) Culturi de cartof. Cartoful se cultivă de obicei pe terenurile din apropierea locuinţelor cu soluri fertile,
ceea ce favorizează dezvoltarea sinantropelor necultivate azotofile, ca: Amaranthus sp., Chenopodium sp.,
Cyclachaena xanthiifolia, Xanthium sp. etc. Din totalul de 121 de specii semnalate în culturile de cartof 89
sunt anuale-bianuale, iar 32 – perene. După indicii de reacţie a solului majoritatea speciilor sunt eurionice
(57 sp.), urmate de cele slab acidoneutrofile (33 sp.). Se consideră invazive cca 18 specii (Tab.2).
C. Specifice ecosistemelor legumicole. Flora sinantropă necultivată din aceste tipuri de ecosisteme este
destul de bogată şi enumeră cca 205 specii (Tab.2). Predomină speciile anuale (111 sp.), iar după indicii de
umiditate – mezofite (47 sp.) şi xeromezofite (47 sp.). Ca plante invazive au fost înregistrate 19 specii.
Pentru prima dată în flora R. Moldova în culturile de prăşitoare au fost depistate două specii: Amaranthus
hypochondriacus şi Neslia paniculata şi înregistrate staţiuni noi de creştere pentru speciile: Dracocephalum
moldavica, D. thymiflorum, Euphorbia dentata, Galihsoga ciliata, Panicum milliaceum, Ranunculus arvensis,
Senecio vulgaris, Setaria italica, Spergula arvensis, Zygophyllum fabago etc.
În baza observaţiilor de teren putem conchide că:
în unele culturi de prăşitoare, cum sunt floarea-soarelui, tutunul, cartoful, destul de frecvente sunt spe-
ciile de plante sinantrope necultivate parazite (Orabanche, Cuscuta);
pe solurile aluvionare se întâlnesc mai frecvent Galinsaga ciliata, G. parviflora, Portulaca oleracea,
Stachys annua, Artemisia annua, Echinochloa crusgali;
în culturile de prăşitoare în ultimii ani a crescut considerabil abundenţa speciilor de plante sinantrope
necultivate perene.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
160
D. Specifice ecosistemelor lucerniere. În următorii 2-4 ani după semănat lucerna acoperă bine solul,
elimină majoritatea speciilor sinantrope necultivate, iar cele perene, care nu pot fi înlăturate, sunt stârpite
treptat prin operaţia de recoltare a plantei, încât practic lucerna nu ridică problema de combatere a plantelor
sinantrope necultivate. Din observaţiile pe care le-am efectuat putem afirma că lucerna poate constitui chiar
un mijloc de luptă contra plantelor sinantrope necultivate, cu excepţia unor specii (Elytrigia repens, Cynodon
dactylon, Taraxacum officinale, Rapistrum petrenne). Începând cu a V-VI-a perioadă de vegetare, puterea de
regenerare a lăstarilor de lucernă descreşte, o parte din plante dispar, iar locul lor este ocupat de plantele sinan-
trope necultivate. Flora sinantropă necultivată din lucerniere include cca 218 specii (Tab.2). Dintre acestea,
ca forme biologice, 50,2% sunt terofite anuale, 17,2% – hemicriptofite, 11,6% – terofite anuale-bianuale şi
6,0% – bianuale.
E. Specifice ecosistemelor viticole. Imediat după topirea zăpezii se dezvoltă plantele care au iernat sub
formă de plante verzi (Capsella bursa–pastoris, Chondrilla juncea, Erophila verna, Erodium cicutarium,
Fumaria sp., Glechoma hederacea, Lamium amplexicaule şi a.) şi unele specii vernale, ca: Allium rotundum,
A. vineale, Muscari neglectum etc.
În aceste ecosisteme au fost indentificate 382 de specii (Tab.2), care fac parte din 209 de genuri şi 52 de
familii. În spectrul fitogeografic rolul edificator revine elementelor de origine sudică (36,9%), la care se mai
adaugă un număr însemnat de elemente eurasiatice (36,1%), cosmopolite (7,8%) şi adventive (10,9%). Se
constată predominarea terofitelor anuale şi bianuale (58,9%).
F. Specifice ecosistemelor pomicole. Din cele 439 de specii de plante sinantrope necultivate identificate
în plantaţiile pomicole (Tab.2) 155 sunt specii care însoţesc şi culturile de cereale păioase, 198 însoţesc cul-
turile de prăşitoare, iar 199 de specii vegetează în lucerniere. Analiza florei necultivate relevă predominarea
terofitelor anuale şi bianuale (59,2%). Spre deosebire de celelalte tipuri de ecosisteme, în plantaţiile pomicole
în proporţie apreciabilă participă hemicriptofitele (26,4%).
G. Specifice ecosistemelor naturale
a) Pajişti şi fâneţe. Din cele 226 specii (Tab.2) identificate în pajişti cca 37 invadează puternic păşunile,
impunând luarea unor măsuri de combatere. Pe pajiştele de coastă, plantele sinantrope, care impun stârpirea
lor cât mai rapidă, aparţin genurilor: Carduus, Cirsium, Euphorbia, Onopordum, Xanthium şi a.
Pe pajiştile utilizate ca fâneţe mai agresive sunt: Cardus acanthoides, C. nutans, Centaurea difusa, Cirsium
arvense, C. setosum, Erigeron canadensis, Euphorbia sp., Onopordum acanthium, iar pe pajiştile de luncă cu
depuneri de aluviuni – Cyclachaena xanthiifolia, Erigeron annus, E. canadensis, Xanthium sp.
Pe un teren folosit ca pajişte a fost depistată pentru prima dată în flora R. Moldova Setaria viridis, subsp.
pycnocoma (Steud.) Tzvel.
În spectrul fitogeografic domină speciile Eurasiatice – 23,5%, Eurocentrale (cont.) – 12,4% şi Meditera-
neene – 6,2%. Este destul de înalt procentul speciilor cosmopolite (9,6%) şi adventive (15,2%), ceea ce denotă
gradul înalt de sintropizare a păşunilor şi fâneţelor.
b) Păduri. Gradul mediu de împădurire este de 9,6%, în zona de nord – de 7,2%, în zona de centru – 13,5%
şi în cea de sud – 6,7%. În diferite tipuri de păduri şi tăieturi plantele sinantrope necultivate au o instalare
diferită ce depinde de gradul de acoperire a arboretului, de umiditatea şi compoziţia solului, de nivelul de
degradare şi de gradul de fragmentare a pădurilor.
Pe terenurile silvice au fost depistate 3 specii de plante sinantrope necultivate neindicate până în prezent
pentru flora R. Moldova: Hesperis sibirica, Potentilla bifurca, Galeopsis pubescens, iar pentru 2 specii –
indicate staţiunii noi de creştere: Centaurea iberica şi Ambrosia artemisiifolia.
În comunităţile silvice au fost edificate 251 specii de plante sinantrope necultivate (Tab.2), dintre care
146 sunt anuale-bianuale, 11 anuale-bianuale – perene, 89 – perene, 2 arbori şi 3 arbuşti.
c) Viroage. Materialul transportat în timpul ploilor torenţiale se depune în conurile de dejecţie sau la con-
fluenţa râurilor şi pâraielor, formând viroage de diferite dimensiuni. Pe ele se instalează specii care totuşi nu
reuşesc să formeze un covor verde continuu, deoarece la o nouă ploaie torenţială apele depozitează straturi
noi. Studiul de teren mărturiseşte că speciile care vegetează pe aceste terenuri prezintă în anumită măsură
asemănări cu flora terenurilor de unde a fost importat materialul erodat împreună cu rămăşiţele de plante. În
total pe aceste suprafeţe au fost depistate 312 specii de plante sinantrope necultivate (Tab.2).
H. Specifice ecosistemelor degradate. Aceste terenuri sunt, de regulă, localităţi cu caracter urban: tere-
nurile virane şi cele cu construcţii nefinisate, depozitele de deşeuri, marginea căilor de comunicaţie. Pe ele se
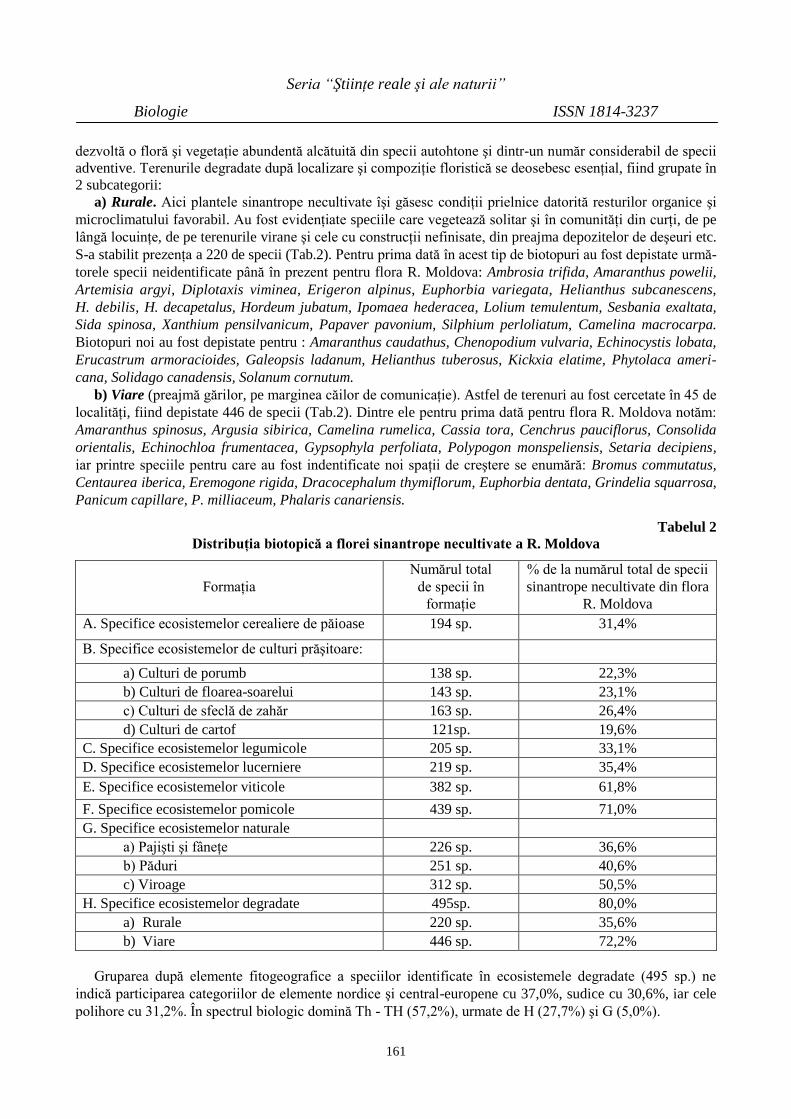
Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
161
dezvoltă o floră şi vegetaţie abundentă alcătuită din specii autohtone şi dintr-un număr considerabil de specii
adventive. Terenurile degradate după localizare şi compoziţie floristică se deosebesc esenţial, fiind grupate în
2 subcategorii:
a) Rurale. Aici plantele sinantrope necultivate îşi găsesc condiţii prielnice datorită resturilor organice şi
microclimatului favorabil. Au fost evidenţiate speciile care vegetează solitar şi în comunităţi din curţi, de pe
lângă locuinţe, de pe terenurile virane şi cele cu construcţii nefinisate, din preajma depozitelor de deşeuri etc.
S-a stabilit prezenţa a 220 de specii (Tab.2). Pentru prima dată în acest tip de biotopuri au fost depistate urmă-
torele specii neidentificate până în prezent pentru flora R. Moldova: Ambrosia trifida, Amaranthus powelii,
Artemisia argyi, Diplotaxis viminea, Erigeron alpinus, Euphorbia variegata, Helianthus subcanescens,
H. debilis, H. decapetalus, Hordeum jubatum, Ipomaea hederacea, Lolium temulentum, Sesbania exaltata,
Sida spinosa, Xanthium pensilvanicum, Papaver pavonium, Silphium perloliatum, Camelina macrocarpa.
Biotopuri noi au fost depistate pentru : Amaranthus caudathus, Chenopodium vulvaria, Echinocystis lobata,
Erucastrum armoracioides, Galeopsis ladanum, Helianthus tuberosus, Kickxia elatime, Phytolaca ameri-
cana, Solidago canadensis, Solanum cornutum.
b) Viare (preajmă gărilor, pe marginea căilor de comunicaţie). Astfel de terenuri au fost cercetate în 45 de
localităţi, fiind depistate 446 de specii (Tab.2). Dintre ele pentru prima dată pentru flora R. Moldova notăm:
Amaranthus spinosus, Argusia sibirica, Camelina rumelica, Cassia tora, Cenchrus pauciflorus, Consolida
orientalis, Echinochloa frumentacea, Gypsophyla perfoliata, Polypogon monspeliensis, Setaria decipiens,
iar printre speciile pentru care au fost indentificate noi spaţii de creştere se enumără: Bromus commutatus,
Centaurea iberica, Eremogone rigida, Dracocephalum thymiflorum, Euphorbia dentata, Grindelia squarrosa,
Panicum capillare, P. milliaceum, Phalaris canariensis.
Tabelul 2
Distribuţia biotopică a florei sinantrope necultivate a R. Moldova
Formaţia
Numărul total
de specii în
formaţie
% de la numărul total de specii
sinantrope necultivate din flora
R. Moldova
A. Specifice ecosistemelor cerealiere de păioase 194 sp. 31,4%
B. Specifice ecosistemelor de culturi prăşitoare:
a) Culturi de porumb 138 sp. 22,3%
b) Culturi de floarea-soarelui 143 sp. 23,1%
c) Culturi de sfeclă de zahăr 163 sp. 26,4%
d) Culturi de cartof 121sp. 19,6%
C. Specifice ecosistemelor legumicole 205 sp. 33,1%
D. Specifice ecosistemelor lucerniere 219 sp. 35,4%
E. Specifice ecosistemelor viticole 382 sp. 61,8%
F. Specifice ecosistemelor pomicole 439 sp. 71,0%
G. Specifice ecosistemelor naturale
a) Pajişti şi fâneţe 226 sp. 36,6%
b) Păduri 251 sp. 40,6%
c) Viroage 312 sp. 50,5%
H. Specifice ecosistemelor degradate 495sp. 80,0%
a) Rurale 220 sp. 35,6%
b) Viare 446 sp. 72,2%
Gruparea după elemente fitogeografice a speciilor identificate în ecosistemele degradate (495 sp.) ne
indică participarea categoriilor de elemente nordice şi central-europene cu 37,0%, sudice cu 30,6%, iar cele
polihore cu 31,2%. În spectrul biologic domină Th - TH (57,2%), urmate de H (27,7%) şi G (5,0%).

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
162
Analiza cerinţelor speciilor sinantrope necultivate faţă de factorii ecologici principali (U,T,R,) indică pre-
dominarea plantelor xeromezofile (39,9%), mezofile (27,3%) şi amfitolerante (14,5%); mezoterme (42,6%)
şi moderat-termofile (29,6%); slab acido-neutrofile (35,5%) şi eurionice (34,3%).
Legăturile florogenetice şi particularităţile zonal-regionale de răspândire
Studierea, evidenţierea şi analiza legăturilor florogenetice ne-a permis, în primul rând, să identificăm în
componenţa florei sinantrope necultivate a R. Moldova două fracţiuni principale: aborigenă – 429 şi cea
adventivă – 189 specii, iar, în al doilea rând, să stabilim regiunile floristice din contul cărora s-a îmbogăţit şi
se îmbogăţeşte flora sinantropă necultivată aflată în studiu. Raportul numeric al speciilor din aceste două
fracţiuni (aborigenă şi adventivă) formate în procesul dezvoltării istorice este, după părerea noastră, o impor-
tantă trăsătură a florei sinantrope necultivate a oricărei regiuni şi caracterizează nivelul ei de sinantropizare.
Este cunoscut faptul că acţiunea intensivă gospodărească a omului duce la aridizarea climatului local şi,
în legătură cu aceasta, favorizează extinderea zonelor aride, creând pe unele teritorii ale Terrei, după cum
afirmă M.Попов [49], „primejdia devastării antropice”.
Conform cercetărilor efectuate, s-a constatat că repartiţia florei sinantrope necultivate în cele două regiuni
botanico-geografice ale republicii se deosebeşte după numărul şi componenţa speciilor, caracterul lor de răs-
pândire şi rolul lor în învelişul vegetal. În regiunea de silvostepă vegetează cca 90 de specii de plante sinan-
trope necultivate care nu se întâlnesc în cea de stepă, iar în cea de stepă vegetează cca 40 de specii care nu se
întâlnesc în cea de silvostepă. Din numărul total de specii sinantrope necultivate cca 480 (77,6%) sunt răs-
pândite în ambele regiuni, celelalte au o răspândire mai limitată.
Pentru R. Moldova problemele ce ţin de ridicarea aridităţii ecotopurilor sub acţiunea factorului antropic
au o însemnătate primordială. În legătură cu aceasta apare, evident, necesitatea identificării zonelor floristice
şi a regiunilor bioclimatice din care provin principalele grupe de specii ce completează flora sinantropă ne-
cultivată şi, concomitent, care este dinamica, direcţia generală de restructurare a spectrului geoelementelor în
condiţiile noastre. Structura fitogeografică a florei sinantrope necultivate a fost elucidată prin distribuirea
speciilor pe fitohorioane. Rezultatele analizei demonstrează că pe ecotopurile antropice ale R. Moldova se
creează condiţii favorabile pentru stabilirea speciilor regiunilor aride ale Terrei. Un rol de seamă în acest
grup îl ocupă speciile aborigene, care total sau parţial au migrat pe biotopurile antropice. Л.Малышева [36]
presupune că în raioanele de sud ale Europei este posibilă creşterea rolului apofitelor din cauza apartenenţei
sau vecinătăţii acestor ţări cu zona agriculturii străvechi, care include Asia Orientală şi Regiunea Meditera-
neană. Însă, privitor la corelaţia dintre speciile antropohore şi apofite în raioanele de sud şi de nord ale Europei
acest indice rămâne necunoscut. Apofitele din flora R. Moldova au fost divizate de către noi în trei grupe:
Evapofite – speciile care complet sau aproape complet au migrat pe ecotopurile antropice; Hemiapofite –
speciile care activ se răspândesc pe ecotopuri antropice, dar mai menţin o poziţie dură în componenţa florei
locale zonale şi intrazonale; Apofite – specii nestabile, care reprezintă un element antropofob ocazional al
biotopurilor antropice. De obicei, componenţa lor este eterogenă şi specifică pentru florele sinantrope necul-
tivate din diferite regiuni.
Elementul autohton al florei în studiu se caracterizează printr-o abundenţă majoră a speciilor, arealul
cărora corespunde mai mult sau mai puţin cu cel Mediteranean Antic.
Predominarea speciilor arealogic legate de regiunea Mediteraneană Străveche ne dă temei să presupunem
că şi elementul antropofil local poartă caracterul unei flore xerofile. Aceste presupuneri se confirmă de rezul-
tatele analizei fitocenologice, conform cărora majoritatea apofitelor (53%) sunt componente ale comunităţi-
lor xeroterme.
Plantele comunităţilor mai mult sau mai puţin mezofile (de luncă, litoral, silvice şi de tufărişuri) alcătuiesc
43,5%, dar cele mai numeroase dintre grupele amintite sunt speciile de luncă şi de tufărişuri (până la 74%).
Numeric mai puţine sunt elementele de pădure (11,7%). Celelalte specii demult au pierdut legătura cu bioto-
purile naturale: evapofitele ce alcătuiesc 0,4% şi halofitele – 1,9%.
Studiile efectuate ne-au dat posibilitatea nu doar de a clarifica particularităţile de repartiţie a elementului
autohton şi alohton, dar şi de a stabili care este rolul lor în formarea biotopurilor antropice studiate. Pe bioto-
purile antropice predomină elementul autohton, însă procentual numărul de specii adventive creşte spre nord,
iar al apofitelor – spre sud. De menţionat că spre sud relativ se măreşte nu doar numărul de specii aborigene,
dar şi rolul acestui element în formarea florei din biotopurile antropice. Una dintre cauzele care duc la mărirea

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
163
procentului de specii adventive în raioanele de nord ale R. Moldova poate fi considerată sărăcirea relativă a
florei naturale odată cu creşterea xerofilizării.
În baza datelor obţinute concluzionăm că în rezultatul sintropizării flora Moldovei s-a îmbogăţit cu ele-
mente floristice, care genetic şi geografic sunt legate cu teritoriile xerice (xericus) ale Terrei. Intensificarea
regimului xerotermic al biotopurilor antropice, în comparaţie cu cele naturale, provoacă, pe de o parte, for-
marea pe ele a unei pături vegetale după structură şi compoziţie echivalentă cu cea a florei subalpine şi de
semideşerturi din regiunea Mediteraneană Străveche, iar, pe de altă parte, eliminarea speciilor mezofile.
Prin urmare, procesul de sintropizare contribuie la xerofilizarea florei regionale, astfel fiind una dintre
cauzele care împiedică regenerarea naturală a florei spontane.
Pentru o regiune atât de sinantropizată ca R. Moldova o însemnătate primordială capătă, în ansamblu,
conservarea eficientă a învelişului vegetal spontan concomitent cu reconstrucţia ecologică a landşafturilor
naturale.
Structura ecologo-cenologică şi complexele floristice evidenţiate
La etapa actuală flora sinantropă necultivată a R. Moldova este reprezentată printr-o totalitate de elemente
vegetale destul de diverse, ale căror componenţă floristică, durată de existenţă, direcţii de dezvoltare depind
de particularităţile istorice, economice şi ecologice. În prezent, delimitarea tradiţională a speciilor sinantrope
necultivate şi a comunităţilor lor în „ruderale” şi „segetale” creează un şir de confuzii, neclarităţi – fapt con-
diţionat de majorarea eterogenităţii şi diferenţierii terenurilor agricole cauzate de regimul de folosire, meto-
dele de lucrare a solului, caracterul culturilor şi de modul de întreţinere.
Noi susţinem părerea expusă de В.Протопопова [50], potrivit căreia speciile, care preferă solurile afânate
(săpate, arate, erodate, aluviale), urmează să fie atribuite la grupa „eroziofitelor”, iar speciile care preferă so-
lurile bătătorite – la grupa „conspissofitelor”. Desigur, schema prezentată este într-o măsură oarecare conven-
ţională, însă ea este mai adecvată repartiţiei actuale a speciilor care vegetează pe anumite ecotopuri. Tocmai
prin reacţia plantei faţă de caracterul deformaţiei structurii solului se explică repartiţia unor şi aceloraşi specii
pe diferite ecotopuri: gunoişti, rambleuri de cale ferată şi autostrăzi, terenuri lucrate, păduri, lunci şi stepe
degradate, aluviuni, maluri de râuri şi pâraie, pante erodate, culturi de prăşitoare etc. Această structură flo-
ristică specifică anumitului biotop corespunde noţiunii de florocomplex propuse de către M.Kлоков [32].
Astfel, un complex floristic antropic este o totalitate de plante sinantrope necultivate aborigene, adventive şi
„restante”, care supravieţuiesc în comun pe terenurile unde flora aborigenă a fost distrusă şi care se formează
şi se dezvoltă sub acţiunea gospodărească dirijată a omului; ele reprezintă stadii primare de adaptare în comun
a diferitelor elemente ale compexelor floristice, care întru totul diferă între ele şi sunt capabile să se acomo-
deze la condiţiile actuale de existenţă. Odată cu slăbirea sau diminuarea influenţei factorului antropic, care se
manifestă în menţinerea stabilităţii antropofitonului, începe să predomine procesele de demutaţie a covorului
vegetal până la etapa, când în condiţii mai mult sau mai puţin favorabile se apropie de cele naturale.
În rezultatul cercetărilor efectuate, considerăm că flora sinantropă necultivată a R. Moldova poate fi ordo-
nată în trei grupe de complexe, iar în limetele fiecărui complex se pot evidenţia şi ecofitoni subalterni. Unităţile
elementare de clasificare au fost stabilite în funcţie de criteriile generale adoptate de B.Протопопова [50].
În continuare prezentăm schema generală de clasificare şi caracterizarea succintă a complexelor ecologo-
floristice ale plantelor sinantropice necultivate a R. Moldova.
I. EDAPHOFODIOERODOPHYTON II. CONSPISSOPHYTON
1. Ruderophyton 1. Viaphyton
2. Aggeratiophyton 2. Pascuaptyton
a) Faciopetrophyton 3. Derelictophyton
3. Erodoalluviophyton III. SCYADOPHYTON
4. Agrophyton 1. Confragophyton
a) Interariophyton 2. Runcatiodrymophyton
b) Segetophyton
c) Faciopratophyton
I. EDAPHOFODIOERODOPHYTON – grup ecologo-floristic care reuneşte complexele floristice ale
terenurilor complet transformate. Numai o parte a componenţilor abiotici şi biotici ai ecotopurilor pe care se
formează astfel de complexe de specii îşi păstrează o independenţă relativă, celelalte sunt totalmente contro-

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
164
late de către om. Această grupă, din punct de vedere floristic, este cea mai bogată. În componenţa ei sunt in-
cluse un număr considerabil de plante adventive, iar ca forme vitale predomină cele anuale şi bianuale.
1. Ruderophyton – reuneşte speciile caracteristice ecotopurilor cu soluri afânate, redzine sau soluri cu canti-
tăţi ridicate de nitraţi. Speciile caracteristice: Amaranthus retroflexus, Atriplex nitens, Capsella bursa-pastoris,
Chenopodium album, Ch. hybridum, Datura stramonium, Galium molugo, Hyscyamus niger, Leonurus
quinquelobatus, Solanum nigrum, Utrica dioica, U. urens etc. Numărul mare de arheofite, ce alcătuieşte
nucleul acestui ecofiton, atestă originea străveche a florei în studiu.
2. Aggeratiophyton – se dezvoltă pe rambleuri lipsite de vegetaţie naturală, de-a lungul căilor de comuni-
caţie, în locuri virane ale construcţiilor curent începute, pe margini de ogoare, în cariere, pe malurile canale-
lor şi lacurilor. Din punct de vedere floristic predomină explorenţii şi prezintă etapa iniţială de dezvoltare a
vegetaţiei de pârloagă, cu legături instabile dintre specii. Nucleul floristic este alcătuit din specii adventive
caracteristice regiunilor aride, apofite cu areale vaste, evapofite, care au pierdut legăturile cu comunităţile
vegetale naturale, şi din specii de stepă, alelopatic şi ecotopologic active cu arealul de tip progresiv. Caracte-
ristice sunt: Atriplex tatarica, Cenchrus pauciflorus, Centaurea diffusa, Cyclachaena xanthiifolia, Grindelia
squarrosa, Panicum milliaceum, Setaria glauca, Sysimbrium altissium, S. loeselii, Xanthium albinum, X. cali-
fornicum, X.strumarium etc.
O varietate ecologică specifică a acestui complex este Faciopetrophyton, care reuneşte speciile specifice
terasamentelor (rambleurilor) pietroase. Specii caracteristice: Cenchraus pauciflorus, Centaurea diffusa,
Grindelia squarrosa, Panicum miliaceum etc.
3. Erodoalluviophyton – reuneşte comunităţi de specii ecologic asemănătoare, care se formează pe tere-
nuri neînţelenite apărute în urma alunecărilor sau pe pantele abrupte ale râpilor, malurilor râurilor, aluviuni-
lor argiloase. Specii caracteristice: Bothriochloa ischaemum, Centaurea diffusa, Cyclachaena xanthiifolia,
Erigeron annus, E. canadensis, Equisetum arvense, Inula britanica, Lythrum salicaria, L. virgatum, Phrag-
mites australis, Sisymbrium loeselii, Typha angustifolia, T. latifolia, Xanthium californicum. În dependenţă
de adâncimea orizontului apelor freatice pot fi identificate un şir de varietăţi ecologice.
4. Agrophyton – complex floristic mixt, care întruneşte speciile adaptate la condiţiile de soluri lucrate,
unde acţiunea antropică este permanentă şi puternică, structura solului fiind complet schimbată. Poate fi îm-
părţit în mai multe grupe bioecologice, după cum urmează.
a) Interariophyton – grup adaptat să vegeteze în culturile de prăşitoare. Reuneşte specii care vegetează pe
terenuri mai mult sau mai puţin deschise cu soluri afînate. Din cauza acţiunii antropice permanente şi puter-
nice au un ritm de dezvoltare specific. Ciclul de dezvoltare într-o perioadă de vegetaţie este întrerupt de ne-
numărate ori. Predomină paucienele. Speciile caracteristice: Amaranthus hybridus, A. retroflexus, Capsella
bursa-pastoris, Chenopodium album, Cirsium arvense, Consolida regalis, Convolvulus arvensis, Cynodon
dactylon, Digitaria sanguinalis, Echinochloa crusgali, Erigeron annus, E. canadensis, Euphorbia helio-
scopia, E. peplus, Galisoga parviflora, Lactuca serriola, Lathyrus tuberosus, Portulaca oleracea, Setaria
viridis, Sonchus oleraceus, Stellaria media, Xanthium californicum etc.
b) Segetophyton – grup adaptat la monoculturi dense, îndeosebi la cele de cereale cu care formează un
complex integru, străvechi şi stabil. Aici predomină paucienele, numărul de plante adventive este mult mai
mare decât al apofitelor. Reprezentanţii caracteristici sunt: Bronus secalinus, Centaureae cyanus, Cirsium
arvense, C. setosum, Erigeron annus, Lathyrus tuberosus, Matricaria perforata, Papaver rhoeas, Polygonum
aviculare, Vicia craca, V. tetrasperma etc.
c) Faciopratophyton – grup de specii acomodate să vegeteze în culturi mixte. Formează comunităţi de
plante sinantrope necultivate mai stabile, dar de o structură mai simplă. Componenţa floristică este dominată
de apofitele perene; de regulă, sunt plante de luncă determinate de particularităţile lor zonal-regionale. Ca
reprezentanţi specifici se enumără: Amaranthus albus, Arenaria serphyllifolia, Aristolochia clematitis, Arte-
misia absinthium, Bromus arvensis, B. commutatus, Capsella bursa-pastoris, Chenopodium album, Consolida
regalis, Digitaria sanguinalis, Falcaria vulgaris, Fumaria scheleichiri, Lamium amplexicaule, Lathyrus
tuberosus, Lotus corniculatus, Medicago falcata, Melandrium album, Melilotus albus, M. officinalis, Micro-
thlaspi perfoliatum, Thlaspi arvense, Torilis arvense, Trifolium arvense T. repens, Verbena officinalis, Vero-
nica polita, Vicia angustifolia etc.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
165
II. CONSPISSOPHYTON – grup de complexe floristice formate din plante sinantrope necultivate, care
vegetează pe soluri mai mult sau mai puţin bătătorite şi din punct de vedere ecologic se apropie de comuni-
tăţile vegetale naturale.
1. Viaphyton – ecovariant, consituit din plante sinantrope necultivate plasate pe marginea drumurilor, că-
rărilor, prin curţi, pe marginea terenurilor sportive etc. Este alcătuit în majoritate din specii sinantrope necul-
tivate străvechi. Printre ele se enumără: Amaranthus albus, Atriplex tatarica, Canabis ruderalis, Cardaria
draba, Chenopodium album, Berteroa incana, Diplotaxis muralis, Lepidium ruderale, Melilotus albus,
M. officinalis, Polygonum aviculare, Ranunculus repens, Saponaria officinalis, Stelaria media, Urtica dioica.
La aceste şi alte specii se alătură şi reprezentanţi ai comunităţilor de luncă, ca Trifolium repens, iar în sudul
republicii şi reprezentanţi ai celor de semipustiu – Tribulus terrestris.
2. Pascuaphyton – ecobiovariant caracteristic terenurilor de luncă şi de stepă degradate, bătătorite şi păşu-
nate excesiv reprezentate prin comunităţi de Arctium lappa, A. tomentosum, Artemisia annua, Cyclachaena
xanthifolia, Conium maculatum, Lamium purpureum, Leonurus quenquelobatus, Lycium barbatum, Sonchus
oleraceus.
3. Derelictophyton – complex floristic al pârloagelor. În condiţii favorabile, când acţiunea antropică este
redusă până la minimum, structura solului şi a covorului vegetal natural se restabileşte, vădit predomină
plantele perene. Nucleul floristic este alcătuit din specii locale.
III. SCYADOPHYTON – întruneşte două ecovariante, în a căror componenţă intră specii sciofile.
1. Confragophyton – ecovariant format din comunităţi de specii de tufărişuri, de liziere, plantaţii forestiere
tinere şi parcuri, care se află sub o acţiune antropică puternică. Nucleul floristic al etajului ierbos este alcătuit
din specii de provenienţă din regiunile mediteraneene şi cele din Europa Centrală. Specii întâmplătoare locale
sunt cu mult mai multe decât cele migratoare. Reprezentanţii caracteristici sunt: Agrimonia eupatoria, Alliaria
petiolata, Anthriscus sylvestris, Asperugo procumbens, Ballota nigra, Poa annua, Torilis japonica etc.
2. Runcatiodrymophyton – ecovariant ce reuneşte grupa de specii nespecifice florei aborigene de pădure.
Nucleul floristic este alcătuit din specii eurasiatice, europene de pădure şi centralasiatice, care vegetează în
complex cu speciile cenozelor naturale. Predomină apofitele şi speciile perene, aproape lipsesc arheofitele.
Caracteristice sunt: Geranium pusillum, Geum urbanum, Chelidonium majus, Urtica dioica.
Este evident că, din punct de vedere istoric, florocomplexele ecotopurilor antropice sunt grupări vegetale
tinere, care supravieţuiesc în condiţiile presingului antropic şi care oglindesc etapa contemporană de dezvol-
tare a florei sinantrope necultivate, procesul de dezvoltare asociat al speciilor sinantrope necultivate, tendinţa
lor de a se reuni într-un complex unic pentru a-şi ridica existenţa şi stabilitatea covorului vegetal.
Din punct de vedere istoric, cele mai vechi şi specifice sunt complexele de specii care vegetează în cultu-
rile de cereale, iar ca ecofiton străvechi al terenurilor degradate poate fi socotit ecofitonul, ale cărui specii
selectiv reacţionează la solurile bogate în nitraţi.
Concluzii
1. Flora sinantropă necultivată include 618 specii din 292 de genuri şi 63 de familii. În calitate de taxoni
noi pentru flora republicii au fost înregistrate şi descrise 32 de specii (inclusiv 4 specii noi pentru flora bazi-
nului de nord-vest al Mării Negre).
2. Componenţa taxonomică a florei sinantrope necultivate a R. Moldova este dominată de reprezentaţii
familiilor: Asteraceae, Brassicaceae, Fabaceae, Lamiaceae, Apiaceae, Caryophyllaceae, Scrophulariaceae,
Boraginaceae, care constituie 66,18%.
3. În baza analizei structurii biotopurilor compoziţiei floristice şi fitocenotice specifice biotopurilor florei
sinantrope necultivate au fost evidenţiate şi descrise 3 grupe de complexe ecologo–floristice (Edaphofodio-
erodophyton, Conspissophyton, Sciadophyton), 9 ecofitoni (Ruderophyton, Aggeratiophyton, Erodoalluvio-
phyton, Agrophyton, Viaphyton, Pascuaphyton, Derelictophyton, Confragophyton, Runcatiodrymophyton) şi
4 ecobiogrupe.
4. Flora sinantropă necultivată constituită din specii aborigene predomină substanţial şi include 429 de
specii (69,4%), iar grupa plantelor sinantrope necultivate adventive enumeră 189 de specii (30,6%).
5. Răspândirea florei sinantrope necultivate în zonele botanico-geografice ale R. Moldova nu este similară
după numărul de specii, structura compoziţională, particularităţile ecologice şi rolul lor în formarea comuni-
tăţilor fitocenotice. Din numărul total de specii (618) numai 480 (77%) sunt răspândite în ambele zone, iar

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
166
138 de taxoni specifici au fost înregistraţi fie numai în zona de silvostepă – cca 90 de taxoni, fie numai în cea
de stepă – peste 40 de taxoni.
6. După apartenenţa ecosistemică, abundenţă-dominanţă, compoziţie taxonomică, plantele sinantrope ne-
cultivate se ordonează în anumite grupe specifice următoarelor tipuri de ecosisteme: agrare de culturi păioase,
prăşitoare şi lucerniere; agrare de culturi multianuale viticole şi pomicole; naturale de pajişti şi fâneţe; natu-
rale forestiere; terenuri degradate (rurale, viare, pârloage etc).
7. Din tipurile de agroecosisteme, mai bogate în specii sinantrope necultivate sunt cele viticole şi pomi-
cole, în cadrul cărora au fost identificate, respectiv, 439 şi 382 de specii. În flora ecosistemelor degradate –
495 de specii. Tipurile de ecosisteme viare (marginea căilor de comunicaţie) cu peste 400 de specii reprezintă
refugii transformate în focare de naturalizare, aclimatizare şi expansiune potenţială a speciilor adventive şi
invazive.
RECOMANDĂRI PRACTICE
în vederea ameliorării particularităţilor structural-funcţionale ale ecosistemelor
deteriorate şi afectate de plantele sinantrope necultivate
În baza cercetărilor efectuate şi rezultatelor obţinute ce ţin de taxonomia, biologia, ecologia, corologia,
rolul şi valoarea economică a florei şi vegetaţiei sinantrope necultivate a Republicii Moldova, au fost elabo-
rate şi propuse spre realizare în practică următoarele recomandări:
1. Reconstrucţia ecologică a tuturor ecosistemelor de pajişti şi fâneţe deteriorate prin activităţi de amelio-
rare a biotopurilor, de substituire a plantelor sinantrope necultivate cu plante furajere tipice biotopului
respectiv, de utilizare raţională şi gospodărire judicioasă a resurselor vegetale din lunci şi pajişti de
coline.
2. Întocmirea hărţii biotehnologice şi respectarea ei cu stricteţe, efectuarea măsurilor agrotehnice de
intreţinere a culturilor şi de combatere a plantelor sinantrope necultivate în tipurile de agroecosisteme:
viticole, pomicole, legumicole, cerealiere etc.
3. Organizarea monitoringului naţional al plantelor adventive, invazive şi de carantină, efectuarea lucrări-
lor de combatere a lor, mai cu seamă în limitele ecosistemelor degradate transformate în focare de
aclimatizare, înmulţire şi extindere a agenţilor vegetali agresivi necultivaţi.
4. Informarea, prin diverse activităţi, a publicului larg despre pericolul şi daunele provocate ecosisteme-
lor naturale de plantele de carantină şi cele invazive; instruirea tineretului studios cu privire la procesele
de fitoinvazie.
5. Includerea în Baza republicană de date cu privire la biodiversitatea Republicii Moldova a unui compar-
timent separat consacrat plantelor sinantrope necultivate.
Bibliografie:
1. Biological pollution: the control and the impact of exotic species (Proceeding of a symposium held at the University) /
Ed. B. N. Mcknight. – Purdue University al Indianapolis on October 25 & 26 1991 – Indianapolis: Indiana Academy
of Science, 1993. 262 p.
2. Biological invasions of ecosystems by pests and beneficial organism. Proceeding of an international Workshop
(Tsukuba, 26-27 February 1997) /Eds. E.Yano, K.Matsuo, M.Shiyomi & D.A. Andow. – Tscuba: National Institute
et Agro-Environmental Sciences 1997 Niaes /series 3. 224 p.
3. CHIFU, T., MÂNZU, C., ZAMFIRESCU, O. Flora şi vegetaţia Moldovei (Romania). Iaşi: Ed. univ. „Alexandru
Ioan Cuza”, 2006. I Flora, 367 p. II vegetaţia. 698 p.
4. CIOCÎRLAN, V. Flora ilustrată a României. Vol.1, 2. Bucureşti: Cereş, 1988, 512 p.; 1990, 598 p.
5. CUHARSCHI, L., Mârza, M. Flora staţiei feroviare Chişinău şi împrejurimile ei. În: Materialele Conferinţei corpu-
lui didactico-ştiinţific „Bilanţul activităţii ştiinţifice a USM pe anul 1994-1995. Chişinău, 1995, p.234.
6. Flora R.P. Română, vol. I-XIII. Bucureşti: Editura Academiei, 1952-1976.
7. Harmful non-indigenous species in the United States /U.S. Congress, Office of Technology Assessment. Washington,
D.C.: US. Government Printing Office, 1993. 392 p.
8. MITITELU, D. Flora şi vegetaţia judeţului Vaslui. În: Stiinţe şi Comunicări ale Muzeului de Ştiinţe Naturale.
Bacău, 1975, nr.7, p.67-162.

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
167
9. MORARIU, I. Asociaţii de plante din jurul Bucureştilor cu observaţii asupra răspândirii lor în ţară şi mai ales în
Transilvania. În: Buletinul Grădinii Botanice şi al Muzeului Botanic de la Universitatea din Cluj-Timişoara, 1943,
23, nr.2, 3-4, p.131-212.
10. MYRZA, M.V. Atlas florae Europaeae (Distribution of Vascular plants. Crucifferae. (Sysymbrium-Aubrieta).
Helsinki, 1994, vol.10. 224 p.
11. MYRZA, M.V. Atlas florae Europaeae (Distribution of Vascular plants. Crucifferae. (Resedaceae to Platanaceae).
Helsinki, 1999, vol.12. 250 p.
12. MÂRZA, M. Naturalizarea plantelor adventive în condiţiile Republicii Moldova. În: Biodiversitatea vegetală a
R. Moldova: Culegere de articole ştiinţifice. Chişinău, 2001, p.131-134.
13. MYRZA, M. Atlas florae Europaeae (Distribution of Vascular plants. Rosaceae (Spiraea to Fragaria, excl. Rubus).
Helsinki, 2004, vol.13. 320 p.
14. MYRZA, M. Atlas florae Europaeae (Distribution of Vascular plants. Rosaceae (Alchemilla and Aphanes). Helsinki,
2007, vol.14. 200 p.
15. MÂRZA, M., ŞABANOVA, G., CUHARSCHI, L., MÂRZA, C. Cercetări asupra buruienilor din rezervaţia naturală
„Pădurea Domnească”. În: Rezumatele Simpozionului jubiliar „Rezervaţia naturală Codru – 25 de ani”. Lozova,
1996, p.158-160.
16. MÂRZA, M., UNGUREANU, I. Contribuţii la cunoaşterea vegetaţiei nitrofile din R. Moldova. În: Materialele
Conferinţei ştiinţifico-practice consacrate împlinirii a 125 de ani de la naşterea academicianului N.Dimo. Vol.II.
Chişinău, 1998, p.86-88.
17. MÂRZA, M., UNGUREANU, I. Complexe floristice sinantrope din R. Moldova. În: Materialele Conferinţei
ştiinţifico-practice consacrate împlinirii a 125 de ani de la naşterea academicianului N.Dimo. Vol.II. Chişinău,
1998а, p.84-86.
18. NEGRU, A. Determinator de plante din flora Republicii Moldova. Chişinău, 2007, 391 p.
19. Plant invations. General aspects and special problems /Eds. P.Pysec, K.Prach, M.Reymanek & M.Wade. Amsterdam:
SPV Academic Publishing, 1995. 257 p.
20. Plant invations. Studies from North America and Europe /Eds. H.Brock, M.Wade, P.Pysek & D.Green. Leiden:
Backhuys Publishers, 1997. 221 p.
21. POSTOLACHE, Gh. Vegetaţia Republicii Moldova. Chişinău: Ştiinţa, 1995. 340 p.
22. PRODAN, I. Flora pentru determinarea şi descrierea plantelor ce cresc în România. Ed. a 2-a, vol.1, partea 1,
Cluj, 1939, 624 p + CIX; vol.1, partea 2, Cluj, 1939, p.625-1278; vol.2, Cluj, 1939, 467 p.
23. SANDALĂ, V., POPESCU, A., BARABAŞ, N. Cenotaxonomia şi caracterizarea grupărilor vegetale din România.
Bacău: „I. Borcea”, 1998. 366 p.
24. SĂVULESCU, Tr. Die vegetation von Bessarabien Mit Besonderee Berucksichtigung der Steppe. Bucureşti:
Buchruckerei Bucovina, 1927. 53 p. + XLV pl.
25. SĂVULESCU, Tr., RAYSS, T. Materiale pentru flora Basarabiei. Bucureşti, 1924. 80 p. (Supliment la „Buletinul
agriculturii”, vol.2, 1924, p.1-80; vol.3, 1926, p.81-250).
26. TOFAN-BURAC, T., Chifu, T. Flora şi vegetaţia din valea Prutului. Iaşi: Corson, 2002. 437 p.
27. ГЕЙДЕМАН, Т.С. Определитель высших растений Молдавской ССР. Кишинев: Штиинца, 1986. 637 с.
28. ГРОСГЕЙМ, А.А. Анализ флоры Кавказа, т.1. В: Труды Ботанического института Азербайджанского
филиала АН СССР, 1936, т.1, с.1-260.
29. ДОБРОЧАЕВА, Д.Н. и др. Определитель высших растений Украины. Киев: Наукова думка, 1987. 544 с.
30. ДУБЫНА, Д.В., ПРОТОПОПОВА, В.В. Новые адвентивные виды флоры УССР, принципы их проникнове-
ния и расселения. В: Тезисы докладов VII съезда ВБО, Донецк, 1983. Москва: Наука, 1983. 44 с.
31. ДУБЫНА, Д.В., Протопопова B.B. Euphorbia maculatа L. – новий для флори Украiни вид. В: Украинский
ботанический журнал, 1984, том 41, №3, с.33-36.
32. КЛОКОВ, M.B. Рiд будяк – Carduus L. В: Флора УРСР, 1962, №11, с.494-520.
33. КУЗЬМИНОВА, Н.Н. Находки обугленных остатков семян сорных растений на территории трипольских
памятников Молдавии. В: Флора, геобот. и палеоэтнобот. (Бот. иссл.), Кишинев: Штиинца, 1988, с.3-12.
34. КУХАРСКАЯ, Л.Г., МЫРЗА, М.В. Cenchrus pauciflorus Dutl. – новое в Молдавии адвентивное растение. В:
Флора и растительность. Кишинев, 1989, №5, с.112-115.
35. КУХАРСКАЯ, Л.Г., МЫРЗА, М.В. Состояние и перспективы изучения адвентивных растений флоры Мол-
давии. В: Теоретические и прикладные аспекты изучения флоры Молдавии (Материалы Республиканской
научной конференции). Кишинев, 1989а, с.18-23.
36. МАЛЫШЕВА, В.Г. Адвентивная флора Калининской области: Автореф. дисc... канд. биол. н. -Л., 1978. 17 с.
37. МЫРЗА, М.В. Сегетальная флора северных районов Молдавии. В: Тезисы Всесоюзного совещания «Агро-
фитоценозы и экология. Пути повышения их стабильности и продуктивности». Ижевск, 1988, с.143-144.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
168
38. МЫРЗА, М.В. Современное состояние и перспективы изучения агрофитоценозов ССР Молдовa. В: Научная
конференция профессорско-преподавательского состава и сотрудников по итогам научно-исследователь-
ской работы за XII пятилетку. (Тезисы докладов). Кишинев, 1990а, с.223.
39. МЫРЗА, М.В. О некоторых редких и адвентивных растениях Молдавии. В: Ботанический журнал, 1991,
76, №1, с.129-134.
40. МЫРЗА, М.В. Биология и распространение Cirsium arvense L. на территории Молдовы. В: Материалы
конференции по онтогенезу высших растений. Ужгород, 1992, с.76-78.
41. МИРЗА, M.В., КУХАРСЬКА, Л.Г., Гочу, Д.I. Поширення Grindelia squarrosa (Pursh.) Duck. на тарриторii
Молдавii. В: Украинский ботанический журнал, 1987, том 44, №6, с.42-44.
42. МЫРЗА, М.В., КУХАРСКАЯ, Л.Г. Новый для Молдавии адвентивный сорняк. В: Сельское хозяйство
Молдавии, 1988, №8. 23 с.
43. МЫРЗА, М.В., КУХАРСКАЯ, Л.Г. Некоторые адвентивные растения Молдавии. В: Актуальные вопросы
ботаники в СССР. Алма-Ата, 1988а, с.70.
44. МЫРЗА, M.B., КУХАРСКАЯ, Л.Г. Особенности распространения некоторых адвентивных растений Мол-
давии. В: Исследования по экологии, флористике, биохимии и физиологии растений Молдавии. Кишинев.
1988б,в, с.60-68.
45. МЫРЗА, М.В., КУХАРСКАЯ, Л.Г. Рост и развитие Amaranthus spinosus в условиях Молдавии. В: Онтогенез
высших цветковых растений. Киев, 1989, с.89-90.
46. МЫРЗА, М.В., ЧЕКОЙ, В.Н. Естественное зарастание золоотвальников Молдавской ГРЭС. В: Теоретиче-
ские и практические аспекты изучения флоры Молдавии (Материалы научной конференции). Кишинев,
1989, с.26-28.
47. ПАЧОСКИЙ, И.К. Основные черты развития флоры юго-западной России. В: Записки Новороссийского
Общества Естествоиспытателей, 1910, №34, Прилож: 1-430.
48. ПАЧОСКИЙ, И.К. О сорно-полевой растительности Херсонской губернии. В: Труды Бюро по прикладной
ботанике, 1910а, с.71-125.
49. ПОПОВ, М.Г. Основы флорогенетики. Москва, 1963. 143 с.
50. ПРОТОПОПОВА, В.В. Натуралізація адвентивних рослин Україні. В: Украинский ботанический журнал,
1988, том 45, №4, с.9-15.
51. ЦВЕЛЕВ, Н.Н. Определитель сосудистых растений северо-западной России (Ленинградская, Псковская,
Новгородская области). Санкт-Петербург, 2000. 780 с.
52. ЧЕРЕПАНОВ, С.К. Сосудистые растения СССР. Ленинград: Наука, 1981. 509 с.
53. ШАБАНОВА, Г.А., МЫРЗА, М.В., ВАНИНА, П.В. Новые и редкие виды для некоторых районов Молдавии.
В: Теоретические и практические аспекты изучения флоры Молдавии. Кишинев, 1989, с.41-43.
54. ШМАЛЬГАУЗЕН, И.Ф. Флора юго-западной России (Киевской, Волынской, Подольской, Черниговской и
смежных местностей). Киев, 1886. 783 с.
55. ЯНУШЕВИЧ, З.В. Видовой состав сельскохозяйственных растений в XVI-XIX вв. на территории Молдавии
по археологическим данным. В: Тезисы докладов Межреспубликанского симпозиума по аграрной истории
Восточной Европы (Рига, 1970). Москва, 1970, с.152-155.
56. ЯНУШЕВИЧ, З.В. Остатки культурных растений на археологических раскопках средневековых памятников
Молдавии. В: Археологические исследования в Молдавии в 1968-1969 гг. Кишинев, 1972, с.250-257.
57. ЯНУШЕВИЧ, З.В. Культурные растения первобытного периода на юго-западе СССР: Автореф. дисc. докт.
биол. наук. Кишинев, 1978. 31с.
58. ЯНУШЕВИЧ, З.В., БЫРНЯ, П.П. Из истории земледелия на территории Старого Орхея. В: Вопросы эконо-
мической истории Молдавии эпохи феодализма и капитализма. Кишинев, 1972, с.267-276.
Prezentat la 10.09.2013

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
169
МОДЕРНИЗАЦИЯ БИОЛОГИЧЕСКОГО ОБРАЗОВАНИЯ
В 12-летней ШКОЛЕ КАЗАХСТАНА
Р.Ш. ИЗБАСАРОВА, А.Ш. ИЗБАСАРОВА*
Казахский национальный педагогический университет им. Абая *Казахский национальный медицинский университет им. С. Асфандиярова
MODERNIZAREA ÎNVĂŢĂMÂNTULUI BIOLOGIC
ÎN ŞCOALA DE 12 ANI DIN KAZAHSTAN
Modernizarea instruirii biologice în şcoala de 12 ani urmăreşte introducerea în procesul instructiv-educativ a cursurilor
aplicative şi a celor opţionale, care vor ajuta elevul în alegerea profesiei, precum şi vor contribui la specializarea instruirii.
Cursul aplicativ „Genetică umană” are drept obiectiv de a aprofunda cunoştinţele elevilor în acest domeniu al ştiinţei.
Cuvinte-cheie: învăţământ biologic, modernizarea învăţământului, discipline aplicative, unităţi de conţinut.
RENEWAL OF THE CONTENT OF BIOLOGICAL EDUCATION
IN THE KAZAKHSTAN 12-YEAR SCHOOL
Perfection of biological education at 12-years-old school is seen in introduction in the educational process of the applied
courses and courses on a choice, which will help a student in the choice of profession, and also will play a role in profiling of
teaching. Before the applied course of «Geneticist of man» a task costs in deepening of knowledge of students in this area of
science.
Keywords: biological education, modernization of education, applied disciplines, content units.
Введение
Динамика развития системы образования в Казахстане с 12-летним обучением предполагает смену
социально-педагогических ценностей, децентрализацию всех уровней управления, предполагающую
введение в систему образования норм и правил, регулирующих деятельность всех участников образо-
вательного процесса.
Результаты исследований и их обсуждение
В Казахстане тактические шаги по реализации педагогической концепции учета интересов, спо-
собностей учащихся посредством дифференциации процесса обучения в школе, вариативности форм
и средств образования предпринимаются в условиях внедрения государственных стандартов, приня-
тых в 2010 году.
Концепцией развития общеобразовательной школы определенó завершение обучения на старшей
ступени общеобразовательной школы. При этом обучение учащихся с учетом их интересов на основе
дифференциации в 12-летней школе можно организовать по следующим направлениям: естественно-
математическому, гуманитарно-эстетическому и профессионально-техническому.
Формирование определенного уровня избирательности учащихся в предпочтении и усвоении тех
или иных областей знаний, в предоставлении им возможности и права выбора в процессе обучения
отражало стремление «решительного поворота современной школы к гуманизации и персонализации
содержания образования».
Рассмотрим подробнее содержание биологического образования в школе согласно ГОСО переход-
ного периода и учебной программе по биологии.
1. Четко представлено по каждому уровню базовое содержание, выстроенное на основе содержа-
тельных линий образовательных областей, ожидаемые результаты которых даны путем интег-
рации ключевых и предметных компетенций. Сформированные в начальной школе ключевые
компетенции развиваются в среднем уровне средствами всех учебных дисциплин.
2. Соблюдена структура базового содержания, состоящая из инвариантной и вариативной частей.
Вариативная часть школьного компонента позволяет организовать занятия по выбору, а уче-
нический компонент направлен на реализацию новых форм и приемов организации учебно-
воспитательного процесса.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
170
3. Учебная программа (авторы: Жумагулова К.А., Избасарова Р.Ш.) основывается на психолого-
педагогическом и инновационном опыте педагогов-практиков, предполагая переориентацию
образования в соответствии с идеями развивающего обучения.
4. Ведущей целью программы является создание благоприятных условий для разностороннего
развития личности как основы психического и физического здоровья детей, повышения каче-
ства образования.
5. В основу реализации программы положен принцип личностно-ориентированного взаимодей-
ствия учителя с учащимися, принцип гуманистической психологии (создания условий для раз-
вития ученика и его максимальной самореализации), принцип культуросообразности. Обозна-
чены и научно обоснованы приоритеты развивающего обучения, обусловленные развитием
ряда фундаментальных способностей учащихся – творческого воображения, постигающего
мышления, произвольности, коммуникативности и др.
Приоритетами организационно-методического характера является рассмотрение учебных про-
грамм прикладных курсов и курсов по выбору. Прикладные курсы дают возможность вхождения в
мир профессий через расширение или углубление базового содержания профильного компонента.
Еще одна особенность прикладных курсов – предметы должны иметь междисциплинарный интегра-
тивный характер. Выбор тем прикладных курсов зависит от контингента учащихся, их интересов, по-
требностей.
Выход видится нам в целенаправленной подготовке и стимулировании педагогов к разработке
курсов по выбору и привлечении имеющегося опыта. Нами были разработаны курсы по выбору (при-
кладные курсы) для учащихся естественно-математического профиля обучения. Так, разработанный
нами курс «Генетика человека» для естественнонаучного направления общеобразовательных школ
предназначен для учащихся 10 класса.
С нашей точки зрения, в программе «Общая биология» недостаточно времени уделяется изучению
вопросов, касающихся наследственности человека, поэтому учащимся, интересующимся генетикой,
мы предлагаем данный курс. В этом курсе рассматриваются как теоретические, так и практические
работы по решению генетических задач, знакомящих обучающихся с многообразием наследственных
заболеваний, их лечением и профилактикой. Решение задач в качестве учебно-методического приема
при изучении вопросов наследственных заболеваний способствует качественному усвоению знаний,
получаемых теоретически, развивает умение рассуждать и обосновывать выводы, расширяет круго-
зор учащихся. Решение задач развивает у школьников логическое мышление, позволяет им глубже
понять учебный материал.
Данный курс преследует определенные цели:
1. Углубить и расширить знания учащихся, интересующихся биологией, по наиболее важным и
значимым проблемам наследственности человека как факторе здоровья.
2. Ознакомить обучающихся с основными методами изучения генетики человека на конкретных
заболеваниях, рассмотреть последствия мутаций, затрагивающих генотип человека.
3. Показать учащимся, что знание законов генетики позволяет предупредить наследственные бо-
лезни или ослабить их проявление.
Для обеспечения эффективного медико-генетического консультирования необходима пропаганда
генетических знаний, осведомленность населения в вопросах наследственных болезней.
В основу данного курса положены принципы углубления и систематизации знаний, полученных
при изучении основного курса, прикладной направленности курса, развитие интереса учащихся к са-
мостоятельному приобретению знаний посредством подготовки сообщений, написания рефератов,
поиска серьезных источников информации, в которых знания излагаются в точном соответствии с со-
временным состоянием науки. Это поможет реализовать исследовательский подход, вовлечь учащих-
ся в поисковую, творческую деятельность.
Курс поможет учащимся в усвоении основных понятий, терминов и законов генетики, генетиче-
ской символики, в объяснении жизненных ситуаций с точки зрения генетики, в подготовке к ЕНТ, а
возможно и в выборе профессии биологической направленности.
В качестве диагностики предлагаются следующие формы:
• вводный контроль;

Seria “Ştiinţe reale şi ale naturii”
Biologie ISSN 1814-3237
171
• тестирование;
• выходной контроль;
• защита проектов и презентаций.
Были разработаны также требования к результатам изучения курса. Так, учащиеся должны знать
основные понятия, термины и законы генетики, генетическую символику, уметь правильно формули-
ровать условия, решения и ответы генетических задач, а также решать типичные задачи и логически
рассуждать и обосновывать выводы.
Критерии оценки успешности представляются в следующем виде:
Ученик получает зачет при условии выполнения 75-100% заданий. В задания входят решение за-
дач, письменные ответы по карточкам, тестирование, успешные ответы. Дополнительные баллы уче-
ник получает за
• использование Интернет-технологий;
• выполнение заданий сверх обязательного минимума;
• доклады в школе или за еѐ пределами.
Курс «Генетика человека» составляет 34 часа, в расчѐте 1 занятие в неделю. Содержание курса
представлено следующими темами:
1. Введение. Здоровье как состояние полного физического, психического, репродуктивного, соци-
ального и духовного благополучия.
2. История изучения человеческого организма и его наследственности от Аристотеля до наших дней.
3. Наследственность как фактор здоровья.
4. Наследственный аппарат соматических и генеративных клеток человека.
5. Методы изучения наследственности.
6. Болезни человеческого организма.
7. Влияние вредных привычек на здоровье человека.
8. Итоговое занятие (тестирование).
Программой курса предусмотрено выполнение практических работ, а также написание и защита
рефератов по предложенным тем.
Учителем может быть предусмотрено продолжение реферативных работ, переходящих в научный
проект, если наблюдается заинтересованность учащихся в проведении дальнейших исследований.
Выводы
При включении данного курса в учебный процесс меняется и деятельность учителя. Учитель 12-
летней школы должен выполнять функции координатора, консультанта, организатора самостоятель-
ной познавательной деятельности и творческой активности учащихся. Педагог должен организовать
учебный процесс так, чтобы помимо предметных знаний у учащихся формировались ключевые ком-
петенции. Грамотный подход к процессу обучения учеников предоставляет широкие возможности не
только для развития интереса учащихся к предмету и изучаемой научной области, но позволяет и са-
мому учителю постоянно совершенствовать свои знания и идти в ногу с наукой и временем.
Prezentat la 25.11.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
172
PROCEDEELE DE AGITARE ŞI CENTRIFUGARE – POSIBILE SURSE
DE ERORI METODOLOGICE ÎN STUDIUL ADSORBŢIEI
SUBSTANŢELOR DIN SOLUŢII
Alexei MAFTULEAC, Nina ŢÎMBALIUC
Institutul de Chimie al AŞM
În lucrare a fost analizată informaţia privind metodele de studiere a adsorbţiei substanţelor din lichide pe diferiţi ad-
sorbanţi. În centrul atenţiei au fost condiţiile de contactare şi separare a fazelor (componentelor) sistemului de adsorbţie.
S-a observat că aceste condiţii diferă foarte mult la diferiţi cercetători, ceea ce creează dificultăţi la compararea rezulta-
telor obţinute de ei. A fost înaintată ideea că este necesară o unificare şi optimizare a proceselor de contactare şi sepa-
rare a componentelor sistemului de adsorbţie. În baza datelor noastre experimentale (adsorbţia în câmp centrifugal), s-a
presupus că aceste operaţiuni pot fi o sursă de erori metodologice în studiul adsorbţiei.
Cuvinte-cheie: adsorbţie din soluţii, agitare/centrifugare, erori metodologice, adsorbţie centrifugală, Congo roşu,
cărbune activ.
SHAKING AND CENTRIFUGATION PROCESSES – POSSIBLE SOURCES OF METHODOLOGICAL
ERRORS IN THE STUDY OF ADSORPTION FROM SOLUTIONS
The information regarding the methods of studying the adsorption of substances from liquids on different adsorbents
has been analyzed. In the spotlight were the conditions of the contacting and separation of the phases (components) of
the adsorption system. It was noted that these conditions vary greatly from different researchers, which makes difficult
to compare the result obtained by them. The paper forward idea that is necessary to unify and optimize the contact and
separation process of the component of adsorption system. Basing on our experimental data (adsorption in centrifugal
field), we assume that these operations may be a source of methodological errors in the study of adsorption.
Keywords: adsorption from solutions, shaking/centrifugation, methodological errors, centrifugal adsorption, Congo
red, activated carbon.
Introducere
Metodele de studiere a adsorbţiei substanţelor din soluţii la utilizarea adsorbanţilor minerali, carbonici sau
de altă natură prevăd contactarea pe o anumită perioadă de timp a componentelor sistemului destinat studierii,
până la atingerea echilibrului de adsorbţie [9], în condiţii statice sau la o agitare uşoară pe instalaţia-agitator.
Această operaţiune se efectuează în scopul creării unor condiţii favorabile pentru difuzia moleculelor din
soluţie spre particulele de adsorbant, influenţând astfel şi asupra cineticii procesului de adsorbţie. După atin-
gerea echilibrului de adsorbţie, pentru separarea adsorbantului epuizat de faza lichidă, sistemul de adsorbţie
este supus, de regulă, centrifugării, pentru ca apoi, în supranatant, să fie determinată concentraţia de echilibru
a substanţei-adsorbat. În baza datelor experimentale se calculează valorile adsorbţiei specifice pentru un
set de eşantioane, iar din ele se obţine izoterma de adsorbţie, care şi caracterizează procesul de adsorbţie
propriu-zis.
Metodele de studiere a adsorbţiei substanţelor din gaze şi lichide sunt utilizate în prezent nu doar de fizi-
cieni şi chimişti, ci şi de ecologi, medici, biologi [5-7]. Reieşind, însă, din scopurile propuse, experimentele
sunt efectuate în condiţii specifice, ceea ce face dificilă compararea rezultatelor obţinute de cercetători,
mai ales ale celor care activează în domenii diferite.
Rezultate şi discuţii
Scopul prezentei lucrării a fost analiza condiţiilor în care se studiază adsorbţia unor substanţe bine cunos-
cute şi compararea rezultatelor obţinute de cercetători, punând în evidenţă modul de efectuare a unor sau
altor operaţiuni în procesul de studiere a adsorbţiei. La efectuarea analizei s-a operat cu aşa indici ca tipul,
intensitatea şi durata operaţiunii (stadiului) în cercetarea procesului de adsorbţie. Informaţia obţinută este
prezentată în tabel.

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
173
Tabel
Operaţiuni ce necesită a fi efectuate în procesul de studiere a adsorbţiei din lichide
Operaţiuni Intensitatea Timpul Referinţa
Agitare
Centrifugare
**
**
În 10…30 zile,
agitat periodic
[16]
Centrifugare* 3000 rot/min. 15 min. [13]
Agitare
Centrifugare
120 rot/min.
14000 rot/min.
48 ore
**
[5]
Centrifugare * 4500 rot/min. 10 min. [9]
Agitare * 400…800 rot/min. ** [7]
Agitare * 160 osc/min. 24 ore [6]
Agitare
Centrifugare
**
3000 rot/min.
12 ore
3 min.
[8]
Agitare
Centrifugare
150 rot/min.
3000 rot/min.
24 ore
30 min.
[4]
Agitare
Pauză
Centrifugare
**
-
**
15 min.
30 min.
2-3 min.
[1]
Notă: Informaţii despre o altă operaţiune (*) sau despre caracteristica respectivă (**) lipsesc.
Tabelul reflectă diversitatea condiţiilor de efectuare a cercetărilor şi faptul că, uneori, nu este respectată
chiar necesitatea de a informa clar în ce condiţii au fost efectuate cercetările. Este, însă, cunoscut că adsorb-
ţia depinde de mai mulţi factori – temperatură, intensitatea şi durata agitării sistemului de adsorbţie, gradul
de dispersie a adsorbantului ş.a. [3].
Experimentele efectuate, având ca scop studierea adsorbţiei în câmp centrifugal, au demonstrat că la cen-
trifugarea eşantioanelor, în dependenţă de viteza de rotaţie şi durata procesului, au loc aşa fenomene ca ridi-
carea temperaturii (1...10ºC) în probe şi intensificarea adsorbţiei de circa 2...3 ori [2], faţă de cea efectuată în
condiţii statice. Ridicarea temperaturii, însă, măreşte viteza reacţiilor chimice [4]; ca urmare, pot avea loc
schimbări necontrolate în sistemul studiat, iar mărirea adsorbţiei la centrifugare conduce la furnizare de date
eronate referitor la valoarea adsorbţiei, considerate ca fiind corecte (în condiţiile planificate, dar nu în cele reale).
Pentru a clarifica în ce măsură influenţează condiţiile de centrifugare asupra valorii adsorbţiei, au fost efec-
tuate cercetări suplementare. În calitate de adsorbant a fost utilizat cărbune activ medicinal, iar ca substanţă-
adsorbat – colorantul Congo roşu. Caracteristicile cărbunelui activ, obţinute prin metoda adsorbţiei azotului,
sunt următoarele: suprafaţa totală sorbtivă S=712,4 m2/g, volumul porilor Vs=0,429 cm
3/g, raza lor efectivă
(Ref) fiind cuprinsă între 0,2-0,8, 1,2-1,3 şi 1,8-2,2 nm. Distribuirea porilor cărbunelui după dimensiuni este
în aceeaşi ordine, adică structura adsorbantului conţine preponderent micropori.
Colorantul Congo-roşu se caracterizează ca având inele benzenice şi grupe de N=N, NH2 şi SO3Na [8],
dimensiunile moleculei (lungimea, lăţimea, grosimea, nm) fiind 2,85x0,90x0,37 [1], masa moleculară rela-
tivă – respectiv, 696,663.
Procesul de adsorbţie a fost efectuat contactând cărbunele activ cu soluţii de colorant, în păhare de sticlă
amplasate în cuiburile rotorului centrifugii T52.1 (Germania). Viteza de rotaţie era programată şi vizualizată
cu ajutorul tahometrului centrifugii, iar durata procesului controlată cu un secundometru. După expirarea
timpului de contactare, rotorul centrifugii era oprit, iar soluţia-reziduu din păhare – separată. În soluţii, până
la şi după adsorbţie, erau măsurate concentraţiile colorantului, utilizând în acest scop spectrofotometrul
JENWAY 6505 (numărul de undă – 505 nm, cuveta – 1 cm). În baza diferenţei concentraţiei soluţiei până la
şi după contactarea cu adsorbantul era calculată valoarea adsorbţiei specifice şi obţinută izoterma adsorbţiei.
Rezultatele sunt prezentate în Figura 1. După cum se observă din această figură, izotermele de adsorbţie
obţinute în câmp centrifugal sunt asemănătoare celor de tipul II după clasificarea BDDT (Brunauer, Demmis,
Demmis, Teller). Până la concentraţii medii de echilibru (200-300 mkmol/L) ele caracterizează procesul ca
fiind o adsorbţie a moleculelor neasociate. La mărirea concentraţiei colorantului are loc asocierea molecule-
lor lui în micelii, care, fiind adsorbite de către cărbune, duc la creşterea valorii adsorbţiei.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
174
0 100 200 300 400 500
0
2
4
6
8
10
12
14
a, m
km
ol/g
Ce, mkmol/L
1
2
3
Fig.1. Adsorbţia colorantului Congo roşu pe cărbune activ în condiţii de centrifugare,
4000 rot/min.; durata procesului (1,2,3), respectiv, una, două şi trei ore.
0,0 0,5 1,0 1,5 2,0 2,5 3,0
0
1
2
3
4
5
6
7
8
9
a,m
km
ol/g
timpul, ore
1
2
3
Fig.2. Dependenţa adsorbţiei colorantului Congo roşu la centrifugare de viteza rotorului centrifugii:
1,2 şi 3 corespund vitezelor 0 (static),1500, şi 4000 rot/min.
Valorile adsorbţiei sunt exprimate pentru concentraţia de echilibru 100 mkmol/L.
Procesul de adsorbţie depinde de durata centrifugării (Fig.1 şi 2). La o durată relativ mică a procesului
(câteva zeci de minute) şi la concentraţii ale adsorbatului de până la circa 100 mkmol/L, la „începutul” izoter-
mei această dependenţă este practic liniară. Din Figura 2 observăm că valoarea adsorbţiei depinde în mare
măsură de viteza de rotaţie – turaţia rotorului centrifugii, fapt ce se explică prin creşterea considerabilă (de
sute şi mii de ori) a presiunii în lichidul centrifugat. Aceasta, însă, cauzează o împingere forţată a molecule-
lor colorantului în porii adsorbantului, influenţând în aşa mod cinetica procesului de adsorbţie. Din datele
prezentate se observă o clară dependenţă a adsorbţiei de modul de efectuare a cercetărilor experimentale, în-
deosebi la executarea operaţiunilor considerate ca auxiliare, deci neesenţiale. Dacă analizăm datele prezentate
în tabel şi la informaţia prezentată de ele adăugăm diversitatea proprietăţilor fizico-chimice ale adsorbanţilor
(fie chiar şi de aceeaşi natură), atunci intervalul posibilelor erori impresionează.

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
175
În opinia noastră, condiţiile de agitare şi centrifugare sunt factori importanţi şi trebuie luaţi în considerare
în procesul de studiere a adsorbţiei substanţelor din soluţii. Astfel, aceste procedee pot deveni surse de erori
metodologice în cercetări.
Concluzii
Din cele expuse pot fi formulate următoarele concluzii:
1. Este necesară o unificare şi optimizare a operaţiunilor de tipul agitării, centrifugarii ş.a., deoarece ele
pot influenţa fiabilitatea datelor obţinute experimental.
2. Erorile ce pot apărea la agitare şi/sau separare în procesul de studiere a adsorbţiei depind de durata de
expunere a eşantioanelor operaţiunilor respective, precum şi de intensitatea acestora. Probabil, ar putea fi
elaborate condiţii optime de efectuare a acestor procedee, care apoi să fie unanim recunoscute şi utilizate în
cercetări, dar aceste aspecte ale problemei deocamdată n-au fost puse în discuţie.
Bibliografie:
1. LUPAŞCU, T. Cărbuni activi din materii prime vegetale. Chişinău: Ştiinţa, 2004. 129 p.
2. MAFTULEAC, A., ŢÎMBALIUC, N. Adsorbţia colorantului Congo-roşu pe cărbune activ în condiţii obişnuite şi în
câmp centrifugal. În: Materialele celei de-a 31-a Conferinţe Naţionale de Chimie, 2010, 6-8 octombrie, Râmnicu -
Vâlcea, România, p.207.
3. КЕРДИВАРЕНКО, М. Молдавские природные адсорбенты и технология их применения. Кишинев: Картя
Молдовеняскэ, 1975. 191 с.
4. ЛЕВЧЕНКО, С. Физическая и коллоидная химия. Ч. 2. Химическая кинетика и катализ. Ростов-на-Дону:
Ростовский госуниверситет, 2004, с.13-17. (bookre. Org/reader?file=13397168&pg=13)
5. ЛЕРНЕР, М., БАКИНА, О., ГЛАЗКОВА, Е. и др. Адсорбция микроорганизмов и бактериального эндоток-
сина на модифицированных полимерных волокнах. B: Перспективные материалы, 2011, №3, с.53-58.
6. САРТАКОВ, М. Адсорбционная способность гуминовых кислот торфов среднего Приобья. B: Вестник
Алтайского государственного аграрного университета, 2011, №4(78), с.60-64.
7. СЕРОВА, А., ПЕХЕНЬКО, В., ТИХОНОВА, И. и др. Адсорбционная и поглотительная способность мате-
риала, включающего наноструктурный оксигидроксид алюминия. Сибирский медицинский журнал (Томск),
2012, т.27, вып.2, с.127-130.
8. Химический энциклопедический словарь (ред. Кнунянц И.П.). Москва: Советская Энциклопедия, 1983, с.271-
272.
9. Экспериментальные методы в адсорбции и молекулярной хроматографии (под ред. А.В. Киселевa и В.П.
Древингa). Москва: Изд-во МГУ, 1973, с.382- 400; 2-ое издание, 1990 (ред. Ю.С. Никитин и Р.С. Петрова),
с.256-265.
Prezentat la 14.06.2013

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
176
INTERCALAREA BENTONITULUI LĂRGUŢA (R. MOLDOVA)
CU OLIGOMERI DE ALUMINIU
Vasile RUSU, Alexei MAFTULEAC, Oleg PETUHOV
Institutul de Chimie al AŞM
Au fost efectuate analize cu raze X şi determinate caracteristicile sorbţional-structurale din izotermele de adsorbţie a
azotului pentru bentonitul Lărguţa (R. Moldova), intercalat cu oligomeri de aluminiu. S-a constatat o majorare de 1,5-2 ori
a suprafeţei specifice (după BET) a adsorbanţilor intercalaţi. Adsorbanţii sintetizaţi utilizând bentonitul Lărguţa posedă
o stabilitate termică satisfăcătoare, caracteristicile sorbţionale menţinându-se înalte după calcinare la temperaturi de
350-450°C, ceea ce prezintă perspective reale de utilizare în practică.
Cuvinte-cheie: bentonit Lărguţa, adsorbanţi intercalaţi, caracteristici sorbţional-structurale.
ALUMINUM-PILLARING OF BENTONITE LARGUTZA (R. MOLDOVA)
The x-ray analysis and sorptional-structural characteristics obtained from nitrogen adsorption izoterms for bentonite
Largutza (R. Moldova) pillared with aluminium oligomers were elucidated. Specific surface area (after BET) of pillared
adsorbents increases by 1.5-2 times as compared to the initial bentonite. On the whole, the synthesized adsorbents on the
basis of bentonite Largutza possess high thermal stability after calcination on 350-450°C which presents real prospects
for practical usage.
Keywords: bentonite Largutza, pillared adsorbents, sorptional-structural characteristics.
Introducere
Pentru elucidarea factorilor geometrici (textura adsorbanţilor) şi a chimiei suprafeţei, o caracteristică mai amplă a parametrilor sorbţional-structurali ai adsorbanţilor se obţine aplicând mai multe metode, cum se exem-
plifică într-un şir de lucrări [11,12,17-19]. Suprafaţa specifică este determinată, aplicând modelele BET [4] şi Langmuir [9]. Volumul total al porilor este determinat din valoarea adsorbţiei la presiunea relativă P/PS 0,95 [19]
sau 0,98 [17,18]. Volumul microporilor este determinat din modelul Dubinin-Radushkevich [6,13]. Metoda t-graficii lui de Boer este utilizată frecvent [7,11,19] pentru determinarea suprafeţei specifice exterioare şi a
volumului microporilor. Distribuirea mezoporilor după dimensiuni este analizată conform modelului Barrett-Joyner-Halenda (BJH) [2], iar distribuirea microporilor după dimensiuni este analizată conform modelului
Saito-Foley (SF) [16]. Montmorilonitul în stare pură este mai puţin răspândit în natură; mai frecvent, acest mineral constituie
componentul principal al bentonitelor. Zăcăminte mai însemnate de bentonite în Republica Moldova sunt în localităţile nordice (Prodăneşti şi Naslavcea), de asemenea la sud-vestul republicii (localităţile Cociulia şi Lărguţa).
Zăcămintele de bentonite din localitatea Lărguţa conţin (în stare naturală) cca 40-50% de montmorilonit, restul aparţinând ilitului (cca 20%), caolinitului (cca 5-10%), altor componente neargiloase (carbonaţi, gips, oxizi de fier etc.).
Anterior au fost efectuate studii [14,15] în vederea stabilirii condiţiilor optime pentru sinteza adsorbanţi-lor intercalaţi pe baza montmorilonitului Ascangel (R. Georgia).
Scopul prezentei lucrări constă în elucidarea caracteristicilor sorbţional-structurale, din analize cu raze X
şi izotermele de adsorbţie a azotului (77 K), ale bentonitului Lărguţa intercalat cu oligomeri de aluminiu, în precizarea schemei de determinare a parametrilor sorbţional-structurali ai adsorbanţilor şi în analiza compara-
tivă cu adsorbanţii obţinuţi pe baza montmorilonitului Ascangel (R. Georgia).
Material şi metode Montmorilonitul prezent în bentonitul Lărguţa (natural) se atribuie formaţiunilor alcalino-pământoase,
în complexul de schimb cationic predominând ionii de Ca2+
şi Mg2+
. Pentru transformare în formă sodică, bentonitul a fost dispersat în apă distilată şi omogenizat pe amestecător magnetic. Suspensia de argilă (după
diluare pentru asigurarea sedimentării particulelor) a fost supusă fracţionării, fiind separată cu un sifon frac-
ţiunea <1 μm a suspensiei, folosind procedura de fracţionare prin sedimentare în mediu apos, conform legii Stokes. Obţinerea argilei în formă monoionică de sodiu s-a efectuat prin tratări repetate cu soluţie de NaCl 1 N

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
177
în raport solid : lichid de 1:25. Na-forma obţinută a fost spălată de surplusul de electrolit (prin centrifugare, testul cu AgNO3) şi utilizată în continuare pentru sinteza adsorbanţilor intercalaţi, cum s-a descris anterior [14,15].
Izotermele de adsorbţie a azotului (N2, 77 K) au fost obţinute prin metoda volumetrică, utilizând analiza-
torul automat Quantachrome AUTOSORB-1 (firma Quantachrome, SUA). Analizele roentgenografice ale
adsorbanţilor au fost efectuate utilizând instalaţia DRON-3.
Rezultate şi discuţii
1. Analiza roentgenografică
Fracţiunea <1 m, obţinută după spălare şi fracţionarea granulometrică a bentonitului Lărguţa, este consti-tuită, conform analizelor cu raze X (Fig.1), preponderent din montmorilonit (reflexul bazal, d001, 15,6 Å) şi, în cantităţi neînsemnate, din ilit (d001 10,1 Å) şi caolinit (d001 7,17 Å). Montmorilonitul prezent în bentonitul natural se atribuie formaţiunilor alcalino-pământoase, în complexul de schimb cationic predominând ionii de Ca
2+ şi Mg
2+. După transformare în formă sodică, Na-bentonitul (Na-forma, Fig.1) este caracterizat prin
reflexul bazal 13,8 Å, care după calcinarea mostrei la 350°C se contractă până la mărimea ~10 Å, suprapu-nându-se pe reflexul ilitului (10,1 Å). După calcinarea mostrei la 550°C, reflexul d001 7,17 Å dispare, ceea ce este caracteristic pentru mineralele din grupa caolinitului.
Pentru stabilirea gradului de intercalare a oligomerilor în spaţiul interlamelar al argilei şi determinarea stabilităţii produsului intercalat se aplică, cel mai frecvent, metoda analizei cu raze X (analiza roentgenogra-
fică). Introducerea speciilor oligomerice de aluminiu în spaţiul interlamelar al montmorilonitului duce la creşterea distanţei între planele 001 ale montmorilonitului; în consecinţă, cresc valorile reflexului bazal d001.
Fig.1. Difractogramele roentgenografice (preparate orientate,
iradiere FeKα) ale bentonitului Lărguţa (RM), fracţiunea <1 m.
Bentonit natural (Ca,Mg-forma) şi în Na-formă.
Preparate necalcinate (25°C) şi după
calcinare la 350°C şi 550°C.
Fig.2. Difractogramele roentgenografice (preparate
orientate, iradiere FeKα) ale bentonitului Lărguţa
intercalat cu oligomeri de aluminiu la raportul molar
iniţial OH-/Al
3+ 2:1. Preparate necalcinate (25°C)
şi după calcinare la 350°C, 450°C şi 550°C.
Intercalarea bentonitului Lărguţa cu oligomeri de aluminiu duce la dilatarea spaţiului interlamelar al montmorilonitului şi la creşterea grosimii stratului produsului intercalat până la 19,2 Å (Fig.2). Calcinarea la 350°C duce la dehidratarea/dexidroxilarea polihidroxipolimerilor de aluminiu, la configurarea şi rearanjarea lor, etapă numită „pilonarea” sau „pivotarea” argilei (pillared clay, PILC). În consecinţă, grosimea stratului produsului intercalat se micşorează, atingând mărimi în domeniul 16-18 Å (preponderent 17,3 Å, Fig.2). Calcinarea în continuare la 450°C practic nu modifică structurile oligomerice, localizate în spaţiul interlame-lar al montmorilonitului (Fig.2). Modificări mai esenţiale în structurile oligomerice se produc după calcinare la 550°C, înregistrându-se reflexii în domeniul 14-17 Å (preponderent 15,2 Å cu sateliţii 14,2 Å şi 16,6 Å).
În ansamblu, adsorbanţii sintetizaţi, utilizând bentonitul Lărguţa, posedă o stabilitate termică satisfăcă-toare, rezistând după calcinare la temperaturi în domeniul 350-550°C, ceea ce nu este caracteristic pentru bentonitul iniţial (Fig.1).
2. Caracteristici sorbţional-structurale Ramura sorbţională a izotermei adsorbţiei azotului pentru bentonitul Lărguţa corespunde tipului II de
izoterme după clasificarea BET (Fig.3). Ramura desorbţională păstrează alura ramurii sorbţionale până la
40 30 20 10 0
Na-, 25oC
Ca,Mg-, 25oC
7,17
3,31
3,57
2
5,03
7,17
10,1
10,1
13,8
15,6
Na-, 550oC
Na-, 350oC
20 15 10 5 0
40 30 20 10 0
40 30 20 10 02
2
3,31
3,57
5,03
7,17
10,1
19,2
10,1
7,17
5,07
4,96
Al-, 550oC
Al-, 450oC
Al-, 350oC
Al-, 25oC
16,6
15,2
17,3

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
178
valorile P/PS ~0,5, închizând histerezisul prin o treaptă bruscă. Asemenea histerezis este atribuit tipului H3 după clasificarea IUPAC [8], fiind caracteristic pentru mineralele argiloase. Acesta are o textură din particule lamelare paralel-orientate, ceea ce poate favoriza prezenţa pe suprafaţa exterioară a porilor secundari în formă de fisuri. După intercalarea bentonitului cu oligomeri de aluminiu, alura izotermelor şi histerezisului păstrează tipul II de izoterme după clasificarea BET şi tipul H3 de histerezis, evidenţiind totodată creşterea bruscă a ramurii sorbţionale pe sectorul iniţial (Fig.4).
Fig.3. Izoterma adsorbţiei azotului (77 K) pentru bentonitul Lărguţa (Na-forma,
fracţiunea <1 μm).
Fig.4. Izotermele adsorbţiei azotului (77 K) pentru bentonitul Lărguţa (Na-forma, fracţiunea <1 μm), intercalat cu oligomeri de aluminiu (obţinuţi la raportul molar iniţial OH
-/Al
3+ 2:1).
Mostre calcinate la diferite temperaturi.
Curbele distribuirii microporilor după dimensiuni pentru bentonitul Lărguţa, obţinute conform modelului Saito-Foley (SF [16]), indică predominarea microporilor cu dimensiuni (semilăţimea, x) de ordinul 5,5 Å, pe fondalul unor cantităţi nesemnificative de micropori cu dimensiuni în domeniul 7-9 Å (Fig.5). Intercalarea bentonitului cu oligomeri de aluminiu practic nu afectează structura microporoasă a adsorbanţilor, menţinân-du-se stabilă după calcinare la 450°C (Fig.5). Volumul microporilor, estimat după modelul SF şi conform modelului Dubinin-Radushkevich (DR [6,13]), concordă satisfăcător în majoritatea cazurilor (Tab.1). Ambele modele indică volumul maxim al microporilor atins după calcinare la 350°C, evidenţiind totodată stabilitatea termică şi după calcinarea adsorbanţilor la 450°C.
Distribuirea mezoporilor după dimensiuni, analizată conform modelului Barrett-Joyner-Halenda (BJH [2]), indică prezenţa mezoporilor cu razele efective de ordinul 20 Å atât pentru bentonitul iniţial, cât şi după inter-calare cu oligomeri de aluminiu (Fig.6). În comparaţie cu bentonitul iniţial, adsorbanţii intercalaţi posedă volume ale microporilor (modelele SF şi DR) considerabil mai mari, însă volume mai mici ale mezoporilor, conform modelului BJH (Tab.1).
Fig.5. Distribuirea microporilor după dimensiuni, obţinută
conform modelului Saito-Foley (SF). Bentonit Lărguţa iniţial (Na-Lar) şi intercalat cu oligomeri de aluminiu
la raportul molar iniţial OH-/Al
3+ 2:1 (Al-Lar).
Mostre calcinate la diferite temperaturi.
Fig.6. Distribuirea mezoporilor după dimensiuni, obţinută conform modelului Barrett-Joyner-Halenda (BJH).
Bentonit Lărguţa iniţial (Na-Lar) şi intercalat cu oligomeri de aluminiu la raportul molar iniţial OH
-/Al
3+ 2:1 (Al-Lar).
Mostre calcinate la diferite temperaturi.
0,0 0,2 0,4 0,6 0,8 1,0
10
20
30
40
50
60
70
80
90
VADS
, cm3/g
P/PS
0,0 0,2 0,4 0,6 0,8 1,0
0
10
20
30
40
50
60
70
80
90
0,0 0,2 0,4 0,6 0,8 1,0
0
10
20
30
40
50
60
70
80
90
0,0 0,2 0,4 0,6 0,8 1,0
0
10
20
30
40
50
60
70
80
90
450oC350
oC105
oC
VADS
, cm3/g
P/PS
P/PS
P/PS
0 4 8 12 16 20
0,000
0,002
0,004
0,006
0,008
0,010
0 4 8 12 16 20 0 4 8 12 16 20 0 4 8 12 16 20
7-9
5,5
Na-Lar, 105oC
ox, A
odV(r), cm
3/A/g
Al-Lar, 105oC
7-9
5,5
7-9
5,5
6
350oC 450
oC
10 20 30 40 50
0,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
10 20 30 40 50 10 20 30 40 50 10 20 30 40 50
odV(r), cm
3/A/g
Na-
Lar
, 105o C
20
20
o
Al-
Lar
, 105o C
2020
x, A
Al-
Lar
, 350o C
Al-
Lar
, 450o C
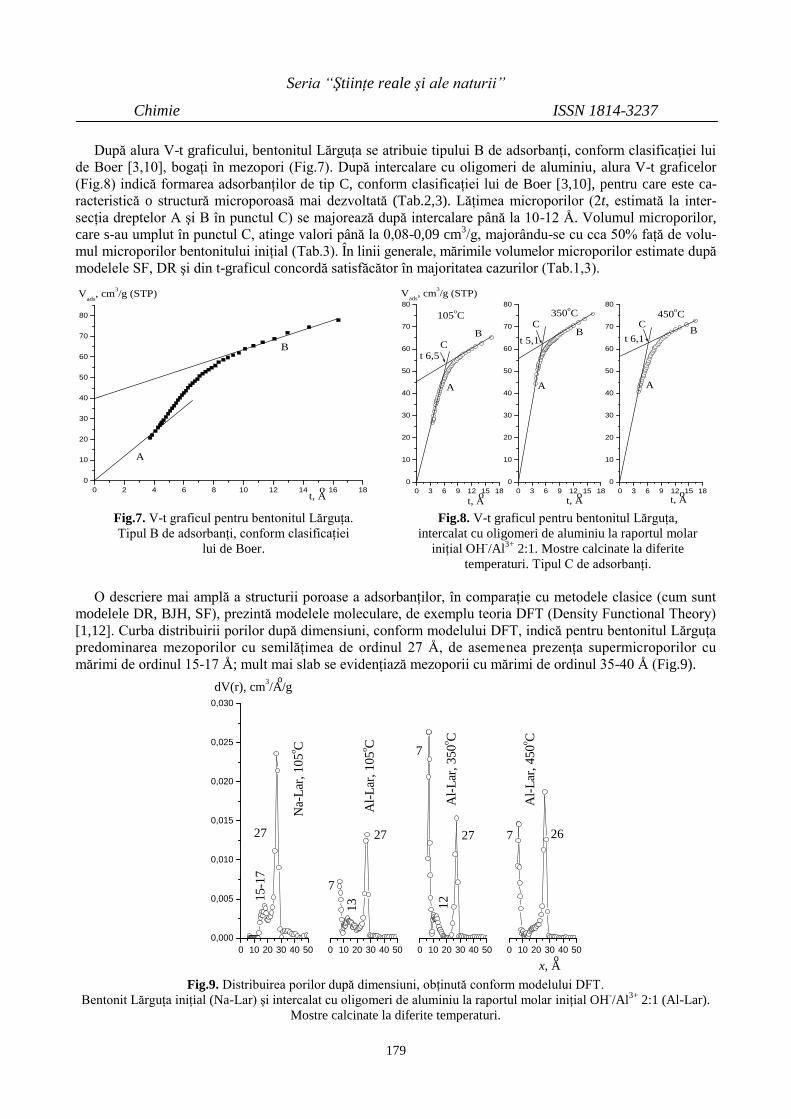
Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
179
După alura V-t graficului, bentonitul Lărguţa se atribuie tipului B de adsorbanţi, conform clasificaţiei lui
de Boer [3,10], bogaţi în mezopori (Fig.7). După intercalare cu oligomeri de aluminiu, alura V-t graficelor
(Fig.8) indică formarea adsorbanţilor de tip C, conform clasificaţiei lui de Boer [3,10], pentru care este ca-
racteristică o structură microporoasă mai dezvoltată (Tab.2,3). Lăţimea microporilor (2t, estimată la inter-
secţia dreptelor A şi B în punctul C) se majorează după intercalare până la 10-12 Å. Volumul microporilor,
care s-au umplut în punctul C, atinge valori până la 0,08-0,09 cm3/g, majorându-se cu cca 50% faţă de volu-
mul microporilor bentonitului iniţial (Tab.3). În linii generale, mărimile volumelor microporilor estimate după
modelele SF, DR şi din t-graficul concordă satisfăcător în majoritatea cazurilor (Tab.1,3).
Fig.7. V-t graficul pentru bentonitul Lărguţa.
Tipul B de adsorbanţi, conform clasificaţiei
lui de Boer.
Fig.8. V-t graficul pentru bentonitul Lărguţa,
intercalat cu oligomeri de aluminiu la raportul molar
iniţial OH-/Al
3+ 2:1. Mostre calcinate la diferite
temperaturi. Tipul C de adsorbanţi.
O descriere mai amplă a structurii poroase a adsorbanţilor, în comparaţie cu metodele clasice (cum sunt
modelele DR, BJH, SF), prezintă modelele moleculare, de exemplu teoria DFT (Density Functional Theory)
[1,12]. Curba distribuirii porilor după dimensiuni, conform modelului DFT, indică pentru bentonitul Lărguţa
predominarea mezoporilor cu semilăţimea de ordinul 27 Å, de asemenea prezenţa supermicroporilor cu
mărimi de ordinul 15-17 Å; mult mai slab se evidenţiază mezoporii cu mărimi de ordinul 35-40 Å (Fig.9).
Fig.9. Distribuirea porilor după dimensiuni, obţinută conform modelului DFT.
Bentonit Lărguţa iniţial (Na-Lar) şi intercalat cu oligomeri de aluminiu la raportul molar iniţial OH-/Al
3+ 2:1 (Al-Lar).
Mostre calcinate la diferite temperaturi.
0 2 4 6 8 10 12 14 16 18
0
10
20
30
40
50
60
70
80
A
B
ot, A
Vads
, cm3/g (STP)
0 3 6 9 12 15 18
0
10
20
30
40
50
60
70
80
0 3 6 9 12 15 18
0
10
20
30
40
50
60
70
80
0 3 6 9 12 15 18
0
10
20
30
40
50
60
70
80
t 6,5C
B
A
ot, A
Vads
, cm3/g (STP)
ot, A
A
Bt 5,1
C450
oC350
oC105
oC
A
B
ot, A
t 6,1
C
0 10 20 30 40 50
0,000
0,005
0,010
0,015
0,020
0,025
0,030
0 10 20 30 40 50 0 10 20 30 40 50 0 10 20 30 40 50
Na-
Lar
, 105
o C
27
15-1
7
7
7
7
27
13
Al-
Lar
, 105
o C
27
12
Al-
Lar
, 350
o C
26
o
ox, A
dV(r), cm3/A/g
Al-
Lar
, 450
o C

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
180
Intercalarea cu oligomeri de aluminiu duce la modificarea structurii poroase a bentonitului, micşorând de
cca 2 ori volumul mezoporilor cu dimensiuni de ordinul 26-27 Å, dar mai cu seamă a supermicroporilor de
ordinul 15-17 Å, care practic dispar după calcinare la 450°C. Totodată, se înregistrează majorarea volumului
microporilor de ordinul 7 Å. Volumul total al porilor este determinat din valoarea adsorbţiei la presiunea rela-
tivă P/PS apropiată de unitate. În linii generale, se constată o concordare satisfăcătoare între volumul porilor
(cumulativ), estimat după modelul DFT, şi volumul total al porilor estimat la P/PS 0,95 (Tab.1).
Tabelul 1
Caracteristica structurii poroase a adsorbanţilor, conform modelelor Saito-Foley (SF),
Dubinin-Radushkevich (DR), Barrett-Joyner-Halenda (BJH) şi DFT
Adsorbant
Modelul
SF
Modelul
DR
Modelul
BJH
Modelul
DFT
Volumul total
al porilor la
P/PS 0,95,
cm3/g
Proporţia*
mezo-
porilor, Vme,
%
Volumul
micro-
porilor,
cm3/g
Volumul
micro-
porilor,
cm3/g
Volumul
mezo-
porilor,
cm3/g
Volumul
porilor
(cumulativ),
cm3/g
Na-Lar, 105oC 0,061 0,050 0,115 0,127 0,120 65
Al-Lar, 105oC 0,069 0,063 0,064 0,104 0,101 48
Al-Lar, 350oC 0,091 0,089 0,061 0,121 0,117 40
Al-Lar, 450oC 0,079 0,074 0,073 0,114 0,113 48
* Vme, % = VBJH
me · 100 / (VSF
mi+ VBJH
me)
3. Analize comparative
Izotermele adsorbţiei azotului sugerează o structură preponderent mezoporoasă, atât pentru montmoriloni-
tul Ascangel (R. Georgia), caracterizat ca o formaţiune monominerală [14], cât şi pentru bentonitul Lărguţa
din Republica Moldova (Fig.3), constituit preponderent din montmorilonit. Pentru ambele formaţiuni argi-
loase, izotermele adsorbţiei azotului corespund tipului II de izoterme după clasificarea BET, cu histerezis de
tipul H3 după clasificarea IUPAC.
Intercalarea cu oligomeri de aluminiu modifică izotermele, evidenţiind majorarea structurii microporoase
a adsorbanţilor. Saltul pe sectorul iniţial al izotermelor (Fig.3,4) este un indiciu al prezenţei microporilor,
pentru cuantificarea lor fiind aplicată metoda t-graficului, în îmbinare cu metoda BET [8]. Suprafaţa micro-
porilor adsorbanţilor, estimată în aşa mod, este mai verosimilă, în comparaţie cu modelele DR şi Langmuir
care supraestimează suprafaţa microporilor, cum rezultă din compararea cu valorile suprafeţei cumulative
determinate din modelul DFT (Tab. 2,3).
Tabelul 2
Suprafaţa adsorbanţilor, conform modelelor Dubinin-Radushkevich (DR),
Barrett-Joyner-Halenda (BJH), DFT, BET şi Langmuir
Adsorbant
Modelul
DR
Modelul
BJH
Modelul
DFT
Modelul
BET
Modelul
Langmuir
Sme *,
din
modelele
DFT şi
t-graficul,
m2/g
Suprafaţa
micro-
porilor,
m2/g
Suprafaţa
mezo-
porilor,
m2/g
Suprafaţa
(cumulativă),
m2/g
Suprafaţa,
m2/g
Suprafaţa,
m2/g
Na-Lar, 105oC 141 97 111 118 136 32
Al-Lar, 105oC 179 52 178 152 172 47
Al-Lar, 350oC 250 46 257 213 242 65
Al-Lar, 450oC 209 69 221 178 202 59
* Sme = SDFT
– Smit
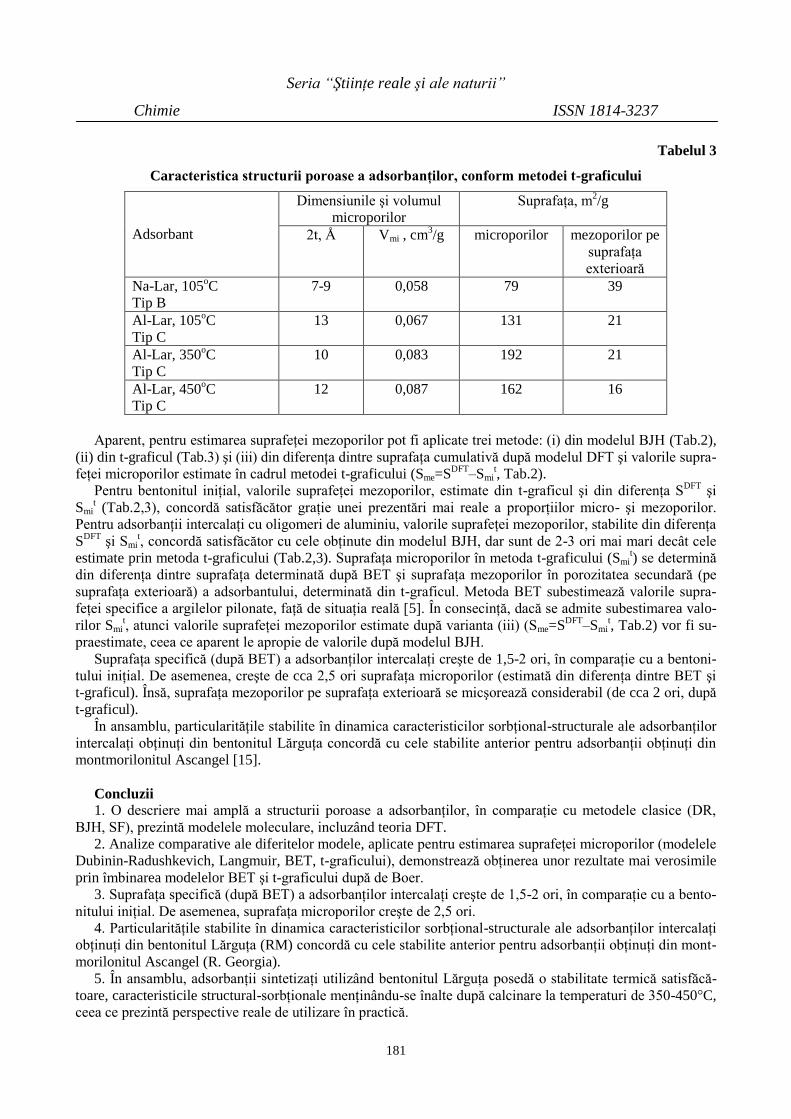
Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
181
Tabelul 3
Caracteristica structurii poroase a adsorbanţilor, conform metodei t-graficului
Adsorbant
Dimensiunile şi volumul
microporilor
Suprafaţa, m2/g
2t, Å Vmi , cm3/g microporilor mezoporilor pe
suprafaţa
exterioară
Na-Lar, 105oC
Tip B
7-9 0,058 79 39
Al-Lar, 105oC
Tip C
13 0,067 131 21
Al-Lar, 350oC
Tip C
10 0,083 192 21
Al-Lar, 450oC
Tip C
12 0,087 162 16
Aparent, pentru estimarea suprafeţei mezoporilor pot fi aplicate trei metode: (i) din modelul BJH (Tab.2),
(ii) din t-graficul (Tab.3) şi (iii) din diferenţa dintre suprafaţa cumulativă după modelul DFT şi valorile supra-
feţei microporilor estimate în cadrul metodei t-graficului (Sme=SDFT
–Smit, Tab.2).
Pentru bentonitul iniţial, valorile suprafeţei mezoporilor, estimate din t-graficul şi din diferenţa SDFT
şi
Smit (Tab.2,3), concordă satisfăcător graţie unei prezentări mai reale a proporţiilor micro- şi mezoporilor.
Pentru adsorbanţii intercalaţi cu oligomeri de aluminiu, valorile suprafeţei mezoporilor, stabilite din diferenţa
SDFT
şi Smit, concordă satisfăcător cu cele obţinute din modelul BJH, dar sunt de 2-3 ori mai mari decât cele
estimate prin metoda t-graficului (Tab.2,3). Suprafaţa microporilor în metoda t-graficului (Smit) se determină
din diferenţa dintre suprafaţa determinată după BET şi suprafaţa mezoporilor în porozitatea secundară (pe
suprafaţa exterioară) a adsorbantului, determinată din t-graficul. Metoda BET subestimează valorile supra-
feţei specifice a argilelor pilonate, faţă de situaţia reală [5]. În consecinţă, dacă se admite subestimarea valo-
rilor Smit, atunci valorile suprafeţei mezoporilor estimate după varianta (iii) (Sme=S
DFT–Smi
t, Tab.2) vor fi su-
praestimate, ceea ce aparent le apropie de valorile după modelul BJH.
Suprafaţa specifică (după BET) a adsorbanţilor intercalaţi creşte de 1,5-2 ori, în comparaţie cu a bentoni-
tului iniţial. De asemenea, creşte de cca 2,5 ori suprafaţa microporilor (estimată din diferenţa dintre BET şi
t-graficul). Însă, suprafaţa mezoporilor pe suprafaţa exterioară se micşorează considerabil (de cca 2 ori, după
t-graficul).
În ansamblu, particularităţile stabilite în dinamica caracteristicilor sorbţional-structurale ale adsorbanţilor
intercalaţi obţinuţi din bentonitul Lărguţa concordă cu cele stabilite anterior pentru adsorbanţii obţinuţi din
montmorilonitul Ascangel [15].
Concluzii
1. O descriere mai amplă a structurii poroase a adsorbanţilor, în comparaţie cu metodele clasice (DR,
BJH, SF), prezintă modelele moleculare, incluzând teoria DFT.
2. Analize comparative ale diferitelor modele, aplicate pentru estimarea suprafeţei microporilor (modelele
Dubinin-Radushkevich, Langmuir, BET, t-graficului), demonstrează obţinerea unor rezultate mai verosimile
prin îmbinarea modelelor BET şi t-graficului după de Boer.
3. Suprafaţa specifică (după BET) a adsorbanţilor intercalaţi creşte de 1,5-2 ori, în comparaţie cu a bento-
nitului iniţial. De asemenea, suprafaţa microporilor creşte de 2,5 ori.
4. Particularităţile stabilite în dinamica caracteristicilor sorbţional-structurale ale adsorbanţilor intercalaţi
obţinuţi din bentonitul Lărguţa (RM) concordă cu cele stabilite anterior pentru adsorbanţii obţinuţi din mont-
morilonitul Ascangel (R. Georgia).
5. În ansamblu, adsorbanţii sintetizaţi utilizând bentonitul Lărguţa posedă o stabilitate termică satisfăcă-
toare, caracteristicile structural-sorbţionale menţinându-se înalte după calcinare la temperaturi de 350-450°C,
ceea ce prezintă perspective reale de utilizare în practică.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
182
Referinţe:
1. AUTOSORB AS-1. AS1Win (21 CFR-Part 11 compliant). GAS SORPTION SYSTEM. OPERATION MANUAL.
Firmware: ver 2.55. AS1Win Software: ver 2.0 and newer. Quantachrome Instruments, 2008.
2. BARRETT, E.P., JOYNER, L.G., HALENDA, P.P. The Determination of Pore Volume and Area Distributions in
Porous Substances. I. Computations from Nitrogen Isotherms. In: J. Amer. Chem. Soc., 1951, vol.73, p.373.
3. de BOER, J.H., LIPPENS, B.C., LINSEN, B.C. In: J. Colloid. Interface Sci., 1966, vol.21, p.405.
4. BRUNAUER, S., EMMETT, P.H., TELLER, E. Adsorption of gases in multimolecular layers. In: J. Am. Chem.
Soc., 1938, vol.60, p.309.
5. COOL, P., VANSANT, E.F. Pillared Clays and Porous Clay Heterostructures. In: SCOTT, M., CARRAD, K.A.,
DUTTA, P.K., eds. Handbook of layered materials. Part II Other Layered Materials. New York Basel: MARCEL
DEKKER, Inc. 2004, p.261-311.
6. DUBININ, M.M. Active carbons. In: GREG, S.G., SING, K.S.W., STOECKLI, H.F. P. eds. Caracterisation of
Porous Solids. London: Soc. Chem. Ind., 1979, p.67-91.
7. GREGG, S.J., SING, W. Adsorption, Surface Area and Porosity. 1st ed. New York: Academic Press, 1967.
8. GREGG, S.J., SING, K.S.W. Adsorption, Surface Area and Porosity. Second edition. London: Academic Press,
1982.
9. LANGMUIR, I. The adsorption of gases on plane surfaces of glass, mica and platinum. In: J. Amer. Chem. Soc.,
1918, vol.40, p.1368.
10. LIPPENS, B.S., LINSEN, B.J., de BOER, J.H. In: J. Catalysis, 1964, vol.3, p.32.
11. MANJU, K., SUGUNAN, S. Selective methyl tert-butyl ether synthesis over metal exchanged pillared clays. In:
Journal of Porous Materials, 2005, vol.12, p.323-336.
12. ROQUE-MALHERBE, R.M.A. Adsorption and Diffusion in Nanoporous Materials. Taylor and Francis Group:
CRC Press, 2007.
13. ROUQUEROL, J., ROUQUEROL, F., SING, K. Adsorption by Powder and Porous Solids. New York: Academic
Press, 1999.
14. RUSU, V., NASTAS, R., OBREJA-NISTOR, L., MAFTULEAC, A., PETUHOV, O., PUŞCAŞU, B. Particularităţi
de sinteză a adsorbanţilor intercalaţi pe bază de montmorilonit. În: Buletinul Institutului de Geologie şi Seismologie
al AŞM, 2011, nr.1, p.94-105.
15. RUSU, V., MAFTULEAC, A., PETUHOV, O. Caracteristici sorbţional-structurale ale montmorilonitului intercalat
convenţional şi competitiv cu oligomeri de aluminiu. În: Studia Universitatis. Seria „Ştiinţe reale şi ale naturii”,
2012, nr.6(56), p.87-95. ISSN 1814-3237
16. SAITO, A., FOLEY, H.C. Curvature and parametric sensivity in models for adsorption in micropores. In: AIChE
Journal, 1991, vol.37, p.429.
17. SALERNO, P., MENDIOROZ, S. Preparation of Al-pillared montmorillonite from concentrated dispersions. In:
Applied Clay Science, 2002, vol.22, p.115-123.
18. SALERNO, P., MENDIOROZ, S., LOPEZ, A. Al-pillared montmorillonite-based Mo catalysts: effect of the
impregnation conditions on their structure and hydrotreating activity. In: Applied Clay Science, 2003, vol.23,
p.287-297.
19. SERGIO, M., MUSSO, M., MEDINA, J., DIANO, W. Aluminum-pillaring of a montmorillonitic clay: textural
properties as a function of the starting min. In: Advances in Technology of Materials and Materials Processing,
2006, vol.8(1), p.5-12.
Prezentat la 29.09.2013

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
183
PROPRIETĂŢI UTILE ALE UNOR COMPUŞI COORDINATIVI
ÎN BAZA LIGANZILOR DIOXIMICI
Eduard COROPCEANU
Institutul de Chimie al AŞM
Tendinţele contemporane în dezvoltarea socioeconomică implică un parteneriat viabil în domeniul cercetare-tehnologii.
Dioximaţii unor metale tranziţionale manifestă proprietăţi utile în diferite domenii: prezintă modele de molecule cu im-
portanţă fiziologică, sunt folosiţi drept catalizatori în procesele industriale, pot fi utilizaţi pentru obţinerea hidrogenului
etc. Proprietăţile utile variază în dependenţă de natura generatorului de complex, a liganzilor, a bioelementelor constitu-
ente, a structurii moleculare şi cristaline etc.
Cuvinte-cheie: dioximaţi, metale tranziţionale, proprietăţi utile, aplicare practică.
SOME USEFUL PROPERTIES OF COORDINATION COMPOUNDS BASED ON DIOXIME LIGANDS
The modern trends in socio-economic development involves a viable partnership research and technology areas.
Dioximates of some transition metals exhibit useful properties in various fields: serve as models of molecules with
important physiological impact, are used as catalysts in industrial processes, can be used for hydrogen production etc.
Useful properties vary depending on the nature of the central atoms, ligands, the constituent bioelements, molecular and
crystal structure etc.
Keywords: dioximates, transition metals, useful properties, practical application.
Introducere
Identificarea domeniilor de aplicare practică a noilor compuşi chimici este una dintre obiectivele impor-
tante ale ştiinţei contemporane axate pe transferul cunoştinţelor din sfera cercetării în cea a elaborării tehno-
logiilor moderne în baza noilor materiale. Compuşii coordinativi ai metalelor tranziţionale cu liganzi chelanţi
ocupă un loc important în chimia contemporană. Din această clasă fac parte şi dioximaţii metalelor tranziţionale.
Capacitatea de complexare a -dioximelor cu metale de tip d atrage atenţia cercetătorilor nu doar din perspec-
tiva sintezei modelelor vitaminei B12 sau a hemoglobinei [1], dar şi a spectrului larg de posibilităţi sintetice,
analitice şi structurale. Dioximaţii pot fi utilizaţi în calitate de catalizatori ai proceselor industriale [41], pre-
parate antihipoxice [38], preparate cu proprietăţi de antidot [40], pentru separarea şi purificarea metalelor
generatoare de complecşi etc. O importantă contribuţie în sinteza şi studiul dioximaţilor unor metale tranzi-
ţionale a adus-o şcoala de la Chişinău fondată de academicianul A.Ablov. A fost studiat comportamentul
unor reprezentanţi ai acestei serii în soluţii şi s-a constatat că, datorită stabilităţii înalte, fragmentul ecuatorial
Co(DioxH)2 nu se modifică, însă pot decurge reacţii de substituţie a liganzilor din poziţiile 1,6 [28,29,42].
Varietatea compoziţiei şi structurii dioximaţilor se datorează condiţiilor de sinteză diferite (pH-ul soluţiei
etc.), naturii liganzilor axiali, anionilor din sfera externă şi altor factori. Una dintre direcţiile promiţătoare de
utilizare a dioximaţilor reprezintă cercetarea proprietăţilor catalitice ale compuşilor coordinativi ai metalelor
în diferite procese chimice. Ca rezultat al acestor cercetări, s-a constatat că unii dioximaţi pot fi utilizaţi drept
catalizatori la vopsirea stofei. Aceştia au fost propuşi pentru utilizare la combinatele din Kalininsk, Iarţev,
Ternopol şi Tiraspol [35]. Dioximatul Co(III) cu nicotinamida a fost testat experimental în procesul de tratare
a anemiei hemolitice şi a altor maladii [39].
Diversitatea dioximaţilor metalelor tranziţionale este prezentată de compuşi mono- şi polinucleari, hetero-nucleari, cu structură di- şi polimerică etc. Dioximaţii mononucleari sunt bine cunoscuţi şi reprezintă compuşi
cu o stabilitate înaltă, oferită de pseudo-metalociclurile formate prin coordinarea monoanionilor dioximelor în planul ecuatorial, cimentate prin intermediul legăturilor de hidrogen intramoleculare dintre grupele oximice.
Ulterior s-a reuşit sinteza diferitelor tipuri de dioximaţi heteronucleari [2,43], în care iese în evidenţă capaci-tatea diversă a afinităţii de coordinare a metalelor tranziţionale faţă de atomul de azot sau oxigen al grupei
oximice. Un interes deosebit în domeniul designului molecular şi al obţinerii complecşilor cu compoziţie va-riată prezintă tris-dioximaţii clatratochelaţi ai unor metale tranziţionale [25], care se obţin prin unirea compu-
şilor borului(III), staniului(IV), germaniului(IV) etc. la trei radicali bideprotonaţi ai dioximelor, precum şi alte încercări de a diversifica compoziţia şi arhitectura reprezentanţilor acestei serii de complecşi. Trecerea

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
184
de la dioximaţii cu structură monomerică la cei cu structură di- şi polimerică este realizată pe baza utilizării diferitelor liganzi-punte care leagă atomii generatori de complex între ei [6]. Sinteza noilor dioxime, cu diverse
grupări funcţionale, care oferă noi posibilităţi pentru compoziţia moleculară şi arhitectura structurală, este una
dintre direcţiile de perspectivă ale domeniului [14]. Deoarece studiile au demonstrat că unii dioximaţi pot manifesta proprietăţi de stimulatori ai unor procese
biochimice, de catalizatori în procese industriale etc., prezintă interes stabilirea noilor domenii potenţiale de utilizare a reprezentanţilor acestei clase de compuşi coordinativi.
Rezultate şi discuţii Din punctul de vedere al aplicabilităţii practice a dioximaţilor metalelor tranziţionale, în prezent se relie-
fează clar mai multe direcţii în care reprezentanţii acestei clase pot fi utilizaţi cu succes: elaborarea modele-lor artificiale ale unor molecule biologice cu importanţă vitală; elaborarea biotehnologiilor eficiente pentru obţinerea preparatelor enzimatice utilizate în diferite ramuri ale industriei alimentare şi farmaceutice; obţinerea unor compuşi cu proprietăţi utile pentru medicină; sinteza materialelor cu proprietăţi fizice valoroase (semi-conductori, materiale fotoluminescente); obţinerea unor sisteme utilizate în producerea hidrogenului etc.
Utilizarea compuşilor coordinativi în calitate de stimulatori ai activităţii biologice a diferitelor organisme este una dintre direcţiile solicitate pentru soluţionarea unor probleme din medicină, industrie alimentară, agri-cultură etc. Biotehnologia modernă acordă o atenţie deosebită sintezei orientate a substanţelor bioactive de către microorganisme, care, datorită unor particularităţi specifice, ca: reacţie sporită la schimbările ambiante, metabolismul adaptiv, ciclul scurt de dezvoltare, sunt obiecte comode pentru diverse cercetări. Din alt punct de vedere, microorganismele sunt recunoscute ca surse economic avantajoase de obţinere a unei game largi de substanţe bioactive importante.
O clasă de substanţe biologic active cu o însemnătate deosebită în biologie, dar şi cu multiple aplicări prac-tice, sunt enzimele. Preparatele enzimatice cunosc o amplă utilizare în cele mai diverse procese tehnologice, precum şi în alte domenii de activitate socioumană. O importanţă majoră în industrie şi în medicină au hidro-lazele – enzimele care catalizează reacţiile de hidroliză ce stau la baza multor procese tehnologice moderne. Numeroasele implicări practice ale enzimelor hidrolitice (pectinaze, amilaze, celulaze, proteaze, lipaze) în diferite sfere economice şi sociale determină necesitatea elaborării unor procedee efective de sporire a capa-cităţii biosintetice a microorganismelor-producători. Din acest punct de vedere, prezintă interes utilizarea stimulatorilor de natură chimică, în special a compuşilor coordinativi ai metalelor tranziţionale [19]. Dintre toţi factorii mediului exterior, care conduc la modificarea proceselor biologice în celula microbiană, remar-cabil este mediul nutritiv. Anume compoziţia şi abundenţa mediului nutritiv reglează, mai mult decât oricare alt factor, dezvoltarea microorganismelor şi, implicit, activitatea de biosinteză. Sub acest aspect prezintă interes utilizarea compuşilor coordinativi în calitate de biostimulatori ai procesului de sinteză a enzimelor.
Însuşirile biologice ale metalocomplecşilor în mare parte pot fi atribuite prezenţei în componenţa lor a metalelor Fe, Cu, Mo, Co, Zn , Mn, Ni etc., care reprezintă microelemente legate nemijlocit cu atomi şi grupe de atomi din moleculele substanţelor organice. Aceste elemente, intrând în cantităţi foarte mici în compo-nenţa celulelor, joacă un rol important în activitatea lor vitală. Influenţa multilaterală a microelementelor este importantă prin participarea lor în reacţiile fine ale schimbului celular de substanţe, mai cu seamă în proce-sele fermentative. Сuprul, zincul, cobaltul, manganul, molibdenul, calciul, fierul participă în procesele de oxidoreducere care decurg în organismele vegetale şi animale, sunt parte componentă a unei serii de fermenţi oxidanţi importanţi, participă la metabolismul glucidic şi proteic în organisme. Microelementele, îndeosebi biometalele (precum: cobaltul, cuprul, zincul, manganul), participă la formarea sau activarea fermenţilor, vitaminelor, hormonilor, reglează schimbul de substanţe, determinând creşterea, dezvoltarea, înmulţirea, productivitatea şi calitatea producţiei. Ionii metalelor îndeplinesc o serie de funcţii: prezintă un grup electrofil al centrului activ al fermentului şi facilitează interacţiunea cu sectoarele cu sarcină negativă ale moleculelor substratului; formează o conformaţie catalitic activă a structurii unor molecule (la constituirea structurii în formă de spirală a ARN participă ionii de zinc şi de mangan); participă în transportul electronilor etc. Capaci-tatea ionului metalului de a forma centrul activ al fermentului depinde de capacitatea de coordinare a ionului metalului, geometria şi stabilitatea complexului format.
Un rol important în manifestarea proprietăţilor metalocomplecşilor joacă şi liganzii din componenţa lor care, datorită setului larg de atomi donori, creează cu ionii metalelor tranziţionale complecşi stabili, diferiţi după compoziţie, structură şi proprietăţi. Substanţele biologic active, incluse în compoziţia complecşilor metalelor în calitate de liganzi, sporesc esenţial eficienţa lor [30].

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
185
Sinteza complecşilor ce prezintă modele ale obiectelor biologice şi testarea acţiunii lor asupra proceselor
metabolice ale microorganismelor constituie o direcţie promiţătoare de reproducere şi dirijare a biosintezei
naturale. Este suficient de menţionat că hemoglobina (atomul central – Fe2+
), clorofila (atomul central – Mg2+
),
vitamina B12 (atomul central – Co2+
) sunt metalocomplecşi. Elementele legate coordinativ sunt mai puţin
toxice şi au o capacitate reagentă mai înaltă.
După cum s-a stabilit, compuşii coordinativi care conţin cobalt pot servi în calitate de stimulatori ai bio-
sintezei fermenţilor hidrolitici ai micromicetelor [37]. În urma testelor biologice, în cadrul cărora dioximaţii
Co(III) ce conţin fluor au fost introduşi în mediul nutritiv al microorganismelor, s-a constatat că ei manifestă
proprietăţi de stimulatori ai biosintezei vitaminei B12 de către alga Spirulina platensis [33]. Aceste studii au
creat premise pentru iniţierea sintezelor în vederea elaborării analogilor sintetici ai sistemelor naturale, mode-
lării moleculelor biologice şi analizei influenţei lor asupra proceselor ce decurg în celulă.
Analiza datelor din literatură a arătat că biomasa de Spirulina platensis, obţinută prin cultivarea în prezenţa
compuşilor coordinativi ai cobaltului, se caracterizează printr-un conţinut înalt de ciancobalamină, proteină,
carotenoizi şi este balansată după celelalte componente [21]. De asemenea, s-a constatat că compuşii cobaltu-
lui cu fluorul posedă proprietăţi stimulatoare asupra unor microorganisme. Formarea compuşilor cu cobaltul
diminuează acţiunea toxică a ionilor de fluor prezenţi în mediul nutritiv. Complexitatea structurală şi de com-
poziţie, prezenţa metalelor în calitate de atom central atestă perspectiva utilizării compuşilor coordinativi în
calitate de stimulatori şi reglatori ai proceselor biologice în celula microbiană.
A fost realizată sinteza orientată a unei serii de dioximaţi ai Co(III) cu formula generală [Co(DioxH)2L2]nX nH2O,
în care DioxH este radicalul dioximei: dimetilglioxima (DH2), metilglioxima (MH2), 1.2-ciclohexandiondioxima
(NioxH2), -benzildioxima (DfH2); L – tiocarbamida (Thio), piridina (Py), anilina (An), nicotinamida (PP),
sulfanilamida (Sam); X – [BF4]-, [PF6]
-, [SiF6]
2-, [ZrF6]
2-, [TiF6]
2- etc., structura cristalină şi caracteristicile
spectrale ale cărora au dovedit configuraţia trans-octaedrică a complecşilor. Au fost studiate proprietăţile chi-
mice şi fizice. S-a constatat că în formarea structurii cristaline un rol important au anionii din sfera externă şi
moleculele de apă de cristalizare. Complecşii analizaţi au fost utilizaţi în calitate de adaos în mediul nutritiv
al unor tulpini de fungi. Testarea dioximaţilor cobaltului(III) în cadrul Laboratorului Enzimologie al Institu-
tului de Microbiologie şi Biotehnologie al AŞM pe tulpinile genurilor Aspergillus, Rhizopus, Penicillium,
Fusarium a demonstrat că aceşti complecşi influenţează benefic desfăşurarea unor procese fiziologice la
fungii respectivi [11,13,31].
Complexitatea structurală şi de compoziţie, prezenţa metalelor în calitate de atom central atestă perspectiva
utilizării compuşilor coordinativi drept stimulatori şi reglatori ai proceselor biologice în celula microorganis-
melor. Includerea în componenţa aceluiaşi complex a mai multor microelemente (spre exemplu: Co, Si, F;
Co, P, F; Co, B, F sau Co, Ti, F) contribuie la manifestarea proprietăţilor de biostimulatori de către dioximaţii
respectivi. Cobaltul este un element necesar sistemelor biologice. Alte elemente sunt la fel de necesare pentru
organisme. Borul, bunăoară, participă la reglarea funcţiilor membranelor, a biosintezei acizilor nucleici. Tita-
nul este un element indispensabil al multor organisme, având funcţii cu importanţă vitală: sporeşte eritropoeza,
catalizează sinteza hemoglobinei, imunogeneza [36]. Complecsonaţii titanului sunt nu doar agenţi de stimu-
lare a fagocitozei, dar şi substanţe ce activează reacţiile de imunitate celulară şi umorală. Chelaţii titanului
influenţează, precum se ştie, asupra funcţiilor de reproducere a unor specii animale. Experimental s-a demon-
strat că titanul este un element care se elimină uşor din organism şi nu prezintă pericolul acumulării în canti-
tăţi mari. Mai mult chiar, fiind considerat un element biocompatibil, se utilizează în medicină pentru unirea
regiunilor fracturate ale ţesutului osos.
Rezultatele obţinute în baza testării influenţei dioximaţilor Co(III) asupra proceselor fiziologice ale unor
tulpini de micromicete permit formularea concluziei că din seria complecşilor analizaţi pot fi selectaţi:
biostimulatori ai proceselor enzimogenetice ale fungilor (spre exemplu: complexul
[Co(DH)2(Thio)2]2[SiF6] 3H2O sporeşte activitatea pectolitică a fungilor Rhizopus arrhizus cu 184,08%
faţă de control) [34];
catalizatori ai proceselor de acumulare a biomasei (spre exemplu: la introducerea compusului
[Co(DH)2(Thio)2][BF4] 3H2O în concentraţie de 5-10 mg/l în mediul de cultivare a tulpinii de fungi
Aspergillus niger 412 s-a înregistrat o sporire dublă a productivităţii biomasei – cu 188,3-197,07% faţă
de control);

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
186
stabilizatori ai proceselor biochimice cu importanţă vitală în condiţii nefavorabile (spre exemplu: în condiţii termice nefavorabile, introducerea complexului [Co(DH)2(Py)2][BF4] în mediul nutritiv al fungilor Aspergillus niger 412 condiţionează sporirea procesului de acumulare a biomasei cu 132,93% faţă de control) [4];
acceleratori ai dezvoltării biologice a microorganismelor, reducând ciclul tehnologic cu 24-48 ore, adică până la ~30% din durata întregului ciclu (la introducerea complexului [Co(DH)2(An)2]2[TiF6] în mediul nutritiv al micromicetei Aspergillus niger 33-19 CNMN FD 02A), fapt ce prezintă interes din punct de vedere economic [5].
Efectul exercitat de metalocomplecşi variază în funcţie de originea microorganismului, tipul sistemului enzimatic sintetizat, componenţa şi concentraţia compusului. Sumând rezultatele înregistrate, putem con-cluziona că dioximaţii testaţi manifestă proprietăţi de stimulatori (acumularea biomasei, sporirea activităţii enzimatice), stabilizează procesele biosintetice în condiţii nefavorabile de activitate, reduc ciclul tehnologic, proprietăţi care sunt influenţate de prezenţa în calitate de generator de complex a ionului Co
3+, precum şi de
natura liganzilor din sfera internă şi a anionului din sfera externă. Aceste rezultate experimentale creează premise favorabile pentru elaborarea unor tehnologii economic avantajoase de cultivare a unor tulpini de fungi cu importanţă industrială.
Pentru elucidarea mecanismului de acţiune a metalocomplecşilor asupra capacităţii de producere a tulpinii
Aspergillus niger 33-19 s-a recurs la studiul influenţei separate a componentelor compuşilor coordinativi în
comparaţie cu complexul. Componentele metalocomplecşilor au fost adăugate la mediul de cultivare în con-
centraţii echivalente concentraţiei lor în compus. S-a constatat că sarea de cobalt, ligandul ecuatorial şi cel
axial, introduşi separat în mediul nutritiv, nu contribuie la sporirea activităţii amilolitice în comparaţie cu mar-
torul, pe când în cazul introducerii compusului coordinativ are loc o sporire considerabilă, fapt ce permite for-
mularea concluziei că efectul benefic este cauzat şi de acţiunea cumulativă a componentelor complexului [22].
Chiar dacă liganzii din poziţiile 1,6 pot fi supuşi unor procese de solvatare, fragmentul ecuatorial care se ca-
racterizează prin stabilitate înaltă nu se distruge. Eficacitatea cea mai înaltă este înregistartă pentru concen-
traţiile 1-15 mg/l de complex, fapt ce poate fi legat de influenţa pozitivă a fluorului în cantităţi mici şi de ac-
ţiunea lui toxică odată cu creşterea concentraţiei. Dintre anionii fluoruraţi cel mai înalt efect se înregistrează
pentru cei care, în afară de fluor, mai conţin un bioelement: BF4-, PF6
-, TiF6
2-.
S-a hotărât de a verifica influenţa dioximaţilor cobaltului(III) asupra proceselor fiziologice la unele
alge. În rezultatul testării capacităţii de stimulare a unor procese biosintetice la alga roşie Porphyridium
cruentum s-a constatat că complecşii studiaţi manifestă efect stimulator, cel mai înalt fiind înregistrat
pentru [Co(DH)2(Thio)2]2F[PF6]. De asemenea, s-a stabilit că acumularea produselor oxidării lipidelor în
biomasă, urmată de intensificarea biosintezei lipidelor, determină capacitatea celulelor de Porphyridium
cruentum de a-şi menţine viabilitatea [20]. A fost studiată şi acţiunea unor dioximaţi ai Co(III) asupra culturilor unor plante superioare: sfecla de
zahăr, porumbul etc. Efectul pozitiv al complexului asupra sfeclei de zahăr duce la majorarea conţinutului de pigmenţi asimilatori, a recoltei şi a producţiei totale de zahăr la unitate de suprafaţă. La tratarea semin-ţelor de sfeclă de zahăr înainte de semănat şi a plantelor în timpul creşterii vegetative cu soluţie apoasă de [Co(DH)2(PP)2][BF4]·2H2O în concentraţie de 0,001% are loc sporul recoltei de rizocarpi cu 11,5% com-parativ cu martorul şi cu 6,1% comparativ cu sporul recoltei asigurat de tratarea cu cea mai apropiată soluţie tehnică [12].
Unii dioximaţi de fier(II) şi fier(III) manifestă proprietăţi bine pronunţate de catalizatori în reacţiile de for-mare a compoziţiilor poliuretanice, destinate lipirii unor piese metalice în industria de construcţie a maşinilor şi fabricării materialelor plastice decorative. Compoziţiile pe baza dioximaţilor de fier se caracterizează prin viabilitate, proprietăţi fizico-mecanice, adeziune şi stabilitate împotriva distrucţiei termooxidante ridicate [3].
Compuşii cis-[MX2(NioxH2)2](M=Co(II), Ni(II), Cu(II); X=Cl-), [Cu
II(NioxH2)] şi [Fe(DfH)2(DMSO)2]
posedă proprietăţi de material dielectric bine pronunţate şi pot găsi aplicare practică în electrotehnică şi micro-electronică.
Reţelele metal-organice reprezintă materiale multifuncţionale luminescente, deoarece atât resturile orga-nice, cât şi cele anorganice pot oferi platforme ce generează luminescenţa. Dioximaţii di- şi polimerici ai zincului şi cadmiului manifestă proprietăţi luminescente (Fig.1) [7], iar în cavităţile intermoleculare ale unor complecşi din această serie sunt înglobate molecule necoordinate, fapt ce permite presupunerea că aceste substanţe ar putea fi precursorii unei clase de materiale cu proprietăţi adsorbante.

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
187
Complecşii macrobiciclici celulari cu ion de metal incapsulat
(clatratochelaţi) constituie o clasă relativ nouă de compuşi care
posedă proprietăţi chimice şi fizice deosebite. A fost realizată
sinteza unei serii de tris-dioximaţi clatratochelaţi în calitate de
potenţiale preparate în terapia cancerului. Reacţia de autoasam-
blare a clatratochelaţilor şi interacţiunea lor cu acizii nucleici
poate fi utilizată pe larg în imunologie şi în biologia moleculară
[16,32].
În ultimul timp, sunt publicate o serie de lucrări în care se
descriu posibilităţile de utilizare a dioximaţilor metalelor tranzi-
ţionale în procesele de descompunere a apei în cadrul fotosinte-
zei artificiale [15]. Dioximaţii cobaltului(III) pot fi utilizaţi în
procesul de obţinere electrocatalitică a hidrogenului [8-10,18,23,
26,27]. Cobaloximele prezintă una dintre cele mai reuşite clase de compuşi sintetici ai metalelor tranziţionale
cunoscute în producerea hidrogenului, care pot fi relativ uşor sintetizate, sunt stabile faţă de oxigen, se pot
cupla în sistemele fotosintetice naturale şi artificiale [17,24].
Concluzii
Dioximaţii metalelor tranziţionale prezintă complecşi cu arhitectură moleculară stabilă şi, în funcţie de
natura generatorului de complex, a liganzilor, a bioelementelor constituente, a structurii moleculare şi a
reţelei cristaline, manifestă diferite proprietăţi chimice, fizice, biologice care-şi pot găsi aplicare în diverse
ramuri ale economiei.
Bibliografie:
1. BRESCIAN-PAHOR, N., FARCOLIN, M., MARZILLI, L.G. et al. Organocobalt B12 models: axial ligand effects
on the structural and coordination chemistry of cobaloximes. In: Coord. Chem. Rev., 1985, vol.63, p.1-125.
ISSN: 0010-8545
2. BURDINSKI, D., BIRKELBACH, F., WEYHERMULLER, T. et al. Encapsulation by Chromium(III)-Containing
Bicyclic Ligand Cage. Synthesis, Structure and Physical Properties of Heterometal Complexes. In: Inorg. Chem.,
1998, 37, p.1009-1020. ISSN 0020-1669
3. BULHAC, I. Sinteza, proprietăţile fizico-chimice şi structura compuşilor coordinativi de fier, cobalt, nichel şi
cupru cu -dioxime / Autoreferat al tezei de doctor habilitat în chimie. Chişinău, 2000.
4. COROPCEANU, E., BOLOGA, O., DESEATNIC, A. et al. Сobalt(III) dioximate fluorine containing compounds
as stabilizers of biosynthesis processes. In: Buletinul Institutului Politehnic din Iaşi, T. XLIX (LIII), fasc. 5, 2003,
p.293-298. ISSN 1223-8139
5. COROPCEANU, E., DESEATNIC, A., RIJA, A. et al. The synthesis and the study of the biological activity of some
cobalt(III) dioximates with fluorine. In: Chemistry Journal of Moldova, 2008, vol.3, no.2, p.70-80. ISSN 1857-1727
6. CROITOR, L., COROPCEANU, E., JEANNEAU, E. et al. Anion-induced generation of binuclear and polymeric
Cd(II) and Zn(II) coordination compounds with 4,4-bipyridine and dioxime ligands. In: Crystal Growth & Design,
2009, vol.9, p.5233-5243. ISSN 1528-7483
7. CROITOR, L., COROPCEANU, E., SIMINEL, A. et al. Synthesis, structures, and luminescence properties of mixed
ligand Cd(II) and Zn(II) coordination compounds mediated by 1,2-bis(4-pyridyl)ethane. In: Inorganica Chimica
Acta, 2011, 370, p.411-419. ISSN: 0020-1693
8. DEMPSEY, J.L., BRAUNSCHWING, B.S., WINKLER, J.R. et al. Hydrogen Evolution Catalyzed by Cobaloximes.
In: Accounts of chemical research, 2003, vol.42, no.12, p.1995-2004. ISSN 0001-4842
9. DEMPSEY, J.L., WINKLER, J.R., GRAY, H.B. Kinetics of Electron Transfer Reactions of H2-Evolving Cobalt
Diglyoxime Catalysts. In: J. Am. Chem. Soc., 2010. 132 (3), p.1060-1065. ISSN 0002-7863
10. DOLGANOV, A.V., BELOV, A.S., NOVIKOV, V.V. et al. Iron vs. cobalt clathrochelate electrocatalysts of HER:
the first example on a cage iron complex. In: Dalton Trans., 2013, 42, p.4373-4376. ISSN 1477-9226
11. INSTITUTUL DE CHIMIE AL AŞM. Fluorura-hexafluorofosfat-bis[di(tiocarbamid)bis(dimetilglioximato)cobalt(III)],
care posedă proprietăţi de biostimulator. Brevet MD nr.2833. Inventatori: Gărbălău N., Simonov Yu., Bouroş P. et
al. Publ. BOPI, 2005, nr.8.
12. INSTITUTUL DE GENETICĂ, FIZIOLOGIE ŞI PROTECŢIE A PLANTELOR AL AŞM. Procedeu de cultivare
a sfeclei de zahăr. Brevet MD nr.510. Inventatori: Ştefîrţă A., Bulhac I., Lisnic S. ş. a. Publ. BOPI, 2012, nr.5.
Fig.1. Fragment al lanţului polimeric în
{[Zn(NioxH2)(SO4)(bpy)]·0.5H2O·DMF}n.

S T U D I A U N I V E R S I T A T I S M O L D A V I A E
Revistă Ştiinţifică a Universităţii de Stat din Moldova, 2013, nr.6(66)
188
13. INSTITUTUL DE MICROBIOLOGE AL AŞM. Mediu nutritiv pentru cultivarea tulpinii de fungi Aspergillus
niger 33-19 CNMN FD 02A. Brevet MD nr.3943. Inventatori: Deseatnic A., Stratan M., Coropceanu E. ş.a. Publ.
BOPI, 2009, nr.7.
14. KURTOGLU, M., ISPIR, E., KURTOGLU, N., SERIN, S. Novel vic-dioximes: Synthesis, complexation with
transition metal ions, spectral studies and biological activity. In: Dyes and Pigments, 2008, vol.77, no.1, p.75-80.
ISSN 0143-7208
15. MCCORMIC, T.M., CALITREE, B.D., ORCHARD, A. et al. Reductive Side of Water Splitting in Artificial Photo-
synthesis: New Homogeneous Photosystems of Great Activity and Mechanistic Insight. In: J. Am. Chem. Soc.,
2010, 132 (44), p.15480-15483. ISSN 0002-7863
16. MOKHIR, A., KRAMER, R., VOLOSHIN, Y.Z., VARZATSKII, O.A. Synthesis and DNA binding properties
of dioxime–peptide nucleic acids. In: Bioorganic & Medicinal Chemistry Letters, 2004, 14, 11, p.2927-2930.
ISSN: 0960-894X
17. MULFORT, K.L., TIEDE, D.M. Supramolecular cobaloxime assemblies for H2 photocatalysis: an initial solution
state structure-function analysis. In: J. Phys. Chem. B., 2010, 114 (45), p.14572-14581. ISSN 1520-6106
18. NIKLAS, J., MARDIS, K.L., RAKHIMOV, R.R. et al. The Hydrogen Catalyst Cobaloxime – a Multifrequency
EPR & DFT Study of Cobaloxime’s Electronic Structure. In: J. Phys. Chem. B., 2012, 116(9), p.2943-2957.
ISSN 1520-6106
19. RUDIC, V. Aspecte noi ale biotehnologiei moderne. Сhişinău: Ştiinţa, 1993. 140 р. ISBN 9789975989251
20. RUDIC, V., CEPOI, L., RUDI, L. ş.a. Acţiunea compuşilor coordinativi ai cobaltului cu dioximele asupra unor
procese biosintetice la alga roşie Porphyridium cruentum. În: Buletinul AŞM. Ştiinţele vieţii, 2012, nr.1, p.144-151.
ISSN 1857-064X
21. RUDIC, V., CODREANU, S., GULEA, A., NOVIŢCHI, G. Influenţa compuşilor coordinativi ai metalelor asupra
biosintezei ciancobalaminei şi porfirinelor de către propionibacterii. Buletinul AŞM. Ştiinţele vieţii, 1995, nr.4, p.23-26.
ISSN 0568-5192
22. STRATAN, M. Biotehnologii de cultivare a tulpinii Aspergillus niger 33-19 CNMN FD 02A – producător de
amilaze / Autoreferat al tezei de doctor în biologie. Chişinău, 2012.
23. VALDEZ, C.N., DEMPSEY, J.L., BRUNSCHWIG, B.S. et al. Catalytic hydrogen evolution from a covalently
linked dicobaloxime. In: Proceedings of the National Academy of Sciences of USA, 2012, vol.109, no.39, p.15589-
15593. ISSN 1091-6490
24. VOLOSHIN, Y.Z., BELOV, A.S., VOLOGZHANINA, A.V. et al. Synthesis, structure, properties and immobiliza-
tion on a gold surface of the monoribbed-functionalized tris-dioximate cobalt(II) clathrochelates and an electro-
catalytic hydrogen production from H+ ions. In: Dalton Trans., 2012, 41(20), p.6078-93. ISSN 1477-9226
25. VOLOSHIN, Y.Z., KOSTROMINA, N.A., KRAMER, R. Clathrochelates: synthesis, structure and properties.
Amsterdam: Elsevier, 2002. 419 p. ISBN 0-444-51223-3
26. WEN, F., YANG, J., XU, Z. et al. Photocatalytic H2 production on hybrid catalyst system composed of inorganic
semiconductor and cobaloximes catalysts. In: Journal of Catalysis, 2011, 281, p.318-324. ISSN 0021-9517
27. ZHANG, P., JACQUES, P.A., CHAVAROT-KERLIDOU, M. et al. Phosphine coordination to a cobalt diimine-
dioxime catalyst increases stability during light-driven H2 production. In: Inorg. Chem., 2012, 51(4), p.2115-2120.
ISSN 0020-1669
28. АБЛОВ, А.В., БОВЫКИН, Б.А. Изучение равновесия диоксиминов кобальта(III), содержащих тиомочевину.
В: Журнал неорганической xимии, 1965, том.10, №1, с.53-60. ISSN 0044-457Х
29. АБЛОВ, А.В., САМУСЬ, Н.М., ДАМАСКИНА, О.Н. Кинетика замещение мочевины на воду в диоксиминах
кобальта(III). В: Журнал неорганической xимии, 1971, том.16, №7, с.1939-1942. ISSN 0044-457Х
30. АЗИЗОВ, М.А. О комплексных соединениях некоторых микроэлементов с биоактивными веществами.
Ташкент: Медицина, 1969. 199 с. ISBN 5-0825323-А
31. БОУРОШ, П.Н., КОРОПЧАНУ, Э.Б., ДЕСЯТНИК, А.А. и др. Супрамолекулярная организация структуры
кристаллов соединения [Co(DH)2(PP)2][BF4].2H2O и его биологические свойства. В: Координационная химия,
2009, том.35, №.10, с.761-767. ISSN 0132-344X
32. ВОЛОШИН, Я.З., ВАРЗАТСКИЙ, О.А., БУБНОВ, Ю.Н. Клеточные комплексы переходных металлов в
биохимии и медицине. В: Известия АН. Серия «Химия», 2007, №4, с.555-582. ISSN 0002-3353
33. ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ МОЛДОВЫ. -пероксо-бис[бис(диметил-глиоксимато)
аквакобальта(III)] гидрофторидгидрат, проявляющий свойства стимулятора биосинтеза витамина B12
сине-зеленой микроводорослью Spirulina platensis. А.С. №1616111 (СССР). Изобретатели: Гуля A.П.,
Рудик В.Ф., Гэрбэлэу Н.В. и др., 1990.
34. ДЕСЯТНИК, А.А., ГЭРБЭЛЭУ, Н.В., КОРОПЧАНУ, Э.Б и др. Использование диметилглиоксиматов
Со(III) при биосинтезе пектиназ Rhizopus arrhizus. Координационная химия, 2002, том.28, №2, с.144-145.
ISSN 0132-344X

Seria “Ştiinţe reale şi ale naturii”
Chimie ISSN 1814-3237
189
35. ЕГОРОВА, 3.Н., ПРОСКИНА, Н.Н., ХОРОШУН, И.В., БУЛУШЕВА, И.Е. Исследование взаимодействия
диоксиминов кобальта(III) с восстановителями кубовых красителей. В: Известия ВУЗов. Химия и химическая
технология, 1984, том.27, №4, с.393-396. ISSN 0579-2991
36. ЕРШОВ, Ю.А., ПОПКОВ, В.А., БЕРЛЯНД, А.С. и др. Общая химия. Биофизическая химия. Химия биогенных
элементов. Москва: Высшая школа, 2000. 560 с. ISBN 5-06-003626-X
37. КРИСС, Е.Е., ВОЛЧЕНCКОВА, И.И., ГРИГОРЬЕВА, А.С. и др. Координационные соединения металлов в
медицине. Киев: Наукова Думка, 1986. 216 с. ISBN 5-1615647
38. МАТКОВСКИЙ, К.Л. О противогипоксических свойствах диоксиминов кобальта. В: Известия АН МССР.
Серия «Биологические и химические науки». 1971, №5, с.88-89. ISSN 1857-064X
39. МАТКОВСКИЙ, К.Л., БАЛАН, Н.А., БАТЫР, Д.Г. и др. Изучение физиологической активности диокси-
минов кобальта(III). В: Известия АН МССР. Серия «Биологические и химические науки». 1969, №4, с.3-5.
ISSN 1857-064X
40. МАТКОВСКИЙ, К.Л., БОЛОГА, О.А. Об антидотных свойствах диоксиматов трехвалентного кобальта. В:
Глобус науки, 2006, том.6, с.34-36. ISSN 1561-4190
41. РОГАЧЕВ, Б.Г.; ХИНДЕКЕЛЬ, М.Л. Селективное восстановление бутадиена в присутствии бисдиметил-
глиоксиматного комплекса родия. В: Известия АН СССР, 1969, №1, с.141-142. ISSN 1066-5285
42. САМУСЬ, Н.М., ДАМАСКИНА, О.Н., ЛУКЬЯНЕЦ, Т.С. Реакции замещения в координационных соедине-
ниях кобальта. Кишинев: Штиинца. 1979. 167 с.
43. САМУСЬ, Н.М., ЮЩЕНКО, С.П., ХОРОШУН, И.В. и др. Железосодержащие трисдиоксиминаты кобальта(III).
В: Журнал неорганической xимии, 1989, том.34, №4, с.892-897. ISSN 0044-457Х
Prezentat la 25.09.2013