cercetĂri privind selecŢia asistatĂ de markeri … · clamidosporii sunt de culoare neagră,...
TRANSCRIPT

1
UNIVERSITATEA DE ŞTIINŢE AGRICOLE
ŞI MEDICINĂ VETERINARĂ DIN CLUJ-NAPOCA
FACULTATEA DE AGRICULTURĂ
ŞCOALA DOCTORALĂ
ING. DIPL. COŢA LAURA CRISTINA
CERCETĂRI PRIVIND SELECŢIA ASISTATĂ
DE MARKERI MOLECULARI PENTRU REZISTENŢA
GRÂULUI (TRITICUM AESTIVUM L.) LA MĂLURA COMUNĂ
(TILLETIA SPP.)
(REZUMATUL TEZEI DE DOCTORAT)
CONDUCĂTOR ŞTIINŢIFIC PROF. DR. CONSTANTIN BOTEZ
CLUJ-NAPOCA 2011

0
CUPRINS
INTRODUCERE................................................................................................................................ 3 CAPITOLUL I CONSIDERAŢII GENERALE PRIVIND IMPORTANŢA ŞI CULTIVAREA GRÂULUI...........................................................................................................................................
3 1.1. GENOMUL GRÂULUI................................................................................................................ 3 1.2. ASPECTE GENERALE PRIVIND AGENŢII PATOGENI CARE CAUZEAZĂ PRINCIPALELE BOLI LA GRÂU.....................................................................................................
3
CAPITOLUL II STUDII CU PRIVIRE LA SPECIILE DE TILLETIA CARE PROVOACĂ MĂLURA COMUNĂ A GRÂULUI...................................................................................................................
4 2.1. SPECIILE GENULUI TILLETIA. TAXONOMIE. MORFOLOGIE........................................... 4 2.2. BIOLOGIA MĂLURII COMUNE............................................................................................... 4 2.3. RASE FIZIOLOGICE DE MĂLURĂ........................................................ .......................... 4 2.4. SIMPTOMELE PRODUSE LA PLANTELE INFECTATE........................................................ 5 2.5. STRATEGII DE COMBATERE A MĂLURII COMUNE.......................................................... 5 CAPITOLUL III ASPECTE GENERALE PRIVIND REZISTENŢA GRÂULUI LA MĂLURA COMUNĂ......
6
3.1. GENETICA REZISTENŢEI PLANTELOR LA BOLI................................................................ 6 3.1.1. Tipuri de rezistenţă............................................................................................................ 6
3.2. CONTROLUL GENETIC AL REZISTENŢEI GRÂULUI LA MĂLURA COMUNĂ.............. 6 3.3. SURSE DE REZISTENŢĂ A GRÂULUI LA SPECIILE DE TILLETIA.................................... 6 CAPITOLUL IV MARKERII MOLECULARI CA NOI METODE UTILIZATE ÎN AMELIORAREA PENTRU REZISTENŢA GRÂULUI LA BOLI.............................................................................
7 4.1. TIPURI DE MARKERI MOLECULARI..................................................................................... 7 4.2. AMELIORAREA PLANTELOR PENTRU REZISTENŢA LA BOLI CU AJUTORUL MARKERILOR MOLECULARI........................................................................................................ 7
4.2.1. SELECŢIA ASISTATĂ DE MARKERI MOLECULARI............................................... 7 CAPITOLUL V SCOPUL ŞI OBIECTIVELE CERCETĂRILOR.......................................................................... 8 CAPITOLUL VI MATERIAL ŞI METODE...............................................................................................................
8
6.1. MATERIAL BIOLOGIC UTILIZAT PENTRU IDENTIFICAREA DE MARKERI DE REZISTENŢĂ A GRÂULUI LA MĂLURA COMUNĂ................................................................... 8 6.2. METODE UTILIZATE................................................................................................................. 9
6.2.1. Metoda cosegregării.............................................................................................................. 9 6.2.2. Metoda segregării în amestec (BSA).................................................................................... 9 6.2.3. Izolarea ADN-ului genomal.................................................................................................. 9 6.2.4. Izolarea ADN-ului plasmidial............................................................................................... 9 6.2.5. Determinarea cantităţii şi a calităţii ADN-ului izolat........................................................... 9
6.3. METODE DE ANALIZĂ MOLECULARĂ................................................................................ 9 6.3.1. Analiza cu primeri specifici de rezistenţă pentru gena Bt10................................................. 9 6.3.2. Amplificarea fragmentelor de ADN prin reacţia în lanţ a polimerazei (PCR)..................... 10 6.3.4. Analiza cu markerii RAPD................................................................................................... 10 6.3.5. Clonarea şi secvenţierea benzilor polimorfe RAPD............................................................. 10 6.3.6. Analiza cu markerii microsateliţi (SSR)............................................................................... 10
CAPITOLUL VII REZULTATE OBŢINUTE............................................................................................................... 10 7.1. ANALIZA GENETICĂ A REZISTENŢEI GRÂULUI LA MĂLURA COMUNĂ ÎN POPULAŢIILE SEGREGANTE F2..................................................................................................... 10
7.1.2. Transmiterea rezistenţei la mălura comună a grâului la descendenţii segreganţi din populaţia F2........................................................................................................................................... 13

1
7.2. REZULTATE PRIVIND CANTITATEA ŞI PURITATEA ADN-ULUI IZOLAT..................... 14 7.3. REZULTATELE ANALIZEI CU MARKERII SPECIFICI GENEI BT10 (FSD şi RSA).......... 14 7.4. REZULTATELE ANALIZEI CU MARKERII RAPD................................................................ 15 7.5. REZULTATE PRIVIND TRANSFORMAREA MARKERILOR RAPD ÎN MARKERI SPECIFICI DE TIP SCAR .................................................................................................................. 17
7.5.1. Izolarea benzilor polimorfe RAPD din gelul de migrare................................................... 17 7.5.2. Clonarea şi secvenţierea benzilor RAPD polimorfe.......................................................... 17
7.6. REZULTATELE ANALIZEI CU MARKERII SSR ................................................................... 18 7.6.1. Atribuirea markerilor moleculari SSR identificaţi genelor de rezistenţă la mălura
comună................................................................................................................................................. 19 CONCLUZII ŞI RECOMANDĂRI.................................................................................................. 20 BIBLIOGRAFIE SELECTIVĂ............................................................................................... 22

Rezumatul tezei de doctorat _____________________________________________________________________________________________
3 3
INTRODUCERE
Grâul (Triticum aestivum L.) este planta din grupul cerealelor care este cultivată pe cele mai mari suprafeţe. Datorită importanţei sale deosebite în alimentaţia oamenilor, cultura grâului este extinsă pe toate continentele. Dintre agenţii patogeni care cauzează boli la grâu, speciile de fungi care atacă spicul de grâu reprezintă agenţi patogeni de importanţă majoră. Micozele grâului afectează atât producţia de grâu dar şi calitatea seminţelor recoltate. Mălura comună a grâului este o boală a grâului cauzată de două specii de Tilletia (Tilletia foetida şi Tilletia tritici). Această boală se transmite prin seminţele infectate sau prin solul contaminat cu spori. Mălura comună este combătută în prezent prin tratarea seminţelor cu fungicide. Rezistenţa la mălura comună este bazată pe gene majore de rezistenţă care au fost identificate, în prezent fiind cunoscute un număr de 15 gene de rezistenţă (Hoffmann şi Metzger, 1976). În studiile anterioare, încercările de a găsi markeri genetici linkaţi cu genele de rezistenţă la speciile de Tilletia au dus la identificarea câtorva markeri specifici linkaţi cu genele de rezistenţă dintre care unii sunt utilizaţi în selecţie (Laroche şi colab., 2000).
Obiectivul propus în cadrul lucrării de doctorat a fost identificarea de markeri genetici pentru rezistenţa grâului comun (Triticum aestivum L.) la speciile de Tilletia care cauzează mălura comună a grâului şi care să poată fi utilizaţi în selecţie. În cadrul acestui demers principalul obiectiv de cercetare urmărit a fost testarea unor markeri existenţi în literatură (markeri RAPD şi SSR) precum şi identificarea de noi markeri de rezistenţă pentru a evidenţia polimorfismul molecular la soiurile şi liniile utilizate în această lucrare.
CAPITOLUL I
CONSIDERAŢII GENERALE PRIVIND IMPORTANŢA ŞI CULTIVAREA GRÂULUI
Grâul comun (Triticum aestivum L.) aparţine ordinului Graminalis, familia Gramineae (Poaceae), genul Triticum L. care cuprinde un număr mare de specii, subspecii, varietăţi şi soiuri. Genul Triticum formează o serie poliploidă care cuprinde specii diploide, tetraploide şi hexaploide toate având numărul de bază de cromozomi de 2n=7. (Feldman, 2001). Grâul comun are o importanţă economică deosebită, fiind cultivat pe suprafeţe foarte mari la nivel mondial, şi este utilizat atât în hrana oamenilor, cât şi pentru furajarea animalelor. 1.1. GENOMUL GRÂULUI
Grâul comun (Triticum aestivum L em. Thell) este un alohexaploid (2n=6x=42) compus din trei genoame distincte dar strâns înrudite (A, B, D) care provin de la trei specii genitori fiecare având numărul de bază de cromozomi (n=7). Cele trei genoame provin de la Triticum urartu (Genomul A), o specie necunoscută de Sitopsis (genomul B) iar de la Triticum tauschi provine genomul D (Ganal şi Röder, 2007). Genomul grâului este de 16x109 perechi de baze pe cromozomi, de aproape 100 de ori mai mare decât genomul la Arabidopsis, de 40 de ori mai mare decât genomul la orez şi de aproximativ 6 ori mai mare decât genomul porumbului (Amuruganathan şi Earle, 1991). 1.2. ASPECTE GENERALE PRIVIND AGENŢII PATOGENI CARE CAUZEAZĂ PRINCIPALELE BOLI LA GRÂU
Cultura grâului este foarte sensibilă la atacul unor boli infecţioase cauzate de un număr foarte mare de agenţi patogeni: virusuri, bacterii, ciuperci, precum şi la insecte vătămătoare cum sunt musca de Hessa şi afidele ruseşti care pot transmite viroze la grâu. Au fost identificate peste 40 de cipuerci, 32 de virusuri, 81 de boli bacteriene care atacă plantele de grâu în diferite stadii de dezvoltare (Bobeş şi colab., 1977).
Dintre bolile spicului de grâu, la ora actuală cele mai răspândite sunt mălura pitică (Tilletia controversa) şi mălura comună (Tilletia spp.), alături de fuzarioză (Fusarium head blight) care produc pierderi considerabile de producţie. Alături de acestea sunt semnalate frecvent ciupercile parazite: rugina

Rezumatul tezei de doctorat _____________________________________________________________________________________________
4 4
galbenă, rugina brună, rugina neagră, tăciunele zburător, înnegrirea cerealelor, făinarea grâului, septorioza, alternarioza (McIntosh, 1998).
CAPITOLUL II
STUDII CU PRIVIRE LA SPECIILE DE TILLETIA CARE PROVOACĂ MĂLURA COMUNĂ A GRÂULUI
2.1. SPECIILE GENULUI TILLETIA. TAXONOMIE. MORFOLOGIE
Genul Tilettia face parte din familia Tiletiaceae, ordinul Ustilaginales, clasa Baziodiomycetes. Speciile de Tilletia care cauzează mălura comună sunt T. laevis Kühn (sinonim Tilletia foetida (Wall.) Liro) şi Tilletia carries (DC.) Tul. (sinonim Tilletia tritici (Bjerk.) Wint (Dumitraş şi Bontea, 1991). Mălura comună se mai numeşte tăciunele mirositor sau tăciunele îmbrăcat şi este una dintre cele mai răspândite şi mai păgubitoare boli ale grâului. Speciile de Tilletia (T. foetida şi T. tritici) sunt foarte asemănătoare între ele deosebindu-se doar din punct de vedere morfologic, prin forma teliosporilor: sporii speciei T. tritici au pereţi reticulaţi iar sporii de T. foetida au pereţii netezi (Saari şi colab., 1996). Agentul patogen al mălurii comune are ca gazdă grâul dar este prezent şi pe alte specii de Triticum, se întâlneşte la speciile care sunt sensibile la una sau mai multe rase de mălură comună precum şi la majoritatea speciilor sensibile la mălura pitică. 2.2. BIOLOGIA MĂLURII COMUNE
Mălura comună este răspândită prin seminţele infectate cu spori. În timpul lucrărilor de recoltare sporii sunt răspândiţi pe suprafaţa seminţelor şi în sol unde supravieţuiesc de la un sezon la altul. În timpul treieratului boabele mălurate se sparg iar sporii sunt răspândiţi în atmosferă şi la suprafaţa solului dar şi pe seminţe. Infecţia este iniţiată atunci când teliosporii de pe suprafaţa seminţelor sau din sol germinează în acelasi timp cu boabele. Prin germinarea teliosporilor se produce (cariogamia acestora) diviziunea reducţională formată din una sau două diviziuni meiotice, chiar înainte de germinaţie. Prin germinare se formează un promiceliu neseptat, hialin, multinucleat pe care se formeaza terminal un numar nedefinit de basidiospori (sporidii primare). Din sporidiile primare sau din conidii, prin copulare se dezvoltă filamente dicariotice de infecţie care pătrund prin axa hipocotilă, succesiv prin toate tesuturile plantei şi formează clamidospori toxici. Clamidosporii sunt de culoare neagră, sferici şi înlocuiesc conţinutul boabelor în spic.
2.3. RASE FIZIOLOGICE DE MĂLURĂ
Au fost ientificate rase patogene atât pentru mălura comună, cât şi pentru mălura pitică. În funcţie de reacţia la mălură a anumitor cultivare numite cultivare diferenţiatoare şi de relaţia genă-pentru-genă care are loc între fungi şi planta gazdă au fost identificate şi definite un număr de 30 de rase pentru T. tritici, 16 rase la T. laevis, şi 17 rase pentru T.controversa (Goates, 1996). În plus faţă de rasele enumerate, au fost identificate numeroase alte patotipuri (Hoffmann, 1982).
În România, primele cercetări cu privire la specializarea fiziologică a speciilor de mălură comună şi mălură pitică au fost efectuate de Săvulescu în 1957 şi Sandu-Ville (1934) care au evidenţiat 20 de rase pentru Tilletia laevis, 9 rase pentru Tilletia triticoides (Gassner) Săvulescu, 8 rase pentru Tilletia tritici şi 7 rase pentru Tilletia controversa. Răspândirea raselor de mălură depinde de soiurile de grâu cultivate, iar menţinerea în cultură a soiurilor cu rezistenţă verticală duce la apariţia de rase noi, mai virulente deoarece rasele de mălură au posibilitatea de a realiza nelimitat combinaţii noi ale genelor de virulenţă cu mult mai agresive (Kendrick şi Holton, 1961).
Prezentarea virulenţei raselor de mălură faţă de fiecare din genele de rezistenţă cunoscute, arată că cele mai eficace gene de rezistenţă la mălură sunt: genele Bt8, Bt11 şi Bt12 faţă de care nici o rasă de mălură comună nu a manifestat virulenţă şi genele Bt13, Bt14 şi Bt15 faţă de care manifestă virulenţă

Rezumatul tezei de doctorat _____________________________________________________________________________________________
5 5
Fig.1. Simptome ale infecţiei cu mălura comună pe spic – dreapta (Sursa: Matanguihan şi colab., 2011). Cariopse (boabe) de grâu infectate cu spori de mălura comună – stânga (Original)
doar una respectiv două rase de mălură. Se remarcă de asemenea cea mai mare virulenţă a rasei de Tilettia tritici (T-30), manifestată faţă de şapte dintre genele de rezistenţă.
2.4. SIMPTOMELE PRODUSE LA PLANTELE INFECTATE
Simptomele sunt manifestate deplin şi sunt vizibile la maturitatea plantelor prin transformarea întregului conţinut al boabelor de grâu într-o masă de spori negri (figura 1). Infecţia plantelor de grâu cu sporii de Tilletia este influenţată de o serie de factori dintre care reacţia şi tipul solului, cantitatea de teliospori, temperatura, lumina, umiditatea sunt cei mai importanţi (Dumitraş şi Bontea, 1991; Ghibu, 2002). 2.5. STRATEGII DE COMBATERE A MĂLURII COMUNE Combaterea chimică De-a lungul timpului mălura a fost controlată prin tratarea intensivă a materialului de însămânţare cu fungicide iar prin această metodă pierderile au fost mult diminuate (Nagy şi Moldovan, 2007). Dacă aceastea sunt aplicate regulat pierderile sunt eliminate în totalitate deoarece sunt eficace atât asupra sporilor de pe seminţe cât si asupra sporilor din sol (Dumitraş şi Bontea, 1991). Utilizarea soiurilor rezistente. Cutivarea soiurilor rezistente la mălură poate fi cea mai bună metodă de combatere a bolii, când sursele de rezistenţă sunt disponibile. Deoarece în ameliorarea clasică nu a existat interes pentru această boală, multe dintre soiurile actuale de grâu de toamnă, cultivate în Europa au fost identificate ca fiind foarte susceptibile la mălură (Tilletia tritici şi Tilletia laevis). Dar au fost identificate şi genotipuri cu rezistenţă la unele soiuri (Dumalasová şi Bartoš, 2006a; Wächter şi colab., 2007). În urma testărilor efectuate, pe plan european sunt menţionate soiurile de origine cehă (cultivarul Bill, cu un grad de infecţie de sub 15% la infecţia artificială), (Dumalasová şi Bartoš, 2007), soiurile europene Tjelvar şi Stava (Bt8, Bt9, Bt10) cu un nivel foarte scăzut de infecţie de 0,6 % şi respectiv 0,7 % (Dumalasová şi Bartoš, 2006b). Testările efectuate au evidenţiat şi alte genotipuri rezistente, de exemplu linia de grâu 0028162-II cu o medie a nivelului de atac de 0.1 % (Huber şi Buerstmayr, 2006). În România au fost identificate mai multe linii de grâu rezistente la mălură a căror sursă de rezistenţă provenite de la Triticum monococum şi de la Triticale (Oncică şi Săulescu, 2008). De asemenea unele soiuri au manifestat rezistenţă crescută în testările efectuate: Flamura 85, Arieşan, Apulum, Transilvania, Voyage şi Estica (Ghibu, 2002).

Rezumatul Tezei de Doctorat
6 6
CAPITOLUL III
ASPECTE GENERALE PRIVIND REZISTENŢA GRÂULUI LA MĂLURA COMUNĂ
3.1. GENETICA REZISTENŢEI PLANTELOR LA BOLI 3.1.1. Tipuri de rezistenţă
În funcţie de răspunsul plantei gazdă la atacul patogenului rezistenţa a fost împărţită în două grupe: rezistenţa specifică şi rezistenţa nespecifică. Rezistenţa specifică se exprimă în stadii timpurii de dezvoltare a plantelor (stadiul de plantulă) şi se caracterizează prin specificitatea rasă – cultivar Rezistenţa specifică este în general monogenică şi dominantă şi este rapid învinsă de patogeni, deci este puţin durabilă. Rezistenţa nespecifică se caracterizează prin sensibilitatea plantelor în stadiul de plantulă dar rezistenţa se manifestă în stadiul de plantă adultă atunci când plantele sunt inoculate cu un amestec de rase ale patogenului. Aceste soiuri conservă un bun nivel de rezistenţă, durabilă. Rezistenţa nespecifică a fost detectată la soiurile de grâu ce nu posedă nici o genă majoră sau au gene majore care au devenit ineficace. 3.2. CONTROLUL GENETIC AL REZISTENŢEI GRÂULUI LA MĂLURA COMUNĂ
Rezistenţa la mălura comună este controlată de gene majore de rezistenţă dar multe dintre ele au fost învinse de virulenţa raselor patogenului (Hoffman, 1982). Au fost identificate un număr de 15 gene majore de rezistenţă (notate de la Bt1 la Bt15) care asigură o rezistenţă specifică (Hoffman şi Metzger, 1976; Goates, 1996). Acestea se găsesc singure sau în combinaţii cu alte gene în anumite soiuri rezistente sau tolerante la mălură aparţinând speciilor de Triticum. Gena Bt8 este responsabilă de rezistenţa celor mai multe soiuri dar unele soiuri au gena Bt12 în combinaţie cu alte gene. O altă gena de rezistenţă (Bt-Z), originară dintr-o translocaţie de la Agropyron intermedium, a fost folosită pentru combaterea mălurii comune în Uniunea Sovietică (Mozgovoi şi colab., 1987).
Din punct de vedere al tipului de rezistenţă, rezistenţa grâului la Tilletia spp este o rezistenţă specifică, controlată monogenic, bazată pe relaţia genă-pentru-genă între genele de rezistenţă ale grâului şi genele de avirulenţă ale patogenului (Hoffman, 1982). Cu toate că genele de rezistenţă sunt cunoscute de mult timp, o rezistenţă durabilă se poate obţine numai prin introgresia mai multor gene de rezistenţă într-un soi (Hoffman şi Metzger, 1976).
Rezistenţa la mălură comună şi pitică este controlată de aceleaşi gene de rezistenţă (Hoffmann şi Metzger, 1976). Un cultivar rezistent la o rasă de mălură comună va fi de asemenea rezistent la o rasă de mălură pitică care are aceleaşi sau mai puţine gene de virulenţă.
3.3. SURSE DE REZISTENŢĂ A GRÂULUI LA SPECIILE DE TILLETIA Deşi tratarea seminţelor cu fungicide este o metodă foarte eficace de combatere totuşi sunt necesare cultivarale de grâu rezistente la mălură mai ales pentru combaterea bolii în agricultura organică, pentru producerea unor culturi ecologice şi ca o alternativă mai puţin costisitoare la tratarea cu fungicide (Saari şi colab., 1996). Metoda de bază în lupta cu mălurile grâului rămâne crearea de soiuri rezistente. Rezistenţa la mălura comună a fost identificată la câteva specii de Triticum şi Aegilops şi la alte specii înrudite: Triticum, Aegilops şi Agropyron, grâul emmer, Triticale, grâul spelta, grâul durum (Rubiales şi colab., 2001, Mamluk,1998). Surse de rezistenţă la malură au fost identificate si la Hordeum chilense o specie sălbatică de diploidă de orz din Chile si Argentina.
În prezent, centrele de origine a rezistentei grâului la mălura comună au fost investigate şi s-au identificat surse de rezistenţă a grâului la mălură în Serbia, Iran şi Muntenegru la unele soiuri sălbatice (Bonmann, 2006).

Rezumatul Tezei de Doctorat
7 7
CAPITOLUL IV
MARKERII MOLECULARI CA NOI METODE UTILIZATE ÎN AMELIORAREA PENTRU REZISTENŢA GRÂULUI LA BOLI
Un marker molecular este reprezentat de diferenţele genetice dintre organismele individuale sau
specii (Collard şi colab., 2005). În comparaţie cu markerii morfologici sau biochimici, markerii moleculari corespund polimorfismelor descoperite la nivel de ADN. Analiza acestor polimorfisme prin tehnicile de biologie moleculară se adresează întregului genom, regiunilor care sunt traduse sau nu în proteine şi este independentă faţă de condiţiile de mediu.
4.1. TIPURI DE MARKERI MOLECULARI Clasele de markeri genetici utilizate în genetica şi ameliorarea plantelor pot fi grupate astfel:
markerii fenotipici markerii izoenzimatici proteine markeri markerii RFLP Markerii bazaţi pe PCR (RAPD, AFLP, SSR, ISSR)
Alte tipuri de markeri moleculari (STS, SCAR, CAPS, ESTs, DArT) care există şi sunt utilizaţi. În ultimul timp, o clasă de markeri moleculari utilizată este reprezentată de polimorfismul unei singure nucleotide (Single nuleotide Polymorphism - SNPs).
4.2. AMELIORAREA PLANTELOR PENTRU REZISTENŢA LA BOLI CU AJUTORUL MARKERILOR MOLECULARI
Ameliorarea rezistenţei la boli a început prin încorporarea de gene majore pentru rezistenţă în noile soiuri şi prin utilizarea rezistenţei specifice (oligogenică), a rezistenţei verticale. Practica a demonstrat că aceste cultivare la care s-a încorporat o genă majoră conferă o rezistenţă instabilă care este învinsă de apariţia de noi rase fiziologice a parazitului. Pentru rezistenţă grâului la mălura comună introducerea de gene de rezistenţă Bt la rase specifice la soiurile cultivate a fost cea mai utilizată metodă de ameliorare pentru controlul bolii.
4.2.1. SELECȚIA ASISTATĂ DE MARKERI MOLECULARI
Pe lângă interesul lor fundamental markerii moleculari prezintă importanţă deosebită în domeniul selecţiei.
În ameliorarea convenţională, se utilizează selecţia bazată pe fenotip sau pe markerii morfologici. O astfel de selecţie este de multe ori mai puţin eficientă datorită numărului de gene care controlează un anumit caracter, a gradului în care mediul afectează expresia acestor gene.
Selecţia asistată de markeri moleculari (MAS) este o metodă de selecţie indirectă care presupune selecţia plantelor purtătoare a regiunilor care sunt implicate în expresia caracterelor de interes (plante rezistente) prin markerii moleculari (Landjeva şi colab., 2007). În prezent selecţia asistată de markeri moleculari este axată pe: (1) selecţia caracterelor cu importanţă economică, pentru care selecţia bazată numai pe fenotip este dificilă sau necesită costuri ridicate (2) selecţia caraterelor cantitative, pentru care selecţia bazată pe fenotip este dificilă (3) piramidarea genelor de rezistenţă. Selecţia asistată de markeri moleculari este considerată o metodă foarte utilă în ameliorarea pentru rezistenţa la boli (Langdridge şi colab., 2001). a speciilor cultivate deoarece:
Identificarea genotipului de rezistenţă prin testarea în câmp este costisitoare, necesită timp îndelungat şi este foarte mult influenţată de condiţiile de mediu. Cu ajutorul markerilor se elimină influenţa mediului în selecţie.

Rezumatul Tezei de Doctorat
8 8
Selecţia se poate realiza fără a recurge la teste de inoculare, permiţând astfel evitarea erorilor asociate cu utilizarea acestor proceduri, fiind posibilă ameliorarea rezistenţei în arealuri unde patogenul nu există.
Schema de selecţie este accelerată întrucât selecţionerii pot conchide prezenţa genei prin prezenţa markerului înaintea expresiei fenotipice.
Markeri linkaţi cu diferite gene de rezistenţă oferă posibilitatea combinării mai multor gene (piramida genelor) atunci când selecţia anumitor gene în prezenţa altor gene este dificilă.
CAPITOLUL V
SCOPUL SI OBIECTIVELE CERCETĂRILOR
În studiile anterioare, încercările de a găsi markeri genetici linkaţi cu rezistenţa la Tilletia spp. au dus la identificarea câtorva markeri RAPD linkaţi cu genele de rezistenţă (Laroche şi colab., 2000). Utilitatea lor a fost confirmată pentru identificarea soiurilor care poartă acestă genă iar Importanţa lor derivă aşadar din faptul că pot fi utilizaţi în selecţie pentru a identifica soiurile şi liniile de grâu rezistente ca alternativă la tratarea seminţelor cu fungicide.
În consecinţă şi scopul cercetarii de faţă a fost identificarea de markeri moleculari pentru unele gene de rezistenţă a grâului la mălura comună. Cercetările s-au bazat pe utilizarea markerilor deja existenţi, testarea acestora la soiurile şi liniile cultivate pe plan local. Apoi, cercetările au urmărit identificarea de noi markeri care să poată fi utilizaţi în selecţia pentru rezistenţă la mălura comună.
Ca urmare, cercetarea de faţă a urmărit atingerea următoarelor obiective: (1) identificarea de markeri moleculari nespecifici prin utilizarea tehnici RAPD (Random Amplified Polymorphic DNA - polimorfisme de ADN amplificate aleator) pentru genele de rezistenţă la mălura comună a grâului; (2) transformarea acestor markeri dominanţi în markeri codominanţi specifici de tip SCAR (Sequence Charcterized Amplified Regions - secvenţe caracteristice regiunilor amplificate); (3) obţinerea generaţiilor segregante F2 prin autopolenizarea plantelor F1 rezultate din încrucişarea dintre liniile (soiurile) parentale rezistente cu cele sensibile; (4) caracterizarea fenotipică a plantelor din generaţia F2 prin teste de infecţie artificială; (5) testarea unor markeri specifici de tip SSR în vederea evidenţierii polimorfismului molecular; (6) atribuirea markerilor identificaţi genelor de rezistenţă la mălura comună prin metoda segregării în amestec; (7) stabilirea relaţiei de înlănţuire dintre markeri identificaţi şi genele de rezistenţă la mălura comună.
CAPITOLUL VI
MATERIAL ŞI METODE
6.1. MATERIAL BIOLOGIC UTILIZAT PENTRU IDENTIFICAREA DE MARKERI DE REZISTENŢĂ A GRÂULUI LA MĂLURA COMUNĂ
1. Germoplasmă analizată reprezentată de linii de grâu - forme parentale este reprezentată de 13 linii (soiuri) de grâu de toamnă, liniile aparţinând unor generaţii segregante, dintre care 5 cu rezistenţă de câmp la mălura comună şi 8 linii (soiuri) sensibile la aceasta boală. Acest material biologic provine de la INCDA Fundulea, Bucureşti unde aceste linii şi soiuri au fost onţinute. Următoarele cinci linii de grâu de toamnă sunt cunoscute ca find foarte rezistente la mălura comună, manifestându-şi rezistenţa în testări repetate: 99419G4-1A/1-1, 00274G2-31, 00281G2-11, 00399G2-11, 01450G1-1. Soiurile de grâu de toamnă sensibile la mălura comună: Farmec, Delabrad, Glosa, Boema, Jiana, Crina, Dropia şi linia F96869G1-108 au manifestat de asemenea sensibilitatea la mălura comună în testări repetate efectuate la INCDA Fundulea (Săulescu N., comunicare personală).
2. De asemenea au fost analizate două seturi de linii dihaploide: - 28 linii rezultate din combinaţia GP 369 (P99419G4-1AI1-1/00356G8-1), notate de la 9.1 la 9.28;

Rezumatul Tezei de Doctorat
9 9
- 41 de linii din combinaţia GP 384 (P99419G4-1AI1-1/98047G14-2INC1), notate de la 4.1 la 4.41.
3. Ca material biologic s-au mai utilizat populaţii segregante F2 obţinute prin încrucişarea formelor parentale rezistente şi sensibile la mălură.
6.2. METODE UTILIZATE Pentru a fi utilizaţi în selecţie mai întâi trebuie ca markerii moleculari identificaţi să fie atribuiţi genelor de rezistenţă. În acest scop se analizează diferite populaţii segregante pentru a evidenţia cosegregarea markerilor cu gena de rezistenţă (sau cu caracterul dorit controlat de genă) (Landjeva şi colab., 2007): Analizele au la bază fie cosegregarea markerilor cu gena de rezistenţă, fie alte metode de analiză:
utilizarea liniilor obţinute prin retroîncrucişare (linii RIL) utilizarea liniilor aproape izogenice (NIL) utilizarea metodei segregării în amestec (Bulked Segregant Analysis) utilizarea liniilor dihaploide
În lucrarea de faţă pentru identificarea de markeri moleculari s-au abordat două metode: Metoda cosegregării markerului cu gena de rezistenţă Metoda Segregării în amestec (Bulked Segregant Analysis - BSA)
6.2.1. Metoda cosegregării
Metoda cosegregării are la bază ideea că markerul şi gena de rezistenţă segregă împreună iar prezenţa genei este evidenţiată de prezenţa markerului.
6.2.2. Metoda segregării în amestec (Bulked Segregant Analysis - BSA)
Metoda presupune efectuarea a două amestecuri de ADN provenite de la două fenotipuri extreme (rezistent şi sensibil) provenite de la indivizii unei populaţii segregante (F2 sau dublii haploizi). Amestecurile de ADN (bulk-uri) sunt analizate cu markerii moleculari pentru a testa polimorfismul dintre ele (Michelmore şi colab., 1991). Markerii polimorfi obţinuţi sunt utilizaţi apoi pentru a stabili distanţa de înlănţuire cu gena de interes în populaţiile segregante (Landjeva şi colab., 2007).
6.2.3. Izolarea ADN-ului genomal
Izolarea ADN a fost realizată utilizând protocolul descris de către Lodhi şi colab. (1994) modificat de către Pop şi colab. (2003) protocol bazat pe utilizarea CTAB (cetil trimetilammonium bromide - bromură de cetiltrimetilamonium) cu adaos de acid citric, PVP (policlorura de vinil) şi Dieca, utilizând frunze proaspete.
6.2.4. Izolarea ADN-ului plasmidial
Izolarea ADN-ului plasmidial a fost realizată prin metoda lizei alcaline. Prin această metodă se obţine ADN plasmidial de foarte bună calitate şi în cantitate suficient de mare pentru a putea fi utilizat în analizele ulterioare.
6.2.5. Determinarea cantităţii şi a calităţii acizilor nucleici izolaţi (ADN)
ADN-ul poate fi cuantificat în general prin următoarele metode: electroforeza în gel şi cunatificarea spectrofotometrică a ADN-ului. Pentru a fi utilizat în analizele ulterioare, calitatea ADN-ului extras trebuie să fie corespunzătoare. Calitatea ADN-ului izolat este foarte importantă pentru diferitele analize cum ar fi digestia cu enzime de restricţie sau amplificarea PCR a ADN-ului matriţă.
Verificarea cantităţilor şi a calităţii soluţiei de ADN a fost realizată prin metoda spectofotometrică. Uneori pentru a evalua calitatea ADN a fost migrat şi în gel de agaroză (0,8 %).
6.3. METODE DE ANALIZĂ MOLECULARĂ 6.3.1. Analiza cu primeri specifici de rezistenţă pentru gena Bt10
Perechea de primeri FSA şi RSA sunt specifici pentru gena de rezistenţă Bt10 şi markează un fragment polimorf de 275 pb (Laroche şi colab., 2000). Aceşti primeri au fost testati atât la liniile parentale cât şi la liniile dihaploide de grâu.

Rezumatul Tezei de Doctorat
10 10
6.3.2. Amplificarea fragmentelor de ADN prin reacţia în lanţ a polimerazei (PCR) Amplificarea fragmentelor de ADN prin reacţia în lanţ a polimerazei se bazează pe capacitatea enzimei ADN polimeraza de a sintetiza fragmente de ADN în prezenţa unor primeri cu secvenţa nucleotidică şi a celor patru nucleotide sintetice (dNTP). 6.3.3. Electroforeza acizilor nucleici (ADN-ului)
Fragmentele de ADN amplificate prin PCR au fost separate prin electroforeză în gel de agaroză sau poliacrilamidă. Vizualizarea şi examinarea lor s-a realizat printr-o sursă de lumină UV, iar imaginile au fost captate şi stocate cu aparatul Alpha Innotech Imager.
6.3.4. Analiza cu markerii RAPD
Pentru a testa polimorfismul pentru rezistenţă dintre liniile şi soiurile parentale rezistente şi cele sensibile şi pentru a descoperi markerii polimorfi RAPD au fost utilizaţi un număr de 32 primeri aleşi la întâmplare (primeri Operon).
6.3.5. Clonarea şi secvenţierea benzilor polimorfe RAPD Pentru a putea fi utilizaţi mai departe produşii de amplificare polimorfi RAPD sunt izolaţi din gelul de migrare în vederea purificării şi a obţinerii secvenţelor. După migrarea şi analiza gelului, opt benzi reprezentative au fost izolate şi reamplificate cu ajutorul primerilor universali, după cum urmează:
- 3 benzi obţinute din amplificarea cu primerul OPC 04 (550 pb) - 5 benzi obţinute din amplificarea cu primerul OPA 17 (850 pb)
Clonarea benzilor separate şi izolate din gelul de agaroză a avut ca scop secvenţierea lor şi pe baza secvenţei nucleotidice construirea de primeri specifici care să amplifice această bandă. Fragmentele de ADN recuperate au fost ligate în vectorul pJET1.2/blunt şi au fost transformate în bacteria Eshericha coli, tulpina GM2163 utilizând protocolul indicat de producător. Pentru clonarea benzilor s-a utilizat kitul de clonare CloneJetTM PCR Cloning Kit (Fermentas). Acesta conţine vectorul de clonare pJET1.2/blunt care conţine o genă letală. În momentul inserării fragmentului dorit în situsul de clonare această genă este inactivată. După identificarea clonelor transformate, acestea au fost cultivate peste noapte pe mediu LB cu ampicilină din care au fost utilizat pentru izolarea ADN-ului plasmidial. Din coloniile care au fost pozitive, extracţia ADN-ul plasmidic a fost realizat pe baza protocolului lizei alcaline. ADN-ul plasmidic extras a fost secvenţiat în laboratoarele Macrogen, Olanda. 6.3.6. Analiza cu markerii microsateliţi (SSR)
În lucrarea de faţă au fost testaţi pentru polimorfism un set de markeri microsateliţi localizaţi pe diferiţi cromozomi ai grâului în regiunile în care sunt localizate şi unele gene de rezistenţă (Bt) prezente în germoplasma analizată. Markerii au fost selectaţi pe baza informaţiilor existente în literaturară cu privire la ”linkage-ul” lor cu unele gene de rezistenţă a grâului la mălura comună. Setul de primeri pentru markeri au fost sintetizaţi la firma Microsynth, Elveţia şi cuprinde 11 perechi de primeri.
CAPITOLUL VII REZULTATE OBŢINUTE
7.1. ANALIZA GENETICĂ A REZISTENŢEI GRÂULUI LA MĂLURA COMUNĂ ÎN POPULAŢIILE SEGREGANTE F2
Pentru obţinerea generaţiei F2 în anul 2007 s-au efectuat hibridările între formele parentale rezistente şi cele sensibile (rezistent x sensibil). Liniile de grâu de toamnă: 99419G4-1A/1-1, 00274 G2-31, 00281G2-11, 00399G2-11, 01450G1-1 au fost utilizate ca forme tată în încrucişări. Soiurile şi liniile de grâu de toamnă, sensibile la Tilletia spp. Farmec, Delabrad, F96869G1-108, Glosa, Boema, Jiana, Crina, Dropia au fost utilizate ca forme mamă în încrucişări. Hibrizii F1 obţinuţi au fost semănaţi în câmp în toamna anului 2008 în vederea obţinerii populaţiei segregante F2.
Înainte de semănatul seminţelor acestea au fost supuse infecţiei artificiale cu spori de Tilletia carries şi Tilletia foetida. Rezultatele infecţiei artificiale (fig. 2) au fost evaluate la soiurile şi liniile

Rezumatul Tezei de Doctorat
11 11
Fig. 2. Simptome ale infecţiei cu mălură pe spice, spic infectat cu mălură (stânga), spic sănătos (dreapta)
parentale şi la următoarele 10 descendenţe din populaţiile F2: 99419G4-1A/1-1 x Glosa, 99419G4-1A/1-1 x Boema, 99419G4-1A/1-1 x Dropia, 00274G2-31 x Glosa, 00274G2-31 x Dropia, 00281G2-11 x Boema, 00399G2-11x Boema, 01450G1-1 x Glosa, 01450G1-1 x Boema, 00450G1-1 x Dropia.
Experimentele de inoculare artificială cu spori de (Tilletia caries şi Tilletia foetida) au fost organizate în câmp, în spaţiul agricol al grădinii botanice USAMV Cluj-Napoca, în perioada 2008-2009. Experienţele au fost realizate în blocuri complet randomizate cu trei repetiţii. Au fost semănate în câmp un număr de aproximativ 250 seminţe din fiecare descendenţă F2 iar de la formele parentale (un numar de 20 de seminţe). Sporii au fost obţinuti prin recoltarea spicelor infecate infectate natural în câmp de la Staţiunea de Cercetare şi dezvoltare Agricolă Simnic. Au fost infectate atât cele opt descendenţe F2 cât şi formele parentale.
Evaluarea rezistenţei s-a făcut prin raportarea numărului de plante atacate la numărul total de plante. Analiza varianţei a fost utilizata pentru interpretarea datelor. Pentru analiza χ2 au fost testate o populaţie formată din 12 descendenţe F2, populaţiile testate în număr de 10 iar două variante reprezintă repetiţiile efectuate. De asemenea s-a efectuat şi o analiză genetică pentru evaluarea modului de transmitere a rezistenţei la mălura comună. Cunoaşterea modului de transmitere a rezistenţei la mălura comună este un obiectiv important pentru identificarea genelor de rezistenta şi a markerilor genetici care sa fie utilizaţi în selecţia asistată de markeri moleculari.
Reacţia soiurilor şi liniilor parentale la infecţia artificială
Tabelul 1
Media procentului de infecţie cu mălură la soiurile şi liniile parentale (%) Rezistente Sensibile
Gena de rezistenţă
Linia Media procentului de infecţie (%)
Soiul (Linia) Media procentului de infecţie (%)
Farmec nedetectat ? 99419G4-1A/1-1 0 Delabrad 50 Bt5 00274 G2-31 0 F96869G1-
108 50
Bt11 00281G2-11 0 Glosa nedetectat Bt8, Bt10 00399G2-11 0 Boema nedetectat Bt11 01450G1-1 0 Jiana 33.3 Crina 33.3 Dropia 100
Rezumatul Tezei de Doctorat
12 12
Liniile de grâu parentale rezistente utilizate ca părinţi în hibridări au manifestat constant rezistenţa la Tilletia spp. fără simptome de infecţie cu mălură, acestea s-au dovedit rezistente şi în alte teste efectuate în anii precedenţi (tabel 1). La soiurile şi liniile sensibile, gradul de infecţie a variat între 33.3-100 %. De exemplu, soiul Dropia este cunoscut ca foarte sensibil la speciile Tilletia (Oncică şi Săulescu, 2007) procentul de infecţie a fost de 100 % la doza de 5 g spori/1000 g seminţe. Reacţia descendenţelor de grâu segregante F2 la infecţia artificială
Analiza varianţei a fost realizată pentru a testa diferenţele în ceea ce priveşte procentul de infecţie dintre descendenţele F2 rezultate de la cele 12 încrucişări (tabelul 2). Datele privind procentul de infecţie utilizate pentru analiza varianţei au fost transformate în unghiuri =arc sin √ %.
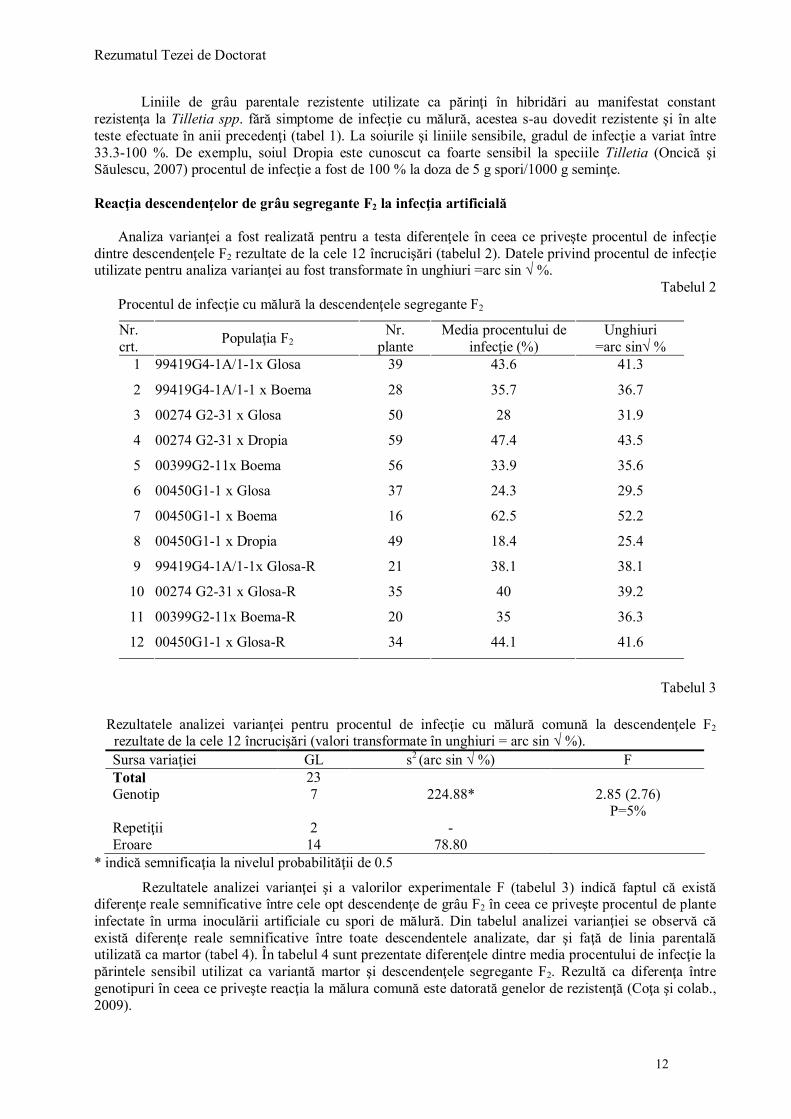
Tabelul 2 Procentul de infecţie cu mălură la descendenţele segregante F2
Nr. crt. Populaţia F2
Nr. plante
Media procentului de infecţie (%)
Unghiuri =arc sin√ %
1 99419G4-1A/1-1x Glosa 39 43.6 41.3
2 99419G4-1A/1-1 x Boema 28 35.7 36.7
3 00274 G2-31 x Glosa 50 28 31.9
4 00274 G2-31 x Dropia 59 47.4 43.5
5 00399G2-11x Boema 56 33.9 35.6
6 00450G1-1 x Glosa 37 24.3 29.5
7 00450G1-1 x Boema 16 62.5 52.2
8 00450G1-1 x Dropia 49 18.4 25.4
9 99419G4-1A/1-1x Glosa-R 21 38.1 38.1
10 00274 G2-31 x Glosa-R 35 40 39.2
11 00399G2-11x Boema-R 20 35 36.3
12 00450G1-1 x Glosa-R 34 44.1 41.6
Tabelul 3
Rezultatele analizei varianţei pentru procentul de infecţie cu mălură comună la descendenţele F2
rezultate de la cele 12 încrucişări (valori transformate în unghiuri = arc sin √ %). Sursa variaţiei GL s2 (arc sin √ %) F Total 23 Genotip 7 224.88* 2.85 (2.76)
P=5% Repetiţii 2 - Eroare 14 78.80
* indică semnificaţia la nivelul probabilităţii de 0.5
Rezultatele analizei varianţei şi a valorilor experimentale F (tabelul 3) indică faptul că există diferenţe reale semnificative între cele opt descendenţe de grâu F2 în ceea ce priveşte procentul de plante infectate în urma inoculării artificiale cu spori de mălură. Din tabelul analizei varianţiei se observă că există diferenţe reale semnificative între toate descendentele analizate, dar şi faţă de linia parentală utilizată ca martor (tabel 4). În tabelul 4 sunt prezentate diferenţele dintre media procentului de infecţie la părintele sensibil utilizat ca variantă martor şi descendenţele segregante F2. Rezultă ca diferenţa între genotipuri în ceea ce priveşte reacţia la mălura comună este datorată genelor de rezistenţă (Coţa şi colab., 2009).
Rezumatul Tezei de Doctorat
13 13
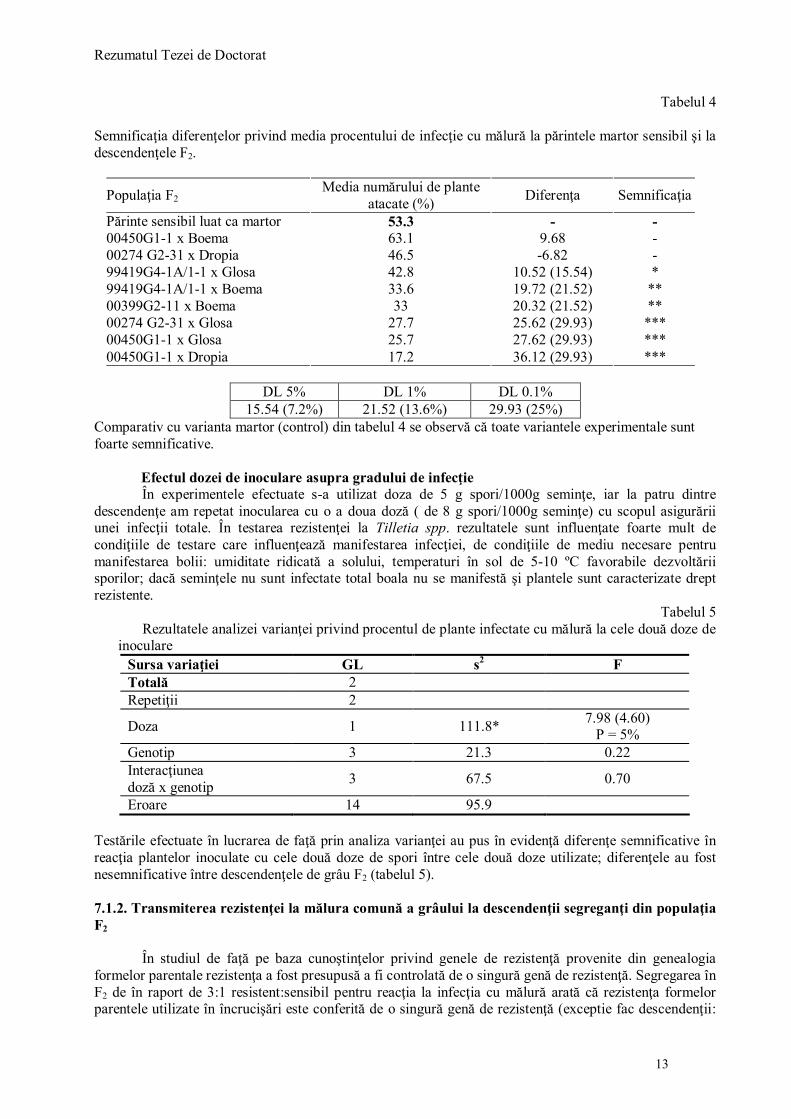
Tabelul 4
Semnificaţia diferenţelor privind media procentului de infecţie cu mălură la părintele martor sensibil şi la descendenţele F2.
Populaţia F2 Media numărului de plante
atacate (%) Diferenţa Semnificaţia
Părinte sensibil luat ca martor 53.3 - - 00450G1-1 x Boema 63.1 9.68 - 00274 G2-31 x Dropia 46.5 -6.82 - 99419G4-1A/1-1 x Glosa 42.8 10.52 (15.54) * 99419G4-1A/1-1 x Boema 33.6 19.72 (21.52) ** 00399G2-11 x Boema 33 20.32 (21.52) ** 00274 G2-31 x Glosa 27.7 25.62 (29.93) *** 00450G1-1 x Glosa 25.7 27.62 (29.93) *** 00450G1-1 x Dropia 17.2 36.12 (29.93) ***
DL 5% DL 1% DL 0.1%
15.54 (7.2%) 21.52 (13.6%) 29.93 (25%) Comparativ cu varianta martor (control) din tabelul 4 se observă că toate variantele experimentale sunt foarte semnificative. Efectul dozei de inoculare asupra gradului de infecţie
În experimentele efectuate s-a utilizat doza de 5 g spori/1000g seminţe, iar la patru dintre descendenţe am repetat inocularea cu o a doua doză ( de 8 g spori/1000g seminţe) cu scopul asigurării unei infecţii totale. În testarea rezistenţei la Tilletia spp. rezultatele sunt influenţate foarte mult de condiţiile de testare care influenţează manifestarea infecţiei, de condiţiile de mediu necesare pentru manifestarea bolii: umiditate ridicată a solului, temperaturi în sol de 5-10 ºC favorabile dezvoltării sporilor; dacă seminţele nu sunt infectate total boala nu se manifestă şi plantele sunt caracterizate drept rezistente.
Tabelul 5 Rezultatele analizei varianţei privind procentul de plante infectate cu mălură la cele două doze de
inoculare Sursa variaţiei GL s2 F Totală 2 Repetiţii 2
Doza 1 111.8* 7.98 (4.60) P = 5%
Genotip 3 21.3 0.22 Interacţiunea doză x genotip 3 67.5 0.70
Eroare 14 95.9 Testările efectuate în lucrarea de faţă prin analiza varianţei au pus în evidenţă diferenţe semnificative în reacţia plantelor inoculate cu cele două doze de spori între cele două doze utilizate; diferenţele au fost nesemnificative între descendenţele de grâu F2 (tabelul 5). 7.1.2. Transmiterea rezistenţei la mălura comună a grâului la descendenţii segreganţi din populaţia F2
În studiul de faţă pe baza cunoştinţelor privind genele de rezistenţă provenite din genealogia formelor parentale rezistenţa a fost presupusă a fi controlată de o singură genă de rezistenţă. Segregarea în F2 de în raport de 3:1 resistent:sensibil pentru reacţia la infecţia cu mălură arată că rezistenţa formelor parentele utilizate în încrucişări este conferită de o singură genă de rezistenţă (exceptie fac descendenţii:
Rezumatul Tezei de Doctorat
14 14
00399G2-11x Boema, 99419G4-1A/1-1 x Boema, 00281G2-11 x Boema, 00274G2-31 x Glosa – R, 01450G1-1 x Glosa - R. La acestea modul de segregare nu a putut fi evaluat probabil datorită numărului mic de plante care a putut fi evaluat; astfel au fost afectate mai mult de eroare decât celelalte (tabelul 6).
Tabelul 6 Analiza segregării fenotipice în generaţia F2. Numărul observat de plante resistente şi sensibile, raportul
de segregare aşteptat pentru o genă de rezistenţă, (χ2).
Nr. plante Raport de asteptat
(rez:sens)
Chi pătrat (χ2)
Valorile χ2 la P=5%
Nr. Descendenţa de grâu F2 testată
Rezist. Sens. 1 00399G2-11x Boema 14 8 3:1 1.5 0.22 2 00274 G2-31 x Glosa 22 17 3:1 7.18 0.007
3 99419G4-1A/1-1x Glosa 18 10 3:1 1.7 0.19
4 99419G4-1A/1-1 x Boema 36 14 3:1 0.24 0.62
5 00274 G2-31 x Dropia 31 28 3:1 15.8 0.00007
6 99419G4-1A/1-1/ Dropia 37 19 3:1 2.37 0.12
7 00281G2-11 x Boema 28 9 3:1 0.0085 0.92
8 00450G1-1 x Glosa 6 10 3:1 12 0.0005
9 99419G4-1A/1-1x Glosa-R 40 9 3:1 1.14 0,28
10 00274 G2-31 x Glosa-R 13 8 3:1 1.92 0.16
11 00399G2-11x Boema-R 21 14 3:1 4.18 0.04
12 00450G1-1 x Glosa-R 13 8 3:1 1.06 0.30 7.2. REZULTATE PRIVIND CANTITATEA ŞI PURITATEA ADN-ULUI IZOLAT
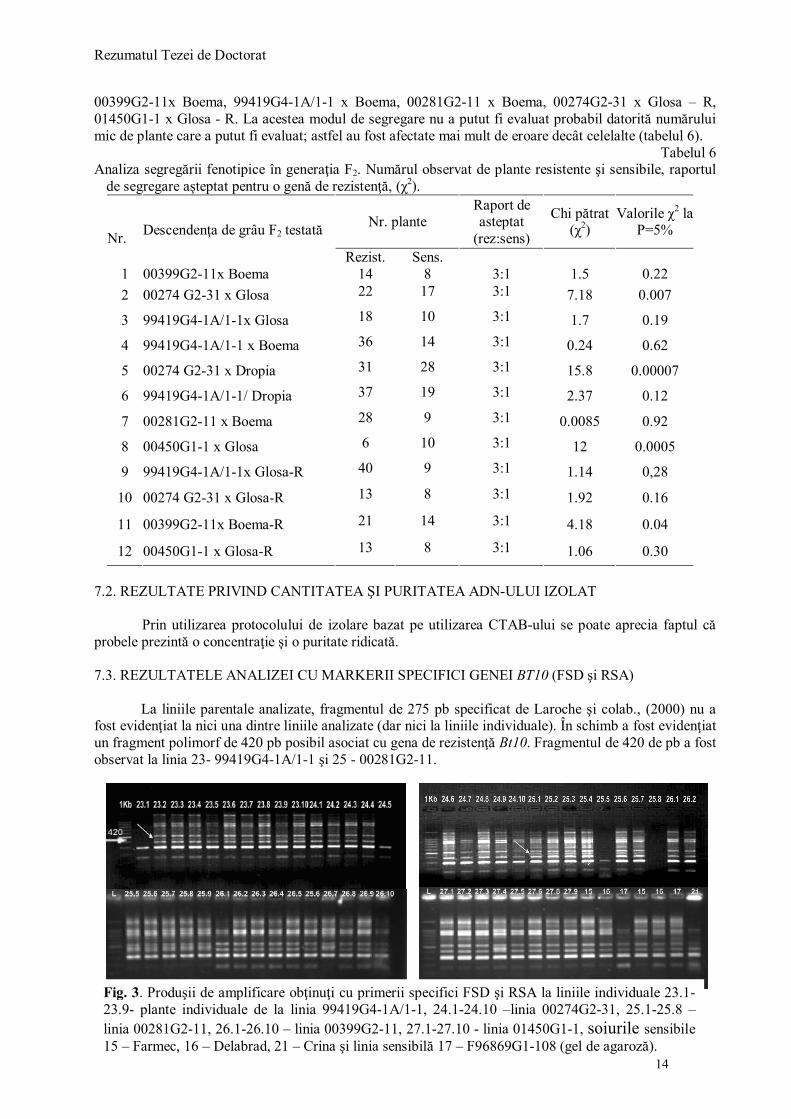
Prin utilizarea protocolului de izolare bazat pe utilizarea CTAB-ului se poate aprecia faptul că probele prezintă o concentraţie şi o puritate ridicată. 7.3. REZULTATELE ANALIZEI CU MARKERII SPECIFICI GENEI BT10 (FSD şi RSA) La liniile parentale analizate, fragmentul de 275 pb specificat de Laroche şi colab., (2000) nu a fost evidenţiat la nici una dintre liniile analizate (dar nici la liniile individuale). În schimb a fost evidenţiat un fragment polimorf de 420 pb posibil asociat cu gena de rezistenţă Bt10. Fragmentul de 420 de pb a fost observat la linia 23- 99419G4-1A/1-1 şi 25 - 00281G2-11.
Fig. 3. Produşii de amplificare obţinuţi cu primerii specifici FSD şi RSA la liniile individuale 23.1-23.9- plante individuale de la linia 99419G4-1A/1-1, 24.1-24.10 –linia 00274G2-31, 25.1-25.8 – linia 00281G2-11, 26.1-26.10 – linia 00399G2-11, 27.1-27.10 - linia 01450G1-1, soiurile sensibile 15 – Farmec, 16 – Delabrad, 21 – Crina şi linia sensibilă 17 – F96869G1-108 (gel de agaroză).

Rezumatul Tezei de Doctorat
15 15
Rezultatele analizei cu primerii specifici de la secară (F3/R3) (Katto şi colab., 2004) Majoritatea liniilor dihaploide analizate obţinute din combinaţia hibridă GP 369, precum şi una
dintre provenienţele de referinţă (21-soiul Crina) au prezentat banda de 1400 pb specifică genomului de secară (fig. 4), ceea ce înseamnă că în genealogia acestora a participat şi secara. Prezenţa unor secvenţe din genomul de secară la provenienţa considerată sensibilă indică faptul că printre grenele preluate de la secară nu sunt şi gene de rezistenţă la mălură
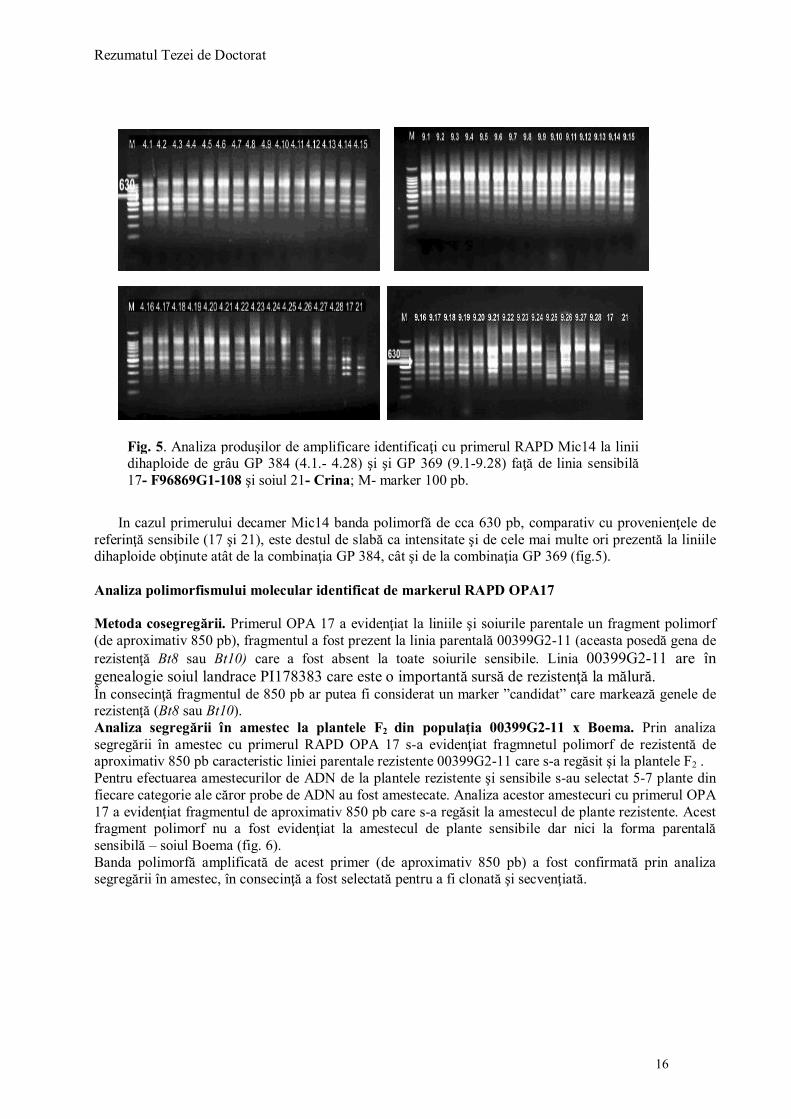
Absenţa benzii la cealaltă provenienţă de referinţă considerată, de asemenea, sensibilă (17-linia F96869G1-108), ne face să credem că materialul genetic provenit de la secară nu a integrat şi gene de rezistenţă la mălura comună. Remarcăm produsul de amplificare foarte bogat la liniile 9.7, 9.8, 9.9, 9.10, precum şi prezenţa altor benzi nespecifice. 7.4. REZULTATELE ANALIZEI CU MARKERII RAPD
Dintre cei 32 de primeri random testaţi un număr de 9 primeri au fost polimorfi. Un polimorfism ridicat s-a obţinut şi cu următorii primeri: UBC 570, Mic 07, Mic 13, OPH 20 (Coţa et al., 2007). O parte din primerii polimorfi au fost eliminaţi deoarece polimorfismul identificat nu a fost reproductibil. Polimorfismul identificat de markerii RAPD testaţi la formele parentale şi la liniile dihaploide pote fi sintetizat astfel:
Primerul OPA 17 a identificat fragmentul de 850 pb la linia parentală rezistentă 00399G2-11; Primerul OPA 16 a identificat fragmentul de 900 pb la linia parentală rezistentă 00274G2-31 şi
la liniile dihaploide; Primerul Mic 14 a identificat fragmentul de 630 pb la linia parentală rezistentă 99419G4-1A/1-1
şi la liniile dihaploide; Primerul OPC 10 a identificat fragmentul de 900 pb la linia parentală rezistentă 99419G4-1A/1-
1 dar şi la liniile dihaploide din combinaţia GP369; Primerul 70.04 a identificat fragmentul de 750 pb la linia parentală rezistentă 00281G2-11 dar şi
la populaţia F2 00281G2-11 x Boema; Primerul OPC 04 a identificat fragmentul de 550 pb doar la liniile parentale rezistente
numerotate 1 -99419G4-1A/1-1 şi 3- 00281G2-11. Analiza polimorfismul molecular identificat de markerii RAPD polimorfi la liniile dihaploide
Dintre primerii care au identificat polimorfism la liniile parentale (OPA 16 şi Mic 14) au fost testaţi apoi la toate liniile dihaploide analizate pentru a vedea cum se regăseste polimorfismul la aceste linii (Coţa şi colab., 2007).
Fig. 4. Produşii de amplificare obţinuţi cu primerii specifici pentru secară (F3/R3), la linii dihaploide GP 369 şi la provenienţele de referinţă 17 şi 21. L - marker 100 pb.
Rezumatul Tezei de Doctorat
16 16
In cazul primerului decamer Mic14 banda polimorfă de cca 630 pb, comparativ cu provenienţele de
referinţă sensibile (17 şi 21), este destul de slabă ca intensitate şi de cele mai multe ori prezentă la liniile dihaploide obţinute atât de la combinaţia GP 384, cât şi de la combinaţia GP 369 (fig.5). Analiza polimorfismului molecular identificat de markerul RAPD OPA17 Metoda cosegregării. Primerul OPA 17 a evidenţiat la liniile şi soiurile parentale un fragment polimorf (de aproximativ 850 pb), fragmentul a fost prezent la linia parentală 00399G2-11 (aceasta posedă gena de rezistenţă Bt8 sau Bt10) care a fost absent la toate soiurile sensibile. Linia 00399G2-11 are în genealogie soiul landrace PI178383 care este o importantă sursă de rezistenţă la mălură. În consecinţă fragmentul de 850 pb ar putea fi considerat un marker ”candidat” care markează genele de rezistenţă (Bt8 sau Bt10). Analiza segregării în amestec la plantele F2 din populaţia 00399G2-11 x Boema. Prin analiza segregării în amestec cu primerul RAPD OPA 17 s-a evidenţiat fragmnetul polimorf de rezistentă de aproximativ 850 pb caracteristic liniei parentale rezistente 00399G2-11 care s-a regăsit şi la plantele F2 . Pentru efectuarea amestecurilor de ADN de la plantele rezistente şi sensibile s-au selectat 5-7 plante din fiecare categorie ale căror probe de ADN au fost amestecate. Analiza acestor amestecuri cu primerul OPA 17 a evidenţiat fragmentul de aproximativ 850 pb care s-a regăsit la amestecul de plante rezistente. Acest fragment polimorf nu a fost evidenţiat la amestecul de plante sensibile dar nici la forma parentală sensibilă – soiul Boema (fig. 6). Banda polimorfă amplificată de acest primer (de aproximativ 850 pb) a fost confirmată prin analiza segregării în amestec, în consecinţă a fost selectată pentru a fi clonată şi secvenţiată.
Fig. 5. Analiza produşilor de amplificare identificaţi cu primerul RAPD Mic14 la linii dihaploide de grâu GP 384 (4.1.- 4.28) şi şi GP 369 (9.1-9.28) faţă de linia sensibilă 17- F96869G1-108 şi soiul 21- Crina; M- marker 100 pb.

Rezumatul Tezei de Doctorat
17 17
Fig. 6 . Produşi de amplificare obţinuţi cu primerul OPA17 la următoarele genotipuri: Pr - linia parentală rezistentă 00399G2-11, Ps - soiul sensibil Boema, Br- amestecul rezistent, Bs – amestecul de ADN sensibil, plante din generaţia F2 (1-16).
7.5. REZULTATE PRIVIND TRANSFORMAREA MARKERILOR RAPD ÎN MARKERI SPECIFICI DE TIP SCAR 7.5.1. Izolarea benzilor polimorfe RAPD din gelul de migrare. Au fost izolate din gel benzile polimorfe obţinute cu primerii OPA 17 şi OPC 04. Pentru a converti banda polimorfă de aproximativ 550 pb obţinută cu primerul OPC04 şi banda de 850 pb obţinută cu primerul OPA 17 în markeri de tip SCAR a fost necesară clonarea benzilor polimorfe. În imaginea de mai jos (fig. 7) sunt prezentate benzile recuperate de la produsul de amplificare polimorf (de 550 pb) amplificat de primerul OPC 04, şi a benzilor polimorfe obţinute cu primerul OPA 17 (cu dimensiunea de aproximativ 850 pb).
Fig. 7. Reamplificarea benzilor recuperate obţinute cu primerul OPC 04
7.5.2. Clonarea şi secvenţierea benzilor RAPD polimorfe.
Aceste benzi au fost clonate în bacteria Escherichia colli (GM2163). S-au analizat colonile care au integrat fragmentul clonat şi care au supravieţuit pe mediu selectiv cu ampicilină (fig. 8).
Fig. 8. Colonii bacteriene pe mediu selectiv care au integrat fragmentul clonat
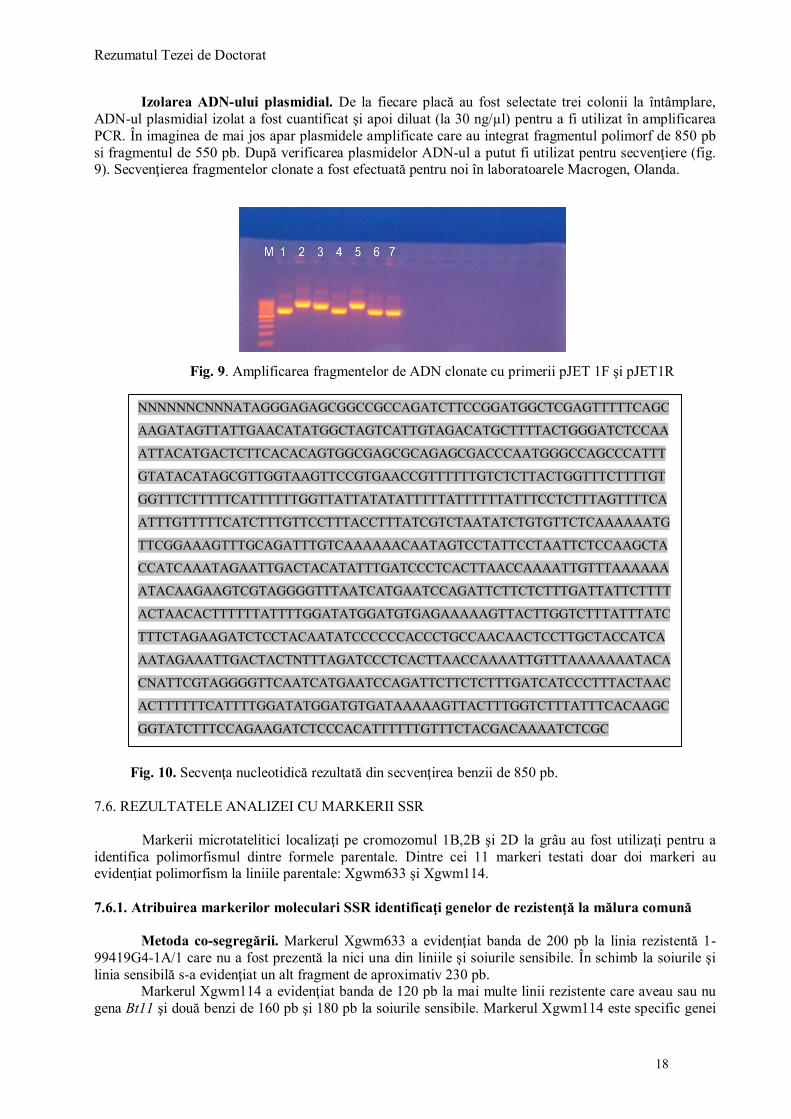
Rezumatul Tezei de Doctorat
18 18
Izolarea ADN-ului plasmidial. De la fiecare placă au fost selectate trei colonii la întâmplare, ADN-ul plasmidial izolat a fost cuantificat şi apoi diluat (la 30 ng/µl) pentru a fi utilizat în amplificarea PCR. În imaginea de mai jos apar plasmidele amplificate care au integrat fragmentul polimorf de 850 pb si fragmentul de 550 pb. După verificarea plasmidelor ADN-ul a putut fi utilizat pentru secvenţiere (fig. 9). Secvenţierea fragmentelor clonate a fost efectuată pentru noi în laboratoarele Macrogen, Olanda.
Fig. 9. Amplificarea fragmentelor de ADN clonate cu primerii pJET 1F şi pJET1R
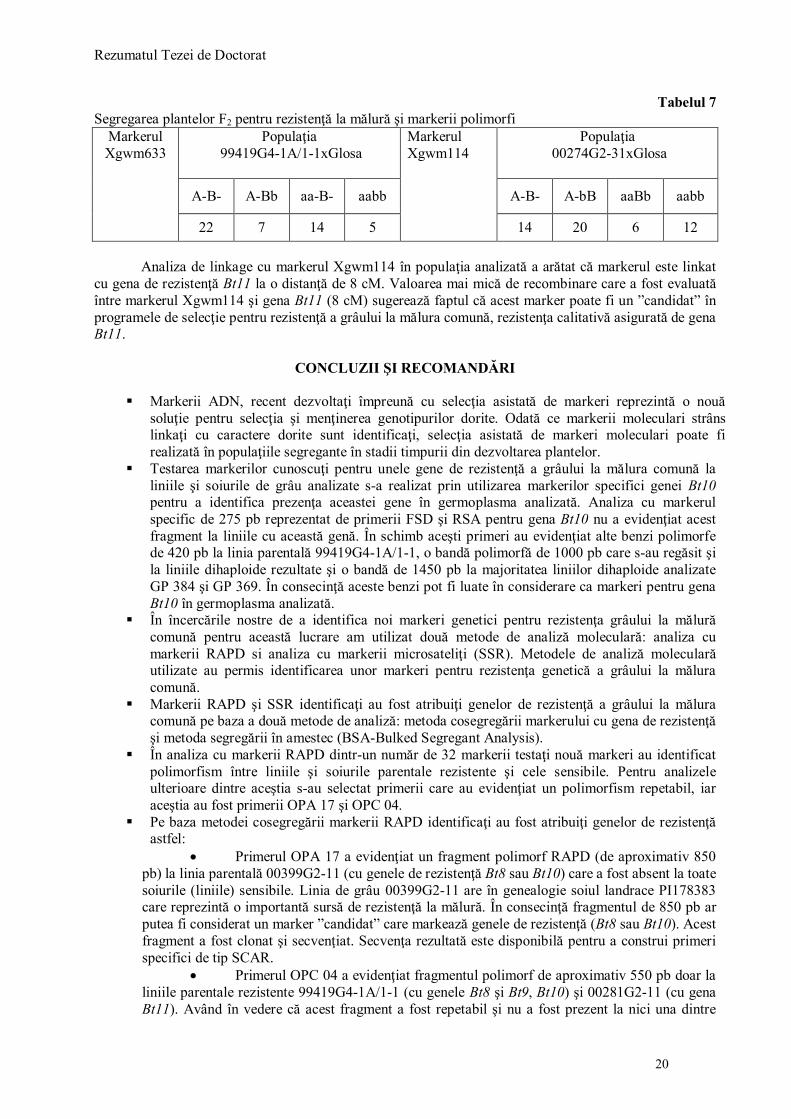
Fig. 10. Secvenţa nucleotidică rezultată din secvenţirea benzii de 850 pb.
7.6. REZULTATELE ANALIZEI CU MARKERII SSR
Markerii microtatelitici localizaţi pe cromozomul 1B,2B şi 2D la grâu au fost utilizaţi pentru a identifica polimorfismul dintre formele parentale. Dintre cei 11 markeri testati doar doi markeri au evidenţiat polimorfism la liniile parentale: Xgwm633 şi Xgwm114.
7.6.1. Atribuirea markerilor moleculari SSR identificaţi genelor de rezistenţă la mălura comună Metoda co-segregării. Markerul Xgwm633 a evidenţiat banda de 200 pb la linia rezistentă 1- 99419G4-1A/1 care nu a fost prezentă la nici una din liniile şi soiurile sensibile. În schimb la soiurile şi linia sensibilă s-a evidenţiat un alt fragment de aproximativ 230 pb. Markerul Xgwm114 a evidenţiat banda de 120 pb la mai multe linii rezistente care aveau sau nu gena Bt11 şi două benzi de 160 pb şi 180 pb la soiurile sensibile. Markerul Xgwm114 este specific genei
NNNNNNCNNNATAGGGAGAGCGGCCGCCAGATCTTCCGGATGGCTCGAGTTTTTCAGC
AAGATAGTTATTGAACATATGGCTAGTCATTGTAGACATGCTTTTACTGGGATCTCCAA
ATTACATGACTCTTCACACAGTGGCGAGCGCAGAGCGACCCAATGGGCCAGCCCATTT
GTATACATAGCGTTGGTAAGTTCCGTGAACCGTTTTTTGTCTCTTACTGGTTTCTTTTGT
GGTTTCTTTTTCATTTTTTGGTTATTATATATTTTTATTTTTTATTTCCTCTTTAGTTTTCA
ATTTGTTTTTCATCTTTGTTCCTTTACCTTTATCGTCTAATATCTGTGTTCTCAAAAAATG
TTCGGAAAGTTTGCAGATTTGTCAAAAAACAATAGTCCTATTCCTAATTCTCCAAGCTA
CCATCAAATAGAATTGACTACATATTTGATCCCTCACTTAACCAAAATTGTTTAAAAAA
ATACAAGAAGTCGTAGGGGTTTAATCATGAATCCAGATTCTTCTCTTTGATTATTCTTTT
ACTAACACTTTTTTATTTTGGATATGGATGTGAGAAAAAGTTACTTGGTCTTTATTTATC
TTTCTAGAAGATCTCCTACAATATCCCCCCACCCTGCCAACAACTCCTTGCTACCATCA
AATAGAAATTGACTACTNTTTAGATCCCTCACTTAACCAAAATTGTTTAAAAAAATACA
CNATTCGTAGGGGTTCAATCATGAATCCAGATTCTTCTCTTTGATCATCCCTTTACTAAC
ACTTTTTTCATTTTGGATATGGATGTGATAAAAAGTTACTTTGGTCTTTATTTCACAAGC
GGTATCTTTCCAGAAGATCTCCCACATTTTTTGTTTCTACGACAAAATCTCGC

Rezumatul Tezei de Doctorat
19 19
Bt11. Acest marker a confirmat prezenţa genei la genotipul liniei purtătoare a acestei gene (linia de grâu 00281G2-11) prin fragmentul polimorf de 120 pb. Totuşi fragmentul a apărut şi la alte linii rezistente (purtatoare a altor gene de rezistenţă) dar şi la două dintre liniile sensibile. Faptul că markerul polimorf a apărut la liniile care nu erau cunoscute ca având gena Bt11 arată că aceste linii posedă şi această genă de rezistenţă. Polimorfismul identificat de markerii a fost testat la populaţiile segregante F2. Cu markerul Xgwm633 a fost testată populaţia rezultată din încrucişarea liniei 99419G4-1A/1-1 la care s-a evidenţiat gena de rezistentă cu soiul sensibil Glosa. Cu markerul Xgwm114 a fost testată populaţia rezultată din încrucişarea liniei 00274G2-31 (cu gena Bt5 şi care a evidenţiat banda polimorfă de 120 pb) cu soiul sensibil Glosa. Metoda segregării în amestec (Bulk Segregant Analysis – BSA) Analiza populaţiei F2 99419G4-1A/1-1 x Glosa cu markerul Xgwm633 Această populaţie este formată dintr-un număr de 51 de plante, fiecare plantă a fost individualizată fenotipic pentru rezistenţa la mălura comună. ADN-ul a fost izolat de la toate plantele F2 şi supus analizei PCR utilizând primerul Xgwm633 şi a fost testată genotipic cu acest marker. Markerul Xgwm633 este cunoscut ca fiind linkat cu loci QTL asociaţi cu rezistenţa la mălură (Fofana şi colab., 2008). În analizele efectuate la plantele F2 markerul a evidenţiat cele două fragmente polimorfe ca şi la liniile parentale de 200 pb şi 230 pb. Gena de rezistenţă prezentă la linia de grâu 99419G4-1A/1-1 nu este cunoscută dar se presupune că ar fi gena Bt8 sau Bt10. Prin analiza segregării în amestec cu markerul Xgwm633 s-a confirmat prezenţa celor două fragmente polimorfe cel de 200 pb pentru amestecul de ADN de la plantele rezistente şi cel de 230 pb pentru amestecul provenit de la plantele sensibile Analiza populaţiei F2 00274G2-31 x Glosa cu markerul Xgwm114 Această populaţie a fost formată dintr-un număr de 52 de plante. Fragmentul polimorf pentru rezistenţă de 120 pb s-a regăsit în populaţia F2 la multe genotipuri individuale care erau rezistente fenotipic dar şi la pante individuale F2 care erau sensibile fenotipic. De asemenea markerul pentru alela de sensibilitate s-a regăsit la multe plante F2 sensibile sau nu fenotipic. Analiza segregării în amestec. Amestecurile au fost efectuate la fel ca şi pentru markerul Xgwm633 în populaţia analizată. Markerul Xgwm114 a arătat la amestecul de ADN al plantelor sensibile acelaşi fragmente polimorfe de sensibilitate de 160 pb şi 180 pb ca şi la linia parentală sensibilă. Iar la amestecul de ADN de la plantele rezistente pe lângă fragmentul polimorf de rezistenţă la dimensiunea de 120 pb s-au evidenţiat şi cele două fragmente de sensibilitate caracteristice formei parentale sensibile. Acest lucru poate fi explicat prin faptul că în alegerea plantelor pentru crearea amestecurilor pe lângă plantele rezistente s-a selectat şi o plantă sensibilă dar care nu manifestat sensibilitatea fenotipic. Trebuie menţionat faptul că fragmentul de 120 pb specific liniei rezistente a fost raportat de asemenea de către Cichy şi Goates (2009), (Cichy K., comunicare personală, April 20, 2010). Dar la genotipurile sensibile ei au identificat un singur fragment cu dimensiunea de 110 pb. Analiza de linkage. Markerii SSR testati in acest studiu au fost utlizati pentru a calcula distanţa de înlăţuire dintre markeri şi genele de rezistenţă. Rezultatele privind segregarea pentru rezistenţă şi cei doi markeri SSR polimorfi (Xgwm633 şi Xgwm114) în populaţiile segregante F2 sunt prezentate în tabelul 7.
Analiza de linkage cu markerul Xgwm633 în populalaţia analizată a aratat că markerul este linkat la o distanţă destul de mare cu gena de rezistenţă de 32 cM. Această distanţa mare este datorată faptului că markerul Xgwm633 este linkat cu un QTL asociat cu rezistenţă la mălura comună în populaţia analizată, rezistenţă controlată de poligene.
Rezumatul Tezei de Doctorat
20 20
Tabelul 7 Segregarea plantelor F2 pentru rezistenţă la mălură şi markerii polimorfi
Populaţia 99419G4-1A/1-1xGlosa
Populaţia 00274G2-31xGlosa
A-B- A-Bb aa-B- aabb A-B- A-bB aaBb aabb
Markerul Xgwm633
22 7 14 5
Markerul Xgwm114
14 20 6 12 Analiza de linkage cu markerul Xgwm114 în populaţia analizată a arătat că markerul este linkat cu gena de rezistenţă Bt11 la o distanţă de 8 cM. Valoarea mai mică de recombinare care a fost evaluată între markerul Xgwm114 şi gena Bt11 (8 cM) sugerează faptul că acest marker poate fi un ”candidat” în programele de selecţie pentru rezistenţă a grâului la mălura comună, rezistenţa calitativă asigurată de gena Bt11.
CONCLUZII ŞI RECOMANDĂRI
Markerii ADN, recent dezvoltaţi împreună cu selecţia asistată de markeri reprezintă o nouă soluţie pentru selecţia şi menţinerea genotipurilor dorite. Odată ce markerii moleculari strâns linkaţi cu caractere dorite sunt identificaţi, selecţia asistată de markeri moleculari poate fi realizată în populaţiile segregante în stadii timpurii din dezvoltarea plantelor.
Testarea markerilor cunoscuţi pentru unele gene de rezistenţă a grâului la mălura comună la liniile şi soiurile de grâu analizate s-a realizat prin utilizarea markerilor specifici genei Bt10 pentru a identifica prezenţa aceastei gene în germoplasma analizată. Analiza cu markerul specific de 275 pb reprezentat de primerii FSD şi RSA pentru gena Bt10 nu a evidenţiat acest fragment la liniile cu această genă. În schimb aceşti primeri au evidenţiat alte benzi polimorfe de 420 pb la linia parentală 99419G4-1A/1-1, o bandă polimorfă de 1000 pb care s-au regăsit şi la liniile dihaploide rezultate şi o bandă de 1450 pb la majoritatea liniilor dihaploide analizate GP 384 şi GP 369. În consecinţă aceste benzi pot fi luate în considerare ca markeri pentru gena Bt10 în germoplasma analizată.
În încercările nostre de a identifica noi markeri genetici pentru rezistenţa grâului la mălură comună pentru această lucrare am utilizat două metode de analiză moleculară: analiza cu markerii RAPD si analiza cu markerii microsateliţi (SSR). Metodele de analiză moleculară utilizate au permis identificarea unor markeri pentru rezistenţa genetică a grâului la mălura comună.
Markerii RAPD şi SSR identificaţi au fost atribuiţi genelor de rezistenţă a grâului la mălura comună pe baza a două metode de analiză: metoda cosegregării markerului cu gena de rezistenţă şi metoda segregării în amestec (BSA-Bulked Segregant Analysis).
În analiza cu markerii RAPD dintr-un număr de 32 markerii testaţi nouă markeri au identificat polimorfism între liniile şi soiurile parentale rezistente şi cele sensibile. Pentru analizele ulterioare dintre aceştia s-au selectat primerii care au evidenţiat un polimorfism repetabil, iar aceştia au fost primerii OPA 17 şi OPC 04.
Pe baza metodei cosegregării markerii RAPD identificaţi au fost atribuiţi genelor de rezistenţă astfel:
Primerul OPA 17 a evidenţiat un fragment polimorf RAPD (de aproximativ 850 pb) la linia parentală 00399G2-11 (cu genele de rezistenţă Bt8 sau Bt10) care a fost absent la toate soiurile (liniile) sensibile. Linia de grâu 00399G2-11 are în genealogie soiul landrace PI178383 care reprezintă o importantă sursă de rezistenţă la mălură. În consecinţă fragmentul de 850 pb ar putea fi considerat un marker ”candidat” care markează genele de rezistenţă (Bt8 sau Bt10). Acest fragment a fost clonat şi secvenţiat. Secvenţa rezultată este disponibilă pentru a construi primeri specifici de tip SCAR.
Primerul OPC 04 a evidenţiat fragmentul polimorf de aproximativ 550 pb doar la liniile parentale rezistente 99419G4-1A/1-1 (cu genele Bt8 şi Bt9, Bt10) şi 00281G2-11 (cu gena Bt11). Având în vedere că acest fragment a fost repetabil şi nu a fost prezent la nici una dintre

Rezumatul Tezei de Doctorat
21 21
liniile şi soiurile sensibile îl considerăm un marker ”candidat” pentru genele de rezistenţă prezente la linia 99419G4-1A/1-1 şi 00281G2-11.
Pe baza metodei de segregare în amestec s-a analizat prezenţa markerului de 850 pb cu primerul OPA 17 în populaţia F2 00399G2-11 x Boema. Polimorfsmul evidenţiat cu acest primer (framentul de 850 pb) a fost confirmat prin analiza segregării în amestec. Astfel, acest marker polimorf poate fi atribuit genelor de rezistenţă (Bt8 sau Bt10) şi utilizat în selecţia pentru rezistenţa grâului la mălura comună.
Obiectivul propus de a transforma markerii RAPD în markerii specifici de tip SCAR a fost îndeplinit. S-a încercat conversia a doi dintre markerii evidentiaţi de primerii RAPD (OPA 17 şi OPC 04) în markeri specifici de tip SCAR. Fragmentele polimorfe au fost clonate si secvenţiate. Pe baza datelor obţinute în urma secvenţierii markerilor RAPD polimorfi se pot construi primeri specifici care sunt codominanţi şi care să fie utilizaţi în analiza cu markerii SCAR.
Markerii SSR au fost identificaţi în număr foarte mare la grâul comun şi au un grad ridicat de polimorfism ceea ce îi face extrem de utili petru identificarea de noi markeri care să fie utilizaţi în selectie, respectiv pentru selecţia asistată de markeri. În analiza cu markerii microsatelitici au fost utilizati în acest studiu markerii SSR care controleaza rezistentă la mălură reprezentaţi de 11 perechi de primeri SSR. Markerii SSR care au evidentiat polimorfism între formele parentale rezistente şi cele sensibile au fost markerul Xgwm633 şi Xgwm114.
Pe baza metodei cosegregării markerii SSR au fost atribuiţi genelor de rezistenţă astfel: Markerul Xgwm633 este asociat cu loci QTL de rezistenţă la mălură, în analizele efectuate
a evidenţiat banda de 200 pb la linia rezistentă 1- 99419G4-1A/1-1 care nu a fost prezentă la nici una din liniile şi soiurile sensibile.
Markerul Xgwm114 a evidenţiat banda de 120 pb la mai multe linii de grâu rezistente care aveau sau nu gena Bt11 şi două benzi de 160 pb şi 180 pb la soiurile sensibile. Cu acest marker s-a confirmat prezenţa genei la genotipul liniilor de grâu purtătoare a acestei gene (00281G2-11 şi 01450G1-1). Acest marker a evidenţiat prezenţa genei de rezistenţă Bt11 şi la alte genotipuri purtătoare a altor gene (Bt8, Bt5, Bt8). Aceşti doi markeri identificati au fost utilizaţi pentru analiza rezistenţei în populaţiile F2.
Prin metoda segregării în amestec markerii SSR au fost atribuiţi genelor de rezistenţă astfel: Markerul Xgwm633 a evidenţiat cele două fragmente polimorfe ca şi la liniile parentale
de 200 pb pentru rezistenţă şi 230 pb pentru alela de sensibilitate. Gena de rezistenţă prezentă la linia de grâu 99419G4-1A/1-1 nu este cunoscută dar se presupune că ar fi gena Bt8 sau Bt10. Cu acest marker s-a confirmat prezenţa celor două fragmente polimorfe cel de 200 pb pentru amestecul de ADN de la plantele rezistente şi cel de 230 pb pentru amestecul provenit de la plantele sensibile. Prin urmare acest marker poate fi atribuit unui locus QTL asociat cu rezistenţa la mălura comună.
Markerul Xgwm114 a arătat la amestecul de ADN al plantelor sensibile acelaşi fragmente polimorfe de sensibilitate de 160 pb şi 180 pb ca şi la linia parentală sensibilă, iar la amestecul de ADN de la plantele rezistente pe lângă fragmentul polimorf de rezistenţă la dimensiunea de 120 pb s-au evidenţiat şi cele două fragmente de sensibilitate caracteristice formei parentale sensibile.
Analiza de linkage dintre aceşti doi markeri SSR şi genele pe care aceştia le markează efectuată în populaţiile F2 nu a evidenţiat un linkage strâns cu genele de rezistenţă.
Markerul Xgwm114 a identificat un linkage mai strâns (8 cM) decât celălalt marker Xgwm633 (32 cM). Astfel, acest marker polimorf poate fi utilizat in selecţia asistată de markeri moleculari pentru rezistenţa grâului la mălura comună, rezistenţă calitativă asigurată de gena Bt11.
Distanţa mare de înlănţuire dintre markerul Xgwm633 şi gena de rezistenţă se explică prin faptul că şi în analizele efectuate în acest studiu acest marker este asociat cu un QTL de rezistenţă a grâului la mălura comună.
Rezultatele obţinute prin utilizarea metodei de segregare în amestec au arătat că această metodă este foarte eficientă pentru identificarea de markeri de rezistenţă la mălura comună a grâului.
În vederea identificării de markeri de rezistenţă a grâului la mălura comună s-au obţinut de la liniile şi soiurile parentale prin încrucişare un număr de zece populaţii F2 care au fost caracterizate fenotipic pentru rezistenţă prin infecţie artificială.

Rezumatul Tezei de Doctorat
22 22
Rezultatele obţinute în urma infecţiei artificiale cu spori de mălură comună la cele zece populaţii analizate a arătat că există diferenţe semnificative între populaţiile testate în ceea ce priveşte rezistenţa la mălura comună.
Analiza segregării fenotipice în populaţiile F2 a arătat că rezistenţa liniilor parentale este controlată de o singură genă de rezistenţă.
Având în vedere importanţa mălurii comune în cultura grâului se recomandă intensificarea cercetărilor privind măsurile de combatere, iniţierea unor programe de amelioare pentru rezistenţă.
Markerii moleculari identificaţi în lucrarea de faţă se recomandă a fi testaţi şi utilizaţi în programele de ameliorare a soiurilor de grâu. Utilizarea lor în combinaţie cu selecţia fenotipică vor îmbunătăţi procesul de amelioare a grâului pentru rezistenţa la mălura comună.
Se recomandă, de asemenea, pentru cercetările viitoare identificarea unui număr cât mai mare de markeri pentru rezistenţa grâului la mălura comună, markeri care să poată fi utilizaţi în selecţie.
BIBLIOGRAFIE SELECTIVĂ
1. Arumuganathan, E., and E. D. Earle, 1991, Nuclear DNA content of some important plant species, Plant Mol. Biol. Rep. 9, 208-218.
2. Bobeş, I., Al. Lazăr, I. Comes, A. Drăcea, M. Hatman, 1977, Fitopatologie, Editura Didactică Pedagogică, Bucureşti.
3. Bonman, J. M., H. Bockelman, B. Goates, D. Obert, P. McGuire, C., Qualset, and R. Hijmans, 2006, Geographic distribution of common and dwarf bunt resistance in landraces of Triticum aestivum subsp. aestivum. Crop Sci. 46:1622-1629.
4. Cichy Karen and B. Goates, 2009, Evaluation of molecular markers for common bunt resistance genes in diverse wheat genotypes. Abstract available at: http://a-c-s.confex.com/crops/2009am/webprogram/Paper54548.html (accessed February 2010).
5. Cichy Karen., personal communication, April 20, 2010. 6. Collard, B. C. Y., M. Z. Z. Jahufer, J. B. Brouwer and E. C. K. Pang, 2005, An introduction
to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts, Euphytica 142:169-196.
7. Coţa Laura Cristina, C. Botez, Dana Curticiu, Meda Lucaci, 2007, Genetic marker assisted selection for common bunt resistance (Tilletia spp.) in some wheat lines, Buletin USAMV-CN, 63: 237-242.
8. Coţa Laura Cristina, C. Botez, M. A. Grigoraş and Dana Curticiu, 2009, Screening for resistance to artificial infection by common bunt (Tilletia caries and Tilletia foetida) in F2 populations of wheat (Triticum aestivum L.), Bull. Univ. Agric. Sci. Vet. Med. (USAMV) 66:24-31.
9. Dumalasová Veronika and P. Bartoš, 2006a, Wheat reaction to common bunt in the field and in the greenhouse, Proc. XVth Biennial Workhop on the Smut Fungi, Prague, June 11-14, 2006. In: Czech J. Genet. Plant Breed. 42. 2006 (Special issue) pp. 37-41.
10. Dumalasová Veronika and P. Bartoš, 2006b, Resistance of winter wheat cultivars to common bunt, Tilletia tritici (Bjerk.) Wint. and T. laevis Kühn, Journal of Plant Diseases and Protection 113(4): 159-163.
11. Dumalasová Veronika and P., Bartoš, 2007, Reaction of winter wheat cultivars to common bunt, Tilletia tritici (Bjerk.) Wint. and T. laevis Kühn, Plant Protect. Sci., 43: 138–141.
12. Dumitraş Lucreţia şi Vera Bontea, 1991, Specii de Tilletia din România: Taxonomie, ultrastructură, biologie, combatere, Editura Academiei Române, Bucureşti.
13. Feldman, M., 2001, The origin of cultivated wheat. In Bonjean, A.P., and W.J. Angus Eds, The World Wheat Book: a history of wheat breeding, Lavoisier Publishing, Paris, 1131 pp.
14. Ganal, M. W. and M. S. Röder, 2007, Microsatellite and SNP markers in wheat breeding In: R. K. Varshney and R. Tuberosa, Eds., Genomics Assisted Crop Improvement: Vol. 2: Genomics Applications in Crops, Springer Netherlands, 2007, pp. 1-24.

Rezumatul Tezei de Doctorat
23 23
15. Ghibu H., 2002, Cercetări privind răspândirea, ciclul biologic şi combaterea mălurii pitice (T. controversa) şi a mălurii comune (T. spp.) a grâului în judeţul Cluj. Teză de doctorat, USAMV Cluj-Napoca.
16. Goates, B. J., 1996, Common bunt and dwarf bunt, In: R.D. Wilcoxson and E.E. Saari, Eds., Bunt and smut diseases of wheat: concepts and methods of disease management, Mexico, DF, CYMMIT, p.12-25.
17. Hoffmann, J. A. and R. J. Metzger, 1976, Current status of virulence genes and pathogenic races of the wheat bunt fungi in the north-western USA, Phytopatology 66:657-660.
18. Hoffmann, J. A., 1982, Bunts of wheat, Plant Disease 66: 979-987. 19. Huber, K., and H. Buerstmayr, 2006, Development of methods for bunt resistance breeding for
organic farming, Czech J. Genet. Plant Breed., 42, Proc. XVth Biennial Workshop on the Smut Fungi, Prague, June 11–14.
20. Katto, M. C., Endo T. R., Nasuda S., 2004, A PCR based marker for targeting small rye segments in wheat background, Genes & Genetics System, Vol 79, Nr. 4 p. 245-250.
21. Kendrick, E. L. and C. S. Holton, 1961, Racial population dinamics in Tilletia carries and Tilletia foetida as influenced by wheat varietal populations in the Pacific Northwest, Plant. Dis. Rep. 45:5-9.
22. Landjeva Svetlana, V. Korzun, and A. Börner, 2007, Molecular markers: actual and potential contributions to wheat genome characterization and breeding, Euphytica 156:271-296.
23. Langridge, P., E. S. Lagudah, T. A. Holton, R. Appels, P. J. Sharp, K. J. Chalmers, 2001, Trends in genetic and genome analyses in wheat: a review. Aust. J. Agric. Res. 52:1043–1077.
24. Laroche, A., T. Demeke, D. A. Gaudet, B. Puchalscki, M. Frick, and R. Mckenzie, 2000, Development of the PCR marker for rapid identification of the Bt-10 gene for common bunt resistance in wheat, Genome 43, 217-223.
25. Lodhi M. A., G. N. Ye, N. F. Weeden, B. I Reisch, 1994, A simpel and efficient method for DNA extraction from grapevine cultivars, Vitis species and Ampelopsis. Plant Molecular Biology Reporter 12 (1), pag 6-13.
26. Mamluk, O.F. & M. Nachit, 1988. Performance and reaction of some durum wheat genotypes against different isolates of common bunt (Tilletia foetida and T. caries). In: Abstracts of papers, the 5th International Congress of Plant Pathology.
27. Mamluk, O. F., 1998, Bunts and smuts of wheat in North Affica and the Near East, Euphytica 100:45–50.
28. Matanguihan Janet Barroga, K. M. Murphy, S. S. Jones, 2011, Control of common bunt in organic wheat, Plant Dis. 95, 92-103.
29. McIntosh, R. A., 1998, Breeding wheat for resistance to biotic stresses, Euphytica 100: 19–34. 30. Michelmore, R. W., I. Paran, and R. V. Kesseli, 1991, Identification of markers linked to
disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations, Proc. Natl. Acad. Sci. USA 88, 9828-9832.
31. Mozgovoi, A. F., I. Y. Saakyan and M. V. Novikova, 1987, Gene pool of sources and donors for breeding winter bread wheat for immunity to bunt, Vavilova 176:11-15.
32. Nagy Elena and V. Moldovan, 2007, The effect on fungicides treatments on the wheat common bunt (Tilletia spp.) in Transylvania – Romania, Romanian Agricultural Research 24: 33-38.
33. Oncică Fraga and N. N. Săulescu, 2007. Sources of resistance to bunt (Tilletia spp.) In modern Semidwarf winter wheat (Triticum aestivum L.). Romanian agricultural research, 24: 29-32.
34. Oncică Fraga and N. N. Săulescu, 2008, Potentially new sources of genes for resistance to common bunt (Tilletia spp.) in winter wheat (Triticum aestivum L.), Proc. Rom. Acad., Series B, 2008, 1–2, p. 97–100.
35. Pop Rodica, M. Ardelean, D. Pamfil, Ioana Marina Gaboreanu, 2003, The efficiency of different DNA isolation and purification protocols in ten cultivars of Vitis Vinifera, Bul. Nr. 59 USAMV, seria ZB, 259-261.
36. Röder MS, Korzun V, Wendehake K, Plaschke J, Tixier M-H, Leroy P, Ganal MW (1998) - A microsatellite map of wheat. Genetics 149:2007–2023.
37. Rubiales, D., R. E. Niks, T. L Carver, J. Ballesteros and A. Martin, 2001, Prospects for exploitation of disease resistance from H. chilense in cultivated cereals, Hereditas 135:161-169.

Rezumatul Tezei de Doctorat
24 24
38. Saari, E. E., O. F. Mamluk, and P. A. Burnett, 1996, Bunts and smuts of wheat p. 1–11. In: R.D. Wilcoxson and E.E. Saari (ed.), Bunt and smut diseases of wheat: Concepts and methods of disease management. CIMMYT. Mexico, D.F.
39. Sandu-Ville C., 1934, Anal ICAR VI, 324-352. 40. Săvulescu, T., 1957, Ustilaginalele din R.P.R, Ed. Academiei Române, I, II, p. 1168. 41. Wächter Rebecca, Franziska Waldow, K. J. Müller, H. Spieß, B. Heyden, Ursula Furth, J.
Frahm, W. Weng, T. Miedaner, D. Stephan and E. Koch, 2007, Resistance of winter wheat varieties and breeding lines against common bunt (Tilletia tritici) and dwarf bunt (T. controversa), Nachrichtenbl. Deut. Pflanzenschutzd. 59(2):30-39.