studia universitatis “vasile goldiş”, seria Ştiinţele ... 21su-2015-lb- 65-71.pdf ·...
TRANSCRIPT

Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25 issue 2, 2015, pp.65-71
© 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
Correspondence: * Constantin Craciun, Ph. D., Babes-Bolyai University Cluj-Napoca, Electron Microscopy Center, 5-7 Clinicilor Street, Cluj-Napoca 400006, Romania, Phone: +40(0)262722, e-mail: [email protected] Article published: May 2015
ANATOMICAL, MORPHOLOGICAL AND CYTOLOGICAL
COMPARATIVE STUDY OF LEAVES AND COTYLEDONS FROM
FORESTRY SPECIES
II. COMPARISON BETWEEN THE MORPHO-ANATOMICAL AND
CYTOLOGICAL STRUCTURES OF COTYLEDONS AND LEAVES OF
ROBINIA PSEUDOACACIA L. Liviu BURESCU
1, Dorina CACHIŢA
2, Constantin CRACIUN
3)*
1University of Oradea, Faculty of Sciences;
2„V. Goldiş” University from Arad, Faculty of Sciences, Engeniering and Informatics;
3„Babeş-Bolyai” University from Cluj-Napoca, Electron Microscopy Center;
ABSTRACT: The role of cotyledons in seed physiology depends on the species it is experimented with. The black locust (Robinia pseudoacacia L) has an embryo with two epigeal cotyledons that, after the plant rises, turn green and their metabolism switches from a heterotrophic regime to an autotrophic photosynthetic process. The cotyledons supply the embryo with water, nutrients and energy that helps the plant to break the seed coat and the hypocotyl together with the cotyledons to raise above the ground. The life span of the cotyledons is approximately 40 days depending on the environmental conditions. After the black locust plantlets emerge, the epycotyl rises between the two cotyledons and produce odd pinnate leaves. They have elliptical-shaped folioles about 2.5 cm in lenght. The folioles are thinner than the cotyledons. At about 14 days after the onset of germination, the cotyledons have an average lenght of 1 cm and a thickness of about 3 - 4 mm. The foliar lamina is crossed by a main visible midvein that is well defined on their lower side. This morpho-anatomical formation is missing in cotyledons. The secondary veins are rare and thin. The folioles epidermis has stomata and the inferior one has live unicellular hairs. Between the two epidermises the foliole laminas have an assimilation mesophyll made of 5 – 7 layers of parenchyma cells. Here is also located the hypoderm whose cells contain secondary metabolites that are stained in blue with „Epoxy tissue stain”. The cotyledons have an elongated shape. The outline of the transversal sections is plano convex, the upper epidermis being planar and the lower epidermis is slightly convex. The cotyledon veins are scarce and are very fine. The cotyledons have a very well ordered tissue structure.
The upper and lower epidermis has rectangle shaped cells and both have only pre-stomata. Between the two epidermises there are 16 -20 assimilating parenchyma layers, the one right below the epidermis is of palisade type.
From a cytological point of view, the assimilating mesophyll of the cotyledons is poor in chloroplasts but both the foliary and cotyledonary mesophyll have in their vacuolar content secondary metabolites which toghether with the staining reagents, aggregate leading to corpuscular formations or electron-dense vacuolar structures.
Keywords: Robinia pseudoacacia L; leaf, cotyledons; morpho-anatomical structure.
INTRODUCTION:
The study of the cotyledons structure was generally
ignored by plant morphoanatomists because these
characteristic organs of embryos and plantlets
respectively although they play an important role in the
first weeks of life, they have a short but intense life.
In the plant kingdom, there are a diversity of
cotyledons depending on the plant species.
Taxonomically there are multicotyledonous (such as
gymnosperms), dicotyledonous (dicots) (for example
rosaceae, leguminosae, etc), monocotyledonous
(monocots) (for example gramineae) and plants that
have embryos with rudimentary cotyledons (such as
orchids and some parasitic plants).
Generally, in seed physiology, the cotyledons play
an important role in triggering the germination and in
embryo and plantlet growth. The role of the cotyledon
depends on the seed structure, if the cotyledon has
stored food reserves or if is in the endosperm
surrounding the cotyledons (for example at
gymnosperms or castor oil plant) or - at monocot plants
– the only cotyledon of the embryo is interposed
between the endosperm and the embryo.
At many dicot species, the two cotyledons of the
embryo are placed at the interface between the
hypocotyl and epicotyl.
Depending on the species, the cotyledons are
epigeal – when the hypocotyl raises above the ground-
or hypogeal when the cotyledons remain below ground.
Furthermore, there are species such as the beech or
cucumber at which the cotyledons – that are epigeal –
raise above ground turn green and became
photosynthetic. At other species such as beans, the
cotyledons will shrivel and fall shortly after raising
above the ground. Of the dicot plants, the black locust
was choosen to examine the morpho anatomical
structure of the cotyledons and to compare it with the
one the leaves of this species

Burescu L., Cachiţă D., Crăciun C.
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71
© 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
66
At the black locust (Robinia pseudoacacia L), the
cotyledons are epigeal (Fig. 1 B), plano convex (with
the planary part orientated upwards and with the curved
part located below); they are green and about 3 - 4 mm
thick. The physiological role of the black locust
cotyledons is different from that of the albuminous
seeds because the storage parenchyma is lacking the
endosperm.
At few dicot plants – for example castor oil plant –
the cotyledons are foliaceous and are surrounded by an
endosperm that provides nutrients to the embryo. Most
of the dicot species lack endosperm and the stored
reserved substances are in the cotyledons.
After the black locust plant raises above the ground,
the cotyledons turn green and begin photosynthesis.
Thus, as in other species, the black locust cotyledons
have a complex role as long as the true leaves have not
yet emerged, situation hat involves a transformation of
their morphoanatomy. The black locust leaves are odd
pinnate (Fig. 1 A) and a foliole is more than double
compared with the cotyledon and as observed has a
different structural makeup than that of the folioles.
Fig. 1 A şi B. Morphological aspect of a odd pennate,
young black locust (Robinia pseudoacacia L) leaf (A);(B) black locust plantlet photographed on the 14
th
day of seeds germination in a plastic container on filter paper moistened with tap water (abbreviations: c (rst) – root-stem transition region; cot – cotyledons; ep – epicotyl; f – foliole; re – embryonary root; h – hypocotyl; b – budlet; R - rachis
In plant biotechnology, the explants taken from the
embryos (Cachiţă et al., 2004) and the plantlets
respectively, are valorised either as phyto – innocules
with high regenerative capacity or as experimental
models to examine their reaction as a function of
vitroculture conditions. Interesting are the
micrografting researches with the purpose of grafting
the top of the vitrocultivated plantlets when an apical
part of the hypocotyl with the pertaining cotyledons as
well as the epicotyl, formation that is used as a graft,
that is implanted in a parent stock (according to
Turnbull et al., 2002) (Fig. 2).
This operation is performed „in vitro” and there is a
good chance to obtain from the graft virus free plants
including the woody plant species at which the disease
free plants regeneration – by meristem culture
techniques – is extremely difficult.
Fig. 2 A – D. Micrografting technique of a plantlet
performed under aseptic conditions (according to Turnbull et al., 2002 –modified image) A – plantlet from
a sterile environment ( the arrow indicates the place where the plantlet was sectioned for detaching the graft); B „a” – trimming of the graft basal area and “b” splitting of the parent stock to introduce the graft; C –
placement of the graft on a parent stock ( with the apex removed and with an “infections free” certificate) D –
grafting zone (gz) can be consolidated with an autoclavable silicone sleeve (ss) (abbreviations: g – graft; ct – cotyledons; s – split; h – hypocotyls; b – budlet; pa – parent stock; Re – embryonary root; B „a” – graft; B „b” – parent stock; D „a” – fixation of the grafting area with a sterile silicone sleeve; D „b” – grafted area after the removal of the silicone sleeve)
The advantage of cotyledon-type phyto-innocules
in not only in the youthfulness of the tissues that make
up this type of explant but also in the fact that it’s cells
have nutrient reserves necessary to start and sustain the
growth processes of the embryo to which the cotyledon
pertains. This is very important mainly in the case of
vitrocultivated forestry species that can be hard to
propagate using the meristem culture technique
(Enescu et al., 1994).
In the literature (Stănescu, 1979), there are no
studies regarding the resemblances or differences
between the anatomy of the black locust cotyledons an
leaves. Therefore, the knowledge of these aspects is
important for explant prelevation in vitroculture
techniques.
MATERIALS AND METHODS: Black locust seeds were placed for germination in
plastic containers (colorless and transparent) on a filter
paper moistened with tap water, at room temperature.
The germination was carried out for 14 days at 23ºC,
the plantlets being exposed to a day/night cycle specific
for June, period in which the experiments were
performed. In the 14th
day of germination, the
cotyledons were taken from the plantlets (Fig. 1 B)
whereas the black locust folioles were take from
nature.
Anatomical, morphological and cytological comparative study of leaves and cotyledons from forestry species II. Comparison between the the morpho-anatomical and cytological structures
of cotyledons and leaves of black locust (Robinia pseudoacacia l)
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71 © 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
67
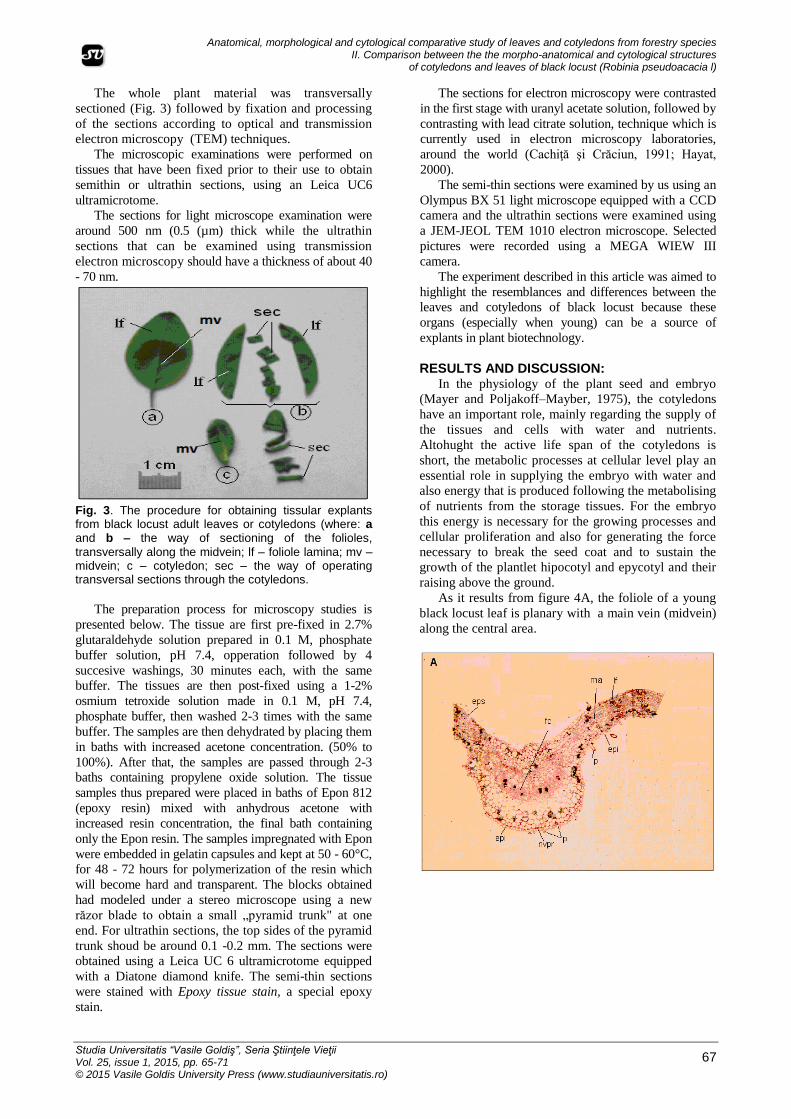
The whole plant material was transversally
sectioned (Fig. 3) followed by fixation and processing
of the sections according to optical and transmission
electron microscopy (TEM) techniques.
The microscopic examinations were performed on
tissues that have been fixed prior to their use to obtain
semithin or ultrathin sections, using an Leica UC6
ultramicrotome.
The sections for light microscope examination were
around 500 nm (0.5 (µm) thick while the ultrathin
sections that can be examined using transmission
electron microscopy should have a thickness of about 40
- 70 nm.
Fig. 3. The procedure for obtaining tissular explants from black locust adult leaves or cotyledons (where: a and b – the way of sectioning of the folioles,
transversally along the midvein; lf – foliole lamina; mv – midvein; c – cotyledon; sec – the way of operating transversal sections through the cotyledons.
The preparation process for microscopy studies is
presented below. The tissue are first pre-fixed in 2.7%
glutaraldehyde solution prepared in 0.1 M, phosphate
buffer solution, pH 7.4, opperation followed by 4
succesive washings, 30 minutes each, with the same
buffer. The tissues are then post-fixed using a 1-2%
osmium tetroxide solution made in 0.1 M, pH 7.4,
phosphate buffer, then washed 2-3 times with the same
buffer. The samples are then dehydrated by placing them
in baths with increased acetone concentration. (50% to
100%). After that, the samples are passed through 2-3
baths containing propylene oxide solution. The tissue
samples thus prepared were placed in baths of Epon 812
(epoxy resin) mixed with anhydrous acetone with
increased resin concentration, the final bath containing
only the Epon resin. The samples impregnated with Epon
were embedded in gelatin capsules and kept at 50 - 60°C,
for 48 - 72 hours for polymerization of the resin which
will become hard and transparent. The blocks obtained
had modeled under a stereo microscope using a new
răzor blade to obtain a small „pyramid trunk" at one
end. For ultrathin sections, the top sides of the pyramid
trunk shoud be around 0.1 -0.2 mm. The sections were
obtained using a Leica UC 6 ultramicrotome equipped
with a Diatone diamond knife. The semi-thin sections
were stained with Epoxy tissue stain, a special epoxy
stain.
The sections for electron microscopy were contrasted
in the first stage with uranyl acetate solution, followed by
contrasting with lead citrate solution, technique which is
currently used in electron microscopy laboratories,
around the world (Cachiţă şi Crăciun, 1991; Hayat,
2000).
The semi-thin sections were examined by us using an
Olympus BX 51 light microscope equipped with a CCD
camera and the ultrathin sections were examined using
a JEM-JEOL TEM 1010 electron microscope. Selected
pictures were recorded using a MEGA WIEW III
camera.
The experiment described in this article was aimed to
highlight the resemblances and differences between the
leaves and cotyledons of black locust because these
organs (especially when young) can be a source of
explants in plant biotechnology.
RESULTS AND DISCUSSION: In the physiology of the plant seed and embryo
(Mayer and Poljakoff–Mayber, 1975), the cotyledons
have an important role, mainly regarding the supply of
the tissues and cells with water and nutrients.
Altohught the active life span of the cotyledons is
short, the metabolic processes at cellular level play an
essential role in supplying the embryo with water and
also energy that is produced following the metabolising
of nutrients from the storage tissues. For the embryo
this energy is necessary for the growing processes and
cellular proliferation and also for generating the force
necessary to break the seed coat and to sustain the
growth of the plantlet hipocotyl and epycotyl and their
raising above the ground.
As it results from figure 4A, the foliole of a young
black locust leaf is planary with a main vein (midvein)
along the central area.
Burescu L., Cachiţă D., Crăciun C.
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71
© 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
68
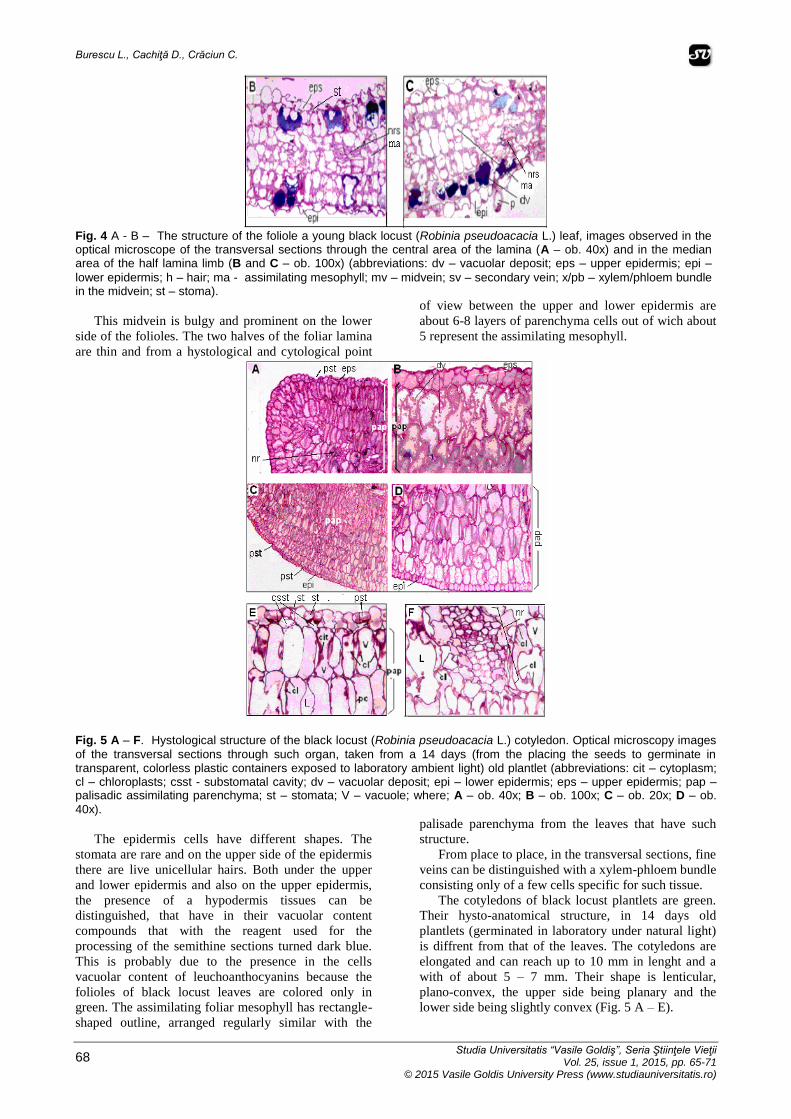
Fig. 4 A - B – The structure of the foliole a young black locust (Robinia pseudoacacia L.) leaf, images observed in the optical microscope of the transversal sections through the central area of the lamina (A – ob. 40x) and in the median area of the half lamina limb (B and C – ob. 100x) (abbreviations: dv – vacuolar deposit; eps – upper epidermis; epi –
lower epidermis; h – hair; ma - assimilating mesophyll; mv – midvein; sv – secondary vein; x/pb – xylem/phloem bundle in the midvein; st – stoma).
This midvein is bulgy and prominent on the lower
side of the folioles. The two halves of the foliar lamina
are thin and from a hystological and cytological point
of view between the upper and lower epidermis are
about 6-8 layers of parenchyma cells out of wich about
5 represent the assimilating mesophyll.
Fig. 5 A – F. Hystological structure of the black locust (Robinia pseudoacacia L.) cotyledon. Optical microscopy images
of the transversal sections through such organ, taken from a 14 days (from the placing the seeds to germinate in transparent, colorless plastic containers exposed to laboratory ambient light) old plantlet (abbreviations: cit – cytoplasm; cl – chloroplasts; csst - substomatal cavity; dv – vacuolar deposit; epi – lower epidermis; eps – upper epidermis; pap – palisadic assimilating parenchyma; st – stomata; V – vacuole; where; A – ob. 40x; B – ob. 100x; C – ob. 20x; D – ob.
40x).
The epidermis cells have different shapes. The
stomata are rare and on the upper side of the epidermis
there are live unicellular hairs. Both under the upper
and lower epidermis and also on the upper epidermis,
the presence of a hypodermis tissues can be
distinguished, that have in their vacuolar content
compounds that with the reagent used for the
processing of the semithine sections turned dark blue.
This is probably due to the presence in the cells
vacuolar content of leuchoanthocyanins because the
folioles of black locust leaves are colored only in
green. The assimilating foliar mesophyll has rectangle-
shaped outline, arranged regularly similar with the
palisade parenchyma from the leaves that have such
structure.
From place to place, in the transversal sections, fine
veins can be distinguished with a xylem-phloem bundle
consisting only of a few cells specific for such tissue.
The cotyledons of black locust plantlets are green.
Their hysto-anatomical structure, in 14 days old
plantlets (germinated in laboratory under natural light)
is diffrent from that of the leaves. The cotyledons are
elongated and can reach up to 10 mm in lenght and a
with of about 5 – 7 mm. Their shape is lenticular,
plano-convex, the upper side being planary and the
lower side being slightly convex (Fig. 5 A – E).

Anatomical, morphological and cytological comparative study of leaves and cotyledons from forestry species II. Comparison between the the morpho-anatomical and cytological structures
of cotyledons and leaves of black locust (Robinia pseudoacacia l)
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71 © 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
69
It is interesting that in the cotyledons, after the
processing of the sections, there are no more cells with
vacuoles having their content colored in blue and the
epidermis cells (upper and lower epidermis) are very
ordered and lack hairs. The epidermises have many
stomata that have a substomatal cavity or, some
stomata are not fully developed and were considered
pre-stomata.
From the hystoanatomical conformation of black
locust cotyledons, the hypodermis is missing; instead
under the epidermis, all around, is a palisade
assimilation parenchyma very well organized made of
3 – 4 layers of elongated successive parenchyma cells
(Fig. 5 A – E)
That have small chloroplasts and big vacuoles.
The cotyledonary mesophyll has approximately 14
– 16 layers of well ordered assimilating parenchyma,
the first 3 – 4 layers from the immediate vicinity of the
epidermises being of palisate type. The gaps and
intercellular spaces are not very large (Fig. 5 E – F).
From place to place, in the foliar mesophyll, small
xylem-phloem veins can be distiguished, some of them
being placed towards the cotyledonary apex (Fig. 5 A) .
Regarding the ultrastructural aspects that can be
distinguished in the lamina of the folioles (Fig. 6 A –
F), it can be stated that during the processing of the
ultrathine preparations, for examination, the vacuolar
content of many cells have suffered a corpusculation
process, in the vacuoles appearing spherical formations
of different sizes that are electron-dense and confirm
the presence in the vacuole of either anthocyanins or
phospholipids. These formations were named vacuolar
deposits. In the cells cytoplasm, normally structured
chloroplasts can be seen (Fig. 6 A – D) and also some
large nuclei in which a very well represented nucleolus
can be observed (Fig. 6 F).
Fig. 6 A –F. Cytological aspects observed in the epidermis and foliar mesophyll of young black locust (Robinia pseudoacacia L.) from the ultrathin transversal sections examined with the transmission electron microscope (A and B – upper epidermis and the first two mesophyll cells layers; C – F – lower epidermis and the neighboring mesophyll cells layers; D – F – details of the aspects preserved in the central foliar mesophyll) (abbreviations: cl – chloroplasts; cit –
cytoplasm; dv – vacuolar deposits; eps – upper epidermis; epi – lower epidermis; h – hair; cw- cellular wall; N- nucleus; n – nucleolus; V – vacuole).
The epidermis of the folioles, on the lower side has
short unicellular live hairs whereas is lacking hairs.
Also, the epidermis of the folioles has a large number
of well structured stomata whereas the one of the
cotyledons has some undifferentiated stomata (pre-
stomata). Besides, depending on the environmental
conditions after about 3 – 4 weeks from the onset of the
germination, the cotyledons start to turn yellow and
finally fall whereas the leaves folioles remain active
until autumn.
Burescu L., Cachiţă D., Crăciun C.
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71
© 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
70
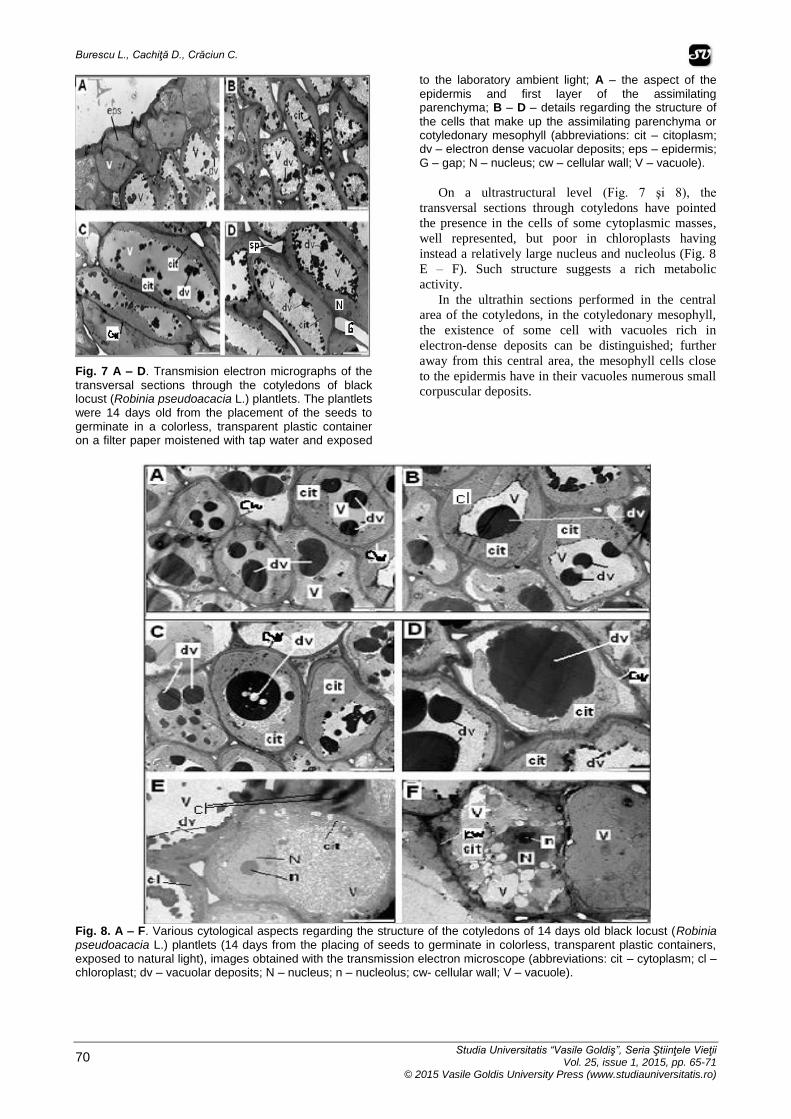
Fig. 7 A – D. Transmision electron micrographs of the
transversal sections through the cotyledons of black locust (Robinia pseudoacacia L.) plantlets. The plantlets were 14 days old from the placement of the seeds to germinate in a colorless, transparent plastic container on a filter paper moistened with tap water and exposed
to the laboratory ambient light; A – the aspect of the
epidermis and first layer of the assimilating parenchyma; B – D – details regarding the structure of
the cells that make up the assimilating parenchyma or cotyledonary mesophyll (abbreviations: cit – citoplasm; dv – electron dense vacuolar deposits; eps – epidermis; G – gap; N – nucleus; cw – cellular wall; V – vacuole).
On a ultrastructural level (Fig. 7 şi 8), the
transversal sections through cotyledons have pointed
the presence in the cells of some cytoplasmic masses,
well represented, but poor in chloroplasts having
instead a relatively large nucleus and nucleolus (Fig. 8
E – F). Such structure suggests a rich metabolic
activity.
In the ultrathin sections performed in the central
area of the cotyledons, in the cotyledonary mesophyll,
the existence of some cell with vacuoles rich in
electron-dense deposits can be distinguished; further
away from this central area, the mesophyll cells close
to the epidermis have in their vacuoles numerous small
corpuscular deposits.
Fig. 8. A – F. Various cytological aspects regarding the structure of the cotyledons of 14 days old black locust (Robinia pseudoacacia L.) plantlets (14 days from the placing of seeds to germinate in colorless, transparent plastic containers, exposed to natural light), images obtained with the transmission electron microscope (abbreviations: cit – cytoplasm; cl – chloroplast; dv – vacuolar deposits; N – nucleus; n – nucleolus; cw- cellular wall; V – vacuole).

Anatomical, morphological and cytological comparative study of leaves and cotyledons from forestry species II. Comparison between the the morpho-anatomical and cytological structures
of cotyledons and leaves of black locust (Robinia pseudoacacia l)
Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii Vol. 25, issue 1, 2015, pp. 65-71 © 2015 Vasile Goldis University Press (www.studiauniversitatis.ro)
71
ACKNOWLEDGEMENTS: This work was supported by the strategic
grant POSDRU/CPP107/DMI1.5/S/80272, Project
“Doctoral and Post-doctoral programs of excellence
for highly qualified human resources training for
research in the field of Life sciences, Environment and
Earth Science” cofinanced by the European Social
Found within the Sectorial Operational Program
Human Resources Development 2007 –
2013. University from Oradea.
CONCLUSIONS:
The folioles of the black locust (Robinia
pseudoacacia L.) leaves are different from the plantlets
cotyledons. This difference comes from both an
morphoanatomical and physiological point of view as
well as ultrastructural.
Because the black locust is a dicot species with
epigeal cotyledons, as the cotyledons raise above the
ground, they turn green and apparently for a short
period of time (about 40 days from placing the seeds to
germinate) they photosynthesize. As the cells of the
cotyledonary mesophyll exhaust their stored reserve
substances, they move from a heterotophic to a
autotrophic nutrition and the energy resulted from the
catabolic metabolic processes serves for the growth of
the embryo.
The folioles of the black locust leaves are odd
pennate have an elipsoidal shape are flat, thin and
about 1.5 – 2.5 cm in lenght. Centrally, across them
thay have a prominent midvein on the back of the
folioles.
The cotyledons of the black locust embryos are
about 10 cm in lenght, plano-convex with a planar
upper side and a convex lower side.
From an anatomical point of view, the folioles –
between the upper and lower epidermis – have a foliar
mesophyll composed of 4 – 5 layers of assimilating
parenchyma with a palisade type structure. The
cotyledonary mesophyll has 16 – 26 layers of cells. In
cotyledons, the cells in the immediate vicinity of the
epidermises have a classical palisade tissue structure,
highly ordered with small intercellular spaces.
REFERENCES: Cachiţă C.D., Crăciun C., Ultrastructural studies on
some ornamentals, in: Ammirato P.V., Evans
D.R., Sharp W.R., Bajaj Y.P.S., 1990,
Handbook af Plant Cell Culture, vol. 5,
Ornamental Species, Ed. Mc. Grow-Hill Publ.
Company, New-York, 1990, pp. 57-94.
Cachiţă C.D., Deliu C., Tican R.L., Ardelean A., Tratat
de biotehnologie vegetală, vol. I. Editura Dacia,
Cluj-Napoca, 2004.
Danciu, M., Parascan, D., Botanică forestieră, Ed. II-a
Ed „Pentru viaţă”. Braşov, 2002,. Enescu, V.,
Ioniţă, L., Palada, N,M,. Înmulţirea vegetativă a
arborilor forestieri. Metode „convenţionale”.
Culturi de ţesuturi in vitro. Ed. Ceres,
Bucureşti, 1994.
Hayat, M.A., Principles and technique for electron
microscopy. Biological Application. Cambridge
University Press , 2000.
Mayer A.M., Poljakoff-Mayber A., The germination of
seeds (Second Edition), în: International Series
of Monographs Pure and Applied Biology,
Division: Plant Physiology, General Ed.
Wareing P.F. and Galston A.Y., Perganon
Press-Oxford New-York Toronto, Sydney, Paris
Braunschweig, 1975.
Stănescu, V., Dendrologie, Ed. Did. şi Ped. Bucureşti,
1979.
Toma, C., Anatomia plantelor II Structura organelor
vegetale şi de reproducere. Univ. „Al. Cuza”
Iaşi, Fac. de Biologie-Geografie, 1977.
Toma C., Gostin I., Histologie vegetală, Ed. Junimea,
Iaşi, 2000.
Turnbull, C.G.N., Booker, J.P., Ottoline, V., Leyser,
H.M., Micrografting techniques for testing long-
distance signalling in Arabidopsis. Plant J., nr.
32, pp. 255-262, 2002.