microbiologie generala b ii
TRANSCRIPT

1
MICROBIOLOGIE GENERALÃ � NOTE DE CURS
1. CONCEPTUL DE BACTERIE
Primele bacterii au fost evidenþiate ºi descrise de cãtre A. Leeuwenhoek în 1675. Recunoaºterea lor ca grup distinct a fost fãcutã în perioada 1850-1875 de cãtre Cohn,
pe baza proprietãþilor grupului omogen al eubacteriilor. Stanier ºi Lwoff (1973) considerã cã F. Cohn este adevãratul fondator al
bacteriologiei ca ramurã a biologiei. Descoperirea diferitelor grupuri de bacterii (actinomicete, spirochete, mixobacterii
etc.) a scos în evidenþã multe caractere care nu sunt unitare, de exemplu: au fost derscrise bacterii imobile sau mobile prin unul sau mai mulþi flageli,
prin alunecare, târâre sau prin contracþia unui folament axial; s-a evidenþiat cã multiplicarea se poate face prin diviziune, înmugurire sau
formare de celule specializate (spori), sau prin segmentare ºi fragmentare (la
actinomicete); unele bacterii (actinomicetele) prezintã, datoritã structurii lor miceliene,
analogii frapante cu grupul fungilor în care au fost încadrate multã vreme; mixobacteriile prezintã un ciclu de dezvoltare complex, care include un stadiu
de creºtere vegetativã ºi unul de fructificaþii policelulare, adesea cu dimensiuni
macroscopice � ca protistele amoeboide.
Conceptul de bacterie a fost definit în perioada 1960-1964, oferind posibilitatea de a grupa pe criterii obiectiv-ºtiinþifice, discriminatorii, microorganismele reunite pânã atunci
numai pe bazã de acord pur convenþional. Stanier (1962, 1964, 1967) a demonstrat primul cã diversificarea prin evoluþie a
lumii bacteriene are la bazã un caracter comun � celula procariotã. Conceptul de bacterie, în forma sa actualã poate fi definit în funcþie de aceastã
organizare procariotã ºi prin antitezã cu tipul de organizare eucariot.
Celua procariotã, mai puþin complexã, este unitatea de structurã a bacteriilor sensu lato ºi a �algelor� albastre-verzi.
Celula eucariotã, mai complexã, este unitatea de structurã a tuturor celulelor animale, a
plantelor vasculare, a briofitelor, a fungilor ºi a tuturor celorlalte grupuri de alge.
Divergenþa fundamentalã care separã bacteriile ºi �algele� albastre-verzi de toate celelalte organisme, corespunzând diferenþei dintre tipul procariot ºi eucariot, reprezintã cea
mai mare discontinuitate evolutivã prezentã azi în lumea vie. Nu se cunosc intermediari între procariote ºi eucariote, adicã nu se cunosc organisme
cu o structurã celularã de tranziþie. Dodge (1965, 1971), pornind de la observaþia cã dinoflagelatele au unele
caracteristici ale procariotelor ºi altele aparþinând eucariotelor propune includerea lor într-un grup intermediar Mesokariota.
Cercetãrile de microscopie electronicã ºi cele de biologie molecularã au dezvãluit ºi
accentuat discontinuitatea dintre procariote ºi eucariote ºi au dus la acumularea unui numãr
important de caractere discriminatorii, care permit sã se stabileascã ceea ce este realmente
tipic ºi ceea ce este numai aparent ºi superficial similar, pentru încadrarea pe baze ºtiinþifice a
diferitelor entitãþi într-un grup sau altul.
id2762156 pdfMachine by Broadgun Software - a great PDF writer! - a great PDF creator! - http://www.pdfmachine.com http://www.broadgun.com

2
Poziþia microorganismelor în lumea vie
Diferenþele fundamentale dintre bacterii ºi celula vegetalã (ca prototip de celulã
eucariotã) demonstreazã absenþa oricãrei punþi de legãturã între lumea bacterianã ºi cea a
plantelor, de aceea încadrarea bacteriilor în rândul plantelor este arbitrarã ºi nelogicã (Stanier
ºi Van Niel, 1962, 1977). Menþinerea acestei încadrãri a bacteriilor în grupa plantelor de-a lungul anilor a reprezentat rezultatul �refuzului îndãrãtnic de a privi lucrurile în faþã�, singurul
argument care poate fi invocat fiind acela al aprecierii preºtiinþifice, dupã care în lumea vie ar
exista doar douã categorii de organisme � mutual exclusive � plantele ºi animalele. Pe mãsurã ce lumea microscopicã a fost cunoscutã mai aprofundat a devenit evident cã
unele microorganisme nu pot fi clasificate � fãrã rezerve � în nici unul din cele douã regnuri
tradiþionale ºi cã existã grupuri întregi � cu caractere intermediare � revendicate în egalã
mãsurã ºi de botaniºti ºi de zoologi (de ex., Euglena, Chlamydomonas, Volvox etc.). Incnvenientele majore ale sistemului tradiþional de clasificare au fost sesizate iniþial
de Hogg (1860) care a propus gruparea microorganismelor într-un regn nou, regnul Primigenum, denumit ulterior Protoctista.
Haeckel (1866) propune crearea unui regn menit sã grupeze toate formele inferioare
de viaþã, regnul Protista, definit ºi delimitat în mai multe variante, pentru ca în formele sale
finale (1894-1904) sã includã numai microorganismele unicelulare sau unicelular-coloniale. Stanier (1970) împarte regnul Protista în douã subgrupuri:
protiste inferioare, corespunzând microorganismelor procariote (bacterii ºi
alge albastre-verzi); protiste superioare, corespunzând microorganismelor eucariote, care includ
algele, fungii ºi protozoarele. În aceastã accepþiune, cadrul conceptului de Protista este exins la toate organismele
unicelulare, cenocitice sau multicelulare care nu formeazã þesuturi. Sistemul celor patru regnuri propus de Copeland (1938) ºi modificat de Hutchinson
(1967) ºi Weisz (1971) are ca trãsãturã esenþialã diviziunea lumii microorganismelor în patru
regnuri: 1. Regnul Monera (Mychota), care include bacteriile propriu-zise ºi �algele�
albastre-verzi; 2. Regnul Protoctista (Protista), incluzând organismele eucariote inferioare cu
organizaþie unicelularã, unicelular-colonialã, sinciþialã sau multicelularã, fãrã diferenþieire
celularã avansatã (de ex., algele cu excepþia celor douã procariote, fungii, mixomicetele ºi
protozoarele). 3. Regnul Plantae (Metaphyta, Weisz) - corespunzând plantelor terestre ºi
acvatice � organisme eucariote superioare, multicelulare, cu perete celular ºi diferenþiere
celularã variabilã de la limitatã la intermediarã. 4. Regnul Animalia (Metazoa, Weisz) - incluzând organisme multicelulare
eucariote, conþinând celule fãrã perete celular ºi fãrã plastide, având ºi prezentând o
diferenþiere avansatã, tisularã ºi de organe. Deºi are o serie de avantaje mari faþã de sistemele anterioare ºi reprezintã o
contribuþie majorã la interpretearea lumii vii, sistemul celor patru regnuri, în diferitele sale variante, prezintã o serie de imperfecþiuni, care, deºi sunt proprii lumii vii ca obiect de
clasificare, apar cu evidenþã maximã, legate tot de lumea organismelor simple. Între aceste imperfecþiuni sunt menþionate urmãtoarele:
Regnul Protoctista (Protista), foarte diferit în raport cu grupul Monera (Meychota), este lipsit de unitate ºi de claritate, din cauza faptului cã limitele
de organizare ale organismelor sunt foarte largi, iar diferenþierea lui de
organismele superioare nu este clar trasatã. Aceastã situaþie se datoreazã

3
faptului cã nu existã nici o cale bunã pentru a separa organismele eucariote
inferioare de cele superioare, ci numai diferite alternative de opþiune, cu
dificultãþi ºi incnveniente diferite. Luând drept criteriu nivelul de diferenþiere
tisularã, apartenenþa algelor ºi a fungilor superiori la regnul Protista este o consecinþã imediatã, care atrage dupã sine o a doua, eterogenitatea lui. Linia de
demarcaþie între Protista, pe de o parte, ºi plantele ºi animalele superioare, pe
de alta, este trasatã, în primul rând de gradul de diferenþiere tisularã situaþie
care creeazã o delimitare destul de neclarã. În fapt, regnul Protoctista reprezintã mai curând o confederaþie de organisme excluse din celelalte trei
regnuri (Monera, Plantae, Animalia), care cuprinde o foarte largã gamã de
organisme cu diferite niveluri intermediare de organizare, superioare în raport
cu procariotele, dar inferioare plantelor vasculare ºi animalelor superioare. Sistemul celor cinci regnuri, propus de Whittacher (1969), încearcã sã ocoleascã
deficienþele sistemelor precedente, utilizând urmãtoarele criterii de grupare: 1. Stabilirea a trei niveluri de organizare: a) procariot; b) eucariot unicelulur;
c) eucariot � multicelular; 2. Existenþa a trei modalitãþi principale de nutriþie, determinante a trei direcþii majore
de evoluþie, care la nivelul multicelular (multinuclear) se exprimã prin divergenþele evolutive
ale celor tri regnuri superioare: a. nutriþia fotosinteticã (secundar absorbtivã) � caracteristicã plantelor; b. nutriþia ingestivã � tipicã pentru animale; c. nutriþia absorbtivã � caracteristicã fungilor.
Consecinþele adoptãrii acestor criterii sunt în esenþã � pe lângã unele restructurãri ale
grupelor mai mici � urmãtoarele: 1. apariþia regnului Fungi ca al treilea regn de organisme superioare,
alãturi de plante ºi animale; 2. trecerea algelor verzi în regnul Plantae, alãturi de plantele verzi
superioare; 3. plasarea liniei de demarcalie dintre organismele superioare ºi protiste la
trecerea de la unicelular la multicelular � multinuclear. În acord cu aceste principii, sistemul celor cinci regnuri elaborat de Whittaker are
urmãtoarea structurã: 1. Regnul Monera include organisme unicelulare de tip procariot, corespunzând
bacteriilor, organisme cu existenþã unicelular-solitarã sau unicelular-colonialã, cu excepþia
actinomicetelor care au organizare de tip micelial. Modul de nutriþie este predominant
absorbtiv, iar metabolismul de tip foto- sau chimiosintetic. Reproducerea se face prin diviziune asexuatã, mai rar prin înmugurire. Rar, prezintã fenomene de protosexualitate.
Imobile sau mobile (prin flageli simpli, de tip special, sau prin alunecare-târâre). 2. Regnul Protista include organisme primar unicelulare sau unicelular-coloniale
de tip eucariot. În unele cazuri sunt prezente stadii de viaþã cu structurã simplã sau
multinuclearã. Modul de nutriþie este diferit de la un grup la altul (absorbþie, ingestie,
fotosintezã sau combinaþii ale acestor tipuri). Reproducerea este, de asemenea, de tip diferit,
incluzând diviziunea asexuatã (la nivel haploid) ºi procese sexuale adevãrate, cu cariogamie ºi
meiozã. Sunt imobile sau mobile prin flagel de tip superior sau prin alte mijloace. 3. Regnul Plantae include organisme multicelulare cu celule eucariote, care au
peretele celular celulozic, frecvente vacuole în citoplasmã ºi pigmenþi fotosintetici în plastide.
Modul principal de nutriþie este cel fotosinteic; unele linii au devenit absorbtiive. Esenþial
imobile, trãiesc ancorate pe un substrat. Prezintã diferenþiere structuralã care duce la apariþia
unor organe de fotosintezã, ancorare ºi de sprijin (suport), iar la formele superioare la
formarea unor þesuturi specializate fotosintetice, vasculare ºi de acoperire. Reproducerea este

4
în primul rând sexuatã, cu cicluri de generaºþii alternative, haploide ºi diploide, primele fiind progresiv reduse spre membrii superiori regnului.
4. Regnul Fungi. Exceptând subregnul Gymnomycota, include organisme multinucleare, cu nuclei de tip eucariot, dispersaþi într-un sinciþiu micelial, acoperit de un
perete celular, adesea septet, caracterizate prin lipsa plastidelor ºi a pigmenþilor fotosintetici.
Nutriþia este de tip absorbtiv. Diferenþierea somaticã este absentã sau limitatã; în schimb,
diferenþierea þesutului reproductiv ºi complicare ciclului de viaþã sunt foarte marcate la formele superioare. În primul rând imobile (dar cu flux citoplasmatic în miceliu), trãiesc
înclavate în mediu sau în sursa de hranã. Prezintã cicluri reproductive tipice, incluzând
procese sexuale ºi asexuale. Micelii, cel mai des haploide la formele inferioare ºi dicariotice
la multe forme superioare. 5. Regnul Animalia include organisme multicelulare de tip eucariot, fãrã perete
celular, fãrã plastide ºi fãrã pigmenþi fotosintetizanþi. Nutriþia este în primul rând de tip
ingestiv, cu digestie într-o cavitate internã; unele forme sunt absorbtive; la unele grupuri
lipseºte cavitatea digestivã internã. La formele superioare nivelul de organizare ºi de
diferenþiere tisularã depãºeºte de departe celelalte regnuri, ducând la apariþia a diferite sisteme
senzoriale neuromotorii. Motilitatea organismelor sau a pãrþilor lor (la formele sesile) se
bazeazã pe existenþa unor fibrile contractile. Reproducerea predominant sexuatã; stadiile
haploide, diferite de cele ale gameþilor, aproape lipsesc la organismele situate deasupra filumurilor inferioare.
Sistemul lui Whittaker, deºi foarte recent creat, este agreat de un mare numãr de
specialiºti, care considerã cã el reprezintã cel mai bine relaþiile dintre organisme, atât în ceea
ce priveºte nivelurile de organizare ale lumii vii, cât ºi modul de nutriþie care afecteazã tipul
de organizare. El are avantaje evidente în sensul unei mai pregnante coerenþe a regnurilor, ca
unitãþi de clasificare decurgând din precizarea mai netã a caracterelor lor definitorii, dar ºi o
serie de limitãri care sunt evidenþiate chiar de Whittaker, dar care nu sunt legate de lumea
bacteriilor.

5
2. ANATOMIA BACTERIILOR
2.1. Morfologie
Forma bacteriilor este controlatã genetic. Deºi variazã destul de mult, în funcþie de
condiþiile de mediu, polimorfismul acestor microorganisme este relativ limitat ºi caracterizat,
de cele mai multe ori, prin predominanþa formei tipice pentru specia datã. Deoarece forma bacteriilor este greu de apreciat direct din mediile naturale,
cunoºtinþele de morfologie bacterianã se referã, de obicei, la celulele cultivate, în condiþii
artificiale de laborator, în care caz forma celulei poate fi influenþatã de vârsta culturii, de
compoziþia mediului, de temperaturã etc. Datoritã acestui fapt, forma bacteriilor � care reprezintã un criteriu taxonomic important � se apreciazã în mod convenþional pe celule
provenite din culturi tinere, în faza de creºtere activã, pe medii de culturã corespunzãtoare ºi
în condiþii optime de temperaturã, tensiunea O2, pH etc. În culturi vechi, în care cele mai
multe bacterii sunt pe cale sã degenereze sau sã moarã, apar celule cu forme aberante (cu
aspecte de Y, ramificate, filamentoase etc.), care se observã ºi în cazul cultivãrii în condiþii
improprii de mediu (temperaturã nepotrivitã, concentraþii mari de sãruri anorganice, doze
subletale de substanþe antibacteriene); acestea rezultã din interferenþa unor acþiuni nocive cu
procesul de diviziune normalã sau din altererarea mecanismelor de permeabilitate selectivã,
urmatã de imbibiþia cu apã ºi autoliza structurilor celulare prin acþiunea enzimelor proprii. Dupã forma celulei, bacteriile pot fi grupate în cinci mari categorii:
1. sferice; 2. cilindrice; 3. spiralate sau elicoidale; 4. filamentoase; 5. pãtrate. La cela mai multe specii, celulele-fiice se separã ºi rãmân independente datoritã
agitãrii mecanice, curenþilor de convecþie din mediu, miºcãrii browniene, forþelor rezultate din
presiunea de turgor a celulelor care cresc, cât ºi a acþiunii flagelilor, în cazul bacteriilor mobile.
La unele specii aparþinând grupului de bacterii sferice ºi uneori chiar al celor
cilindrice, celulele-fiice nu se despart la sfârºitul diviziunii, ci formeazã grupãri caracteristice
cu valoare texonomicã. Rareori însã toate celulele speciei respective sunt grupate tipic. Caracteristic este modul de grupare dominant. Modul de grupare a celulelor dupã diviziune
este în funcþie de raportul geometric dintre diferite planuri succesive de diviziune ºi de
tendinþa celulelor-fiice de a rãmâne unite. În unele cazuri, gruparea nu reprezintã un caracter
morfologic propriu, ci este o consecinþã a stadiului de creºtere sau a condiþiilor de culturã.
Spre exemplu, Corynebacterium diphteriae are tendinþa sã se grupeze în palisadã, iar
Mycobacterium tuberculosis în grupuri de trei bacili, care dau impresia de structurã ramificatã. Aceste moduri de grupare reprezintã însã excepþii de la regulã, deoarece cea mai
mare parte dintre bacteriile respective apar izolate unele de altele.

6
1. Bacteriile sferice (cocii) au formã sfericã, ovoidalã, elipsoidalã, reniformã, uneori
neregulatã, sau parþial poliedricã, diametrele celulei fiind aproximativ egale. Deoarece aceastã
formã este adesea modificatã de diferiþi factori de mediu este discutabil dacã în stare vie
celula este realmente sfericã. În funcþie de poziþia celulelor-fiice dupã diviziune, cocii prezintã urmãtoarele ºase subtipuri
morfologice: a) cocul simplu sau izolat, la care celulele rezultate din diviziune rãmân
independente; b) diplococul, la care diviziunea se face dupã planuri succesive paralele, celulele
rezultate rãmânând grupate câte douã (Streptococcus pneumoniae); c) streptococul la care diviziunea se face dupã planuri succesive paralele, dar
celulele rezultate formeazã lanþuri de lungimi variabile, ca un ºirag de mãrgele (Streptococcus pyrogenes);
d) tetracocul sau tetrada, la care planurile succesive de diviziune sunt perpendiculare unele faþã de altele, iar celulele rezultate sunt dispuse în grãmezi de patru
elemente (Micrococcus tetragenes); e) sarcina, la care planurile de diviziune sunt orientate în trei direcþii diferite,
perpendiculare unul pe alrul (al doilea pe primul, ºi al treilea pe primele douã), de unde
rezultã o grupare în cuburi sau pachete (Sarcina flava, S. aurantiaca); f) stafilococul la care planurile succesive de diviziune sunt dispuse în câteva
direcþii, iar organismele rezultate se aranjeazã în grupãri neregulate în formã de ciorchine
(Staphylococcus aureus).
Lampropedia are forma unor celule rotunde sau aproape cubice, grupate în ºiruri
regulate, formând �tablete� rectangulare de 16, 32 sau 64 de celule agregate, datoritã unei
matrice extracelulare. Celulele care formeazã �tablete�, având în interior incluziuni
refringente strãlucitoare, sunt acoperite de un înveliº structurat complex, distinct de peretele
celular, care nu înconjurã celulele individuale, ci numai ansamblul grupãrii lor. Izolatã din
rumen, ape mâloase, stagnante, Lampropedia formeazã pelicule pe suprafaþa mediilor lichide. 2. Bacteriile cilindrice cunoscute mai ales sub denumirea comunã de bacili, au formã de
bastonaºe. Raportul dintre cele douã axe variazã mult, unele bacterii putând lua formã aproape
sferic-ovalã (cocobacili); acestea din urmã fiind greu diferenþiate de coci, dar în culturi pure
existã mereu câteva celule suficient de lungi, pentru a identifica natura lor cilindricã. Bacilii sunt drepþi sau uºor încurbaþi la mijloc sau la una din extremitãþi. Capetele pot fi tãiate drept (Bacillus antracis) sau rotunjite ca la majoritatea celorlalþi. Marginile laterale ale celulei sunt de obicei paralele, dar pot fi ºi apropiate la
extremitãþi, în firmã de suveicã (Fusiformis fusiformis), sau îndepãrtate ºi rotunjite la una sau
ambele extremitãþi, în formã de mãciucã sau de piºcot (Corynebacterium). Bacilii pot fi grupaþi:
câte doi (diplobacili); în lanþuri cu lungimi variabile (streptobacili); în palisadã � ca scândurile unui gard, celulele rãmânând apropiate ºi paralele în
sensul axului lung, aºezarea fiind rezultatul unei miºcãri de basculare a celulei-fiice, având ca punct de sprijin peretele transvers recent separat;
sub formã de rozetã sau de stea (Agrobacterium stellulatum, Ag. radiobacter, Phyllobacterium stappi).

7
3. Bacteriile spiralate sau elicoidale cuprind trei subtipuri morfologice: a) vibrionul, în formã de virgulã (Vibrio cholerae); b) spirilul, în formã de spiralã cu mai multe ture de spiralã rigide (Spirillum
volutans); c) spirocheta, în formã de spiralã cu mai multe ture flexibile, care se pot strânge sau
relaxa (Borrelia, Treponema ºi Leptospira). 4. Bacteriile filamentoase au ca prototip actinomicetele, microorganisme cu asemãnãri morfologice asemãnãtoare cu fungii, având particularitatea de a forma hife ramificate (de
unde aspectul de miceliu). În unele cazuri aspectul filamentos este determinat de aºezarea
celulelor individuale în lanþuri de celule reunite printr-o teacã delicatã cu perete neted, care în
condiþii favorabile pot acumula hidroxid de fier sau mangan.
5. Bacteriile pãtrate evidenþiate în apa hipersalinã din unele bãlþi din Peninsula Sinai
(Walsby, 1980) au forma unor pãtrate cu latura de 1,5-11 ìm ºi o grosime inegalã (0,1 ìm sau chiar mai micã în regiunea centralã ºi 0,2-0,5 ìm la periferie, Parkes, 1981). Uneori formeazã placarde de 8-16 pãtrate la care se vãd net planurile de diviziune. Celulele conþin vacuole cu gaze, care dispar la presiune, situate frecvent de-a lungul marginilor pãtratelor. În unele cazuri sunt dispersate la întâmplare în celulã. Multiplicarea se face prin diviziune. În unele cazuri, fiecare bacterie pãtratã creºte pânã ia
forma unui dreptunghi, care se divide în douã pãtrate egale. Alteori, diviziunea se face în douã
planuri ce alterneazã în unghi drept iar celulele rezultate din diviziune formeazã placarde. Bacteriile pãtrate au fost încadrate în genul Quadra aparþinând probabil grupului Archaebacteria (Parkens ºi Walsbz,1981).
În afara acestor cinci tipuri morfologice de bazã existã bacterii cu forme particulare,
rezultate din gruparea lor într-un trichom, prezenþa unor prelungiri celulare (prostecã), a unor
aprndice acelulare etc.
Bacteriile care formeazã trichoame. Trichomul este o grupare de celule sub forma unui filament multicelular, uniseriat, rezultat din diviziune, în care celulele adiacente au o suprafaþã
relativ mare de contact strâns ºi sunt menþinute într-un înveliº parietal comun (Starr,1973) (Leucothrix, Beggiatoa, Caryophanon latum, Sphaerotilus natans).
Bacteriile prostecate. Unele bacterii prezintã o complicaþie morfologicã � prosteca (gr. = adaos, apendice, �coadã�) sub forma unui apendice semirigid, situat în continuarea unei celule procariote ºi având totdeauna un diametru mai mic decât cel al celulei mature. Funcþiile prostecii:
- implicatã în reproducerea bacteriilor care înmuguresc; - mãreºte suprafaþa celulei ºi implicit a membranei plasmatice, permiþând
sporirea activitãþilor asociate (respiraþia ºi absorbþia de sunbstanþe nutritive); - are rol de fixare (Caulobacter), deºi frecvent a fost gãsitã ºi la celule plutitoare.
Bacteriile cu apendice acelulare. Bacteriile prezintã un aspect filamentos datoritã unor
apendice acelulare, formate din substanþe secretate sau excretate de ele, nedelimitate de perete
celular. Gallionella ferruginea, prezentã în ape feruginoase, are forma unui bob de fasole de
0,5x1,2 ìm ºi este situatã la extremitatea unui apendice filamentos acelular, format dintr-un produs secretat de ea însãºi prin porii situaþi pe partea sa concavã.

8
Celulele bacteriene miniaturale (minicelulele). Minicelulele sunt corpusculi mici, aproximativ sferci, care nu cresc ºi sunt formaþi printr o septare neobiºnuitã, în apropierea uneia dintre extremitãþile unei bacterii cilindrice. Au cel mai adesea un diametru mai mic decât al unei celule bacilare parentale. Sunt lipsite de
material nuclear sau deficiente în ADN. De aceea sun incapabile de creºtere ºi diviziune. Au fost descrise ca un proces de diviziune inegalã (asimetricã) la Escherichia coli, Salmonella, Bacillus subtilis, Haemophilus influenzae, Erwinia amylovora º.a.
2.2. Proprietãþi fizice
Dimensiuni Bacteriile au dimensiuni foarte mici, în medie 0,5-1 ìm x 3-6 ìm. Cele mai mici
aparþin genului Mycoplasma ºi au diametrul 125-250 nm. Cele mai mari pot ajunge la 10-20ìm lungime, iar în cazul formelor filamentoase, în mod excepþional, chiar la 500 ìm (Saprospira grandis).
Sub raportul dimensiunilor cele mai mici bacterii se suprapun virusurilor mari (Poxvirus), vizibile la microscopul fotonic, iar cele mai mari depãºesc mãrimea celor mai mici protiste eucariote. Deoarece celulele diferã mult ca formã, stanier (1970) recomandã volumul ca unicã bazã satisfãcãtoare de apreciere a mãrimii bacteriilor. Volumul celulei bacteriene poate fi
apreciat cu ajutorul formulelor de calcul aplicate corpurilor geometrice regulate.
Suprafaþa celulei bacteriene se poate calcula încadrând bacteriile într-o formã geometricã regulatã, pe baza dimensiunilor celulei. Exemple:
1,104x10-9 cm2 (Mycoplasma), 35,340x10-9 cm2 (Escherichia coli), 12 576 x10-9 cm2 (Saprospira grandis).
Densitatea sau greutatea specificã a celulei bacteriene vii (în stare umedã) variazã
între 1,07 ºi 1,32. Valoarea acestui indice este în funcþie de proporþia relativã a substanþerlor
celulare cu densitate diferitã de cea a apei (D=1). Lipidele au densitatea sub 1,0, glucidele 1,4-1,6, proteinele1,5, acizii nucleici 2,0, sãrurile minerale 2,0-2,5.
Deoarece compoziþia chimicã a bacteriilor este variabilã chiar la aceeaºi specie, în funcþie de condiþiile de mediu sau de vârsta celulei, densitatea lor este caracteristicã numai în raport cu anumite condiþii de creºtere. Celulele tinere, care sunt turgescente, au o densitate mai micã decât cele bãtrâne, iar la bacteriile cultivate pe medii lichide valoarea ei este mai
micã decât la cele cultivate pe medii solidificate. Datoritã densitãþii lor mici, bacteriile rezistã
la sedimentare spontanã în medii gazoase ºi lichide, din care însã pot fi separate prin
centrifugre. Greutatea unei bacterii poate fi calculatã împãrþind greutatea unei anumite mase de celule
la numãrul unitãþilor componente. Celulele de Escherichia coli, care sunt de 500 de ori mai mici decât o celulã vegetalã sau animalã de dimensiuni medii (Ø 10 ìm), au o greutate de 10
12 dal, respectiv de 6 x 106 dal ori mai mare decât greutatea unei molecule de apã (g.m.=18 dal.)
Raportul dintre suprafaþa celulei ºi greutate are o valoare foarte ridicatã la bacterii. În cazul unui coc cu diametrul de 1 ìm acest raport este de ~ 55 000, la E. coli ~ 81 000, la Chlamydia ~ 193 000, iar la Mycoplasma ~ 290 000. Acelaºi raport la om (24000 cm2/70 000 g)

9
este de 0,342, respectiv de 84 146 ori mai mic decât la Bacillus megaterium, de 239 233 ori mai mic decât la Escherichia coli, ºi de 850 611 ori mai mic decât la Mycoplasma
Datoritã raportului foarte mare dintre suprafaþa ºi volumul bacteriilor, comparativ cu
alte organisme, asemãnãtor aceluia al particulelor de substanþe în state de dispersie coloidalã,
o cantitate micã de bacterii, de exemplu 1 g de Escherichia coli, care conþine 1,8 x 1012 celule
însumeazã o suprafaþã totalã de contact cu mediul de circa 56 000 cm2; pentru comparaþie
menþionãm cã 1 g celule de drojdie conþine 8,3 x 109 celule, care au o suprafaþã de 9 100 cm
2. Mãrimea considerabilã a acestei suprafeþe reactive de la interfaþa celularã � mediu este
deosebit de importantã, deoarece, practic toate substanþele care intrã ºi ies din celulã trec prin
aceastã zonã. Teoretic, pe baza unor considerente de ordin biologic ºi biochimic, limita inferioarã
de mãrime a unei celule bacteriene este determinatã de posibilitatea ei de a conþine toþi
constituenþii necesari pentru creºtere ºi diviziune, respectiv un numãr minim de enzime diferite (probabil câteva sute), aflate fiecare într-un numãr variabil de molecule, acizi nucleici,
glucide, lipide etc. Cea mai micã bacterie vizibilã la microscopul fotonic are dimensiuni foarte
apropiate de aceastã limitã molecularã necesarã pentru menþinerea funcþiilor celulare.
Celulele cu dimensiuni mai mici nu ar fi fizic suficient de cuprinzãtoare pentru a conþine
întreg echipamentul enzimatic ºi moleculele necesare pentru viaþã. În cazul microorganismelor eucariote, limita cea mai micã de mãrime este determinatã
de limitãri structurale: cele mai mici protiste eucariote sunt algele din genul Micromonas (M. pulsatilla 1-1,5 ìm) cu un singur cloroplast ºi o singurã mitocondrie, care alãturi de nucleu
ocupã ceaq mai mare parte din celulã. Orice reducere de dimensiuni dincolo de aceastã limitã
nu se poate face decât prin eliminarea unui organit celular, fãrã de care însã funcþiile
metabolice ar înceta de îndatã. Ca regulã generalã, celulele eucariote sunt însã mai mari decât
cele procariote ºi conþin structuri absente la procariote. Celulele procariote nu pot depãºi anumite limite de mãrime ºi, în general, nu pot
ajunge la dimensiunile foarte mari ale celulelor eucariote. Limita superioarã de mãrime a unui organism unicelular este posibil corelatã cu
raportul suprafaþã/volum, având în vedere cã toþi nutrienþii trebuie sã pãtrundã în celulã, iar
produºii de catabolism sã fie eliminaþi. Pe mãsurã ce dimensiunile unei celule cresc, volumul
ei creºte mult mai rapid decât suprafaþa (volumul creºte la cub în raport cu raza celulei, în
timp ce suprafaþa creºte cu raza la pãtrat). Când raza celulei creºte de trei ori, volumul creºte
de 27 de ori (33), iar suprafaþa de numai 9 ori (32). În prezenþa acestui dezechilibru, celula nu
poate îngloba nutrienþi ºi elimina substanþe reziduale în acord cu necesitãþile crescute ale
volumului celular mãrit (Nester ºi colab., 1973). Celulele eucariote mari au rezolvat aceastã problemã fie prin modificarea adecvatã a
formei (aducând astfel pãrþileinterne ale celulei aproape de suprafaþã), fie, mai ales, prin
prezenþa curenþilor citoplasmatici care asigurã �circulaþia� nutrienþilor ºi a produºilor de uzurã
dintr-o zonã în alta. Ca o regulã generalã, valabilã la toate nivelele de complexitate biologicã, rata
metabolismului este invers proporþionalã cu mãrimea organismului, iar rata de creºtere
(determinatã în special de rata globalã a metabolismului) creºte pe mãsurã ce dimensiunile
celulare scad în corelaþie cu raportul suprafaþã/volum. Dimensiunile mici ale bacteriilor apar ca o condiþie esenþialã pentru creºterea ºi
multiplicarea lor rapidã, în comparaþie cu celulele eucariote, ceea ce le conferã un mare
avantaj biologic în naturã, asigurând supravieþuirea lor în competiþie cu alte organisme.
Celulele mai mari au nevoie de mult timp pentru ca nutrienþii sã pãtrundã în celulã ºi sã fie
metabolizaþi pentru a asigura creºterea acesteia. Un alt factor care pare sã determine o limitã superioarã de mãrime a celulelor
bacteriene este dificultatea de menþinere a unei reglãri satisfãcãtoare ºi de coordonare a

10
activitãþii metabolice într-o celulã mare cu organizare de tip procariot. Unele bacterii foarte
mari, Spirillum volutans, Thiospirillum jenense etc., nu pot fi cultivate decât extrem de greu,
în contrast cu bacteriile mici din acelaºi grup fiziologic (Stanier, 1970). Explicaþia constã în
faptul cã la aceste bacterii condiþiile fizico-chimice necesare dezvoltãrii sunt foarte restrânse,
iar capacitatea de reglare a funcþiilor metabolice esenþiale pentru a le conferi flexibilitatea adaptativã la modificãri minore ale mediului foarte limitatã.

11
3. ULTRASTRUCTURA CELULEI BACTERIENE
Luând ca reper peretele celular, constituenþii celulei bacteriene pot fi grupaþi în douã
mari categorii: intraparietali, care formeazã protoplastul (membrana plasmaticã, mezosomii,
citoplasma, nucleoidul, ribosomii, magnetosomii); extraparietali (capsula, stratul mucos, glicocalixul, �spinii�, flagelii, fimbriile
ºi pilii).
3.1. Peretele celular
Celula bacterianã este delimitatã de un perete celular bine definit structural ºi cu
consistenþã rigidã. Situat în afara membranei citoplasmatice, el este acoperit, la unele specii, de o
capsulã sau de un strat mucos pericelular, iar la bacteriile mobile este strãbãtut de flageli. Datoritã indiceluli de refracþie mic, peretele celular este invizibil ori foarte greu
vizibil la celulele vii examinate la microscopul fotonic. Evidenþierea lui este însã posibilã prin
colorare selectivã sau prin examinare la microscopul electronic. La bacteriile tinere, care au citoplasmã abundentã, compactã ºi omogenã, peretele
celular aderã intim la conþinutul celular subiacent. La celulele bãtrâne sau în curs de lizã,
plasmolizã etc., peretele celular devine evident, fiind depãrtat de citoplasmã. Punerea în evidenþã a peretelui celular se poate face fie prin izolarea lui ca structurã
intactã completã, fie prin modificarea raporturilor lui spaþiale cu citoplasma. Aceastã
modificare se poatwe obþine pe cale dezintegrãrii celulare prin ultrasonare sau agitare cu perle de sticlã, cu ajutorul dehnicii de microdisecþie sau prin provocarea unui ºoc osmotic (inducþie
de plasmolizã sau plasmoptizã) ori a autolizei celulare. Examinat la microscopul electronic, peretele celular izolat are aspectul unui sac gol,
care conservã forma bacteriei din care provine, ºi constituþie asemãnãtoare ochiurilor unei
plase, cu tramã regulatã, care apare ca fiind formtã din fascicule de fibre paralele care se
încruciºeazã. Grosimea peretelui celular variazã între 15 ºi 35 nm, excepþional putând sã ajungã la
80 nm, ca la Lactobacillus acidophilus. Este mai subþire la celulele tinere care cresc mai
rapid. În funcþie de prezenþa ºi particularitãþile de structurã ale peretelui celular au fost
propuse trei diviziuni în cadrul regnului Procaryotae (Gibbson ºi Murray, 1978): 1. Firmacutes (l. firmus = tare; cutis = piele, înveliº), corespunzând bacteriilor Gram-
pozitive cu perete celular gros, lipsit însã de membrana externã; 2. Gracilicutes (l. gracilis = subþire), corespunzând bacteriilor Gram-negative cu perete
celular în mod obiºnuit subþire, având în structura sa membranã externã; 3. Mollicutes (l. mollis = moale, delicat), corespunzând genului Mycoplasma, care
cuprinde bacterii Gram-negative fãrã perete celular uniform. În funcþie de structurã ºi de compoziþia chimicã, pereþii celulari ai bacteriilor aparþin
la trei categorii, care coincid cu modul de colorare: Gram-pozitiv, Gram-negativ sau acidorezistent.
Pereþii celulari din cele trei categorii au drept constituient comun peptidogicanul, dar se deosebesc în ceea ce priveºte raportul, prezenþa, natura, cantitatea ºi aranjamentul structural
al proteinelor, polizaharidelor ºi lipidelor.

12
Peretele celular al bacteriilor Gram-pozitive apare la microscopul electronic ca un strat unic, relativ omogen, deºi frecvent pot fi deosebite mai multe structuri contigue,
foarte rar bine definite. Componentul major este peptidoglicanul (80-90% din greutatea uscatã), care apare
foarte net dupã colorare cu sãruri ale metalelor grele ºi poate fi degradat dupã tratare cu
lizozim. În afarã de peptidoglican, peretele celular al bacteriilor Gram-pozitive mai conþine
proteine ºi polizaharide care includ acizii teichoici, teichuronici, neteichoici ºi polizaharide neutre.
Peptidoglicanul (sin. mureinã), glicopeptid, mucopeptid, glicozaminopeptid, mucocomplex este un component parietal caracteristic ºi comun tuturor bacteriilor, cu
excepþia genului Mycolasma, a formelor L ºi a bacteriilor halofile extreme. Este unu heteropolimer compus dintr-o porþiune glican ºi o componentã peptidicã. Porþiunea glicanicã -foarte uniformã- are structura unor lanþuri lineare formate din
resturile alternente a douã N-acetilhexozamine diferite, legate între ele prin legãturi â-1,4. Componenta peptidicã conþine ca unitate de construcþie un tetrapeptid. Unitãþile
tetrapeptidice aparþinând lanþurilor de glican adiacente sunt legate prin intermediul unor
�punþi� specializate, interpeptidice. Astfel alcãtuit sacul peptidoglicanic este în esenþã o moleculã unicã, gigantã ºi rigidã,
având o structurã de reþea tridimensionalã care formeazã în jurul protoplastului bacterian o
reþea ca o plasã rigidã cu �ochiuri� mici. Peptidoglicanul poate fi atacat de lizozim ºi de penicilinã.
Acizii teichoici sunt molecule polimere lungi ºi flexibile (formate din 1,5-poli ribitol-fosfat ºi 1,3-poli glicerofosfat) localizate exclusiv în peretele celular, membrana plasmaticã ºi
straturile capsulare ale bacteriilor. Sunt prezenþi numai la bacteriile Gram-pozitive ºi aparþin la trei categorii:
acizii teichoici parietali (la suprafaþe celulei), legaþi covalent de stratul
peptidoglicanic al peretelui celular; acizi teichuronici legaþi de peretele celular; acizii teichoici membranari (sau lipoteichoici) legaþi covalent de fracþiunea
glicolipidicã a membranei plasmatice, formând o reþea între membrana
plasmaticã ºi peretele celular. Acizii teichoici conferã bacteriilor Gram-pozitive o serie de proprietãþi importante:
au rol esenþial în menþinerea unei concentraþii de ioni metalici, în particular
Mg2+ la suprafaþe externã a membranei plasmatice, importantã pentru activarea
unor sisteme enzimatice membranare dependente de cationi; au rol arhitactural; joacã un rol important ca determinanþi de patogenitate (inhibã fagocitoza ºi
protejeazã bacteriile contra efectului bactericid al fagocitozei); leagã ºi controleazã activitatea autolizinelor cu rol în creºterea ºi diviziunea
peretelui celular; acizii lipoteichoici acþioneazã ca receptori de fag ºi de colicine, creînd avantaje
adaptative bacteriilor Gram-pozitive.
Peretele celular al bacteriilor Gram-negative este mai subþire decât al
bacteriilor Gram-pozitive, este distinct stratificat, are o structurã mai complexã datoritã
prezenþei membranei externe, care apare cu o structurã similarã membranei plasmatice
(triplustrat). Este alcãtuit din urmãtoarele structuri:

13
1. Complexul peptidoglican-lipoproteinã care este situat în zona medianã a peretelui, are
o grosime de 1,5-3,0 nm ºi este electronodens; stratul peptidoglicanic reprezintã 2,4-10% din greutatea peretelui celular. La bacteriile Gram-negative sacul mureinic poate fi conceput ca o reþea
bidimensionalã sau ca un monostrat molecular, cu structurã relativ constantã. 2. Membrana externã a peretelui celular (numitã astfel pentru cã este situatã la exterior
faþã de stratul peptidoglicanic ºi de membrana plasmaticã) are o grosime de 6-20 nm. Membrana externã este alcãtuitã din fosfolipide (35%), care formeazã un strat intern
continuu, proteine (15%) ºi lipopolizaharide (50%), care sunt asociate cu stratul extern.
Fig.1. Reprezentarea schematicã a înveliºuilor celulare, evidenþiind structura membranei
externe a peretelui celular ºi raporturile cu spaþiul periplasmic ºi membrana citoplasmaricã. (G. Zarnea, 1983, dupã DiRienzo ºi Inouye, 1980)
Semnificaþia biologicã a peretelui celular Peretele celular reprezintã sistemul de susþinere mecanicã, un fel de corset neelastic,
al întregii arhitecturi celulare. Datoritã rigiditãþii sale determinã celulelor o formã caracteristicã. Asigurã protecþia faþã de ºocul osmotic. Presiunea osmoticã internã este de ~5-6 atm.
(la Escherichia coli de ~20-30 atm.). Fãrã suportul rigid al peretelui celular membrana
plasmaticã nu ar putea rezista acestei presiuni interne ºi nu ar putea asigura protejarea
protoplastului. Participã la procesul de creºtere ºi diviziune celularã, urmând membrana
citoplasmaticã în formarea septurilor transversale, care, dupã replicarea cromozomului
bacterian separã celula-mamã în douã celule-fiice. Unii constituenþi parietali imprimã suprafeþei celulare o anumitã specificitate, jucând
rolul de receptori pentru unii bacteriofagi, determinând fenomenul de �recunoaºtere� a
celulelor apte de conjugare sau, în cazul bacteriilor patogene, acþionând ca factori antigenici
capabili sã inducã în organismul animal infectat elaborarea de anticorpi specifici care pot servi ca receptori celulari de suprafaþã.

14
Conþine enzime autolitice capabile sã atace structura glicopeptidului, active în
momentul sporulãrii, al eliberãrii sporului prin liza sporangelui, al germinãrii, ca ºi în biosinteza peretelui celular ºi în reglarea acestui proces.
La bacteriile Gram-negative, prezenþa membranei externe ºi a constituenþilor ei
conferã bacteriilor respective proprietãþi deosebite în raport cu mediul înconjurãtor: acþioneazã ca barierã impermeabilã pentru molocule hidrofobe, împiedicând
intrarea în celulã a diferitelor substanþe potenþial nocive ºi pierderea unor
metaboliþi necesari; funcþioneazã ca o �sitã molocularã� asigurând trecerea unor oligopeptide,
oligozaharide ºi a unor substanþe hidrofile; transportul pasiv este mediat de molecule proteice transmembranare, numite porine care formeazã adevãrate
canale moleculare prin membrana externã; membrana extrnã reþine în spaþiul periplasmic enzimele degradative sintetizate
în celulã, dupã ce au traversat membrana plasmaticã, precum ºi o varietate de
molecule nutritive; prin aceastã funcþie activitãþile enzimatice esenþiale pentru
celulã, dar potenþial dãunãtoare pentru constituenþii citoplasmatici, au loc în
afara membranei plasmatice, iar degradarea moleculelor nutritive mari la monomeri simpli este efectuatã în apropierea proteinelor de legare ºi a
permeazelor specifice care permit transportul selectiv în celulã a unor molecule
mai mari de 700 daltoni; este sediul unor sisteme de transport specifice: pentru vitamina B12, maltozã,
maltodextrine, ioni ferici ºi nucleozide); este implicatã în adsorbþia unor fagi.
3.2. Protoplaºtii ºi sferoplaºtii
Protoplastul reprezintã ansamblul structurilor celulare rãmase dintr-o bacterie Gram-
pozitivã dupã îndepãrtarea integralã a peretelui celular sub acþiunea lizozimului. Îmbrãcat în membrana citoplasmaticã, protoplastul se menþine în medii izotonice sau
hipertonice ca formaþiune aproximativ sfericã, pãstrând, în general, proprietãþile ºi activitãþile
vitale ale celulei din care provine (capacitatea respiratorie normalã, sinteza de proteine ºi de
acizi nucleici, viabilitatea în culturi ºi, în anumite condiþii, chiar capacitatea de diviziune ºi
aptitudinea de a asigura multiplicarea fagului al cãrui acid nucleic se gãsea deja în celulã în
momentul pierderii peretelui celular). Protoplastul nu este capabil sã refacã prin sintezã peretele celular dacã acesta a fost
complet îndepãrtat. Funcþia de barierã osmoticã asigutatã de membrana citplasmaticã, este
mult mai labilã în lipsa protecþiei conferite de peretele celular rigid ºi, ca urmare, în medii
hipotonice protoplastul lizeazã. La bacteriile Gram-negative, peretele celular nu poate fi degradat complet, astfel cã
tratarea cu lizozim îi slãbeºte doar structura prin degradarea selectivã a mucocomplexului. În
medii hipotonice celula astfel tratatã se lizeazã, iar în mediile hipertonice se transformã în
sferoplast, corp sferic echivalent unui protoplast, înconjurat de constituenþii peretelui celular
care au rezistat la acþiunea lizozimului. Protoplaºtii aparþinând unor specii diferite de bacterii (B. megaterium ºi B. subtilis)
fuzioneazã în prezenþa unor substanþe ce acþioneazã ca agenþi de fuzionare , producând
formarea de bacterii bi- sau multinucleate care în cursul reversiei la forma bacilarã (prin
refacerea peretelui celular) sau unterior, într-o perioadã scurtã de timp, duc la apariþia unor
bacterii cu genotip haploid, modificat prin recombinare geneticã cu caractere provenind de la
cele douã celule originare (Schaeffer, 1976, Fodor ºi Alföldi, 1976).

15
3.3. Spaþiul periplasmic
Spaþiul periplasmic sau zona periplasmicã este un compartiment întâlnit numai la
bacteriile gram-negative, delimitat spre interior de membrana citoplasmaticã, iar spre exterior
de un strat care acþioneazã ca o sitã molecularã, corespunzând membranei externe a peretelui
celular (Mitchell, 1961). Spaþiul periplasmic ºi complexul peptidoglican-lipoproteinã ocupã aceeaºi zonã a
peretelui celular, în care primul adãposteºte constituenþii periplasmici, iar celãlalt susþine ºi
întãreºte înveliºurile celulei. Spaþiul periplasmic conþine numeroase enzime implicate în nutriþie, proteine
specifice, neenzimatice �proteine de legare�, care leagã specific anumite substanþe (zaharuri,
aminoacizi, ioni anorganici) transportându-le pânã la nivelul proteinelor de transport legate de
membranã. Semnificaþia biologicã
Membrana externã a peretelui celular lasã sã treacã în regiunea periplasmaticã
numeroase substraturi hidrofile (echivalente cu tetra- ºi pentamere de aminoacizi, dimeri ºi trimeri de carbohidraþi). Funcþia principalã a enzimelor periplasmice (fosfataze, sulfataze,
amidaze etc.) este de a pregãti chimic substanþele care difuzeazã prin membrana externã
pentru trecerea lor prin membrana plasmaticã în citoplasmã. Prin acest mecanism, bacteriile
Gram-negative prin intermediul enzimelor degradative acþioneazã asupra unei largi varietãþi
de substraturi întâlnite în naturã, convertind substanþele care difuzeazã în spaþiul periplasmic
în molecule transportabile în celulã, unde sunt accesibile imediat proteinelor de legare ºi
permeazelor. Acest aspect conferã bacteriilor Gram-negative un deosebit avantaj biologic. De aceea bacteriile adaptate la viaþa în mãri, râuri, lacuri etc. sunt în majoritatea lor Gram-negative.
Spre deosebire de bacteriile Gram-negative, cele Gram-pozitive elibereazã cea mai
mare parte a enzimelor lor extracelular, în mediu. Acest mecanism este avantajos pentru
bacteriile care trãiesc în condiþii de mare concentraþie de substrat ºi de mare densitate populaþionalã, deoarece produºii de digestie ai enzimelor extracelulare, neasociate cu peretele
celular, sunt accesibili tuturor celulelor aflate în vecinãtatea locului lor de producere.
3.4. Membrana plasmaticã
Membrana plasmaticã (membranã citoplasmaticã, membranã celularã) este formaþiunea structuralã care acoperã de jur împrejur citoplasma bacterianã, separând-o de suprafaþa internã a peretelui celular, de care este de obicei strâns lipitã, datoritã diferenþei de
presiune osmoticã între conþinutul celular ºi mediul extern. Membrana plasmaticã poate fi pusã în evidenþã la microscopul fotonic, fie dupã o
colorare selectivã cu albastru Victoria, fie prin examinarea bacteriilor vii la microscop, în
câmp întunecat, în care caz apare ca o linie netã, luminoasã, strãlucitoare. Membrana plasmaticã examinatã la microscopul electronic pe secþiuni ultrafine � în
condiþii tehnice standard - apare ca o formaþiune triplu stratificatã, cu o grosime de 7,5-10 nm, alcãtuitã din douã straturi întunecate care separã un strat mai clar.
Robertson a denumit aceastã structurã unitate de membranã (unit-membrane), pornind de la ideea cã ea reprezintã unitatea structuralã din care sunt alcãtuite structurile
membranare complexe. Aspectul de triplu strat a dus la elaborarea modelului de structurã greºit, în acord cu care stratul bimolecular de fosfolipide ar fi mãrginit de fiecare parte de un
strat proteic (Robertson, 1969), variantã a modelului propus iniþial de Danielli-Dawson (1935).

16
Modelul de structurã acceptat în unanimitate � modelul mozaicului fluid (Singer-Nicolson, 1972) � propune o aºezare caracteristicã a componentelor specifice ale membranei
reprezentate de lipide, proteine ºi glucide în acord cu proprietãþile lor, satisfãcând exigenþele
termodinamice ºi furnizând o explicaþie satisfãcãtoare a proprietãþilor generale ale
biomembranelor. Dupã acest model fosfolipidele formeazã un film fluid, discontinuu, în care
�plutesc� proteinele globulare, în timp ce glucidele interacþioneazã fie cu unele fie cu altele.
Fig.2. Evidenþierea structurii internea membranei plasmatice, prin tehnica de îngheþare-fracturare de-a lungul planului central al stratului dublu lipidic (G. Zarnea, 1983, dupã Staehlin ºi Hull, 1979)
Lipidele sunt reprezentate de fosfolipide � molecule amfipatice cu structurã complexã,
având o extremitate polarã, hidrofilã (hidrosolubilã în stare izolatã), alcãtuitã dintr-o grupare fosfat (PO-
4) ºi alþi constituenþi, legaþi printr-o moleculã de glicerol � care formeazã un fel de
punte de �cozile� moleculei, reprezentate de doi acizi graºi, ce constitue regiunea nepolarã,
hidrofobã (insolubilã în apã) a moleculei. În contact cu apa grupãrile polare poartã de regulã
sarcini electrice. Deoarece cele douã extremitãþi ale moleculelor lipidice au solubilitãþi incompatibile,
în suspensie apoasã ele se organizeazã spontan pentru a forma un dublu strat de molecule,
discontinuu ºi fluid-vâscos, în care moleculele sunt aranjate �coalã-la-coadã� în aºa fel încât
capetele ionice polare (hidrofile) sunt expuse spere soluþia apoasã � de ambele pãtþi ale
membranei, iar lanþurile nepolare (hidrofobe) ale acizilor graºi sunt orientate în direcþia opusã
contactului cu apa. În felul acesta, cele douã monostraturi de molecule formeazã împreunã
douã straturi hidrofile periferice separate de porþiunea centralã hidrofobã. Aceastã modalitate
de aranjare reprezintã configuraþia de minimã energie posibilã pentru o suspensie de lipide în
apã ºi este, în acelaºi timp, foarte adecvatã pentru funcþia de barierã între douã soluþii apoase
(cun sunt interiorul celulei ºi mediul extern). Structura de dublu strat fosfolipidic explicã
proprietãþile de permeabilitate ale membranei, deoarece acest strat este impermeabil faþã de
particulele încãrcate ºi faþã de ioni ºi uºor penetrabil de moleculele liposolubile. Fiecare dublu strat este un �lichid bidimensional� în care moleculele lipidice difuzeazã
lateral, schimbându-ºi poziþia pânã la un milion de ori pe secundã. În schimb deplasarea unei
molecule de pe un monostrat pe altul (trenziþia �flip-flop�) se face foarte rar (cel mai des o
datã pe lunã pentru o moleculã datã). Raritatea deplasãrilor �flip-flop� ale lipidelor ºi
proteinelor permite menþinerea compoziþiei membranei ºi a structurii ei caracteriestice. Dublul strat fosfolipidic trebuie sã fie suficient de fluid pentru a permite miºcarea
liberã a proteinelor membranare implicate în procesele de transport activ.

17
Fosfolipidele formeazã matricea structuralã a membranei ºi sunt rãspunzãtoare de
integritatea structuralã a acesteia. Prin structura caracteristicã a dublului strat, ele conferã
membranei impermeabilitatea la cele mai multe molecule hidrosolubile, care sunt insolubile în regiunea �uleioasã� a pãrþii de mijloc a membranei.
Proteinele, în raport cu poziþia lor în structura membranei, sunt de douã tipuri: proteinele integrate (intrinsece); proteinele de suprafaþã (periferice sau extrinsece).
Proteinele integrate, în general insolubile în apã, nu pot fi îndepãrtate fãrã ruperea
dublului strat lipidic. Au o orientare fixã: fiecare proteinã de acelaºi tip este îndreptatã în
aceeaºi direcþie. Cele mai multe strãbat toatã grosimea membranei celulare (�proteinele
transmembranare�) dar unele pot fi expuse fie numai pe suprafaþa internã (citoplasmaticã), fie
spre suprafaþa externã. Regiunile lor dirijate spre interior ºi/sau spre exterior au caracter
hidrofil, ceea ce împiedicã tranziþia lor tip �flip-flop�. Proteinele de suprafaþã, neinserate în dublul strat lipidic, sunt în general hidrosolubile
ºi situate fie pe suprafaþa internã, fie pe cea externã, de regulã legate de proteinele integrate. Din punct de vedere funcþional, proteinele de membranã pot fi:
enzime care fac biosinteza înveliºurilor celulare (membrana plasmaticã,
polimeri parietali, extraparietali), proteine de transport care asigurã transportul moleculelor solubile din mediu în
celulã ºi invers; citocromi ºi alte proteine aparþinând sistemului transportor de electroni; proteine cu activitate adenozinfosfatazicã (ATP-aza); proteine implicate în turnover-ul lipidelor ºi al proteinelor membranare
(fosfolipaze, proteaze, peptidaze).
Fig. 3. Membrana plasmaticã � reprezentarea schematicã a unui model tridimensional (G. Zarnea, 1983, dupã Lodish ºi Rothman, 1979)
Glucidele reprezintã componentul cel mai slab reprezentat în structura membranei ºi
se regãsesc sub forma unor polizaharide legate de proteine (glicoproteine) sau interacþionând
cu anumite lipide (glicolipide).

18
Semnificaþia biologicã Membrana plasmaticã reprezintã singura suprastructurã citoplasmaticã permanentã a
celulei bacteriene, având rolul de a delimita spaþiul intracelular. Ea formeazã un compartiment
închis, dar nu reprezintã o graniþã fizicã inertã a celulei, ci o structurã funcþionalã capabilã sã
asigure o deosebire netã între interiorul ºi exteriorul acesteia. Aceastã proprietate este
consecinþa faptului cã membrana plasmaticã prezintã o asimetrie funcþionalã, cu importanþã
esenþialã pentru viaþa celulei, în sensul cã suprafaþa internã funcþioneazã diferit de cea externã.
Astfel un ion sau o moleculã pompatã la interior printr-un punct al membranei ar putea fi eliminat în altul cu o cheltuialã inutilã de energie. Aceastã asimetrie funcþionalã are la bazã o
asimetrie de structurã molecularã, manifestatã pe mai multe cãi: cele douã monostraturi lipidice includ proporþii variate ale diferitelor tipuri de
molecule lipidice; carbohidraþii sunt prezenþi numai pe suprafaþa externã a membranei, în timp ce
proteinele periferice sunt situate aproape întotdeauna pe faþa internã; fiecare tip de proteinã integratã are o orientare definitã, care este aceeaºi pentru
fiecare moleculã de acelaºi tip. Membrana plasmaticã este asociatã cu toate funcþiile (transport, fosforilare), care la
organismele superioare sunt legate de structuri diferenþiate. Membrana plasmaticã poate sã-ºi mãreascã suprafaþa, furnizând mai mult spaþiu
pentru diferite activitãþi celulare (respiraþie, fotosintezã) prin invaginare, formând sisteme de
membrane, care uneori se ramificã în citolpasmã, sau se pot detaºa, formând entitãþi virtual independente.
Funcþioneazã ca o �barierã osmoticã�, dotatã cu impermeabilitate cvasitotalã faþã de
multe tipuri de mlecule, permiþând trecerea nestânjenitã a altora. Ea asigurã în acest fel
schimburile necesare ºi selective între mediul extern ºi cel intracelular, menþinând constantã
compoziþia chimicã ºi ionicã a celulelor, care, la rândul lor, influenþeazã critic numãrul enorm
de reacþii interdependente ce au loc în citoplasmã. Proprietãþile de permeabilitate ale membranei plasmatice pot fi sintetizate astfel:
substanþele uºor solubile în solvenþii lipidelor, ca ºi unii anioni (ex. Cl-) traverseazã uºor biomembranele;
unii ioni ca Na+, K+, glucidele ºi proteinele, nu o pot traversa uºor, celula
recurgând la mecanisme speciale de transport. Membrana plasmaticã bacterianã ºi structurile derivate din ea formeazã baza
structuralã a sinergonului fotosintezei ºi a celui respirator, reprezentând prin aceasta un
echivalent funcþional al cloroplastelor, respectiv al mitocondriilor din celulele eucariote. Este implicatã în mobilitatea bacterianã, datoritã faptului cã una din structurile
corpului bazal al flagelului este strâns legatã de structura sa. Unele proteine legate de membranã sau aflate în contact lax cu ea (fiind localizate în
spaþiul periplasmic) joacã rolul de chemoreceptori. Participã la formarea ºi eliminarea unor proteine, ca enzimele ºi exotoxinele, care pot
fi sintetizate în membrana plasmaticã, sau pe suprafaþe ei externã. În primul caz, trecerea lor
extracelular s-ar realiza fie printr-un mecanism de tipul pinocitozei inverse, fie datoritã unei
porþiuni glucidice legate, care ar facilita eliberarea. În al doilea caz, moleculele sintetizate ar
difuza liber în mediu.
3.5. Mezosomii
Mezosomii (Fitz � James, 1967) au fost descriºi iniþial sub denumiri diferite: membrane intracitoplasmatice (Hopwoord, 1960), corpi periferici (Chapman, 1953), condrioizi (Van Iterson, 1961) sau plasmalemasomi (Edwards, 1963).

19
Greenawalt (1975) considerã mesozomii structuri membranoase intracitoplasmatice,
caracterizate prin trei particularitãþi definitorii: derivã ultrastructural din membrana celularã ca o invaginare în formã de �sac�
sau �pungã�, care conþine corpi membranoºi; pot fi extrudaþi din sacul mezosomal în spaþiul dintre membrana plasmaticã ºi
peretele celular prin agitare, îndepãrtarea peretelui celular sau dupã plasmolizã; sunt asociaþi fizic ºi/sau topografic cu replicarea ºi segregarea cromosomului,
cu formarea septului de diviziune ºi cu sporularea. Structurile membranare care nu întrunesc aceste condiþii trebuie numite membrane
intracelulare (intracitoplasmatice) sau structuri similare mezosomilor (�mezosome-like�). La microscopul electronic apar sub forma unor structuri cu formã, mãrime, localizare
ºi complexitate foarte diferite, în general putând varia nu numai în funcþie de starea
fiziologicã a celulei, ci ºi de calitatea ºi natura tehnicilor de prfixare ºi fixare, precum ºi de
unghiul de secþionare.
Fig. 4. Structura schematicã a mezosomului bacterian, evidenþiind modul de pãtrundere a citoplasmei în structura acestuia, într-un spaþiu care nu este conectat nici cu cisternele
tubululor, nici cu mediul extern. Tubulii din sacul mezosomal apar pe secþiunea transversalã
circulari. Situsul de legare al genomului este localizat pe faþa citoplasmaticã a sacului
mezosomal (G. Zarnea, 1983, dupã Reusch ºi Burger, 1973).
Au fost descrise trei tipuri morfologice de mezosomi, dupã unii autori
interconvertibile: lamelari (formaþi prin plierea membranei invaginate într-un aranjament în
spiralã încolãcitã ca un ghem); veziculari sau saciformi (vezicule aproape sferice); tubulari (de forma unor tubuºoare lungi).
Dupã localizare mezosomii pot fi: septali; periferici; nucleari.

20
Mezosomii au caracteristicile structurale ale membranei plasmatice din care derivã (structura triplustratificatã ºi grosimea de 7,5 � 8,0 nm).
Sunt mai numeroºi ºi mai bine dezvoltaþi la bacteriile Gram-pozitive, în timp ce la
bacteriile Gram-negative sunt relativ mai greu de observat, fiind în general rudimentari, slab
dezvoltaþi ca mãrime ºi mai puþini ca numãr. Prin expunere în medii hipertonice, mezosomii sunt extrudaþi în spaþiul dintre
membranã ºi peretele celular sub forma unor filamente, ca un ºirag de mãrgele, care poate
ajunge la dimensiunea de 20 nm.
Fig. 5. Reprezentarea schematicã a stadiilor de formare a mezosomilor. A, B. Dezvoltarea unui mezosom sacular turtit, dintr-un mezosom sferic (vezicular). x ºi y reprezintã punctele
terminale ale sacului ºi situsurile potenþiale de creºtere ale membranelor intracelulare. C. Pentru simplificare diagrama ilustreazã numai dezvoltarea în punctul x printr-o secvenþã de
sãgeþi ºi vezicule, deºi procesul are loc ºi în punctul y. D. Mezosom cu structurã complexã
multilamelarã, format prin creºterea ºi plierea sacului turtit prin extinderea lui x la x` ºi a lui y la y`. (G. Zarnea, 1983).
Originea, creºterea ºi diferenþierea mezosomilor Formarea mezosomilor este un proces complex, care începe cu invaginarea
membranei plasmatice ºi sfârºeºte cu legarea de genomul bacterian; are loc în special în zona
în care creºterea membranei se face mai repede decât a peretelui celular, ceea ce are drepr
consecinþã invaginarea acestuia sub forma unor �pungi� pline cu membrane (Rogers, 1970). Invaginarea iniþialã duce la formarea unui sac sferic, conectat cu membrana printr-un
peduncul de lungime variabilã, deschis spere mediul extern ºi/sau spaþiul periplasmic. Mezosomul simplu, sferic, suferã în continuare modificãri rezultate din diferite grade de
turtire însoþite de invaginãri secundare, care duc la formarea de lamele, vezicule ºi tubuli. În
cazul în care complexitatea mezosomului creºte, dezvoltarea se face pe seama membranei nou

21
sintetizate care se adaugã membranei invaginate. Concomitent cu creºterea în complexitate
apare posibilitatea unui grad mai mare de compartimentare a constituenþilor citoplasmatici,
chiar în mezosomi, care apar la microscopul electronic sub forma unor canale rezultate din
invaginãrile secundare. Nu se cunoaºte natura stimulilor implicaþi în invaginare ºi nici a celor care determinã
modificãri de formã, localizare ºi complexitate. Semnificaþia biologicã
Mezosomul nu este o structurã staticã ci este un organit influenþat permanent de
dinamica proceselor celulare. Morfologia mezosomului este influenþatã de condiþiile de prelucrare premergãtoare
examinãrii la microscopul electronic (prefixare, calitatea ºi natura fixãrii chimice etc.), care
afecteazã tipul, forma ºi poate chiar localizarea în celulã. Unii autori considerã mezosomii ca fiind structuri vestigiale sau redundante, versatile
(instabile) ºi multifuncþionale, neesenþiale pentru viabilitatea bacteriilor, având o deosebitã
plasticitate structuralã, ca un rãspuns la nevoi specifice, variate. Formarea mezosomilor este modalitatea prin care celula bacterianã are posibilitatea
de a-ºi mãri, prin invaginare ºi pliere, suprafaþa membranei plasmatice, ca rãspuns la
condiþiile de mediu, de aceea, se considerã cã nu ar avea funcþii diferite sau suplimentare faþã
de cele ale membranei plasmatice. Rolul mezosomilor în replicarea genomului bacterian a fost demonstrat pe baze
morfologice, genetice ºi biochimice. Fiind legat de membrana plasmaticã, cât ºi de materialul
nuclear are un rol important în controlul replicãrii cromosomului ºi al plastidelor bacteriene, prin transmiterea semnalului biochimic care ia naºtere la suprafaþa celulei. La B. subtilis, în
faza iniþialã, fiecare nucleosom este legat de un mezosom, pentru ca pe mãsurã ce celula
creºte ºi se apropie de diviziune mezosomul sã se dividã, iar formaþiunile rezultate sã migreze
spre extremitãþile celulei, antrenând cromosomii bacterieni progeni. Forþa care realizeazã
aceastã deplasare este reprezentatã de sinteza ºi încorporarea de constituenþi noi de membranã
celularã. Mezosomii participã la reacþii de fosforilare, oxidoreducere ºi transport de electroni,
care au loc însã cu o pndere mai micã decât în membrana plasmaticã. Conþin fosfataze acide, esteraze etc. ºi ar putea funcþiona ca �organite� subcelulare
degradative, asimilabile funcþional cu lizosomii din celulele eucariote (Revsch, 1972). Au rol în unele procese secretorii ca, de exemplu, în producerea ºi eliberarea unor
exoenzime ca �penicilinaza�. Mezosomii ar fi implicaþi în sinteza înveliºurilor celulare, în mod particular a
membranei plasmatice, a peretelui celular ºi a septului transversal care separã celulele dupã diviziune. Tichy ºi Landman (1969) considerã cã ar reprezenta o regiune specializatã prin
care, în cursul transformãrii genetice, ADN transformat ar pãtrunde în celulã.
3.6. Citoplasma
Citoplasma bacterianã este, în general, consideratã ca un sistem coloidal complex format din proteine, glucide, lipide, apã ºi substanþe minerale, sistem în care apa ºi compuºii
dizolvaþi în ea acþioneazã ca faza externã a unui sol (Lamanna, 1949). Deoarece în citoplasmã
coexistã stãrile de emulsie ºi de soluþie, aceasta poate fi consideratã ca un complex de stãri
fizice într-o continuã transformare. Se caracterizeazã prin menþinerea permanentã a stãrii de
gel � ceea ce are ca rezultat o imobilitate a conþinutului (lipsa curenþilor citoplasmatici) � ºi reprezintã o condiþie indispensabilã a menþinerii nemicibilitãþii nucleului cu citoplasma, având
în vedere absemþa unor membrane intracelulare.

22
Citoplasma bacterianã nu are o organizare definitã, similarã celulei eucariote, care conþine organite diferenþiate ºi delimitate de membrane. La celulele tinere ºi în condiþii
normale de dezvoltare, citoplasma aderã la peretele celular ºi se prezintã ca o masã densã,
omogenã ºi intens colorabilã. La celulele bãtrâne citoplasma îºi pierde treptat afinitatea
tinctorialã, se retractã centripet îndepãrtându-se de perete ºi capãtã o structurã granularã cu
vacuole din ce în ce mai evidente la microscopul electronic. La bacteriile examinate pe secþiuni mai puþin fine (~ 50 nm), zona citoplasmaticã
apare ca fiind plinã de agregate de ribosomi, aspect rezultat din suprapunerea acestor structuri
cu Ø de 10 nm. Pe secþiuni extrem de subþiri citoplasma apare ca masã amorfã ºi lipsitã de
particule structurale, cu ribozomi rari. În interiorul citoplasmei se gãsesc materialul nuclear, incluziunile, vacuolele ºi
ceilalþi constituenþi ai protoplastului. O caracteristicã a citoplasmei bacteriene este prezenþa unei mari cantitpþi de ARN,
ceea ce explicã bazofilia ei intensã, mai evidentã la celulele tinere. Celulele moarte se
coloreazã mai puþin intens, ca ºi celulele bãtrâne, la care sinteza ARN a încetat, iar cel
existent a fost folosit ca sursã de N ºi P.
3.7. �Nucleul�
Spre deosebire de celulele eucariote care au un nucleu cu structurã bine definitã,
mãrginit de o membranã ºi conþinând un numãr definit de cromozomi capabili de diviziune
mitoticã, �nucleul� bacterian reprezintã o formã primitivã de organizare, lipsitã de membranã
(de tip procariot), inclavatã direct în citoplasmã, în mod obiºnuit în partea centralã a celulei ºi
care nu suferã modificãri de tip mitotic în cursul ciclului de diviziune. Datoritã caracterelor sale particulare, aceastã structurã a fost desemnatã cu numeroºi
termeni ca: nucleoid, nucleosom, material nuclear, nucleoplasmã, echivalent nuclear sau "nucleu" prin analogie funcþionalã cu structura echivalentã a celulelor eucariote.
Deoarece citoplasma bacterienã este puternic bazofilã ca urmare a prezenþei ARN-ului, nu este posibilã diferenþierea la microscopul fotonic cu coloranþi bazici a materialului
nuclear constituit din ADN, la fel de bazofil. Evidenþierea corpului cromatic bacterian la microscopul fotonic nu se poate face prin
colorare selectivã decât dupã îndepãrtarea ARN-ului citoplasmatic prin hidrolizã acidã (tehnica Robino ºi Feulgenw) sau enzimaticã (tehnica Boivin, cu ribonucleazã). Cu aceste
tehnici materialul nuclear apare sub diferite forme (granule sferice sau ovalare, halterã, bastonaºe izolate sau dispuse în V, filament axial etc.), reprezentând 5-16% din volumul celulei.
Materialul nuclear este localizat în mod obiºnuit în partea centralã a celulei. El se
prezintã ca o zonã mai clarã, cu o densitate medie mai micã decât aceea a citoplasmei
înconjurãtoare, astfel cã la bacterii contrastul dintre structurile intracelulare este inversat în
raport cu acela caracteristic celorlalte celule, la cere nucleul este mai dens decât citoplasma.
Acest aspect este determinat de densitatea neobiºnuit de mare a citoplasmei bacteriene. Pe
secþiuni ultrafine se observã cã regiunea nuclearã este plinã cu fibrile fine cu Ø între 2,0 ºi 5,0
nm, uneori aranjate în ºiruri ondulate, paralele, care dispar dupã digestia cu
dezoxiribonucleazã, ceea ce aratã cã sunt formate din ADN ºi corespund cromosomului
bacterian. Cromosomul bacterian denumit ºi lineom, nucleosom sau genofor, s-a dovedit a fi
alcãtuit dintr-o singurã moleculã de ADN, de formã circularã (extremitãþile sunt reunite), cu o
lungime de 1 400 ìm ºi un Ø de 2,5 nm, corespunzând diametrului moleculei de ADN dublu catenar.

23
Organizarea fizicã a nucleului Molecula de ADN depãºeºte de aproximativ 1 000 de ori lungimea linearã a celulei.
Ea este �împachetatã� pentru a forma un corp cromatic de 1 500 de ori mai mic decât propria sa
dimensiune în stare desfãºuratã. Teoretic �împachetarea� unei molecule atât de lungi într-un volum mic implicã plierea moleculei de cel puþin 1000 de ori ºi menþinerea ei în stare compactã.
Au fost propuse mai multe modele pentru a explica mecanismul acestui proces � care dacã s-ar face la întâmplare ar duce la �încuracrea� moleculei de ADN astfel încât o parte din
infermaþia geneticã ar deveni inaccesibilã pentru transcriere. Stonington ºi Pettijohn (1971) au izolat nucleul din E. coli sub forma unei structuri
compacte cu g.m. 2,5 x 109±0,5 x 109 dal (1 600 � 1 700 S). Aceastã structurã este alcãtuitã
din ADN corespunzând genomului pliat, complexat cu ARN ºi proteine ºi este foarte sensibilã
la Rnazã. Complexul ADN-ARN-proteinã conþine 80% ADN; ARN-ul reprezintã 10% din
greutate ºi este format din ARNr ºi ARNm, în stare nãscândã, iar proteina care formeazã tot
10% (mai puþin de 1% din proteina celularã totalã), este ARN-polimerazã (subunitãþile á, â ºi
â�). Dupã tratare cu ribonucleazã, din acest complex se elibereazã o moleculã de ADN dublu
helicatã depliatã, ceea ce demonstreazã cã ARN are rolul de a stabiliza ºi menþine forma
condensatã (compactã) a nucleului.
Fig. 6. Modelul lui Worcel ºi Burgi (1072)
pentru plierea cromosomului la E. coli. Cromosomul formeazã ~ 50 bucle superhelicale în jurul unei structuri celulare formatã din
ARN. Trastarea cu DN-azã, care produce incizii
monocatenare, elibereazã bucle individuale ºi
reduce progresiv valoarea constantei de sedimentare (S) del la 1500 S laa 155 S. Tratarea cu ribonucleazã depliazã complet
cromosomul, dându-i formã care sedimenteazã
mai lent (G. Zarnea, 1983).
Fig. 7. Modelul lui Pettijohn (1974) de împachetare
a cromosomului bacterian. A. Cromosom circular nepliat cu Ø 350 ìm. B. Cromosom pliat cu 7 bucle. În realitate, ar exista 40-80 domenii de pliere, corespunzând unui Ø de 30 ìm. C. Cromosom pliat în care buclele au suferit transformare superhelicatã.
Dacã pasul suprahelici este de 11 nm, Ø nucleului
este de 2 ìm. Modelul bidimensional devine tridimensional prin plierea zonelor superhelicale deasupra ºi dedesubtul planului hârtii. DNaza produce incizii monocatenare ºi desface
superhelicea în bucla respectivã, fãrã a afecta
structura superhelicalã a altor domenii (D). E. Secþionarea a douã molecule de ARN ale unor domenii adiacente uneºte cele douã domenii fãrã
pierderea structurii lor superhelicale (G. Zarnea, 1983).

24
Pettijohn ºi Hecht (1974) au elaborat un model în acord cu care ADN din
cromosomul de E. coli ar fi condensat într-o structurã compactã printr-un proces de pliere ºi formare de superhelice, în care structura condensatã a ADN este menþinutã de molecule de
ARN, ce leagã ºi stabilizeazã buclele de pliere. Pentru a menþine starea condensatã a ADN
este necesarã o sintezã continuã de ARN, fapt care explicã asocierea ARN-polimerazei. Rouviere-Yaniv (1975) a izolat de la E. coli o proteinã specificã, proteina HU,
asociatã cu ADN ºi implicatã în plierea lui în celulã. În mod nomal, bacteriile aflate în faza de repaus în culturi staþionare ºi vechi au un
singur cromosom, astfel cã sunt unicelulare. În faza de creºtere activã, în culturi tinere pe medii optime, ele apar ca multinucleate,
având 2-4 cromosomi, care sunt genetic identici, deoarece provin din replicarea dintr-un singur cromosom parental. De aceea, indiferent de aspectul morfologic al materialului nuclear, din punct de vedere genetic, bacteriile sunt organisme haploide, astfel încât chiar
atunci când celula primeºte un aport de material nuclear exogen � prin procese de transfer genetic � diploidia nu este decât parþialã ºi tranzitorie.
Apariþia bacteriilor multinucleate este în mod obiºnuit rezulatul unei lipse de
sincronizare între ritmul de creºtere ºi ritmul de diviziune celularã.
Semnificaþia biologicã Cromosomul bacterian poartã în structura sa toatã informaþia geneticã esenþialã,
necesarã pentru existenþa unei celule, respectiv setul de determinanþi genetici reprezentând
acel minimum necesar pentru ca o bacterie sã poatã ocupa poziþia s-a ecologicã normalã: gene
necesare pentru metabolismul energetic, pentru biosintezã, creºtere ºi diviziune ºi, în acelaºi
timp, pentru reglarea activitãþilor celulare. El determinã ºi arhitectura celulei bacteriene,
ereditatea ºi capacitatea de evoluþie a acesteia.
3.6. Ribosomii
Sunt particule nucleoproteice intracitoplasmatice de formã aproximativ sfericã. Au diametrul de aproximativ 20 nm. Pot fi caracterizaþi dupã constanta de sedimentare la ultracentrifugã (unitãþi Svedberg
� S) ºi prin apacitatea d a participa la sinteza proteinelor in vitro. La procariote se gãsesc în citoplasmã 15 000-10 000 particule ribozomale, cu
constanta de sedimentare 70 S cu tendinþa de a se disocia rapid în douã subunitãþi inegale cu
constantele de sedimentare de 30 S ºi 50 S. Mãrimea ºi stabilitatea lor este în funcþie de concentraþia ionilor Mg
2+ ºi K+. Subunitatea micã 30 S (g.m. 900 000 dal) este alcãtuitã din 21 molecule de proteine
diferite, notate de la S1 la S21 ºi o moleculã de ARNr, notatã 16 S. Subunitatea ribosomalã micã are trei regiuni: capul (1/3 din subunitatea micã), baza
(formeazã restul de 2/3) ºi platforma, care este separatã de cap printr-o scobiturã numitã
fisurã sau despicãturã. Subunitatea mare 50 S (g.m. 1 000 000 dal) este alcãtuit din 34 de proteine diferite,
notate de la L1 la L34 ºi ouã molecule de ARNr, una de 23 S ºi cealaltã de 5 S. Moleculele 16 S ºi 23 S aleARNr nu sunt transcrise separat, ci derivã din degradarea
catabolizatã de o nucleazã a unui precursor mare de 30 S (pre-ARNr). Modelul de structurã al subunitãþii mari include o protuberanþã centralã, flancatã de
câte o prelungire de fiecare parte, cea mai extinsã fiind denumitã peduncul,iar cealaltã creastã, între ele având o vale.
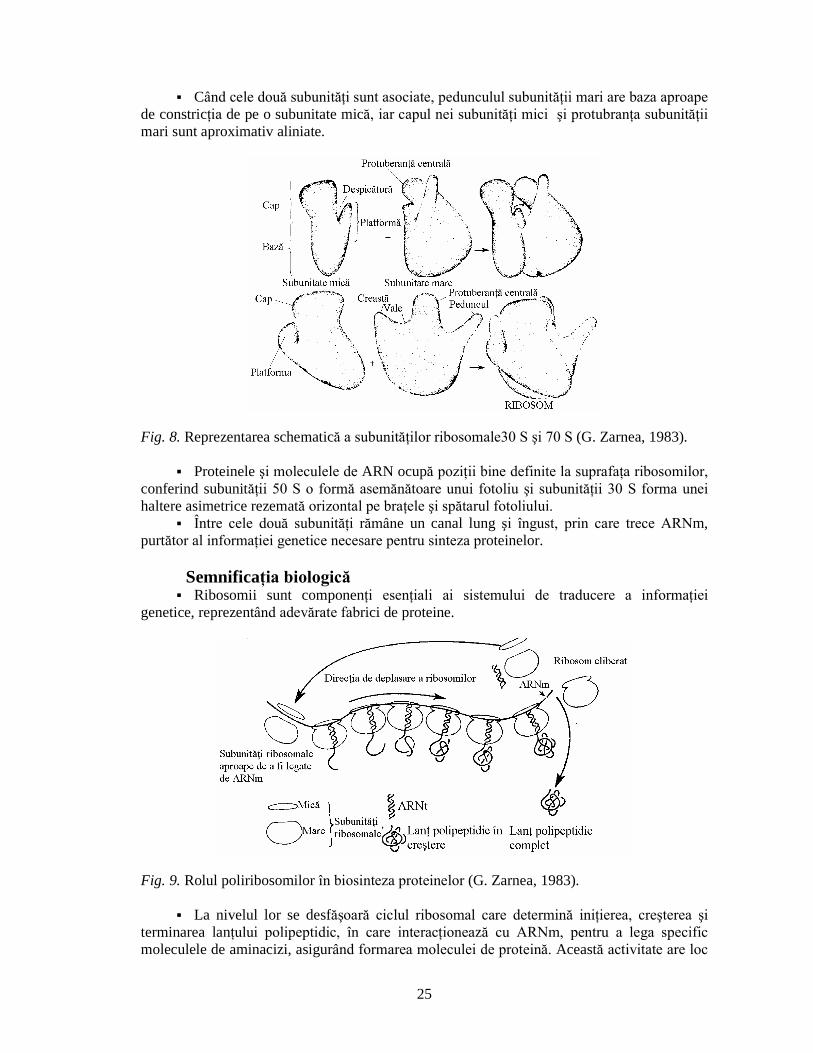
25
Când cele douã subunitãþi sunt asociate, pedunculul subunitãþii mari are baza aproape
de constricþia de pe o subunitate micã, iar capul nei subunitãþi mici ºi protubranþa subunitãþii
mari sunt aproximativ aliniate.
Fig. 8. Reprezentarea schematicã a subunitãþilor ribosomale30 S ºi 70 S (G. Zarnea, 1983).
Proteinele ºi moleculele de ARN ocupã poziþii bine definite la suprafaþa ribosomilor,
conferind subunitãþii 50 S o formã asemãnãtoare unui fotoliu ºi subunitãþii 30 S forma unei
haltere asimetrice rezematã orizontal pe braþele ºi spãtarul fotoliului. Între cele douã subunitãþi rãmâne un canal lung ºi îngust, prin care trece ARNm,
purtãtor al informaþiei genetice necesare pentru sinteza proteinelor.
Semnificaþia biologicã Ribosomii sunt componenþi esenþiali ai sistemului de traducere a informaþiei
genetice, reprezentând adevãrate fabrici de proteine.
Fig. 9. Rolul poliribosomilor în biosinteza proteinelor (G. Zarnea, 1983).
La nivelul lor se desfãºoarã ciclul ribosomal care determinã iniþierea, creºterea ºi
terminarea lanþului polipeptidic, în care interacþioneazã cu ARNm, pentru a lega specific
moleculele de aminacizi, asigurând formarea moleculei de proteinã. Aceastã activitate are loc

26
numai când cele douã subunitãþi sunt asociate ºi combinate cu ARNm, ca ºi cu alþi
componenþi neribosomali ca ARNt ºi unele proteine. În cursul procesului de biosintezã a proteinelor ribosomii individuali au tendinþa de a
se grupa în ºiruri lineare de 4-50 de elemente formând polisomi (poliribosomi sau ergosomi). Mãrimea acestora este determinatã de lungimea moleculelor de ARNm.
Poliribosomii se deplaseazã de-a lungul moleculei de ARNm, permiþâmd ca lanþil
polipeptidic sã creascã progresiv în lungime, pe mãsurã ce ribosomii se deplaseazã spre
extremitatea terminalã a secvenþei de baze traduse, fapt care conferã o eficienþã mult mai mare procesului de biosintezã a proteinelor.

27
4. METABOLISMUL MICROORGANISMELOR
Metabolismul microorganismelor reprezintã totalitatea reacþiilor biochimice
implicate în activitatea lor biologicã, prin intermediul cãrora energia ºi elementele biogene
sunt preluate din mediu ºi utilizate pentru biosintezã, degradare ºi creºtere, ca ºi pentru
activitãþi fiziologice secundare (mobilitate, luminescenþã etc.). Ca urmare a acestor reacþii
substanþele din mediu sunt transformate în constituenþi celulari, energie ºi produºi de
metabolism. Diferitele reacþii ale metabolismului îndeplinesc patru funcþii esenþiale pentru viaþa
celulei: eliberarea de energie ºi stocarea ei sub formã de ATP ºi alþi compuºi
macroergici. producerea subunitãþilor folosite pentru construcþia constituenþilor celulari,
pornind de la substanþele nutritive. activarea subunitãþilor de construcþie monomere, pe seama energiei de legãturã
din compuºii macroergici ºi formarea constituenþilor celulari macromoleculari,
prin asamblarea monomerilor. formarea ºi degradarea biomoleculelor necesare anumitor funcþii specializate
ale celulei. Aceste funcþii sunt realizate printr-o reþea complexã ºi variatã de cãi metabolice, care
funcþioneazã perfect coordonat datoritã intervenþiei unor reacþii speciale (�pace marker
reactions�, Krebs ºi Kornberg, 1957). Cãile metabolice sunt secvenþe de reacþii chimice, catalizate enzimatic, care permit
organismelor vii sã obþinã din nutrienþi, atât compuºii necesari pentru sinteza constituenþilor
celulari, cât ºi energia necesarã pentru aceste sinteze, ca ºi pentru alte procese care necesitã
consum de energie. Au fost descrise patru tipuri de cãi metabolice:
cãile catabolice; cãile anabolice; cãile amfibolice; cãile anaplerotice.
4.1. Cãile catabolice
Procesele biochimice implicate în degradarea diferiþilor compuºi nutritivi din mediu
ºi eliberarea de energie în celulã sunt reunite sub denumirea de catabolism, iar reacþiile
enzimatice implicate în degradare sunt numite reacþii catabolice. Ca regulã generalã procesele de catabolism evolueazã în trei faze succesive
(Kornberg, 1965): Faza I corespunde degradãrii macromoleculelor la unitãþile mici constitutive.
Ex.: proteinele sunt degradate la aminoacizi, glucidele la hexoze, lipidele la glicerol ºi acizi graºi.
Se elibereazã ~1% din energia totalã a macromoleculelor, care se pierde sub formã de
cãldurã. Faza II corespunde etapei în care diferitele micromolecule rezultate în faza precedentã
sunt degradate incomplet, eliberând ~1/3 din energia totalã cu producerea � în afarã de CO2 ºi H2O � a unui numãr relativ mic de produºi diferiþi, numiþi intermediari metabolici ai cãilor
metabolice centrale.

28
Ex.: hexozele pot fi metabolizate pe cãi diferite (calea glicolizei, calea Entner-Doudoroff sau cale hexozomonofosfatului), dar produºii obþinuþi
final nu sunt foarte diferiþi, reprezentând, cu excepþia CO2, intermediari ai cãilor metabolice centrale sau precursori ai lor.
Aminoacizii pot fi utilizaþi pe cãi diferite, dar catabolismul lor duce la formarea de acetil-CoA sau intermediari ai ciclului acizilor tricarboxilici (ciclul Krebs).
Caracteristica esenþialã a acestei faze decurge din faptul cã deºi existã o mare varietate
de cãi catabolice, acestea conduc transformarea surselor primare de carbon la un numãr redus
(~12) de intermediari ai cãilor metabolice centrale. Faza a III-a are o evoluþie diferitã în funcþie de natura microorganismului studiat. În cazul microorganismelor aerobe, care pot metaboliza substraturile nutritive integral
(pânã la CO2 ºi H2O), calea majorã de desfãºurare a acestei faze ºi eliberare de energie este
cea a ciclului acizilor tricarboxilici (ciclul Krebs), cuplat cu fosforilarea oxidativã. Microorganismele care nu pot efectua conversia completã a surselor de C la CO2 ºi
H2O realizeazã aceastã fazã prin reacþii de fermentaþie (alcoolicã, lacticã, butiricã, propionicã,
acetonobutilicã etc.), în care produºii cãilor catabolice servesc � direct sau indirect - ca donori, respectiv acceptori de H, în secvenþe de reacþii de oxidoreducere cuplate. În aceste
cazuri, cantitatea de energie utilã pentru celulã este mult mai scãzutã, în raport cu cea
eliberatã în cursul respiraþiei aerobe, prin mecanismul fosforilãrii oxidative (Kornberg, 1965).
4.2. Cãile anabolice
Procesele de asimilare, corespunzând anabolismului, se realizeazã prin secvenþe de
reacþii enzimatice, care asigurã folosirea intermediarilor cãilor centrale ale metabolismului
pentru sinteza blocurilor de construcþie, componente ale macromoleculelor. Cãile anabolice au ca punct de plecare, de multe ori, intermediari ai cãilor centrale, foarte diferiþi de cei care
rezultã din catabolismul compusului respectiv. Cãile catabolice ºi anabolice pentru douã substraturi chimice date nu sunt complet
reversibile ºi sunt numai parþial comune. Spre exemplu, degradarea glicogenului la acid lactic
este catalizatã de 12 enzime, dintre care numai 9 catalizeazã reacþii reversibile; pentru
celelalte trei cãi, din considerente energetice, se folosesc cãi ocolite, care implicã enzime
diferite ºi intermediari diferiþi. Ca ºi procesele de catabolism, reacþiile de biosintezã, corespunzând reacþiilor
anabolice, se realizeazã tot în trei faze succesive, prin utilizarea micromoleculelor precursoare
produse în faza a III-a a catabolismului ºi conversia lor la molecule mai mari � blocuri de construcþie (faza a II-a), care în final sunt asamblate în macromolecule de proteine, glucide ºi
lipide. Spre deosebire de cãile catabolice, ale cãror începuturi sunt multiple, dar definite ca
punct de plecare (pornind de la mai multe proteine, glucide ºi lipide diferite), pentru ca sã
evolueze convergent spre anumite cãi finale comune, cãile anabolismului au ca punct de
plecare un numãr foarte limitat de molecule precursoare simple, de la care evolueazã divergent ºi ramificat pentru a realiza gama largã de molecule ce caracterizeazã celulele vii.
4.3. Cãile amfibolice
Denumirea genericã de cãi amfibolice (gr. amfi = douã) pentru a desemna cãile
metabolice centrale, care îndeplinesc, în acelaºi timp, funcþia de eliberare de energie ºi de
furnizare a unor precursori pentru biosinteze, a fost propusã de Davis, în 1961.

29
Cãile catabolice ºi cele anabolice funcþioneazã simultan în celule, sub forma unor
secvenþe de reacþii catalizate de enzime ºi însoþite, la fiecare treaptã, de o modificare
caracteristicã, energeticã. În felul acesta, de exemplu, între catabolismul unui substrat organic
ºi procesele de biosintezã se stabilesc, la modul general, douã legãturi biochimice distincte: 1. În anumite trepte specifice ale cãilor catabolice se elibereazã o parte din energia
chimicã a metaboliþilor pentru a fi conservatã sub formã de ATP, în timp ce în anumite
trepte ale anabolismului, energia din ATP este utilizatã pentru biosinteze. 2. Produºii cãilor de catabolism funcþioneazã nu numai ca intermediari în eliberarea
treptatã de energie, ci sunt punctul de plecare al unor reacþii de biosintezã în care
reprezintã precursori ai componenþilor celulari. În felul acesta, calea de catabolism al
unui substrat organic funcþioneazã, cel puþin parþial, ca o cale de iniþiere a unor reacþii
de biosintezã. Ciclul Krebs, spere exemplu, reprezintã nu numai o cale de oxidare totalã a acetil
CoA, ci, în acelaºi timp, ºi o sursã importantã de intermediari (á-cetoglutarat, oxalilacetat, succinat) pentru cãile de biosintezã. Datoritã acestui fapt, el nu funcþioneazã, niciodatã, în
ciclu închis, deoarece anumiþi intermediari trec direct în cãile de biosintezã.
4.4. Cãile anapleurotice
Deoarece intermediarii cãilor centrale amfibolice sunt în permanenþã îndepãrtaþi, pe
de o parte, în cursul fazei a III-a a catabolismului, cu eliberare de energie, ºi, pe de altã parte,
prin utilizarea lor în diferite biosinteze, funcþionarea multor cicluri este condiþionatã de
asigurarea unei reaprovizionãri permanente cu intermediarii utilizaþi, încetul cu încetul, pe
parcursul desfãºurãrii lor. Aceastã funcþie este asiguratã de o serie de cãi metabolice auxiliare
� cãile anapleurotice � diferite de cele ale catabolismului, reprezentate de una sau mai multe etape catalizate enzimatic, care asigurã completarea acestui deficit prin introducerea unor
metaboliþi în anumite cicluri metabolice esenþiale. Funcþionarea ºi interacþiunile celor patru tipuri de cãi metabolice au loc în mod
perfect coordonat, datoritã mecanismelor complexe care asigurã reglarea activitãþii lor, ºi
adaptat la nevoile microorganismelor, determinate, în mare mãsurã, de compoziþia mediului
în care trãiesc. Evoluþia cãilor metabolice sub forma unor reacþii secvenþiale multiple conferã
metabolismului o mai mare flexibilitate ºi diversitate, asigurând, în acelaºi timp, eliberarea
treptatã a energiei ºi efectuarea interconexiunilor cu alte reacþii ale metabolismului ºi în
special cu cele de biosintezã.
4.5. Particularitãþile generale ale metabolismului microorganismelor
Natura ºi diversitatea nutrienþilor folosiþi. Considerate în ansamblu,
microorganismele sunt organismele cele mai tipic omnivore dintre toate cele cunoscute, deoarece îºi pot realiza metabolismul folosind foarte numeroase ºi diferite surse de substanþe
nutritive, de la N molecular, CO2, So pânã la substanþele organice complexe. Teoretic, aproape
orice substanþã din mediu, organicã sau anorganicã, din care se poate obþine energie, este
accesibilã metabolismului bacterian. În lumea bacteriilor existã însã ºi mari diferenþe individuale sub raportul potenþialului
biochimic în sensul cã, unele specii, ca de exemplu cele din genul Pseudomonas pot utiliza ca sursã de C ºi energie 70-200 de substanþe chimice diferite, în timp ce altele sunt foarte �specializate�, utilizând un numãr foarte limitat de nutrienþi. Astfel, bacteriile metilotrofe
utilizeazã numai metanul ºi metanolul, bacteriile celulozolitice, numai celuloza, Nitrosomonas numai NH3 ºi O2, iar Bacillus fastidiosus numai acidul uric ºi compuºii purinici înrudiþi.

30
Bacteriile sulfatreducãtoare (Desulfovibrio desulfuricans) nu utilizeazã ca sursã de C decât un
numãr limitat de compuºi organici (lactat, piruvat ºi acizi dicarboxilici cu C4) ºi spre
deosebire de majoritatea bacteriilor, nu metabolizeazã nici un glucid ºi nici un aminoacid, cu
excepþia cisteinei. Diferitele grupuri sistematice de bacterii se caracterizeazã deci prin anumite
particularitãþi metabolice, în aºa fel încât, în afara unor substanþe folosite de toate
microorganismele (apã, CO2, sãruri minerale etc.), unele specii sau grupuri fiziologice de
specii au nevoie ºi de anumite substanþe nutritive specifice, indispensabile pentru realizarea
metabolismului celular. �Plasticitatea� metabolismului microorganismelor este o altã particularitate,
decurgând din capacitatea acestora de a se adapta la tipul ºi cantitatea de nutrienþi diferiþi,
prezenþi în mediu. E. coli, de exemplu, utilizeazã nu numai glucoza ºi NH3 ca sursã de C ºi N,
ci ºi o gamã largã de nutrienþi, incluzând diferite alte glucide, glicerol, etanol, acetat etc.,
respectiv purine, pirimidine, aminoacizi, datoritã echipamentului enzimatic complex, care
asigurã conversia tuturor acestor substanþe în compuºi care pot intra în cãile metabolice
centrale. Diversitatea mecanismelor enzimatice ºi a produºilor. Microorganismele pot
utiliza acelaºi substrat nutritiv pe mai multe cãi distincte sau parþial intricate, fiecare dintre ele
putând sã ducã, în funcþie de natura microorganismului, la formarea de produºi diferiþi. Diferitele cãi metabolice pot coexista în acelaºi microorganism ºi se pot substitui una
celeilalte, în funcþie de condiþiile fiziologice ºi/sau de mediu. Intensitatea metabolismului bacterian. Una din caracteristicile distinctive ale
activitãþilor metabolice microbiene este intensitatea lor excepþionalã comparativ cu aceea a
activitãþilor omologe ale organismelor superioare. Astfel, activitatea respiratorie a unui gram
(raportat la greutatea uscatã) de bacterii aerobe este de câteva sute de ori mai intensã decât
aceea a omului, iar potenþialul metabolic al microorganismelor din 25 cm superficiali ai solului, pe o suprafaþã de un hectar, este echivalent cu cel la câtorva zeci de mii de oameni.
Un gram de bacterii lactice hidrolizeazã circa 178-14 890 g lactozã pe orã. Pentru a atinge un
asemenea nivel de activitate metabolicã organismul uman ar avea nevoie de mai multe mii de
tone de alimente pe orã. O celulã de E. coli, care se divide la un interval de o orã, sintetizeazã pe secundã 4 000
de molecule de lipide, aproximativ 1 000 tipuri de proteine (fiecare având 300 de aminoacizi)
ºi 4 molecule de ARN (Nester, 1973).

31
5. NUTRIÞIA MICROORGANISMELOR
Microorganismele, ca toate organismele vii, au nevoie pentru creºtere ºi multiplicare,
ca ºi pentru toate celelalte manifestãri ale activitãþii lor biologice, de prezenþa în mediul
înconjurãtor a unor substanþe nutritive, care sã conþinã, pe de o parte, elementele chimice
necesare pentru sinteza constituenþilor celulari, pentru activitatea enzimelor ºi sistemelor de
transport, iar, pe de altã parte, sã le furnizeze substanþele necesare pentru producerea de
energie biologic utilã. Din cauza caracterului foarte heterogen al mediilor naturale ºi dificultãþilor de a pune
la punct tehnici de identificare ºi dozare a diferitelor substanþe prezente în jurul celulelor
bacteriene se cunosc foarte puþine date referitoare la procesele lor de nutriþie în naturã. Mediile de culturã utilizate în practicã conþin, într-o formã adecvatã tipului de
nutriþie al fiecãrui microorganism, nutrienþii esenþiali pentru sinteza de material celular ºi
pentru producerea de energie in vitro, care condiþioneazã procesul de creºtere ºi multiplicare.
Ele includ în mod obligatoriu, pe lângã o serie de ioni anorganici, diferiþi compuºi ai C, H, O,
N, P, S etc.
5.1. Bioelementele
Microorganismele au nevoie pentru creºtere de aceleaºi bioelemente întâlnite în
structura oricãrui sistem biologic. Bioelementele majore (macroelementele) ºi anume C, O, H, S, P, K, Mg, Ca ºi Fe,
sunt necesare în concentraþii relativ mari (> 10-4 M).
Dintre acestea C, O, H, N, S ºi P formeazã ~95% din greutatea celularã uscatã a
bacteriilor ºi reprezintã, ca atare, constituenþii majori ai biomasei celulare a
microorganismelor. Macroelementele îndeplinesc funcþii cu semnificaþie esenþialã în metabolism ºi, în
general, în viaþa microorganismelor. Astfel, sulful este necesar pentru sinteza unor aminoacizi (cisteinã ºi metioninã) ºi a
unui numãr de coenzime; fosforul intrã în structura acizilor nucleici, a fosfolipidelor, a acizilor teichoici, precum ºi în structura unor nucleotide ca ATP, GTP, NAD
+ ºi FAD. K, Mg, Ca, ºi Fe sunt ioni metalici necesari pentru activitatea enzimelor sau intrã, ca
elemente, în compoziþia complexelor metalice. Spre exemplu, fosfolipidele peretelui celular bacterian sunt chelate cu ioni de magneziu. Calciul intrã în structura unor coenzime; ionii
feros ºi feric fac parte din structura unor purtãtori redox, cum sunt citocromii ºi proteinele
FeS. Bioelementele minore (microelementele sau micronutrienþii, �trace elements�)
Zn, Mn, Na, Cl, Mo, Se, Co, Cu, W, ºi Ni sunt necesare în cantitãþi foarte mici, unele (Zn ºi
Mn) ca elemente esenþiale pentru toate microorganismele, în timp ce altele (Se, Mo, Co, Cu,
ºi W) numai pentru unele activitãþi metabolice speciale: zincul este necesar pentru cã ADN-polimerazele ºi ARN-polimerazele sunt
metaloproteide cu zinc; molibdenul intrã în constituþia nitrogenazei, enzimã esenþialã pentru procesul de
fixare biologicã a azotului; cobaltul participã la efectuarea reacþiilor metabolice dependente de vitamina B12; cuprul intrã în structura unor enzime care transferã electroni de la nivelul unui
substrat la oxigen. Nevoia de ioni minerali variazã cantitativ ºi calitativ de la un microorganism la altul, în
primul rând, datoritã diferenþelor de constituþie enzimaticã ºi de eficacitatea mecanismelor de
concentrare activã a lor din mediu.

32
5.2. Sursa de carbon
În funcþie de natura sursei de carbon folositã pentru creºtere ºi multiplicare,
microorganismele formeazã douã mari grupuri: microorganismele autotrofe, care folosesc CO2 ca unicã sau ca principalã
sursã de C celular; microorganismele heterotrofe sau organotrofe, la care substanþele organice
servesc, în acelaºi timp, ca sursã de C pentru biosinteze ºi producere de energie.
Marea varietate a tipurilor nutriþionale întâlnite la microorganisme nu poate fi însã
exprimatã numai în funcþie de sursei de C. Se iau în considerare criterii suplimentare ca:
natura sursei de energie; natura substanþelor folosite ca donatori de hidrogen (sau de electroni).
În cazul microorganismelor care folosesc diferite substanþe organice, rolul de sursã
de energie este asigurat prin faptul cã oxidarea sau dezasimilarea lor se face cu eliberare de
energie utilã pentru celulã ºi eliberarea unei mari varietãþi de produºi de metabolism, de
exemplu CO2 (produsul major al metabolismului respirator) sau ca un amestec de CO2 ºi compuºi organici (în cazul metabolismului fermentativ). În acelaºi timp, compuºii organici
utilizaþi ca nutrienþi sunt folosiþi ca punct de plecare pentru sinteza celor câteva mii de
substanþe organice constitutive, care sunt încorporate prin reacþii consumatoare de energie în
structura celulelor. Existã mari diferenþe în ceea ce priveºte natura ºi numãrul compuºilor care pot fi
folosiþi ca sursã de C. Nutriþia heterotrofã poate fi:
simplã, ca la E. coli, care poate fi cultivatã pe medii de culturã ce conþin numai
glucozã ºi sãruri minerale; complexã, ca la Lactobacillus sp., care are nevoie de o serie de compuºi
organici, inclusiv vitamine. Heterotrofia implicã incapacitatea de sintezã a unor metaboliþi esenþiali ºi necesitatea de
a-i gãsi ca substanþe preformate în mediu, spre deosebire de autotrofe, care sunt capabile sã
sintetizeze constituenþi organici, pornind de la compuºi anorganici simpli. Unele microorganisme heterotrofe au nevoi foarte specifice ºi nu se pot dezvolta
decât în prezenþa unui singur compus organic: Bacillus fastidiosus utilizeazã numai acidul uric; microorganismele celulozolitice utilizeazã numai celuloza; bacteriile metilotrofe folosesc numai metanul, metanolul ºi dimetileterii.
Alte microorganisme, ca, de exemplu, bacteriile din genul Pseudomonas, folosesc o gamã foarte largã, de peste 100 de compuºi organici cu C, pe care nu sunt capabile sã îi
sintetizeze (carbohidraþi, polialcooli, acizi graºi, acizi aminaþi, hidrocarburi, acizi aromatici etc.). Este probabil cã toate microorganismele heterotrofe au nevoie ºi de CO2 în cantitãþi
foarte mici pentru diferite reacþii biosintetice. Dovada o constituie faptul cã îndepãrtarea
completã a CO2 împiedicã sau inhibã creºterea microorganismelor respective în medii
organice. Cel mai adesea, nevoile de CO2 sunt acoperite de însuºi metabolismul normal al
substanþelor organice, care produce cantitãþi importante de CO2. Existã ºi excepþii,
reprezentate de unele bacterii ºi fungi microscopici a cãror dezvoltare heterotrofã este
condiþionatã de prezenþa unei atmosfere bogate în CO2 (5-10%).

33
5.3. Sursa de azot
Azotul este necesar în cantitãþi mari deoarece reprezintã ~10% din greutatea uscatã a
celulei bacteriene, în care este încorporat în general, sub formã redusã, ca grupãri �NH2. Azotul este prezent în mod natural ca N2 sau în diferite combinaþii ca NH3, nitrat, nitrit sau compuºi organici. Sursa preferatã de N pare sã fie NH3, care este utilizat practic de toate microorganismele.
În cazul microorganismelor fixatoare de N2, acesta este redus iniþial la NH3, înainte
de a fi încorporat în compuºi organici. Î mod similar, microorganismele care folosesc nitraþii
sau cele care utilizeazã N organic îi convertesc iniþial la NH3, formã accesibilã pentru
biosinteze.
5.4. Factorii de creºtere
În anul 1901, Wildiers a observat cã levurile nu se pot dezvolta pe medii de culturã
sintetice, formate exclusiv din substanþe minerale ca surse de N ºi C, decât dacã li se adaugã
culturi vechi sau omorâte de levuri sau extracte de levuri, cultivate pe medii complexe sau pe
diverse produse de origine biologicã. Mediul sintetic nu satisface deci nevoile metabolice ale
levurii. Adãugarea de levuri omorâte îl îmbogãþesc cu un factor care stimuleazã creºterea.
Substanþele nedefinite prezente în aceste preparate au fost numite �factorul bios�. S-a demonstrat cã factorul bios este un amestec de vitamine, ºi aminoacizi ca:
mezoinozitol, acid pantotenic, piridoxinã, acid nicotinic, biotinã, aneurinã, â-alaninã, L-leucinã etc. Cercetãrile au demonstrat cã unele microorganisme au nevoie nu numai de surse de
energie, de C ºi de N, ci ºi de anumite substanþe organice oligodinamice, esenþiale pentru metabolismul lor, numite factori de creºtere.
Factorii de creºtere ai unui microorganism sunt acele substanþe pe care
microorganismul dat este incapabil sã la sintetizeze în cursul metabolismului sãu ºi în absenþa
cãrora multiplicarea lui este imposibilã (Lwoff, 1943). La unele microorganisme factorii de creºtere sunt de origine endogenã, fiind
elaboraþi prin biosintezã în cursul metabolismului celular, astfel cã prezenþa lor ca substanþe
preformate în mediu nu este necesarã. În schimb, microorganismele care nu au capacitatea de a sintetiza unul sau mai mulþi dintre aceºti metaboliþi esenþiali nu pot trãi decât dacã mediul
lor este suplimentat cu factorul sau factorii de creºtere pentru a cãror sintezã facultãþile lor
metabolice sunt deficitare. În funcþie de natura lor chimicã, factorii de creºtere pot fi grupaþi în trei categorii:
aminoacizi necesari pentru sinteza proteinelor; purine ºi pirimidine utilizate pentru sinteza ARN, ADN ºi a unor coenzime; vitamine care funcþioneazã ca grupãri prostetice ale unor enzime, ale unor
proteine-purtãtor sau cu funcþie de coenzime. În raport cu celelalte categorii de nutrienþi, care folosesc pentru formarea
constituenþilor celulari, factorii de creºtere sunt necesari în cantitãþi mici, deoarece cei mai
mulþi îndeplinesc funcþii specifice în biosinteze.
5.5. Oxigenul
Oxigenul este furnizat microorganismelor, în primul rând, de diferiþi nutrienþi ºi de
apa din constituþia mediilor de culturã. Din cauza solubilitãþii sale scãzute, oxigenul aflat în
soluþie este utilizat foarte repede de bacteriile aerobe, chiar în cazul unor populaþii puþin
numeroase. De aceea, frecvent, densitatea atinsã de o culturã este adesea limitatã de rata de
difuzie a O2 prin interfaþa aer-apã, chiar când culturile sunt bine aerate.

34
În culturile aerate prin simpla rotaþie a flacoanelor creºterea unor bacterii aerobe este
adesea limitatã la ~1-2 mg greutate uscatã/ml. Creºterea mai abundentã necesitã tehnici speciale, care mãresc aria interfeþei lichid-
aer, ca, de exemplu, barbotarea rapidã cu aer printr-un filtru poros, sau prin recircularea culturii. Pentru necesitãþi de ordin practic, pe baza efectului pe care oxigenul îl exercitã
asupra creºterii ºi metabolismului, microorganismele pot fi grupate în patru categorii: 1. Microorganismele strict sau obligatoriu aerobe (de exemplu, Bacillus anthracis, B.
subtilis, Mycobacterium tuberculosis), care au nevoie absolutã de prezenþa oxigenului
molecular ºi sunt incapabile sã trãiascã în anaerobiozã. 2. Microorganismele strict sau obligatoriu anaerobe (de exemplu, Clostridium tetani, C.
botulinum, Welchia perfringens etc.), care nu se pot dezvolta în prezenþa oxigenului molecular
ºi, ca urmare, pot fi cultivate numai în medii sãrãcite în O2, deoarece chiar la presiuni joase, de ordinul a 10-5 atm, pot avea efect inhibitor.
În practicã, se folosesc înglobãri în medii ale unor substanþe care favorizeazã creºterea
bacteriilor anaerobe, de exemplu tioglicolatul de sodiu 0,1%, care reduce O2 din mediu. Alte procedee: adãugarea unui strat de ulei de parafinã; utilizarea de agar semisolid (0,2-
0,3%), pentru a diminua difuzia O2; cultivarea în recipiente (anaerostate) din care oxigenul a
fost îndepãrtat sau înlocuit cu un amestec controlat de alte gaze. 3. Microorganismele microaerofile (de exemplu, Spirochaetales, Thiobacteriales etc.)
cresc cel mai bine la presiuni de oxigen mai mici (~0,2 atm) decât în aerul atmosferic.
Aceastã particularitate reflectã prezenþa unor enzime care sunt inactivate în condiþii de
oxidare puternicã ºi pot fi menþinute în stare funcþionalã numai la presiuni joase de O2. 4. Microorganismele anaerobe, facultativ aerobe (E. coli, Staphylococcus, Streptococcus
etc.) au capacitatea de a se dezvolta deopotrivã în prezenþa sau în absenþa O2. unele dintre ele (de exemplu bacteriile lactice) desfãºoarã în esenþã un metabolism de tip fermentativ, chiar în
prezenþa aerului, fãrã a fi sensibile la O2. Altele, cum sunt levurile, E. coli ºi Enterobacteriaceae-le, în general, sunt capabile sã-ºi orienteze metabolismul spre respiraþie
sau spre fermentaþie, în funcþie de condiþiile de mediu, respectiv de disponibilitãþile de O2. În esenþã nevoia de O2 reflectã mecanismul molecular prin care microorganismele îºi
satisfac nevoile energetice.

35
6. AUTOTROFIA
În concepþia clasicã, autotrofia reprezintã tipul de metabolism caracteristic organismelor capabile sã creascã în absenþa oricãrui compus organic. Microorganismele
autotrofe utilizeazã CO2 ca sursã unicã sau majorã de carbon celular ºi N sub formã de NH3, nitrat sau chiar N molecular, diferite sãruri minerale ºi apa, de la care îºi construiesc
numeroasele substanþe organice caracteristice organismelor vii. Microorganismele autotrofe
folosesc aceeaºi sursã de C, adicã CO2. Ele se deosebesc, în raport cu natura sursei de energie
care asigurã creºterea, putând fi diferenþiate în: o fotoautotrofe, care pot obþine toatã energia necesarã pentru creºtere de la luminã,
oxidând compuºi anorganici compuºi ai sulfului sau So;
o chimioautotrofe, care îi obþin energia necesarã din oxidarea unor compui anorganici reduºi ca NH3, nitriþi, compuºi ai S, Fe sau H2. Sub raportul capacitãþii lor de biosintezã, autotrofele pot fi considerate ca microorganisme
complete, deoarece posedã sistemele enzimatice cele mai complexe, care asigurã producerea
tuturor componenþilor celulari de la CO2, pe seama oxidãrii unor substanþe anorganice. Au fost descrise urmãtoarele categorii de microorganisme autotrofe:
Microorganisme autotrofe obligate, care fixeazã CO2 ca principalã sursã de C, pe
calea Benson-Calvin (calea ribulozodifosfatului), utilizând o substanþã anorganicã redusã ca
sursã de energie, sau, dacã sunt fototrofe, ca donator de electroni. Ele sunt de douã tipuri:
1. microorganisme chemolitotrofe obligate, obþin toatã energia necesarã pentru
creºtere ºi pentru asimilarea C prin oxidarea specificã (în absenþa luminii) a unor
compuºi anorganici ai S, ai ionilor NH4+, nitrit ºi fier feros sau a H molecular;
2. microorganisme fotolitotrofe obligate, utilizeazã energia luminoasã, asociatã
cu oxidarea compuºilor reduºi ai S sau H2. Microorganisme autotrofe facultative, care au o mult mai mare adaptabilitate
metabolicã, incluzând specii (Acidithiobacillus novelus, Hydrogenomonas etc.) ale cãror
particularitãþi fiziologice merg de la capacitatea de a se dezvolta autotrof, pânã la cea de
creºtere heterotrofã. Chemolitotrofele facultative pot creºte fie autotrof (în medii cu sãruri
minerale, utilizând substanþe anorganice ca sursã de C ºi energie), fie heterotrof, folosind
substanþe organice.
Orientãri noi asupra conceptului de autotrofie Iniþial definiþia autotrofelor a fost legatã numai de metabolismul carbonului,
caracterizând organismele care folosesc energia obþinutã prin oxidarea unor donatori
anorganici de electroni pentru a fixa CO2, ca primã ºi indispensabilã sursã de C. Schlägel (1975) a considerat cel dintâi definiþia autotrofiei legatã de utilizarea CO2
ca sursã majorã obligatorie de C, ca depãºitã. Dupã Schlägel, definitoriu pentru autotrofe este
faptul cã un organism este capabil sã sintetizeze substanþele celulare de la CO2, ca sursã principalã de C, deoarece faptul cã unele autotrofe sunt facultativ heterotrofe ºi pot creºte
ocazional ºi pe substanþe organice este de importanþã minorã. Cum cele mai multe microorganisme autotrofe nu se încadreazã în conceptul riguros
clasic stabilit, s-a încercat sã se redefineascã conceptul de autotrofie. Aceste încercãri au ca
suport, printre altele, urmãtoarele argumente (Whittenbury ºi Kelly, 1977): 1. microorganismele autotrofe nu prezintã nici un mecanism comun de oxidare a
compuºilor anorganici, ceea ce demonstreazã caracterul neuniform al grupului;
diferitele substraturi sunt oxidate de enzime diferite, pe cãi diferite; 2. cele mai multe bacterii autotrofe nu sunt inhibate de anumite substanþe
organice, sau nu sunt singurele organisme inhibate;

36
3. numeroase microorganisme, considerate convenþional ca heterotrofe, oxideazã
anumite substanþe anorganice, ceea ce demonstreazã cã oxidarea compuºilor
anorganici reduºi nu este o proprietate limitatã la autotrofe ºi nici un caracter-cheie obligatoriu pentru autotrofie;
4. existã bacterii care nu cresc în absenþa luminii sau a unor surse anorganice de
energie; nici una nu este însã autotrofã obligatã. Aceste date demonstreazã cã principalele categorii de autotrofe descrise în literatura
de specialitate nu acoperã toate categoriile nutriþionale care pot fi considerate ca autotrofe. Rezultã cã este preferabil sã se menþinã o separare netã între metabolismul energetic ºi
cel al carbonului, deoarece este evident cã producerea de energie de la un substrat anorganic
nu trebuie cuplatã obligatoriu cu autotrofia. În felul acesta, existã posibilitatea clasificãrii fiziologice precise: o în funcþie de sursa de energie (litotrof sau organotrof); o în funcþie de tipul de nutriþie cu C (autotrof sau heterotrof). În timp ce termenul clasic de autotrof poate implica sau nu utilizarea unui donator
anorganic de electroni, termenul chemolitoautotrof este precis ºi înseamnã organism care fixeazã CO2, utilizând energia eliberatã prin reacþii chimice din oxidarea unor donatori
anorganici de electroni. În acest context conceptul de autotrofie include:
1. organismele considerate ca autotrofe ºi în concepþia clasicã, organisme care
asimileazã CO2 pe calea circuitului Calvin; 2. organismele care folosesc alþi compuºi de tip C1 ºi calea
ribulozomonofosfatului, calea serinei sau alte cãi specifice.
Cauzele autotrofiei obligate Nu existã o explicaþie valabilã pentru toate cazurile de autotrofie. Mai mulþi factori,
acþionând separat sau în combinaþie, pot explica autotrofia unui organism sau a unui grup de
organisme: 1. Permeabilitatea limitatã faþã de nutrienþii organici. S-a demonstrat cã bacteria
Acidithiobacillus thiooxidans, adaptatã sã trãiascã într-un mediu toxic, împiedicã pãtrunderea
compuºilor organici nocivi în celulã prin proprietatea de permeabilitate selectivã. În schimb
A. denitrificans este permeabilã pentru anumite substanþe organice (glicerol), deºi nu le
folosesc în metabolism; 2. Incapacitatea de a obþine energie suficientã de la substratul heterotrof; 3. Prezenþa unei �leziuni metabolice�: adicã incapacitatea de a creºte pe nutrienþi organici
datoritã lipsei uneia sau mai multor enzime necesare pentru sinteza anumitor metaboliþi esenþiali; 4. Influenþa �patologicã� a unor nutrienþi organici exogeni: la A. neapolitanus
creºterea este inhibatã de mici cantitãþi de fenilalaninã, metioninã ºi cisteinã; 5. Autoinhibarea ºi �antrenarea� autotrofelor: la creºterea heterotrofã a fost observat cã
anumiþi nutrienþi pot fi metabolizaþi dar nu permit creºterea; de exemplu aspartatul produce compuºi toxici când este prezent în culturile de A. thiooxidans, care cresc în medii cu S ºi CO2;
6. Dependenþa de un anumit produs esenþial specific. Într-un mediu mineral aerob, suplimentat cu un substrat organic, deci în condiþii de heterotrofie, Nitrosomonas oxideazã preferenþial NH3 la nitrit, epuizând sursa de N necesarã pentru scopuri biosintetice.
Au mai fost luaþi în discuþie ºi alþi factori, cum ar fi: capacitatea limitatã de a sintetiza
compuºii necesari pentru creºtere din alte surse diferite de CO2; blocarea biochimicã a
creºterii în prezenþa unui exces de nutrienþi organici; inhibarea creºterii de cãtre anumiþi
produºi ai metabolismului compuºilor organici; dependenþa de un anumit intermediar
anorganic al procesului respirator în chemotrofie sau fototrofie, pentru anumite reacþii specifice în celulã.

37
6.1. Microorganismele chemolitoautotrofe
Bacteriile chemolitoautotrofe au douã particularitãþi de metabolism definitorii: 1. capacitatea de a obþine energie (ATP) ºi putere reducãtoare din oxidarea unor
compuºi anorganici (H2, CO, NH3, NO2-, Fe2+, So, S2O3
2- etc.); 2. capacitatea de a utiliza CO2 ca sursã unicã sau principalã de C, pe care îl reduc la
aldehidã 3-fosfoglicericã (fixarea reductivã a CO2) pentru a participa la metabolismul celular (ciclul Calvin-Barsham).
Unele sunt chemolitotrofe obligate (bacteriile care oxideazã NH3 ºi NH2-), altele sunt
facultative (hidrogenbacteriile) deoarece pot creºte ºi chemoorganotrof utilizând carbonaþi sau
acizi organici. Capacitatea de a oxida compuºi anorganici reduºi nu este limitatã doar la
chemolitotrofe. Unele heterotrofe posedã enzimele necesare pentru oxidarea H2, dar nu se pot dezvolta pe medii complet anorganice, datoritã incapacitãþii de a utiliza CO2 ca unicã sursã de
C, din lipsa enzimelor ciclului Calvin. De asemenea, unele bacterii heterotrofe pot oxida cu mai micã eficienþã compuºii reduºi
ai N (NH3, NH2-), ceea ce demonstreazã cã utilizarea acestora nu este o particularitate
exclusivã a chemolitotrofelor. În raport cu heterotrofele, microorganismele chemolitotrofe prezintã o serie de
dezavantaje legate de metabolismul energetic: o deºi realizeazã reacþii de oxidare care asigurã o conversie energeticã globalã cu o
eficienþã considerabilã, ele cresc slab ºi produc mai puþin ATP, datoritã
randamentului energetic molar relativ scãzut al substanþelor pe care le folosesc; o fixarea reductivã a CO2, principala sursã de C celular, necesitã putere reducãtoare
pentru a aduce C din CO2 la nivelul de oxidare caracteristic glucidelor; Practic, chemolitotrofele (cu excepþia celor care oxideazã hidrogenul) consumã 80% din
bugetul lor energetic total pentru conversia CO2 în constituenþi celulari.
6.2. Mixotrofia
Mixotrofia este capacitatea microorganismelor chemolitotrofe facultative de a combina utilizarea surselor de energie ºi/sau de C autotrofe, cu folosirea substraturilor organice.
Termenul de mixotrofie a fost introdus de Pfeffer (1897) pentru a descrie tipul de metabolism în care compuºii organici sunt fie necesari, fie stimulatori pentru creºterea anumitor
plante, iar Pringsheim (1967) l-a utilizat pentru a caracteriza bacteriile (de exemplu, Beggiatoa) a cãror creºtere autotrofã este mult stimulatã de prezenþa unor compuºi organici specifici.
În accepþiunea actualã, mixotrofia implicã producerea concomitentã de energie, prin
mecanisme alternative, chemolitotrofe ºi chemoorganotrofe sau utilizarea concomitentã de
mecanisme autotrofe ºi heterotrofe. Deºi nutrienþii mixotrofi nu au efect beneficial când sunt în exces, pot oferi un
avantaj deosebit în condiþii de limitare nutriþionalã. Întrucât în naturã condiþiile mixotrofe se întâlnesc foarte frecvent, microorganismele
mixotrofe au un avantaj selectiv evident faþã de heterotrofe ºi autotrofe, a cãror creºtere este
limitatã de cantitãþile insuficiente de substanþe organice, respectiv anorganice. Datoritã acestui
fapt, este probabil cã mixotrofia reprezintã modul de existenþã principal al microorganismelor
chemolitotrofe facultative în naturã. Exemplu: La Acidithiobacillus intermedius rata de creºtere este mai mare în condiþii
de mixotrofie (mediu cu tiosulfat ºi glucozã), decât în condiþii autotrofe sau heterotrofe. La
A.novellus însã nutriþia mixotrofã (mediu cu tiosulfat ºi glucozã) nu prezintã nici un avantaj,
ba chiar este stânjenitoare, dimiunuând rata de utilizare a glucozei, cât ºi rata de creºtere.

38
6.3. Sintrofia
Studiindu-se bacteria Haemophilus influenzae s-a pus în evidenþã un fenomen general:
microorganismele capabile sã-ºi sintetizeze constituenþii celulari de la compuºi simpli din
mediu excretã adesea mici cantitãþi de vitamine, aminoacizi sau alþi factori de creºtere, în afara
celulei lor, unde pot fi folosiþi de alte organisme auxotrofe pentru substanþele respective. Acest fenomen permite dezvoltarea pe medii de culturã solide, în jurul coloniilor
�excretoare� de substanþe organice, a unor colonii �satelite� formate din microorganisme mai pretenþioase (auxotrofe), care gãsesc astfel substanþele esenþiale pentru creºtere.
Exemple: H. influenzae pe gelozã-sânge formeazã colonii mici, punctiforme ºi
translucide. În vecinãtatea coloniilor de Staphylococcus, care contamineazã în mod accidental
mediul de culturã, coloniile de H. influenzae devin mai mari, mai voluminoase, deoarece stafilococul este capabil sã facã sinteza de NAD
+ pe care în excretã în mediu, permiþând
bacteriei H. influenzae sã se dezvolte mult mai abundent decât este posibil în prezenþa
cantitãþilor mici de NAD+ conþinut în sânge.
Bacteriile metanogene nu pot utiliza glucoza ca sursã de C ºi energie, dar pot creºte
uºor pe seama produºilor finali formaþi prin degradarea glucozei de cãtre bacteriile anaerobe,
capabile sã fermenteze acest compus.
6.4. Microorganismele oligotrofe
Termenul de microorganism oligotrof (sau oligocarbofil) caracterizeazã acele
microorganisme a cãror supravieþuire ºi dezvoltare ca populaþii vegetative sunt favorizate de prezenþa unor concentraþii scãzute de substanþe organice în mediu. Încadrarea în aceastã
categorie se poate face pe baza capacitãþii microorganismelor respective de a creºte în mod
evident pe medii de culturã care conþi numai 5-10 mg C/l. Microorganismele oligotrofe se deosebesc de microorganismele copitrofe, a cãror
creºtere este dependentã de o aprovizionare abundentã cu nutrienþi, în medie de o sutã de ori
mai mare decât în habitatele oligotrofe. Din cauza dificultãþilor de studiu, bacteriile oligotrofe sunt grupate provizoriu în
patru grupuri, pe baza relaþiilor lor cu substanþele organice din mediu: 1. primul grup cuprinde organisme care se pot dezvolta la prima cultivare în apa
sterilizatã a bazinului din care provin, dar care nu se dezvoltã în recultivãrile ulterioare,
nici pe medii sãrace ºi nici pe medii standard bogate. Grupul este format din bacterii
cunoscute numai pe baza morfologiei lor, studiatã la microscopul electronic; 2. grupul al doilea este reprezentat de o mare varietate de microorganisme, care nu cresc
deloc sau cresc foarte încet dacã sunt trecute din apa de testat pe medii bogate, dar care
încep sã creascã bine pe medii bogate în nutrienþi, dupã recultivarea pe apã sterilã sau
pe agar sãrac în nutrienþi; 3. grupul al treilea include bacterii cu caractere morfologice ºi fiziologice particulare, care
cresc numai pe medii speciale, uneori fiind nevoie de tehnici particulare de izolare; 4. grupul al patrulea este format din bacterii insuficient studiate ºi care nu au fost încã
cultivate pe medii artificiale. Multe dintre ele sunt prostecate, ceea ce le oferã un
avantaj adaptativ, nu numai pentru fixare, ci ºi pentru transportul nutrienþilor în celulã. Semnificaþia biologicã.
Microorganismele oligotrofe acþioneazã în naturã ca ultimi utilizatori ai substanþelor
organice care rãmân în urma activitãþii mineralizatorilor copiotrofi. Inhibarea creºterii
bacteriilor oligotrofe în medii bogate în nutrienþi pare sã fie datoratã efectului toxic al
produºilor de metabolism ºi, în special, peroxidului de hidrogen, care se acumuleazã în mediu
ºi determinã liza celulelor.

39
7. DIFUZIA ªI TRANSPORTUL SUBSTANÞELOR PRIN MEMBRANELE CELULARE
Creºterea ºi multiplicarea microorganismelor sunt condiþionate de pãtrunderea
nutrienþilor esenþiali prin înveliºurile celulare ºi de eliminare a unor substanþe rezultate din
catabolism, împiedicând însã pierderea din celulã a substanþelor necesare. Toate microorganismele au structuri-barierã ºi adevãrate porþi moleculare care asigurã integritatea
lor ºi limiteazã intrarea ºi ieºirea anumitor compuºi chimici. Datoritã acestor structuri, celula
bacterianã, spre exemplu, nu este niciodatã într-o stare de echilibru în raport cu mediul
înconjurãtor în ceea ce priveºte concentraþia diferitelor substanþe de cele douã pãrþi ale
membranelor celulare. Unele substanþe trec liber de cele douã pãrþi ale membranei, dar cele mai multe nu
pot fi transportate numai sub impulsul gradientului de concentraþie, în aºa fel încât transportul
lor este condiþionat de intervenþia unor mecanisme speciale. În funcþie de mecanismele fizicochimice care stau la baza lor, modalitãþile generale
de transport prin membranele biologice ale electroliþilor ºi neelectroliþilor sunt: difuzia pasivã,
difuzia facilitatã, translocaþia de grup, transportul activ ºi endocitoza (Zarnea, 1984).
Difuzia pasivã Se realizeazã ca urmare a unui gradient de concentraþie, posibil când în exteriorul
celulei concentraþia este superioarã celei din interiorul acesteia. Difuzia pasivã este procesul de trecere liberã a substanþelor solubile prin membrana
plasmaticã, fãrã a interacþiona cu moleculele din structura acesteia ºi fãrã consum de energie.
Moleculele cu grad ridicat de solubilitate în lipidetraverseazã mai uºor regiunea hidrofobã a
membranei. Deorece concentraþia celor mai mulþi metaboliþi este mai mare în interiorul decât în
afara celulei, difuzia pasivã este restrânsã, pe lângã apã, la un mic grup de substanþe (unele
gaze ca O2 ºi CO2, acizii graºi ºi substanþele liosolubile, anumiþi ioni). Difuzia pasivã se produce lent ºi nespecific ºi înceteazã când compusul respectiv
ajunge în aceeaºi concentraþie atât în interiorul, cât ºi în afara celulei.
Difuzia facilitatã Se realizeazã ca urmare a prezenþei în biomembrane a unor proteine receptoare
denumite permeaze, localizate la nivelul plasmalemei sau în spaþiul periplasmic. Aceastã
difuzie este stereospecificã ºi se realizeazã faþã de un gradient de concentraþie, iar celula nu
consumã energie pentru acest transfer.
Transportul activ Are urmãtoarele particularitãþi: transportul în celulã este asigurat chiar în absenþa
gradientului de concentraþie ºi se realizeazã cu consum de energie. Acest transport a fost
demonstrat în cazul drojdiilor care pot acumula intracelular o cantitate de glucide ºi de
aminoacizi mai mare decât cea existentã în mediul de culturã. Se admite cã o moleculã-transportor, care consumã o cantitate de energie pentru a se activa ºi a produce o legãturã
instabilã cu molecula nutrientului printr-o reacþie catalizatã enzimatic, transferã nutrientul ºi îl elibereazã în interior. Transportorul se activeazã ca urmare a energiei eliberate prin
transformarea ATP.
Translocaþia de grup Este un transport activ întâlnit la drojdii ºi mucegaiuri, în care intervine un sistem
enzimatic complex de transferaze ºi transfosfataze, ce permit pãtrunderea glucidelor prin
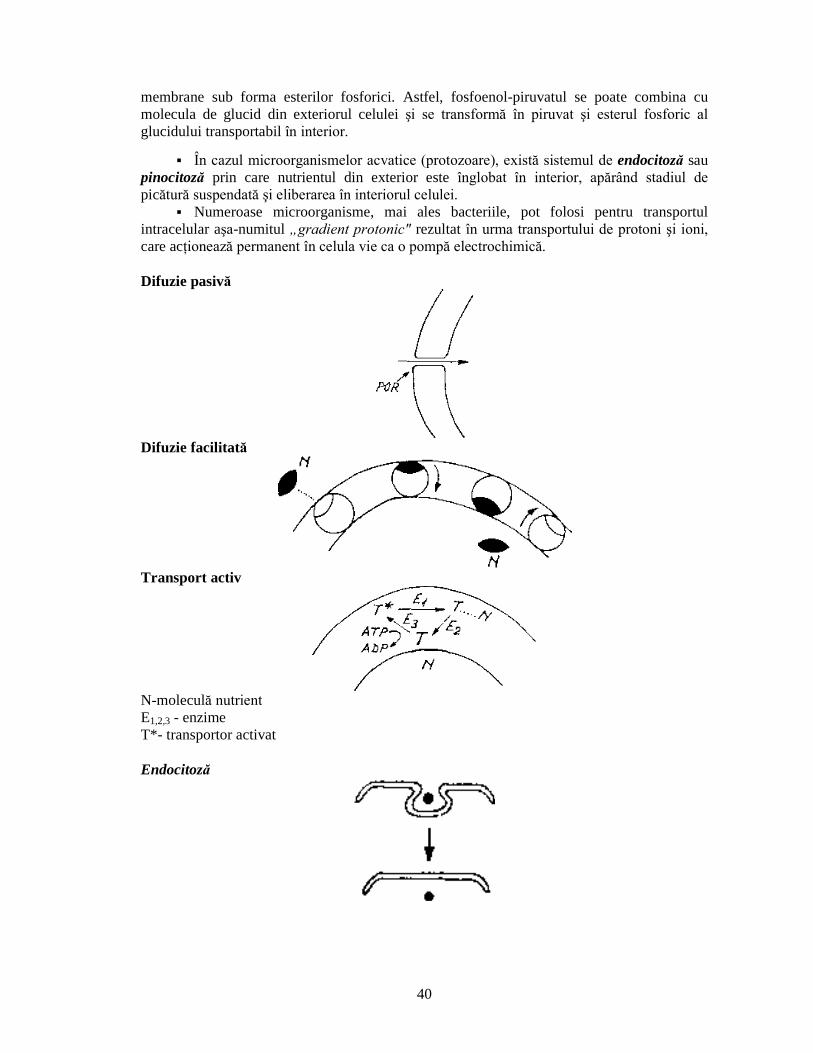
40
membrane sub forma esterilor fosforici. Astfel, fosfoenol-piruvatul se poate combina cu molecula de glucid din exteriorul celulei ºi se transformã în piruvat ºi esterul fosforic al
glucidului transportabil în interior.
În cazul microorganismelor acvatice (protozoare), existã sistemul de endocitozã sau pinocitozã prin care nutrientul din exterior este înglobat în interior, apãrând stadiul de
picãturã suspendatã ºi eliberarea în interiorul celulei. Numeroase microorganisme, mai ales bacteriile, pot folosi pentru transportul
intracelular aºa-numitul �gradient protonic" rezultat în urma transportului de protoni ºi ioni,
care acþioneazã permanent în celula vie ca o pompã electrochimicã. Difuzie pasivã
Difuzie facilitatã
Transport activ
N-moleculã nutrient E1,2,3 - enzime T*- transportor activat Endocitozã

41
8. METABOLISMUL GLUCIDELOR LA MICROORGANISME
Numeroase microorganisme utilizeazã glucidele ca sursã de energie ºi de C celular
pe calea unor secvenþe metabolice de catabolism ºi biosintezã, care uneori variazã de la o
specie la alta, astfel încât nu pot fi prezentate într-o schemã unitarã. Cea mai mare parte a rezervei de C din naturã se gãseºte sub formã de poliglucide,
care în prealabil suferã un proces de degradare sub acþiunea exoenzimelor microbiene la
molecule mici, transportabile prin membranele celulare. Astfel, celuloza, care formeazã cea
mai abundentã rezervã de material organic din naturã, reînnoibilã anual prin fotosintezã, este
degradatã de bacterii, succesiv, la molecule oligoglucidice lungi, sub acþiunea unei
endoglucanaze extracelulare, la oligoglucide scurte, de cãtre endoglucanazele periplasmice, ºi
la celobiozã ºi glucozã, de cãtre â-glucozidazele periplasmice (Ramasamy ºi Verachtert, 1980).
Sistemele enzimatice celulozolitice ale diferitelor microorganisme sunt diferite, fiind formate din enzime extracelulare ºi intracelulare sau legate de suprafaþã. În general,
degradarea celulozei are loc cel mai eficient prin contact direct între microorganisme ºi substrat, deoarece prin acest mecanism se realizeazã o concentraþie enzimaticã localã maximã
ºi o aranjare spaþialã favorabilã (Hofstern, 1975; Tor-Magnus Enari, 1983). În mod similar, amidonul � poliglucid vegetal format prin asocierea a douã specii
macromoleculare distincte, amilopectina (component major formând ~ 80 %) ºi amiloza � este degradat sub acþiunea á-amilazei (la Aspergillus niger, A. oryzae, B. subtilis, B. stearothermophilus etc.) ºi de glucoamilaze, enzime exclusiv microbiene (prezente la Aspergillus, Rhizopus, levuri ºi bacterii), la moleculele monomere componente de D-glucozã.
Utilizarea glucozei în metabolismul microorganismelor
Forma principalã în care glucidele sunt utilizate în metabolismul microorganismelor
este reprezentatã de glucozã. Procesul a fost studiat în detaliu la E. coli, care în condiþii aerobe utilizeazã ~ 50 %
din substrat, oxidându-l la CO2 (C6H12O6 + 6 O2 → 6 CO2 + H2O; ÄG0' = - 686 kcal) printr-o serie de reacþii din care rezultã suficient ATP pentru a asigura conversia restului de substrat (~ 50 %) în material celular.
Au fost descrise patru cãi principale: Calea Embden-Meyerhof-Parnas; Calea hexozomonofosfatului; Calea Entner-Doudoroff; Calea fosfocetolazei.
Primele douã sunt funcþionale deopotrivã în celulele animalelor superioare, ca ºi la
bacterii ºi fungi, în timp ce ultimele douã sunt folosite în exclusivitate de bacterii. Deºi transformãrile chimice efectuate în reacþiile acestor cãi la diferite
microorganisme sunt aceleaºi, sistemele enzimatice efectoare active la un microorganism dat sunt, adesea, întrucâtva diferite de cele care acþioneazã la alte microorganisme.
În toate aceste cãi, piruvatul ocupã poziþia unui intermediar-cheie, deoarece este situat la punctul de intersecþie metabolicã de la care pornesc diferitele cãi terminale.

42
9. METABOLISMUL ENERGETIC AL MICROORGANISMELOR
Bioenergetica microbianã (metabolismul oxidativ energetic)
Procurarea energiei este o proprietate vitalã a celulei microbiene, care se realizeazã
prin eliberarea energiei chimice a diferitelor alimente ºi formarea compuºilor macroergici prin
procesul de fosforilare. În metabolismul energetic, prin procese de oxidare ºi oxidoreducere pe cale
enzimaticã, substratul nutritiv trece în substrat oxidat, cu eliberarea de energie potenþialã (Q1). Substratul oxidat rezultat, având energia potenþialã diminuatã (Q2), suferã în continuare
transformãri pânã la produºii finali:
ATPPADPATPPADP
OHCOoxidatSubstratnutritivSubstrat
ii
21
22EE 21
Substratul oxidat poate servi drept precursor sau, dacã nu este util celulei, se poate
elimina în mediu sub formã de catabolit. Cantitatea de energie obþinutã prin catabolism poate
fi obþinutã direct în biosintezã, iar energia excedentã este stocatã în compuºi macroergici prin
procesul de fosforilare, denumit astfel deoarece componenta macroergicã, care stocheazã
energia rezultatã, conþine grupãri fosfat. Se considerã compus macroergic un compus capabil
sã elibereze o cantitate de energie mai mare sau egalã cu 30 kj/mol. Principalii compuºi macroergici sunt: ATP-ul, acetil fosfatul (Acetat~P), ADP-ul,
fosfoenolpiruvatul, compuºi care pot disponibiliza cantitãþi variabile de energie, prin ruperea
legãturii macroergice. Fosforilarea poate fi de trei tipuri:
fotosinteticã, întâlnitã la bacteriile din diviziunea Photobacteria care folosesc energia radiantã;
oxidativã, când formarea compuºilor macroeregi cu fosfor are loc prin
înmagazinarea energiei eliberate prin reacþii de oxidare, în procesele de
respiraþie aerobã sau anaerobã; de substrat, în urma formãrii de compuºi macroergici prin reacþii de oxido-
reducere, prin care glucidele sunt transformate, prin procese de fermentaþie
anaerobã, în compuºi intermediari, acceptorul de H sau electroni fiind un
compus organic. La fosforilarea oxidativã eliberarea energiei potenþiale din diverse substraturi de
naturã organicã se realizeazã prin procese de oxidare, care se pot desfãºura astfel: reacþii de oxidare prin pierdere de electroni, caz în care are loc un transfer de
electroni de la substanþe cu potenþial de oxidoreducere negativ (maxim �0,41 pentru hidrogen), la substanþe cu potenþial de oxidoreducere pozitiv (maxim
+0,82 pentru oxigen); reacþii de oxidare prin transfer de hidrogen (electroni ºi protoni): acest tip de
oxidare este folosit de unele microorganisme anaerobe ºi aerobe ºi transferul este realizat de enzime active la diferite valori ale potenþialului de
oxidoreducere. Enzimele implicate în reacþiile de oxidoreducere sunt
dehidrogenazele aerobe (aldehid-dehidrogenaza) ºi anaerobe (alcool-dehidrogenazele), care au drept coenzime NAD-ul sau NADP-ul ºi care preiau
atomii de hidrogen trecând din forma oxidatã (NAD+) în forma redusã (NADH
+ H+);

43
reacþii de oxidare prin câºtig de O2, când, în prezenþa enzimelor care
acþioneazã la un potenþial de oxidoreducere pozitiv, are loc transferul de H la O2 provenit din aer ºi se formeazã apa; aceastã reacþie de oxidare este
caracteristicã microbiotei aerobe. Respiraþia aerobã este un metabolism oxidativ dependent de oxigenul din aer,
iar produsele finale sunt CO2 ºi apa, întreaga energie a substratului oxidat eliberându-se prin produsele finale ale respiraþiei.
Respiraþia aerobã este din punct de vedere energetic foarte avantajoasã pentru celula
microbianã; de aceea, atunci când se urmãreºte obþinerea de celule în cantitãþi mari (drojdie
comprimatã) sau de substanþe intracelulare, cultivarea se face în condiþii de aerare. Microorganismele aerobe dispun de o catenã respiratorie diversificatã, în
componenþa cãreia intrã dehidrogenaze, citocromi, citocrom-oxidaze, oxidaze. Numeroase bacterii pot oxida hidrogenul (Pseudomonas), amoniacul pânã la NO2
(Nitrosomonas), sulful ºi H2S pânã la sulfit (Acidithiobacillus thiooxidans), sau Fe2+ la Fe3+ (Acidithiobacillus ferrooxidans), reacþii cu rol important în ciclul natural al elementelor.
Respiraþia anaerobã este un metabolism oxidativ în care substratul este
transformat pânã la CO2, iar electronii sunt cedaþi prin oxidare unor compuºi anorganici. Acest tip de respiraþie este întâlnit la bacteriile strict anaerobe. Astfel, bacteriile denitrificatoare pot transforma NO3 la azot molecular, bacteriile
metanogene pot transforma dioxidul de carbon în metan, bacteriile acetogene ale genului Clostridium pot transforma dioxidul de carbon în acid acetic.
Aceste bacterii obþin o cantitate micã de energie ºi pot creºte în absenþa oxigenului
molecular, la un potenþial de oxidoreducere de �0,2...-0,3 V. Þinând cont cã mediile care vin
în contact cu O2 au un potenþial redox de +0,2...+0,4 la un pH= 7, pentru a asigura dezvoltarea
anaerobilor, în mediu se adaugã substanþe cu caracter reducãtor ca: tioglicolat de sodiu,
cistein-SH, sulfurã de sodiu. Substanþele reducãtoare menþin un potenþial oxidoreducãtor
scãzut, care permite dezvoltarea anaerobilor în plãci, chiar când mediul vine în contact cu
aerul. Pentru cultivarea anaerobilor se pot folosi ºi vase speciale numite anaerostate, în care
O2 este legat chimic, sau cultura se menþine în atmosferã de gaze inerte (CO2, N2). Metabolismul oxidativ anaerob poate fi întâlnit ºi la microorganisme facultativ
anaerobe. Aceste microorganisme au capacitatea de a creºte aerob, utilizând oxigenul din aer
(respitaþie aerobã), sau anaerob, utilizând compuºi organici ca acceptori finali ai electronilor
produºi prin catabolism. În condiþii aerobe microorganismele facultativ anaerobe îºi adapteazã echipamentul
enzimatic pentru procese de oxidare pânã la produºi finali, utilizând preferenþial oxigenul,
când este disponibil, datoritã cantitãþii mai mari de energie rezultatã prin respiraþie aerobã
(36 moli ATP/mol glucozã asimilatã, faþã de 2-3 moli ATP/mol glucozã fermentatã). Celula microbianã prelucreazã nutrienþii obþinând energie ºi diferiþi compuºi, iar
produºii de catabolism sunt eliminaþi. În cursul acestor prelucrãri metabolice, în funcþie de
faza de creºtere a celulei, rezultã produºi de metabolism, care pot fi împãrþiþi astfel: produºi primari, care se formeazã în faza de creºtere exponenþialã a celulelor
(trofofaza) ºi sunt produºi esenþiali pentru celulã; produºi secundari, care sunt formaþi în faza de declin a crºterii celulelor (idiofazã) ºi
nu sunt esenþiali celulei (toxine, alcaloizi).
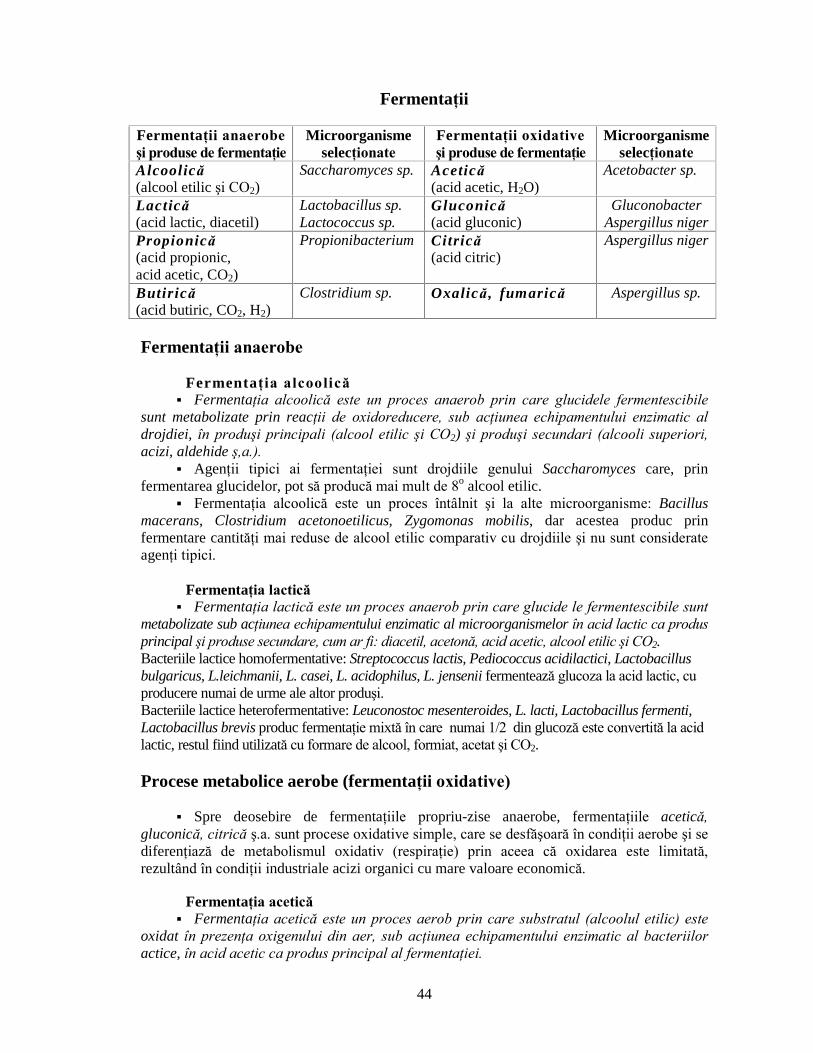
44
Fermentaþii
Fermentaþii anaerobe ºi produse de fermentaþie
Microorganisme selecþionate
Fermentaþii oxidative ºi produse de fermentaþie
Microorganisme selecþionate
Alcoolicã (alcool etilic ºi CO2)
Saccharomyces sp. Aceticã (acid acetic, H2O)
Acetobacter sp.
Lacticã (acid lactic, diacetil)
Lactobacillus sp. Lactococcus sp.
Gluconicã (acid gluconic)
Gluconobacter Aspergillus niger
Propionicã (acid propionic, acid acetic, CO2)
Propionibacterium Citricã (acid citric)
Aspergillus niger
Butiricã (acid butiric, CO2, H2)
Clostridium sp. Oxalicã , fumaricã Aspergillus sp.
Fermentaþii anaerobe
Fermentaþia alcoolicã Fermentaþia alcoolicã este un proces anaerob prin care glucidele fermentescibile
sunt metabolizate prin reacþii de oxidoreducere, sub acþiunea echipamentului enzimatic al
drojdiei, în produºi principali (alcool etilic ºi CO2) ºi produºi secundari (alcooli superiori,
acizi, aldehide º,a.). Agenþii tipici ai fermentaþiei sunt drojdiile genului Saccharomyces care, prin
fermentarea glucidelor, pot sã producã mai mult de 8o alcool etilic.
Fermentaþia alcoolicã este un proces întâlnit ºi la alte microorganisme: Bacillus macerans, Clostridium acetonoetilicus, Zygomonas mobilis, dar acestea produc prin fermentare cantitãþi mai reduse de alcool etilic comparativ cu drojdiile ºi nu sunt considerate
agenþi tipici.
Fermentaþia lacticã Fermentaþia lacticã este un proces anaerob prin care glucide le fermentescibile sunt
metabolizate sub acþiunea echipamentului enzimatic al microorganismelor în acid lactic ca produs
principal ºi produse secundare, cum ar fi: diacetil, acetonã, acid acetic, alcool etilic ºi CO2. Bacteriile lactice homofermentative: Streptococcus lactis, Pediococcus acidilactici, Lactobacillus bulgaricus, L.leichmanii, L. casei, L. acidophilus, L. jensenii fermenteazã glucoza la acid lactic, cu
producere numai de urme ale altor produºi. Bacteriile lactice heterofermentative: Leuconostoc mesenteroides, L. lacti, Lactobacillus fermenti, Lactobacillus brevis produc fermentaþie mixtã în care numai 1/2 din glucozã este convertitã la acid
lactic, restul fiind utilizatã cu formare de alcool, formiat, acetat ºi CO2. Procese metabolice aerobe (fermentaþii oxidative)
Spre deosebire de fermentaþiile propriu-zise anaerobe, fermentaþiile aceticã,
gluconicã, citricã º.a. sunt procese oxidative simple, care se desfãºoarã în condiþii aerobe ºi se
diferenþiazã de metabolismul oxidativ (respiraþie) prin aceea cã oxidarea este limitatã,
rezultând în condiþii industriale acizi organici cu mare valoare economicã.
Fermentaþia aceticã Fermentaþia aceticã este un proces aerob prin care substratul (alcoolul etilic) este
oxidat în prezenþa oxigenului din aer, sub acþiunea echipamentului enzimatic al bacteriilor
actice, în acid acetic ca produs principal al fermentaþiei.

45
Caracterele morfologice ºi fiziologice ale bacteriilor acetice. Bacteriile acetice sunt bacterii strict aerobe, sub formã de bastonaºe, gram-negative, grupate în perechi sau lanþuri,
cu dimensiuni variabile (0,5-0,8)x(80,9-4,2) ìm. Pot fi imobile sau mobile, cu cili polari sau peritrichi. În mediu acid, în timp, pot apãrea forme de involuþie, ramificate, care îºi pierd
capacitatea de reproducere. În medii lichide (staþionar) se dezvoltã sub forma unui voat fragil care, cu creºterea
în dimensiuni, ascensioneazã pe pereþii vasului (Acetobacter ascendens, A. aceti). Alte specii, A.xylinum, A. xilinoides formeazã, în vin oþeþit sau în oþet, un strat gelatinos de naturã â-glucanicã (coloidal ºi fibros).
Bacteriile acetice sunt mezofile (temperatura optimã 30 oC) ºi produc fermentaþia
aceticã într-un domeniu larg de temperaturi, 0...35 oC. Au o termorezistenþã scãzutã în mediu
lichid cu pH acid, inactivitatea lor având loc la 60 oC într-un minut, în timp ce bacteriile
reþinute pe suporturi solide (doage de lemn) sunt inactivate la temperaturi mai ridicate (100 oC).
Fermentaþia gluconicã
Fermentaþia gluconicã este un proces oxidativ simplu prin care glucoza, în prezenþa
oxigenului din aer ºi a sistemului enzimatic al microorganismelor selecþionate, este
transformatã în acid gluconic ca produs principal. Agenþii tipici ai fermentaþiei gluconice sunt bacteriile din genurile Gluconobacter
(Acetomonas) ºi Moraxella ºi mucegaiurile din genurile Aspergillus (A. niger, A. phoenicis, A.Wentii) ºi Penicillium (P. chrysogenum, P. luteum).
Fermentaþia citricã Fermentaþia citricã este un proces oxidativ complex prin care substratul glucidic
(zaharoza) este metabolizat la compuºi intermediari de oxidare, cu acumulare în mediu a acidului citric ca produs principal.
Agenþi tipici ai fermentaþiei citrice sunt tulpinile selecþionate ale speciei Aspergillus niger care produc activ citrat sintetazã. Acidul citric se poate obþine cu un bun randament (52
g/dm3) ºi prin cultivarea tulpinilor de drojdii din specia Candida oleophilla pe medii cu parafine. Transformãri microbiene ale compuºilor organici macromoleculari
Se apreciazã cã prin fotosintezã, energia radiaþiilor solare care cad anual pe pãmânt
(echivalenþã cu 3x1024 J) este convertitã în energia chimicã a combinaþiilor organice, cu
acumularea a 2x1011 tone C/an. Din aceastã biomasã vegetalã, cea mai mare parte o constituie
celuloza, substanþele pectice, amidonul ºi în cantitãþi mai reduse, lipidele, acizii nucleici º.a. În condiþii naturale dupã moartea vieþuitoarelor vegetale ºi animale, în diferitele
habitaturi (sol, apã) se produc degradãri microbiene specifice, cu formare de: produºi intermediari (condiþii de anaerobiozã); produºi finali, ca CO2 ºi H2O (condiþii de aerobiozã).
În afarã de substraturile direct fermentescibile (glucozã, zaharozã, lactozã, acizi
organici) microorganismele pot acþiona ºi asupra altor compuºi organici, fie în condiþii
dirijate, când se obþin produse cu valoare economicã, fie în condiþii naturale, când au loc
procese complexe cu rol vital în circuitul natural al elementelor (C, N, O, S, P) ºi menþinerea
vieþii pe Pãmânt.

46
Descompunerea amidonului ºi a glicogenului Descompunerea acestor polioze au loc datoritã unor microorganisme ce produc
enzime extracelulare, care produc hidroliza acestor compuºi macromoleculari la molecule
simple (glucozã, maltozã) transportabile prin membranã ºi folosite în nutriþie. Dintre enzimele microbiene care hidrolizeazã amidonul fac parte: á-amilaza, â-
amilaza ºi glucoamilaza, enzime extracelulare elaborate de bacterii, mucegaiuri ºi drojdii. Bacteriile genului Bacillus (B.subtilis, B.subtilis var. amyloliquefaciens,
B.licheniformis, B.macerans, B.sthearothermophillus sunt producãtoare de â-amilazã, enzimã
zaharogenã care, prin hidroliza legãturilor á 1-4 glucozidice, elibereazã molecule de maltozã. Mucegaiurile Aspergillus, Mucor, Rhizopus produc mai ales á-amilazã ºi
glucoamilazã. Aspergillus niger ºi A. oryzae produc á-amilaze. Mucegaiurile A.awamori, A.usami, A.niger ºi Rhizopus delemar produc
glucoamilazã. Se cunosc peste 100 de specii de drojdii care produc á-amilazã ºi glucoamilazã,
dintre care mai importante sunt cele din genurile Saccharomycopsis (S.bispora, S.fibuligera), Schwanniomyces, Trichosporon ºi Candida.
Hidroliza enzimaticã a amidonului cu enzime vegetale (din malþ) sau cu enzime
microbiene, cu formare de glucide fermentescibile, este folositã în biotehnologia spirtului, a
berii, a panificaþiei sau pentru obþinerea siropurilor dulci, a dextrinelor, a maltozei, a glucozei cristalizate º.a.
Descompunerea celulozei ºi hemicelulozei Celuloza este un poliglucid rãspândit în materiile prime de origine vegetalã
(aproximativ 1/3) ºi în structura pereþilor celulari ai fungilor. Celuloza ajunsã în sol ºi în ape
este transformatã în timp, sub acþiunea microorganismelor care produc enzime celulozolitice:
micromicete-agenþi ai putrezirii, bacterii aerobe ºi anaerobe. Produºii intermedieari de hidrolizã sunt metabolizaþi în mod diferenþiat, în funcþie de
natura microorganismelor ºi de prezenþa/absenþa oxigenului din aer. Degradarea aerobã a celulozei se caracterizeazã prin formarea din celobiozã sau
glucozã, în prima etapã, a hidroxiacizilordin care, prin oxidare, se elibereazã CO2 ºi H2O. Dintre bacteriile aerobe care asimileazã celuloza fac parte bacterii cu forme spiralate
aparþinând genurilor: Cytofaga, Cellvibrio, Cellulomonas. Importanþã industrialã o au bacteriile din genul Cellulomonas, care pot fi cultivate aerob pe deºeuri vegetale, pe hârtie º.a., iar biomasa rezultatã prin înmulþire este valoroasã prin
conþinutul ridicat în proteine (46,2%). Dintre micromicete au activitate celulozoliticã superioarã specii ale genurilor:
Botryotinia, Fusarium, Mucor, Rhizopus, Trichoderma ºi Tricothecium. Tulpini selecþionate de micromicete ale genurilor Trichoderma ºi Aspergillus sunt
bune producãtoare de celulaze ºi sunt folosite industrial ca surse de enzime. Degradarea anaerobã a celulozei are loc permanent în sedimente ºi în mâlul apelor,
la fermentarea compostului, în rumenul ierbivorelor, în profunzimea solului, în soluri
inundate, lacuri termale. În transformarea celulozei se pot distinge trei etape, în funcþie de natura produselor
de fermentaþie. Dupã hidroliza celulozei la compuºi simpli, sub acþiunea unei microbiote heterogene
alcãtuite din bacterii anaerobe ºi facultativ anaerobe din familiile Enterobacteriaceae, Bacillaceae (genul Clostridium), se acumuleazã alcooli, acizi, CO2 ºi H2.
Produsele rezultate în acest stadiu sunt folosite de bacteriile acetogene producãtoare de acid acetic pe cale anaerobã, în a doua etapã.

47
În etapa a treia, denumitã metanogenã, sunt active bacteriile metanogene strict
anaerobe din genul Methanobacterium cu speciile M.ruminantium, M.formicicum, M.mobilis, M.farkeri º.a.
Bacteriile anaerobe pot folosi acidul acetic ca sursã energeticã ºi produc metan. Se
apreciazã cã 73% din metan dezultã din acidul acetic format în faza acetogenã:
CH3-COOH ikerbarcinaMethanosar CH4 + CO2
Methanobacteriile pot forma metan prin reacþia:
4CO + 2H2O trophicumthermoautocinaMethanosar CH4 + 3CO2
În afara acestor procese naturale, care explicã formarea zãcãmintelor de metan, în
ultimele decenii a luat amploare metanogeneza dirijatã în scopul obþinerii de biogaz, prin
prelucrarea microbiologicã a dejecþiilor animale, când se realizeazã concomitent protecþia
mediului ºi valorificarea energeticã a metanului ºi a hidrogenului.

48
9. DINAMICA MULTIPLICÃRII BACTERIILOR ÎN CULTURI
În general microorganismele sunt cultivate pentru mai multe scopuri: - izolarea ºi identificarea lor; - menþinerea în stare viabilã, în colecþii, fãrã pierderea funcþiilor importante; - studiul structurii ºi funcþiei lor; - studierea rolului în naturã; - obþinerea de produºi rezultaþi din biomasa celularã, biosintezã, fermentaþii, bioconversii etc.
În prezent se cunosc puþine date în legãturã cu dinamica multiplicãrii populaþiilor
bacteriene în naturã. Procesul este însã destul de bine cunoscut raportat la multiplicarea în
condiþii experimentale, de laborator, sau industriale. Existã douã tipuri principale de culturi bacteriene:
culturi discontinue (în mediu de culturã nereînnoit), care se pot realiza în douã
variante o asincrone; o sincrone.
culturi continue, în medii de culturã reînnoite continuu.
Culturi discontinue asincrone Aceste culturi corespund cultivãrii în �sisteme închise�, în care un anumit volum de
mediu corespunzãtor este �însãmânþat� cu bacteriile respective (�batch-culture�, eng. batch =
ºarjã, lot). În aceste condiþii, creºterea este limitatã la un volum fix de mediu, care nu este reînnoit ºi care de multe ori este modificat de acumularea produºilor de metabolism, putând
deveni la un moment dat necorespunzãtor pentru creºtere. Acest mod de cultivare este folosit
frecvent în laborator, deºi are o serie de dezavantaje de care trebuie þinut seama: numãrul
bacteriilor variazã continuu, ritmul de creºtere este impus de compoziþia chimicã a mediului
care variazã de la un moment la altul, vârsta bacteriilor individuale este variabilã, numãrul de
generaþii posibile este limitat. Prin inocularea de celule, aparþinând unei culturi pure, într-un mediu nutritiv steril se
poate stabili dinamica de creºtere prin studiul vitezei de acumulare a biomasei sau prin
creºterea numãrului de celule raportat la unitatea de volum a mediului. În timp ce la
mucegaiurile inferioare, la care prin creºtere are loc prin diviziunea nucleilor ºi nu a celulei
coenocitice, creºterea se apreciazã prin determinarea masicã a biomasei formate, la alte
microorganisme, la care concomitent cu diviziunea nuclearã are loc ºi diviziunea celularã
(înmugurirea la drojdii, sciziune la bacterii, formare de pereþi despãrþitori la mucegaiurile
superioare), creºterea se apreciazã fie prin acumularea de biomasã fie prin determinarea
numãrului de celule. Pentru a aprecia dinamica multiplicãrii, la intervale regulate se recolteazã probe mici
de culturã, care sunt analizate, iar valorile logaritmice ale numãrului de celule sunt înscrise pe
curbe ale cãror modificãri aratã tranziþia de la o fazã de creºtere la alta. În general, se folosesc
logaritmi în baza 2, deoarece fiecare unitate pe ordonatã reprezintã o dublare a populaþiei
bacteriene respective. În aceste condiþii procesul evolueazã în ºase faze succesive, în funcþie
de rata de creºtere (notatã cu r sau ì): faza de lag, faza de accelerare, faza de creºtere
exponenþialã, faza de încetinire ºi faza de declin.
o Faza de lag (eng. to lag = a întârzia), numitã ºi de latenþã, sau de creºtere zero, (ì=0)
în care nu se constatã o creºtere a numãrului de celule, numãrul celulelor din inocul rãmâne
neschimbat, sau chiar scade temporar. În aceastã fazã are loc o adaptare a celulelor la
condiþiile de mediu, biosinteza de ADN/ARN ºi o activizare a sistemelor enzimatice ºi
elaborarea de enzime induse. În cazul drojdiilor, aceastã fazã poate dura 1-2 ore, duratã ce

49
depinde de compoziþia mediului ºi de capacitatea de reglare a metabolismului propriu. Durata fazei de lag poate fi redusã la limite imperceptibile în cazurile în care se foloseºte un
inoculum mare de culturã în fazã logaritmicã, iar transplantarea se face într-un mediu de culturã bogat, identic cu cel în care au fost cultivate bacteriile din inoculum.
o Faza de iniþiere a creºterii, sau faza de accelerare a ritmului de creºtere (ì ∕ ), când
are loc o creºtere în dimensiuni a celulelor prin mãrirea mai rapidã a volumului în raport cu
suprafaþa celularã, ceea ce favorizeazã declanºarea procesului de diviziune;
o Faza de creºtere logaritmicã sau de multiplicare exponenþialã, care reprezintã faza
în care creºterea masei de celule poate fi determinatã cantitativ prin dublarea numãrului de
celule în unitatea de timp (pentru drojdii ºi bacterii), sau ca dublare a biomasei / t pentru
microorganismele filamentoase (streptomicete ºi fungi). Prin exprimarea acestor valori pe o
scarã semilogaritmicã rezultã o dreaptã. Dacã unghiul pantei acestei drepte este mic aceasta se
datoreazã condiþiilor de creºtere, care nu sunt din cele mai favorabile. Deºi celulele produc o
modificare a mediului de culturã prin asimilare de compuºi nutritivi ºi eliberarea compuºilor
de catabolism, în faza logaritmicã, rata de creºtere rãmâne constantã. Rata de creºtere este independentã de concentraþia în nutrienþi atâta timp cât aceºtia se
aflã în exces. Rata de creºtere în biomasã X(g x dm-3) este corelatã cu rata specificã de
creºtere ì, în timp ce rata de creºtere a numãrului de celule este corelatã cu rata specificã de
creºtere ì ºi densitatea celulelor N. Rata specificã de creºtere ì, este dependentã de trei parametri: - concentraþia substratului limitativ S; - rata maximã de creºtere ìm; - constanta specificã dependentã de substrat Ks, conform ecuaþiei stabilitã de Jaques Monod: ì = ìm
S
Ks + S
Constanta Ks este concentraþia substratului la care se obþine ½ din rata specificã maximã
de creºtere (ì = 0,5 ìm ) ºi este echivalentã cu constanta Michaelis în cinetica enzimaticã. Dacã existã un exces al tuturor nutrienþilor, atunci ì = ìm ºi cultura se aflã deci în faza
logaritmicã la rata maximã de creºtere. Dacã unul dintre nutrienþii preferaþi, de exemplu glucoza, s-a epuizat, în prezenþa altor
surse de carbon se observã aºa-numitul fenomen de diauxie când se poate, dupã o nouã fazã
lag, sã aibã loc metabolizarea noului substrat ºi creºterea exponenþialã a celulelor. La sfârºitul fazei logaritmice se acumuleazã o cantitate maximã de biomasã, substratul
este rapid epuizat ºi cultura trece în faza staþionarã de creºtere. Rata specificã de creºtere
maximã ìm, are o importanþã considerabilã în procesele industriale în care se urmãreºte
obþinerea de celule ºi este dependentã de natura microorganismului ºi de condiþiile de
cultivare. De exemplu, pentru mucegaiuri, aceasta poate sã varieze între 0,090 ºi 0,61 h-1.
La cultivarea lui Aspergillus niger pe un mediu cu glucozã la 30oC s-au obþinut o ratã
maximã de creºtere ìm = 0,2 ºi un timp de dublare al biomasei de 3,46 h. Bacteriile aflate în faza logaritmicã au o citoplasmã omogenã, nu conþin substanþe de
rezervã ºi au o mare afinitate pentru coloranþii bazici, datoritã conþinutului lor mare de ARN.
Mãrimea lor este constantã, depãºind însã dimensiunile caracteristice speciei.
o Faza de încetinire a creºterii corespunde perioadei în care, ca urmare a epuizãrii treptate a nutrienþilor, reducerea concentraþiei în oxigen ºi acumularea de compuºi de
catabolism ce pot avea efect inhibitor asupra celulelor, creºterea ia toate valorile intermediare
între creºterea cu o ratã maximã ºi 0.
o Faza staþionarã de creºtere, când se stabileºte un echilibru între numãrul de celule
care se formeazã prin reproducere ºi cel al celulelor care se autolizeazã. Cantitatea de biomasã

50
poate rãmâne constantã, deºi se schimbã compoziþia celulelor. Datoritã lizei unor celule se elibereazã noi substraturi care vor servi drept nutrienþi pentru celulele viabile. Aceastã fazã
poate fi prelungitã atunci când urmãrim pãstrarea culturii pure, prin modificarea unor factori
care scad viteza de metabolism celular;
o Faza de declin sau curba de distrugere ºi inactivare metabolicã a celulelor, are loc ca urmare a urmãtorilor factori:
o lipsa surselor de nutriþie ºi energie; o denaturarea componentelor celulare în prezenþa substanþelor acumulate
(alcooli, acizi, º.a.); o procentului de celule autolizate.
Consecinþa acestor acþiuni conduce, în final, la sterilizarea mediului ºi, deci, la
moartea tuturor celulelor, respectiv la pierderea culturii. Culturi discontinue sincrone
În cazul culturilor bacteriene obiºnuite diviziunile celulelor individuale urmeazã ritmuri proprii, în aºa fel încât rezultã o populaþie eterogenã, formatã din indivizi aflaþi, practic,
în toate fazele ciclului lor de creºtere ºi multiplicare. Chiar în cazuri speciale, în care pornim de
la o singurã celulã, se constatã cã la descendenþii acesteia, dupã o perioadã de diviziune sincronã
apar diferenþe decurgând din timpul diferit dnecesar celulelor progene sa se dividã. Culturile cu creºtere sincronã sunt selecþionate din culturi asincrone ºi conþin
organisme foarte asemãnãtoare deoarece aparþin aceleiaºi categorii de vârstã ºi/sau de
dimensiune ºi ca urmare se divid aproape simultan. Procesul de diviziune sincronã dureazã numai câteva generaþii (2-3) deoarece
diferenþele individuale determinã procese de defazare în ritmul de creºtere, chiar între
descendenþii unei singure celule. Tehnicile de obþinere ale culturilor sincrone pot fi grupate în douã categorii: A. Tehnici bazate pe selecþie dupã dimensiuni de vârstã:
filtrarea prin teanc de hârtii de filtru suprapuse; separarea prin centrifugare în gradienþi de densitate nemetabolizabili de
microorganismul studiat; separarea prin filtrare pe filtre de membranã.
B. Tehnici bazate pe modificãri ale mediului, pot fi utilizate în numerase variante: expunerea microorganismelor auxotrofe la o perioadã de înfometare, în medii
fãrã n factor de creºtere esenþial, înainte de a fi trecute într-un mediu complet; expunerea la doze subletale de radiaþii; expunerea la acþiunea unui antibiotic care inhibã sinteza proteinelor; expunerea microorganismelor fotosintetizante alternativ la întuneric ºi
luminã; expunerea alternativã, unicã sau repetatã, la temperaturi optimale ºi supra-
sau suboptimale pentru creºtere. Culturi continue
Anumite procese experimentale de laborator sau de producþie industrialã necesitã
prezenþa unor bacterii în fazã exponenþialã de creºtere, ceea ce nu se poate realiza timp
îndelungat în condiþiile obiºnuite de cultivare, într-in spaþiu închis, respectiv într-un volum fix de mediu nereînnoit, în care faza exponenþialã are un numãr limitat de generaþii, iar unele
condiþii de mediu devin limitante. Obþinera culturilor continue se realizeazã cu ajutorulunui dispozitiv numit chemostat
baza pe principiul diluþiei ontinue în volum constant.