cursuri histologie
TRANSCRIPT

Histologie
Anul II
Histologie - curs 1
1

TESUTUL EPITELIAL
Tesutul epitelial este tesutul cel mai raspandit in organism.Principalele caracteristici:
· este format din celule ce au un aspect de mase celulare compacte · spatiile intercelulare sunt foarte reduse (au putina substanta extracelulara)· celulele sunt unite prin jonctiuni· orice epiteliu are doua suprafete: libera (orientata catre mediul extern) si atasata la tesutul
conjunctiv, intotdeauna ancorata la el printr-o masa acelulara (bogata in proteine si glicozaminoglicani sulfatati) care formeaza membrana bazala (MB).
Diferente intre tesutul epitelial si tesutul epiteloid: tesutul epiteloid are MB dar nu are suprafata libera. Spre exemplu, in corticosuprarenala, apar celule secretorii, atasate prin jonctiuni la MB dar care nu au suprafata libera. Acesta este un tesut epiteloid.
· are o mare diversitate de forme si functii: · formeaza invelisuri membranare (epiderm, cavitatile inchise ale organismului: pleura,
peritoneu, pericard, peretii cailor de comunicare cu mediul extern: arborele traheo-bronsic, tubul digestiv, caile genito-urinare etc.).
· asigura transportul selectiv de molecule intre mediul intern si extern sau intre doua compartimente ale organismului.
· intra in structura unor organe solide si a unor glande: tesutul epitelial realizeaza unitatea secretorie si ajuta la formarea sistemului canalicular, deci formeaza parenchimul glandei (parenchimul glandular este format din unitatile secretorii si canaliculele secretorii).
· au rol senzorial, in organele de simt (mugurii gustativi, organul Corti, crestele ampulare, aparatul otolitic).
· au rol contractil, care prezinta o importanta minora. Acestea sunt populatii aparte de celule epiteliale, care se numesc celule mioepiteliale (un tesut epitelial care prezinta miofilamente contractile in citoplasma)
Un tesut epitelial poate realiza una sau mai multe functii.Rolul sau este asemanator cu al membranei celulare, dar relativ la organ si anume: separa
selful de non-self si monitorizeaza, controleaza si modifica substantele care il traverseaza.Caracterul ubicuitar al tesutului epitelial este determinat de originea embriologica a sa care
este in oricare dintre cele trei foite embrionare:· din ectoderm deriva:
· epidermul si derivatele sale (par, unghii)· glandele tegumentare (sudoripare si sebacee)· epiteliul cavitatii bucale, nazale si anale· unele glande endocrine: adenohipofiza
· din endoderm deriva:· tubul digestiv si aparatul respirator · parenchimul glandelor anexe digestive · glandele endocrine (paratiroida, tiroida si timusul)
· din mezoderm deriva:· corticosuprarenala· endoteliile · mezoteliileTesutul epitelial este lipsit de vase sangvine si limfatice (este avascular). Vasele sangvine
aflate in tesutul conjunctiv nu traverseaza MB si tesutul epitelial este hranit prin difuziune. Spre
2

exemplu, in cazul epiteliului pluristratificat, celulele aflate in contact cu MB sunt mai bine hranite decat cele aflate la o distanta mai mare de MB. Exista si un epiteliu vascularizat: stria vasculara, care se gaseste in urechea interna, pe peretele lateral al canalului cohlear.
Este bine inervat, prin terminatii nervoase libere, distribuite in mod neuniform, in retea butonata, retea in cosulet sau retea nebutonata. Anumite epitelii nu au terminatii nervoase libere pentru durere, ceea ce permite efectuarea anumitor manevre fara anestezie (raclarea epiteliului gastric si intestinal pentru biopsie, cauterizarea colului uterin).
Tesutul epitelial are capacitatea de regenerare (proces fiziologic prin care sunt inlocuite celulele uzate, prin activitate mitotica neintrerupta a unor celule nediferentiate din epiteliu, numite celule stem). Rata de inlocuire a unui epiteliu variaza in limite foarte largi: epiteliul intestinal se inlocuieste in 2-5 zile (cel mai rapid) iar epidermul se inlocuieste in 30 zile (cel mai lent).
Inlocuirea se poate face in trei moduri:1. pe seama unor celule bazale izolate, intercalate intre celulele epiteliale. Apare in cazul unor
epitelii unistratificate cilindrice (epiteliul trompelor uterine, epiteliul cailor genitale masculine: epididim si canal deferent)
2. pe seama unor celule bazale grupate (de obicei intr-un strat numit strat germinativ)3. pe seama unor zone numite zone germinative. Apare in cazul glandelor intestinale si al celor
gastrice (in fundul criptelor gastrice apar aceste zone germinative).Metaplazia este procesul de transformare a unui tip de epiteliu intr-un alt tip de epiteliu.
Spre exemplu, epiteliul unistratificat se transforma in epiteliu pluristratificat. Metaplazia apare sub actiunea cronica a unor factori nocivi de mediu. Spre exemplu, sub actiunea substantelur toxice din fumul de tigara epiteliul pseudostratificat ciliat din caile respiratorii se transforma in epiteliu pluristratificat si isi pierde functia. Daca actiunea nociva inceteaza, procesul este reversibil.
EPITELIUL DE ACOPERIREAlcatuieste invelisuri membranare, adeseori cu rol de protectie.Poate fi clasificat dupa numarul de straturi in epiteliu simplu (unistratificat) si epiteliu
stratificat (pluristratificat) sau, dupa forma celulelor, in epiteliu cilindric (columnar, prismatic), scuamos (pavimentos) sau cubic. Clasificarile nu tin seama de particularitatile functionale. Deci epiteliile pot fi:· simple
· pavimentoase (scuamoase)· cubice· cilindrice (columnare, prismatice)
· stratificate:· pavimentoase (cheratinizate sau necheratinizate)· cubice· cilindriceClasificarea dupa forma celulei se face dupa stratul superficial, in cazul epiteliilor stratificate.
· pseudostratificat cilindric · de tranzitie (uroteliu)
Epiteliile pavimentoase simple (scuamoase)Seamana cu solzii pestilor, au nuclei alungiti si turtiti, deci acest tesut apare ca un sir de
nuclei turtiti. Acest epiteliu actioneaza ca o bariera simpla permeabila, asigurand un tranzit selectiv de gaze si metaboliti.
Ex.: epiteliul alveolei pulmonare, ramura descendenta a ansei Henle, foita parietala a capsulei Bowman. Mezoteliile si endoteliile sunt tot epitelii simple scuamoase. Endoteliul este epiteliul care captuseste orice tip de vas. Mezoteliile intra in structura seroaselor, alcatuind membrana ce captuseste cavitatile inchise ale organismului (pleura, pericard, peritoneu).
3

Epiteliul simplu cubicLatimea si inaltimea celulelor este aproximativ egala. Nucleul este rotund si situat central de
regula. D.p.d.v. functional acest epiteliu este destinat absorbtiei sau secretiei (in functie de sens). Exista si locuri unde are rol de protectie. Exemple: ramura ascendenta a ansei Henle si segmentul distal al tubului urinifer, canalele glandelor exocrine (intralobulare) care se varsa in ducte (canale interlobulare), epiteliul de acoperire al ovarelor.
Epiteliul simplu cilindricEste format dintr-un rand de celule inalte (inaltimea este de 2-3 ori mai mare decat latimea).
Nucleii sunt alungiti si ovalari, asezati spre MB (dar exista si cazuri cand sunt asezati central sau chiar apical). Poate avea specializari la nivel apical (cili, microvili, stereocili).
Exemple: tubul digestiv (stomac, intestinul subtire si cea mai mare parte a intestinului gros), caile genitale feminine (trompele uterine si uterul), segmentul proximal al tubului urinifer, glandele exocrine - ductele striate.
Functii: · de secretie si absorbtie (absorbtia este marita datorita prezentei microvililor);· de transport al moleculelor de-a lungul celulei (realizat de catre cili)
Epiteliul stratificat scuamosEste alcatuit din mai multe randuri de celule. Doar stratul superficial este scuamos (nucleii
sunt turtiti numai la suprafata, in rest fiind rotunzi).Este de doua tipuri:
1. cheratinizat (cornos)2. necheratinizat (moale) Deosebirile intre cele doua tipuri apar la nivelul stratului superficial.
In cazul epiteliului cheratinizat, celulele de la suprafata acumuleaza proteine (cheratine) si astfel ele isi pierd nucleii. Acesta este cazul epidermului, care este impermeabil pentru apa si are un rol protector impotriva factorilor fizici, chimici si infectiosi.
In cazul epiteliului necheratinizat, celulele superficiale au si ele in citoplasma granule de cheratina dar ele isi pastreaza nucleii. Acest epiteliu este localizat in primele segmente ale tubului digestiv (cavitate bucala, faringe, esofag), epiteliul anterior al corneei., epiteliul vaginal. Acest epiteliu are un rol protector dar este permeabil pentru unele molecule mici si pentru apa.
Epiteliul stratificat cubic si cilindricSunt slab intalnite la adult. La embrion se gaseste frecvent epiteliul stratificat cilindric.Epiteliul stratificat cubic se gaseste la adult in peretii unor ducte interlobulare a unor glande
exocrine, in peretele canalului de secretie a glandelor sudoripare, in peretele foliculilor ovarieni.Epiteliul stratificat cilindric se gaseste pe suprafata respiratorie a epiglotei si in conjunctiva
palpebrala.
Epiteliul pseudostratificat cilindricEste o forma de epiteliu simplu. Toate celulele au raport direct cu MB, dar sunt de inaltimi
diferite astfel incat doar unele ajung la suprafata. Prin urmare, si nucleii ajung la inaltimi diferite. Poate prezenta la nivelul membranei apicale specializari (cili sau stereocili). Epiteliul pseudostratificat cilindric ciliat se mai numeste si epiteliu de tip respirator deoarece
se gaseste numai in arborele traheobronsic. Epiteliul pseudostratificat cilindric cu stereocili se gaseste in caile genitale masculine, in
epididim si canalul deferent. Stereocilii se vad si la microscopul optic si la cel electronic, fiind putin mai lungi decat cilii.
4

Epiteliul de tranzitie (epiteliul polimorf sau uroteliu)Epiteliul se mai numeste si uroteliu deoarece se gaseste numai in caile urinare (calice mici,
calice mari, bazinet, ureter, vezica urinara, partea proximala a uretrei). Epiteliul se mai numeste si polimorf deoarece este format din celule care au diferite forme (celule din stratul bazal - cubice, celule in racheta, celule umbeliforme - in forma de umbrela).
Acest epiteliu este o forma speciala de epiteliu stratificat.Celulele din stratul bazal sunt celule germinative.Celulele in racheta prezinta un pol latit, indreptat spre stratul celulelor umbeliforme si un pol
efilat (ingustat) indreptat spre stratul bazal.Celulele umbeliforme sunt uni- sau binucleate si acopera 2-3 celule in racheta.Epiteliul de tranzitie isi modifica forma celulelor si numarul de straturi. Modificarea
numarului de straturi se datoreste si faptului ca (probabil) toate celulele acestui epiteliu sunt ancorate la MB. Atunci cand se exercita o presiune, celulele gliseaza unele pe langa altele si se reduce numarul de straturi de celule.
Modificarea formei celulare se realizeaza in special la celulele umbeliforme. Prin studii de microscopie electronica s-au observat pe membrana acestor celule “arii dense ale celulelor umbeliforme”. Acestea reprezinta niste invaginari ale membranei care constituie rezerve membranare. Cand celula urinara este goala, ariile se vad clar iar cand celula este plina, ariile dispar, ca urmare a intinderii membranei celulare, proces care se produce cu ajutorul miofibrilelor din citoplasma.
Epiteliul este impermeabil, constituind o bariera osmotica. Aceasta functie este data de organizarea membranei celulare care are o compozitie chimica bogata in sfingolipide.
EPITELIUL GLANDULARReprezinta grupuri de celule epiteliale specializate pentru procese de secretie. Procesul
secretor are 4 faze:1. Asimilare - celula preia din mediul extracelular precursorii de sinteza2. Sinteza - prelucrarea precursorilor3. Ambalarea si stocarea - are loc in granule de secretie4. Secretia propriu-zisa - eliminarea extracelulara a granulelor
Glandele pot fi:· exocrine - isi varsa produsii de secretie in mediul extern (cu sau fara ajutorul canalelor de
secretie)· endocrine - isi varsa produsii de secretie in mediul intern (preferential in sange)
Histogeneza (geneza tesutului)Se porneste de la o lama epiteliala ce vine in contact cu un tesut conjunctiv tanar (tesut
mezenchimal). Aici exista un grup de celule ce formeaza un mugure ce patrunde in tesutul mezenchimal. Evolutia spre glanda exocrina se face atunci cand mugurele pastreaza legatura cu lama iar spre glanda endocrine se face atunci cand mugurele nu mai pastreaza legatura cu lama.
Glandele pot fi exocrine, endocrine sau mixte (amficrine).Glanda reprezinta asocierea dintre epiteliul glandular, vase si nervi.
Glandele exocrinePartea lor profunda alcatuieste unitatea secretorie si se numeste adenomerul glandular. El
pastreaza legatura cu lama epiteliala prin intermediul canalelor de excretie. El vine in raport cu vase limfatice si sangvine si cu membrana bazala.
Clasificarea glandelor exocrine dupa structura se poate face dupa:
5

· numarul de celule din glanda exocrina· aspectul sistemului canalicular· aspectul adenomerului
Dupa numarul de celule din glanda, glandele exocrine pot fi unicelulare sau multicelulare. Exista un singur tip de glanda exocrina unicelulara: celula caliciforma (are forma de cupa), care se gaseste in epiteliul respirator sau in epiteliul digestiv. Restul glandelor exocrine sunt multicelulare.
Uneori celulele din glanda exocrina se diferentiaza intr-un epiteliu de invelis, ceea ce va duce la formarea unei glande intraepiteliale (in mucoasa nazala). Aceasta contine celule ce formeaza un epiteliu de invelis si celule secretorii, formand mase compacte asociate unui sistem canalicular.
Glanda exocrina isi varsa produsii de secretie intr-o cavitate cu sau fara ajutorul canalelor de excretie. Clasificarea glandelor dupa sistemul canalicular se face in simple si compuse. Glandele simple au un canal unic de excretie, neramificat. Glandele compuse au un canal de secretie ramificat.
Clasificarea glandelor dupa forma adenomerului se face in glande tubulare, acinoase sau alveolare.
Adenomerul tubular poate fi:· unic si drept (glande tubulare drepte)· unic dar ramificat (glande tubulare ramificate)· contort (glande tubulare glomerulare)
Adenomerul acinar se aseamana cu cel alveolar, cu precizarea ca cel alveolar are un scurt canal prin care se varsa in canalul excretor, canal care la adenomerul acinar lipseste. Exista si glande tubulo-acinoase si tubulo-alveolare.
Pentru definirea corecta a tipului unei glande se folosesc doua adjective: unul referitor la forma canalelor si al doilea referitor la forma adenomerului. Exista astfel urmatoarele tipuri de glande:
· glanda simpla tubulara dreapta (glanda intestinala - glanda Lieberkühn)· glanda simpla tubulara ramificata (glandele stomacului - fundice si pilorice si glandele duodenale
Brünner)· glanda simpla tubulara glomerulara (glanda sudoripara)· glanda simpla alveolara sau alveolara ramificata (glanda sebacee)· glanda simpla acinoasa (glanda lacrimala)· glanda compusa acinoasa (glanda parotida)· glanda compusa tubuloacinoasa (glandele sublinguale si submaxilare)· glanda compusa tubuloalveolara (glanda mamara)
Glandele exocrine mai pot fi clasificate si dupa produsii de secretie in glande seroase, glande mucoase si glande mixte (seromucoase sau mucoseroase)
Glandele mucoase secreta mucus. Mucusul contine proteine inalt glicozilate (mucine). Celulele glandelor mucoase au un continut ridicat de glicozil transferaze (necesare glicozilarii proteinelor), care se gasesc in aparatul Golgi.
Glandele seroase au o secretie cu aspect apos, care contine protein-enzime. Exemplu: glanda parotida.
Glandele mixte pot fi:· seromucoase (glanda submandibulara) - au o secretie predominant seroasa· mucoseroase (glanda sublinguala) - au o secretie predominant mucoasa
De asemenea, glandele exocrine mai pot fi clasificate si dupa mecanismul de excretie in glande merocrine, apocrine sau holocrine.
Majoritatea glandelor sunt merocrine. Acestea isi elimina produsii prin exocitoza.
6

Glandele apocrine isi acumuleaza produsii de secretie la polul apical si pentru excretie se rupe membrana care se pierde, impreuna cu o parte din citoplasma apicala. Exemple: glanda mamara si glandele ceruminoase, care produc ceara si se gasesc in conductul auditiv extern.
In cazul glandelor holocrine, produsii de secretie invadeaza toata celula, ceea ce duce la moartea celulei. Eliminarea produsilor de secretie se va face prin eliminarea intregii celule moarte. Exemplu: glanda sebacee.
Glandele endocrineIsi elimina produsii de secretie in mediul intern: in sange (preferential), in limfa sau in
lichidul interstitial.Momentul de eliberare a produsilor de secretie poate fi:
· imediat (corticosuprarenala) - glande cu secretie continua· dupa o prealabila perioada de stocare (intracelulara sau extracelulara)
· stocarea intracelulara are loc prin granule de secretie· stocarea extracelulara: tiroida depoziteaza produsul de secretie in coloidMajoritatea glandelor endocrine sunt de origine epiteliala dar exista si glande de origine
nervoasa. O dovada a originii epiteliale este contactul celulelor secretorii cu membrana bazala.Criterii de clasificare a glandelor endocrine:
· dupa modul de dispunere a celulelor:· in cordoane celulare, dispuse fie paralel, fie ramificate si anastomozate (ex.: zona
fasciculata din corticosuprarenala)· in foliculi - o cavitate (coloid) cu un continut delimitat de un epiteliu secretor (ex.:
tiroida)· insule, cuiburi sau gramezi (ex.: insulele Langerhans din pancreasul endocrin)
· dupa natura produsilor de secretie:· glande ce secreta polipeptide mici si amine biogene (ex.: celulele ce alcatuiesc sistemul
neuroendocrin difuz - APUD)· glande ce secreta proteine si polipeptide mari (majoritatea glandelor endocrine)· glande care secreta steroizi (corticosuprarenala, componenta endocrina a gonadelor)
Histologie - curs 2
1. Polaritatea tesutului epitelial2. Specializarile membranare3. Membrana bazala
7

POLARITATEA TESUTULUI EPITELIAL
Indiferent de localizare, tesutul epitelial indeplineste numeroase roluri. El are proprietatea de polaritate, care este de doua feluri:· polaritatea tesutului epitelial (tisulara)· polaritatea celulei epiteliale (celulara)
Polaritatea tisularaTesutul epitelial are polaritate datorita rolului de bariera pe care-l indeplineste, separand doua
medii cu compozitie diferita. Tesutul epitelial are deci doua suprafete:1. suprafata libera orientata spre mediul extern sau lumen2. suprafata orientata spre membrana bazala
Tesutul epitelial este o bariera pentru factori fizici, chimici etc.La unele epitelii, suprafata spre mediul extern are specializari. In cazul tesutului
pluristratificat, stratul liber este cel specializat si deci este bine structurat. La epiteliul pluristratificat pavimentos cheratinizat, suprafata libera este reprezentata de cheratina iar celulele specializate se numesc cheratinocite. La epiteliul simplu, spre mediul extern apar specializari membranare.
Polaritatea celularaPolaritatea celulara consta in existenta la nivelul membranei celulelor epiteliale a unor
domenii membranare distincte, ce difera intre ele d.p.d.v. biochimic, structural si functional. In mod obisnuit, o celula epiteliala are doua domenii membranare:1. apical (spre lumenul organului)2. laterobazal (spre tesutul conjunctiv)
Cele doua domenii vin in contact cu compartimente cu continut diferit d.p.d.v. al lichidului extracelular.
Elemente de definire a polaritatii· cele doua domenii membranare difera d.p.d.v. biochimic si apare astfel asimetria biochimica a
suprafetei membranei (diferenta intre membrana apicala si cea laterobazala).· existenta jonctiunilor stranse (zonulae ocludens) si realizarea contactelor intercelulare ce sunt
asigurate prin molecule de adezivitate celulara si dispozitive jonctionale.· dispozitia caracteristica a organitelor celulare si a componentelor de citoschelet in citoplasma.
AsimetriaFiecare domeniu are functii specifice. Domeniile difera atat d.p.d.v. protidic cat si lipidic.
D.p.d.v protidic:· Domeniul apical are un set caracteristic de proteine membranare ce cuprinde:
enzime;proteine de transport;canale ionice.
Deci, domeniul apical contine setul de proteine ce asigura absorbtia si secretia celulara. Enzima marker a domeniului apical este leucinaminopeptidaza.Numeroase proteine din domeniul apical au atasate si resturi glucidice, care vor forma glicocalixul. Aceasta explica reactia PAS pozitiva.
· Domeniul bazal contine:enzimeproteine de transport
8

canale ioniceproteine receptoareproteine de adezivitate
Existenta receptorilor si moleculelor de adezivitate explica rolul acestui domeniu in recunoasterea si transcrierea informatiilor si respectiv mentinerea integritatatii structurale. Proteine de adezivitate sunt caderina E si integrinele care fac parte din glicoproteinele membranare cu activitate dependenta de Ca2+. Receptorii sunt cei pentru insulina, parathormon si EGF (epitelial growth factor).
Prin examinarea continutului proteic al celor domenii s-a observat ca exista numeroase similitudini intre enzime. Totusi, enzima marker a domeniului laterobazal este Na+/K+ ATP-aza.
D.p.d.v. lipidic: Se constata existenta acelorasi tipuri de lipide in cele doua domenii, dar difera concentratia lor, ceea ce implica diferente functionale si fizico-chimice.
· Domeniul apical prezinta o concentratie mare in colesterol si sfingomielina· Domeniul laterobazal prezinta o concentratie mare in fosfatidil colina si fosfatidil
inozitol (PC&PI).
Aparitia jonctiunilor ocludensJonctiunile stranse separa domeniul apical de cel laterobazal. Integritatea acestor jonctiuni
face posibila existenta celor doua domenii. Jonctiunile stranse sunt situate spre polul apical.Interactiunile celulelor epiteliale pot fi de doua tipuri:
1. cu celule epiteliale adiacente numite interactiuni (contacte) celula - celula2. cu matricea extracelulara numite interactiuni celula - substrat.
Intre moleculele de adezivitate ce asigura contactul celulelor epiteliale, un rol deosebit il are caderina E in cazul interactiunilor celula - celula si integrina in interactiunile celula - substrat.Cele doua domenii difera si prin tipul de jonctiune prin care se realizeaza contactele:· Domeniul lateral ® apare complexul jonctional alcatuit dinspre polul apical al celulei spre cel
bazal din:· jonctiune stransa · jonctiune intermediara (zonula adherens)· desmozom· jonctiune tip gapFiecare asigura o functie:· Jonctiunea stransa separa domeniul apical de cel bazal · Zonula adherens si desmozomul sunt jonctiuni de ancorare si asigura integritatea
functionala si morfologica a epiteliului (asigura interactiunea celulara).· Jonctiunea gap (nexus) asigura comunicarea intre celulele epiteliale.
· Domeniul bazal ® apar alte doua tipuri de jonctiuni care asigura contactul celula - substrat:· jonctiuni focala· hemidesmozomiulDistributia organitelor celulare in citoplasma In cazul unei celule epiteliale glandulare exocrine, la microscopul electronic, exista o anume
distributie:1. Reticulul endoplasmic rugos - la polul bazal al celulei2. Complexul Golgi localizat supranuclear3. Granule de secretie la polul apical
Aceasta repartitie sugereaza succesiunea etapelor din procesul de secretie1. Sinteza proteinelor incepe in reticulul endoplasmic rugos
9

2. Se continua sinteza proteinelor in aparatul Golgi 3. Desavarsirea secretiei are loc in granulele de secretie si se incheie prin exocitoza
granulelorAceasta distributie a organitelor citoplasmatice se datoreaza centrului celular (centrozom).Centrul celular este alcatuit din doi centrioli iar in jurul lor exista o zona din citoplasma
densa la fluxul de electroni. Centrul celular se afla la periferia celulei intr-o anumita pozitie. Zona densa la fluxul de electroni reprezinta centrul de formare a microtubulilor. Acestia determina, sub actiunea centriolilor o anume deplasare a organitelor in citoplasma. Microtubulii sunt alcatuiti din tubuline si sunt supusi unor procese de polimerizare si depolimerizare, fiind niste organite orientate in citoplasma. Organizarea lor se face datorita centriolilor.
Componentele citoscheletului· Filamente intermediare (la celula epiteliala sunt de citocheratina si se numesc tonofilamente)· Microtubuli
Distributia componentelor de citoschelet:1. Microtubulii au o orientare apico-bazala (in lungul celulei). Prin aceasta dispozitie, prin
polimerizari si depolimerizari, ei asigura transportul de vezicule dinspre domeniul bazal spre cel apical si invers.
2. Microfilamentele de actina. Actina din citoplasma interactioneaza cu suprafata membranara la mai multe nivele:3. formeaza fascicule polarizate si interactioneaza cu domeniul apical formand microvili
care cresc suprafata de absorbtie la acest nivel.4. formeaza dispozitive jonctionale. Microfilamentele de actina formeaza la nivelul
domeniului lateral jonctiuni stranse (ocludens) si intermediare (adherens), iar la nivelul domeniului bazal se organizeaza sub forma de contacte focale.
5. actina interactioneaza cu moleculele de adezivitate celulara: “citoscheletul se ancoreaza”. Astfel, la nivelul domeniului bazal actina interactioneaza cu integrinele iar la nivelul domeniului apical interactioneaza cu caderina E. Interactiunea dintre caderina E si actina este mediata de catenine (proteine de tip a, g, D). Interactiunea dintre caderina si actina e datorata Ca2+. Caderina E interactioneaza cu actina F si se ancoreaza prin catenine.
6. Actina interactioneaza cu proteinele membranare de suprafata (Na+/K+ ATP-aza). Daca actina interactioneaza cu proteinele membranare de suprafata, ea mentine aceste proteine intr-o anumita zona, asigurand stabilitatea metabolica a celulei epiteliale. Proteine membranare de suprafata sunt anchirina, folina.
Stabilirea si mentinerea polaritatii celulei epitelialeStabilirea polaritatii celulei epiteliale este un proces complex, determinat genetic, care
parcurge mai multe etape. Afectarea unei functii determina aparitia unei disfunctii celulare care va duce la aparitia unei boli. Deci, daca celula epiteliala are polaritate, ea va functiona normal, iar pierderea polaritatii duce la aparitia unei boli.
Stabilirea polaritatii celulei epiteliale necesita 3 etape.Prima etapa consta in formarea de contacte celula - celula si celula - substrat ceea ce va
duce (in etapa a doua) la organizarea unui grup de celule epiteliale intr-un monostrat. Realizarea acestor contacte se datoreaza caderinei E. Rolul acesteia a fost studiat experimental (in vitro, in vivo). Ea este implicata in contactul celula - celula. Experimentul s-a realizat pe celule L (celule conjunctive tinere, fibroblaste de sobolan). Celulele L sunt nepolarizate, stabilesc intre ele contacte reduse si contin la nivelul membranei Na+/K+ ATP-aza dar ea este distribuita difuz. Membrana celulara nu contine caderina E. In mediul de cultura se introduce ADN complementar pentru caderina E, deci va aparea caderina E. Se constata ulterior formarea de numeroase
10

contacte celulare. Se observa o anume redistribuire a Na+/K+ ATP-azei, pe aumite domenii membranare, dar nu se observa redistributia citoscheletului de actina. Acest experiment explica rolul caderinei.
Caderina nu este suficienta pentru jonctiune dar este necesara integritatea sa. Caderina E este o glicoproteina integrala. Domeniul sau transmembranar leaga domeniul extracelular (care asigura jonctiunea celula - celula), de domeniul intracelular. Daca se indeparteaza domeniul intracitoplasmatic al caderinei, dispar contactele celula - celula, ceea ce dovedeste necesitatea integritatii proteinei pentru a-si putea indeplini functia. De asemenea, daca molecula sa este fragmentata, ea nu mai reactioneaza cu cateninele si nu mai ancoreaza actina.
A doua etapa consta in organizarea celulelor in monostrat. Acest proces duce la individualizarea celor doua domenii membranare, ceea ce are loc in viata intrauterina. Initial proteinele si lipidele au o dispozitie difuza si apoi se realizeaza o fixare selectiva a proteinelor si lipidelor, proces ce sta la baza remodelarii membranei plasmatice.
Pentru remodelare, celula foloseste proteinele din membrana si proteinele din citoplasma (care alcatuiesc impreuna fondul comun de proteine). Fixarea selectiva a proteinelor (deplasarea lor) se poate face prin translatie laterala. Dar aceasta posibilitate este putin probabila. Cel mai probabil este ca aceasta se face prin transcitoza.
Generarea celor doua domenii nu este concomitenta. Primul domeniu care apare este cel apical care se realizeaza rapid, necesitand doar initierea contactelor celula - celula. Al doilea domeniu care apare este cel bazal care apare gradat, lent necesitand existenta jonctiunilor stranse celula - celula.
Cea de-a treia etapa consta in reorganizarea citoscheletului de actina.
Implicatiile polaritatii celulei epiteliale in clinicaS-au descris mai multe afectiuni cu localizare diferita, in care alterarea integritatii epiteliale
nu este cauza dar apare ca o consecinta a bolii. Dintre acestea enumeram: carcinoamele, insuficinta renala acuta prin ischemie, boala Davidson (boala cu microvili inclusi), boala rinichiului polichistic, Pemfigus Vulgaris.
7. Carcinomul este o tumora maligna a tesutului epitelial. Exista denumiri generice precum adenocarcinom (tumora unei glande). Ele sunt cele mai frecvente (reprezinta 90% din totalul cancerelor). Carcinomul presupune o proliferare necontrolata a celulelor epiteliale. Afectarea polaritatii apare din invazivitatea, metastazarea carcinomului. In transformarea unui tesut epitelial normal intr-un tesut malign intervin caderina E si cateninele. In metastazarea carcinomului, caderina E este exprimata genetic putin, ceea ce duce la scaderea cantitatii de caderina. In aceste cazuri exista si o alterare a cateninelor precum si o modificare in citoscheletul de actina. Prin urmare, sunt afectate contactele celula - celula, ceea ce explica migrarea celulelor (metastazarea lor).
8. In insuficienta renala acuta prin ishemie celulele renale sunt afectate prin ischemie si se transforma in celule nepolarizate. Termenul de “acuta” arata faptul ca boala se instaleaza rapid. Ischemia inseamna scaderea fluxului sangvin ceea ce duce la scaderea cantitatii de oxigen si substante nutritive. Cele mai afectate sunt celulele tubului urinifer, unde au loc procese de secretie si reabsorbtie, si in special tubul contort proximal unde exista microvili si absorbtia este crescuta. Leziunile determinate de ischemia tubului proximal depind de factorul timp. La doar 5
minute de ischemie are loc o redistributie a lipidelor si proteinelor membranare din celula. In acest caz Na+/K+ ATP-aza ajunge in polul apical, integrinele se redistribuie pe toata suprafata celulara. Primele modificari constau in redistributia proteinelor membranare. Daca ischemia continua apar fragmentari ale citoscheletului de actina. Cele doua modificari morfologice duc la aparitia principalelor modificari functionale.
11

Redistributia Na+/K+ ATP-azei, care se va gasi atat in domeniul bazal cat si in cel apical, influenteaza cantitatea de molecule absorbite din tubul urinifer. Spre exemplu, in tubul proximal are loc in mod normal o absorbtie crescuta de Na+, strabatand membrana apicala dinspre mediul extracelular spre celula. Daca in domeniul apical apare Na+/K+ ATP-aza aceasta va scoate 3 ioni Na+ si va introduce 2 ioni de K+, ceea ce va duce la aparitia unui ciclu inutil (o parte este absorbit si o parte se reintoarce in lumen) si va fi afectat numarul ionilor absorbiti. Acest rationament este valabil pentru orice tip de ion.Fragmentarea actinei are consecinte asupra microvililor care isi vor pierde integritatea, ceea
ce duce la reducerea absorbtiei. Actina participa si la formarea jonctiunilor iar daca isi pierde integritatea va duce la disparitia jonctiunilor care va duce la deschiderea spatiului intercelular, iar fluidele vor putea trece prin spatiul intercelular.Redistributia integrinelor va duce la disparitia contactelor celula - substrat ceea ce va
determina celulele sa cada in lumenul tubului urinifer si astfel este posibila obstruarea lumenului tubului urinifer.
9. Boala Davidson consta in alterarea citoscheletului de actina care va duce la pierderea microvililor. Boala se manifesta la nou nascut. Atunci cand sunt afectate celulele tubului intestinal are loc reducerea suprafetei de absorbtie, deci scade cantitatea de substante absorbite ceea ce va avea efecte grave asupra copilului: scaderea in greutate si malnutritia, care in final vor conduce la deces.
10. Boala rinichiului polichistic este o boala determinata genetic, in care tubul renal este inlocuit cu chisturi (formatiuni ovalare care contin lichid). Aceasta va duce la pierderea zonei renale, ceea ce va determina insuficienta renala. Aparitia chisturilor este datorata lipsei polaritatii ca urmare a redistributiei Na+/K+ ATP-azei si a EGF.
11. Pemfigus Vulgaris este o dermatoza (dermato = piele) care apare la adult. Ea are un debut brusc. Pe suprafata de piele sanatoasa, sau pe mucoasa apar bule care dupa spargere formeaza cruste (coji) sangerande. Dintre mucoase, cea mai afectata este cea bucala. Daca are loc o alimentatie deficitara se poate ajunge la casexie (slabire accentuata) care va duce la deces. In aceasta boala afectarea polaritatii nu este cauza. In plasma celor bolnavi exista anticorpi pentru catenine (a, g, b). Datorita afectarii acestora, se pierd contactele celula - celula, ceea ce favorizeaza aparitia bulelor (spatii intre celule).
SPECIALIZARI MEMBRANARE
Aceste specializari apar atat in domeniul bazal cat si in cel apical. Exista deci specializari membranare la nivelul membranei plasmatice apicale, laterale si bazale.
Specializari ale membranei celulare apicaleEle au diverse roluri: marirea suprafetei de absorbtie (microvilii si stereocilii), transport de
molecule si celule (cilii), sau rol senzorial (stereocilii).12. Microvilii sunt specializari ce se disting numai la microscopul electronic. Ei reprezinta forme de
organizare stabila a filamentelor de actina. In celula epiteliala sunt frecventi si sunt in general scurti. Aceste specializari sunt cacteristice celulelor implicate in absorbtie. Ei apar fie izolati (la distante variabile, avand forma si dimensiuni diferite) fie grupati (acopera intreaga suprafata membranara si au aceeasi forma si acelasi diametru).
Microvilii egali, cand sunt grupati au corespondenti in microscopia optica: platoul striat (o zona striata la polul apical cu striatii perpendiculare pe suprafata celulei) si margine in perie (aspect de fire la polul apical).
Exista diferente intre platoul striat si marginea in perie. Marginea in perie prezinta microvili mai lungi care au citoscheletul axial mai putin dezvoltat ceea ce permite variatia lungimii lor; ei se rup usor (sunt friabili), ceea ce le da un aspect neregulat. Marginea in
12

perie apare la polul apical al nefrocitului, in segmentul proximal al tubului urinifer. Platoul striat apare la polul apical al enterocitului.
13. Stereocilii se disting si la microscopul electronic si la cel optic. La microscopul optic se aseamana cu niste cili mai lungi. Studiile de microscopie electronica au aratat ca denumirea este improprie, stereocilii fiind de fapt niste microvili. Ei sunt mai lungi decat microvilii obisnuiti, mai subtiri si adeseori ramificati. Rolul stereocililor depinde de localizarea epiteliului. In unele localizari au rol in marirea absorbtiei (epiteliul cailor genitale masculine: epididim si canal deferent). In epiteliile receptoare din urechea interna (organul Corti, crestele ampulare etc) au rol senzorial.
14. Cilii apar si in microscopia optica si in cea electronica. Spre deosebire de microvili si stereocili, cilii sunt alcatuiti din microtubuli. Ei reprezinta specializari membranare cu mobilitate, asigurand miscari ondulatorii (in val). Au rol in transportul moleculelor sau celulelor de-a lungul epiteliului. Ex.:epiteliul de tip respirator, epiteliul trompelor uterine.
Specializari ale membranei plasmatice bazaleAcestea apar la unele celule epiteliale, adica la cele implicate in transportul transcelular si au
rolul de a mari suprafata. Ele apar ca invaginari ale membranei spre citoplasma (in care exista mitocondrii cu axul longitudinal paralel cu invaginatia). Aceste invaginari produc o compartimentare a citoplasmei la nivel bazal. Ansamblul invaginarilor membranei plasmatice bazale se numeste labirint bazal. Datorita acestor invaginari si mitocondriilor exista corespondent in microscopia optica: aspectul striat al citoplasmei la polul bazal. Ex.: tubul urinifer, glandele exocrine (in lobul, in ductele striate). Aceste specializari contin receptori pentru factorii sangvini si se specializeaza pentru jonctiunea celula - matrice extracelulara. Polul bazal se ancoreaza la matrice prin contacte focale (alcatuite din filamente de actina) si prin hemidesmozomi (alcatuiti din filamente intermediare sau tonofilamente). Indiferent de jonctiune, planul de organizare a jonctiunii e similar: in citosol exista proteine de ancorare intracelulare care sunt conectate cu proteine transmembranare prin intermediul carora se face legatura cu proteinele din matricea extracelulara.
Specializari ale membranei plasmatice lateraleAu rol in stabilirea interactiunilor celula - celula, asigura adezivitatea simpla, care se
realizeaza prin spatiile intercelulare si adezivitatea prin jonctiuni speciale. Spatiile intercelulare contin o retea de glicoproteine care interactioneaza cu domeniile extracelulare a proteinelor transmembranare din membrana laterala a doua celule adiacente. Spatiile intercelulare mai contin lichid ce mediaza schimbul dintre celula epiteliala si matricea extracelulara.
Jonctiunile stranse (zonulae ocludens) fac parte din jonctiunile speciale si au rol de a delimita domeniul apical de cel laterobazal. Ele controleaza permeabilitatea spatiului extracelular, impiedicand trecerea libera a moleculelor in spatiul extracelular.
Jonctiunile de ancorare mentin integritatea epiteliului. Ele sunt jonctiuni adherens (formate din filamente de actina) sau desmozomi (formate din filamente intermediare).
Jonctiunile de comunicare (nexus, gap) asigura o difuzie selectiva a moleculelor intre celulele adiacente. Ele apar in epiteliile embrionare si sunt rare la adult. Exista in doua tipuri de tesuturi: endoteliu si mezoteliu.
Desmozomul este caracteristic tesutului epitelial, astfel incat, daca o persoana examineaza o leziune tumorala la microscopul optic si la cel electronic, el nu poate deosebi celula tumorala decat pe baza modificarilor aparute la nivelul desmozomului.
MEMBRANA BAZALA
13

Este o componenta a matricei extracelulare ce stabileste raporturi constante cu orice tip de proteina. Are in componenta proteine si glicozaminoglicani sulfatati.
Caracteristici:15. MB are o grosime variabila (aprox. cateva zeci de microni) ce se evidentiaza la microscopul
optic, care are o putere separatoare de 0,2m. In coloratii uzuale MB nu se vede la microscopul optic. Pentru evidentiere se folosesc metode histochimice. Atfel, prin metoda PAS, datorita continutului ridicat de glucide, MB apare in rosu purpuriu. Prin impregnare argentica cu precipitarea sarurilor de argint, MB este neagra datorita continutului crescut de fibre de reticulina. Cea mai fina MB este cea din epiteliul vezicii urinare, astfel incat pana la aparitia microscopului electronic existenta membranei bazale in raport cu epiteliul era incerta.
16. MB are un aspect variat: in mod obisnuit apare ca o structura liniara, uneori insa are un aspect ondulat datorita unor proiectii ale tesutului conjunctiv spre epiteliu, proiectii numite papile conjunctive.
17. MB are o compozitie chimica variata (in functie de localizarea epiteliului). Astfel, in orice MB exista 5 componente chimice majore (permanente) si anume: colagen tip IV (o proteina fibroasa), laminina (glicoproteina), entactina (glicoproteina), heparansulfat, fibronectina (glicoproteina). Colagenul, laminina, entactina si heparansulfatul sunt secretate de celulele epiteliale iar fibronectina este secretata de celulele din matrice. Aceste componente chimice majore sunt secretate de celulele epiteliale sprijinite pe membrana si in parte de celulele conjunctive din matricea extracelulara.
18. La examinarea prin microscopie electronica de transmisie, membrana bazala are aspect fin, granular, ce are drept corespondent morfologic o retea de fibrina foarte fina.
19. De la polul bazal al celulelor epiteliale la matricea extracelulara, membrana bazala are trei componente care pot fi evidentiate la microscopul electronic: lamina lucida (rara) care apare ca o regiune electronotransparenta, lamina densa (bazalis) care apare ca o banda electronodensa, lamina fibroreticularis (retea de fibre ce contine un alt tip de colagen, colagen tip III, care se organizeaza formand fibre de reticulina care se vad prin impregnare argentica).Functiile membranei bazale:
20. reprezinta o interfata de adezivitate intre epiteliu si matricea extracelulara: celulele epiteliale se ancoreaza la membrana bazala care, la randul ei, are contact cu celulele tesutului conjunctiv.
21. este o bariera permeabila si selectiva. Dintre componentele MB, asigura permeabilitatea GAG sulfatati care, prin multiplele lor sarcini negative, se organizeaza in pori cu diametrul variabil.
22. controleaza organizarea si diferentierea celulara. Ea mediaza interactiunea dintre receptorii de suprafata, ce apartin celulei epiteliale, si diverse molecule din matricea extracelulara.
Histologie - curs 3
PROCESUL DE CHERATINIZARE
Are loc in epiteliul pavimentos stratificat. Procesul principal consta in sinteza si depozitarea intracelulara a cheratinei, proces ce are loc pe masura ce celula inainteaza dinspre stratul bazal spre cel superficial. In celulele superficiale se va forma o masa proteica rezistenta si compacta care este bogata in cisteina, deci are un continut mare de sulf.
Cheratina este rezultatul a doua procese ce au loc in celula:1. sinteza de citocheratina (condensarea filamentelor intermediare de citocheratina = tonofilamente)2. impregnarea citocheratinei cu proteine speciale (cheratohialina si filagrina)
14

Timpul necesar variaza in raport cu localizarea epiteliului si este cuprins intre 20 si 45 de zile.
Prin aceste doua procese se formeaza cheratina moale, care se deosebeste de cea dura (existenta la nivelul derivatelor epidermului - unghii, par). Cheratina moale este saraca in sulf, bogata in grasimi, pastreaza proprietatea de flexibilitate, se poate descuama.
Acest epiteliu are doua corpuri: un corp mucos si un corp cornos. El are mai multe straturi: bazal (generator), spinos, granular, cornos (care are doua substraturi: stratul lucidum si stratul cornos propriu-zis).
Pe langa cheratinocite, celulele specifice epidermului, care au rol in producerea si depozitarea cheratinei, exista si alte tipuri de celule: melanocite, celule Merkel, celule Langerhans, care au alta origine embriologica decat cheratinocitele.
Melanocitele sunt derivate din crestele neurale si au rol in sinteza melaninei. Ele se gasesc in stratul bazal, alaturi de celulele Merkel, care sunt derivate tot din crestele neurale si au rol de receptori tactili. Celulele Langerhans au origine in maduva osoasa hematogena si intervin in procesul imun (recunosc antigenele).
Cheratinocitele alcatuiesc cele doua compartimente ale epidermului: compartimentul proliferativ si cel functional. Cheratinocitele sunt si ele de mai multe feluri, in functie de stratul in care se afla: cheratinocite bazale (care se gasesc numai in compartimentul proliferativ), si cheratinocite spinoase, granulare sau cornoase, care alcatuiesc impreuna compartimentul functional.
Compartimentul proliferativ este alcatuit din celule cubice sau usor columnare, cu citoplasma bazofila. Exista cantitati mari de poliribozomi, mitocondrii si putin aparat Golgi, ceea ce dovedeste sinteza crescuta de proteine structurale. Cheratinocitele bazale stabilesc jonctiuni desmozomale cu celulele supraiacente (celule spinoase) si hemidesmozomi cu membrana bazala. Aceste cheratinocite prezinta mitoze crescute.
La nivelul stratului bazal exista o populatie heterogena de celule. Exista un procent mic de celule stem (celule de origine) care alcatuiesc un rezervor de celule pentru epiderm. Restul celulelor din stratul bazal evolueaza pe linie cheratinocitara (sintetizeaza si depoziteaza cheratina). In aceste celule au loc cele doua procese: sinteza filamentelor de citocheratina (tonofilamente) si sinteza de involucrina (involucrum = invelis).
Compartimentul functional este alcatuit din celelalte tipuri de cheratinocite (granulare, spinoase si cornoase). Cheratinocitele din stratul cornos se numesc corneocite.
La acest nivel mitozele sunt absente. Au loc doar diferentieri celulare, care se petrec in mai multe etape:1. Are loc acumularea treptata de citocheratina la nivelul corneocitului unde reprezinta aprox. 50%
din totalul proteinelor celulare. Citocheratina are tendinta de a se dispune spre periferia citoplasmei.
2. Citocheratina se insolubilizeaza si se dispune ca un invelis subplasmalemal si va aparea deci o ingrosare a membranei celulare. Acest invelis este maxim la nivelul corneocitelor dar este vizibil de la nivelul........
3. Sinteza de cheratohialina debuteaza in cheratinocitul spinos superficial iar cea mai mare cantitate se gaseste in cheratinocitul granular. Cheratohialina se dispune sub forma de granule bazofile care nu au endomembrane.
4. In cheratinocitul spinos superficial incepe formarea de corpi lamelari care sunt delimitati de membrane derivate din aparatul Golgi. La nivelul cheratinocitului granular continutul corpilor lamelari este eliminat la exterior. Acestia contin material fosfolipidic. Eliminarea se face spre stratul cornos, deci materialul fosfolipidic se dispune intre corneocite, formand straturi lamelare. Acest material fosfolipidic are rol de a mentine adezivitatea celulelor si actioneaza ca o bariera pentru apa (impiedica evaporarea apei).
5. Are loc sinteza de filagrina. Proteina este sintetizata ca profilagrina in cheratinocitul granular. Ea se va transforma in filagrina matura in corneocit.
15

6. Urmatoarea etapa consta in sinteza cheratinei. Mai intai citocheratina se impregneaza cu cheratohialina si se formeaza a-cheratina. a-cheratina se combina cu filagrina si rezulta cheratina moale. Acum apar si modificari ale organitelor celulare. Acestea se degradeaza sub actiunea enzimelor lizozomale si dispare si nucleul. Astfel, procesul de cheratinizare este considerat o apoptoza speciala.
Dupa procesul de cheratinizare, celula devine o masa proteica inconjurata de corpi lamelari. Acest strat are mai multe roluri:· asigura rezistenta mecanica· reprezinta componenta epidermului cu rol de bariera: impiedica patrunderea microorganismelor
la nivelul epidermului· este impermeabila pentru apa
Celulele superficiale se pierd, se descuameaza, fiind celule care acumuleaza o mare cantitate de fosfataza acida.
Psoriazisul este o dermatoza eritemato-scuamoasa destul de frecventa. Are loc inrosirea pielii (datorita vasodilatatiei locale) si se formeaza cruste in centru. Boala are perioade de acutizare si perioade de remisiune. Zonele afectate sunt in special zonele supuse unor presiuni (coate, genunchi, incheieturile degetelor). Acutizarile apar in special in perioade de stress. Cauzele acestei boli sunt modificari ale procesului de cheratinizare, in zonele afectate:1. Are loc extinderea compartimentului proliferativ de la cheratinocitele bazale si la cele spinoase.
Prin urmare, apare o populatie de celule mai numeroasa.2. Are loc reducerea ciclului de viata al celulei din epiderma la aprox. 7 zile fata de 20-45 de zile
cat este in mod normal.
TIPURI DE CELULE EPITELIALE
Clasificarea celulelor epiteliale se face in functie de aspectul morfologic, care se afla in stransa legatura cu functia celulei. Astfel exista:1. celule epiteliale specializate pentru transport2. celule epiteliale specializate pentru sinteza si secretia de molecule3. celule epiteliale specializate pentru contractie (celulele mioepiteliale)
Celule epiteliale specializate pentru transport Aceste celule poseda mecanisme de transport selectiv de Na+ si K+. Ele au pompe
membranare cu activitate ATP-azica si anumite particularitati structurale.Prototipul acestui tip de epiteliu este nefrocitul din segmentul proximal al tubului
urinifer. Aceasta celula are mai multe particularitati:· La nivelul membranei bazale exista invaginari citoplasmatice dar care nu sunt insotite de
membrana bazala. Astfel este marita suprafata activa membranara. La acest nivel se gasesc pompele cu activitate ATP-azica.
· La nivelul membranei bazale apar si compartimente, ca urmare a numeroaselor invaginari citoplasmatice. In aceste compartimente exista mitocondrii alungite ce prezinta numeroase criste. Ele realizeaza aportul crescut de ATP necesar pompelor.
· La nivelul membranei laterale, in partea sa apicala exista jonctiuni stranse, care impiedica retrodifuzia ionilor pompati (directioneaza fluxul de ioni).
· Si la nivelul membranei laterale exista interdigitatii.· La nivelul membranei apicale exista microvili care maresc suprafata de absorbtie.
Nefrocitul asigura transportul de Na+ din lumen spre mediul intern. Acest proces cuprinde mai multe etape:
16

· Na+ este preluat din lumenul tubului, prin membrana apicala, prin difuziune prin canale ionice, ca urmare a gradientului de concentratie si electric. Ionul de Na+ este insotit si de ionul de Cl-
si de molecule de H2O, prin osmoza.· Ionul de Na+ este pompat in mediul intern (sange), insotit de ionul de Cl- si de molecule de H2O.
Epiteliul tubului urinifer are mai multe roluri:· reprezinta o bariera activa prin care se controleaza concentratia de apa si de ioni.· are loc concentrarea unor medii prin deplasarea apei.
In functie de sensul de deplasare al ionilor au loc procese de absorbtie sau secretie. Atunci cand deplasarea se face in sens apico-bazal au loc procese de absorbtie, iar cand deplasarea se face in sens bazo-apical au loc procese de secretie.
Reabsorbtia de ioni are loc in:· nefrocitele din segmentul proximal al tubului urinifer· nefrocitele din segmentul distal al tubului urinifer· celulele din ductele striate· epiteliul tubului digestiv· peretii colecistului
Secretia de ioni are loc in:· procesele ciliare - ghemuri de capilare care intra in alcatuirea tunicii medii a globului ocular.
Secretia are loc la nivelul celulelor endoteliale ale capilarelor· plexurile coroide - formatiuni din SNC ce reprezinta invaginari ale piamater spre ventriculii
cerebrali. Prezinta un ax conjunctiv si un epiteliu de acoperire care secreta ioni. Ele sunt responsabile de secretia lichidului cefalorahidian (LCR).Deplasarea ionilor se face mai degraba prin spatiul intercelular decat transcelular, adica ionii
patrund in celula prin membrana apicala si, dupa ce au trecut de zona jonctiunilor stranse, ies prin membrana laterala in spatiul intercelular si ajung pe aceasta cale in mediul intern. Acest traseu este preferabil traseului transcelular datorita consumului mai scazut de energie.
Celule epiteliale specializate pentru sinteza si secretieSe pot secreta:
· proteine· amine si proteine mici· glicoproteine · lipide
Celulele epiteliale specializate pentru sinteza si secretia de proteineAceste celule se pot gasi in glande exocrine, endocrine sau paracrine. Celulele au polaritate
morfologica: citoplasma bazala este bazofila iar citoplasma apicala este acidofila sau bazofila dupa natura proteinei secretate.
Cu ajutorul microscopului electronic s-a observat localizarea diferitelor organite dupa cum urmeaza:
· la polul bazal - organitele specializate pentru sinteza proteinelor: ribozomi si reticul endopasmic rugos
· aparatul Golgi este situat supranuclear· granulele de secretie in zona apicala (apar dense la fluxul de electroni)
Proteinele sintetizate pot fi:· enzime digestive (secretate de celulele seroase) - glanda parotida, submandibulara, pancreasul
exocrin· proteine plasmatice - hepatocitul, care secreta albumine, globuline, fibrinogen.
17

· hormoni - paratiroida, care sintetizeaza si secreta parathormon.Sinteza proteinelor este continua.Secretia proteinelor poate fi continua sau intermitenta. In cazul secretiei continue celula nu
face depozite intracelulare de produs de secretie (hepatocitul). In cazul secretiei intermitente produsul de secretie se acumuleaza in granule de secretie care alcatuiesc depozite intracelulare.
Produsul de secretie poate fi eliberat prin canale de excretie (glanda exocrine), intr-un capilar (glanda endocrina) sau in lichidul interstitial (glanda paracrina).
Prototipul celulei care sintetizeaza si secreta proteine este celula din acinul pancreatic.Etapele ciclului secretor in celula pancreatica sunt urmatoarele:
· Asimilarea aminoacizilor de catre celula epiteliala. Celula poate sintetiza proteine numai pe baza aminoacizilor preluati din mediul intern. Preluarea se realizeaza la polul bazal si se face in mod activ, necesitand un transportor care este specific pentru fiecare tip de aminoacid.
· Sinteza lantului polipeptidic la nivelul ribozomilor atasati la reticulul endoplasmic rugos. Moleculele proteice sintetizate vor patrunde in lumenul RE unde va avea loc segregarea (inlaturarea moleculei semnal).
· Acumularea in aparatul Golgi. Transportul de la reticulul endoplasmic la fata convexa a cisternelor golgiene se face cu ajutorul veziculelor sau microveziculelor de transport ce iau nastere prin inmugurire din cisternele reticulului endoplasmic. In aparatul Golgi are loc condensarea, agregarea moleculelor proteice.
· Formarea granulelor de secretie. Granulele se formeaza prin inmugurirea cisternelor aparatului Golgi. Granulele de secretie devin din ce in ce mai dense la fluxul de electroni pe masura ce se apropie de polul apical al celulei. Granula de zimogen este specifica pancreasului exocrin. Granulele de zimogen contin amestec de protein-enzime active si inactive (amilaza, lipaza, ribonucleaza, colesterol esteraza, tripsinogen, procarboxipeptidaza etc)
· Transportul granulelor de secretie. Se realizeaza cu ajutorul elementelor de citoschelet (microtubuli si microfilamente). S-a dovedit experimental ca substantele care distrug citoscheletul au ca efect incetarea migrarii veziculelor. Colchicina duce la depolimerizarea microtubulilor iar citochalazina B afecteaza microfilamentele. Aceste substante determina oprirea granulelor de secretie la nivelul aparatului Golgi.Procesul de migrare este dependent si de consumul energetic, deci o substanta care produce decuplare proceselor de oxido-reducere are ca efect si oprirea veziculelor de secretie.Procesul este independent de intensitatea sintezei proteice. Astfel, cicloheximida, o substanta care blocheaza transferul unui aminoacid de pe ARNt specific pe lantul polipeptidic, ducand la blocarea sintezei proteice, nu afecteaza transportul veziculelor de secretie.
· Eliminarea produsului de secretie care are loc prin exocitoza. Exocitoza are loc numai la polul apical si niciodata prin membrana bazala sau laterala. Are loc o interactiune intre proteinele din endomembrana granulelor cu proteinele din membrana apicala a celulei. Apoi are loc o rearanjare structurala a celor doua membrane care vor fuziona. Granula de secretie se deschide astfel spre exterior si continutul sau este eliminat din celula.
Celule epiteliale specializate pentru sinteza si secretia de amine si polipeptide miciSe mai numesc si celule APUD (Amine Precursor Uptake and Decarboxilation). Notiunea a
fost introdusa de Pearse. Aminele pot fi preluate din circulatie de catre celule sau pot fi sintetizate de catre celule din
precursori, proces realizat de celulele APUD. Aceste celule au un continut mare in aminoacid decarboxilaza.
Ele sunt celule endocrine sau paracrine, au distibutie difuza si sunt de obicei localizate intraepitelial, fiind izolate sau in grupuri mici. Celulele APUD se gasesc de-a lungul tubului digestiv de la esofag pana la canalul anal si in structura canalelor care se varsa in intestin (canalul coledoc si
18

canalul Wirsung). Aceste celule alcatuiesc axa gastro-entero-pancreatica. Ele se mai gasesc si in epiteliul respirator, in structura glandelor tiroida si paratiroida, in aparatul urogenital si in piele.
La microscopul optic citoplasma celulelor APUD este bazofila si celulele sunt greu de diferentiat de restul celulelor. Deci sunt necesare metode speciale de diferentiere:1. Precipitarea cu saruri de Ag (solutie amoniacala de AgNO3). Celulele pot fi argentafine sau
argirofile. Celulele argentafine retin Ag care precipita direct din solutie. Celulele argirofile necesita un reducator din mediul extern pentru a putea retine Ag.
2. Oxidarea cu dicromat de potasiu (K2Cr2O7) pentru celulele cromafine. Celulele enterocromafine se gasesc in tubul digestiv si se pot oxida cu K2Cr2O7.
3. Metacromazia cu albastru de toluidina dar dupa o hidroliza acida.4. Fluorescenta indusa de formaldehida.5. Imunocitochimie - metoda cea mai folosita actual. Se folosesc anticorpi monoclonali cu ajutorul
carora se fac determinari foarte precise ale tipului de celule APUD studiate. Se mai pot folosi markeri, prin intermediul carora se pune in evidenta o populatie de celule APUD. Markeri pot fi: enolaza neuronala, specifica sistemului neuroendocrin, dar care se gaseste si in sistemul nervos); proteina PG P9.5 (Protein Gene Product 9.5), care este o proteina solubila ce apare in celulele APUD cu exceptia celor intestinale si careia nu i se cunoaste functia; cromograninele si secretograninele (initial se gasesc in medulosuprarenala) care sunt proteine solubile ce contribuie la descarcarea granulelor de secretie.
6. Hibridizare in situ: celulele sunt identificate dupa continutul in ARNm specific.In microscopia electronica celulele APUD apar sub doua forme: inchise sau deschise, in
functie de raportul polului apical cu lumenul organului. La celulele deschise, polul apical vine in contact direct cu lumenul. Uneori au si microvili. Celulele inchise au polul apical acoperit de o alta celula epiteliala, deci nu intra in raport direct cu lumenul.
La celulele APUD organitele sunt slab reprezentate. Concentratia de polipeptide este cuprinsa intre 10-6 si 10-9 g/ml. Granulele de secretie sunt aglomerate spre polul bazal al celulei. Granulele de secretie au forma, dimensiuni si densitati la fluxul de electroni diferite. Produsul de secretie este un amestec de substante format din: amine, alaturi de unul sau mai multi hormoni proteici, cromogranine, secretogranine, Ca2+ si ATP. Eliminarea granulelor se face prin polul bazal atunci cand celula este endocrina sau paracrina dar exista si cazuri cand eliminarea se face prin polul apical. Eliberarea se face prin exocitoza discontinua, in functie de stimuli.
Actualmente, notiunea de celule APUD a fost inlocuita de conceptul de Sistem Neuroendocrin Difuz. Se numesc sistem neuroendocrin deoarece aceste celule au proprietati care amintesc atat de neuron cat si de celula endocrina clasica. Sub actiunea unui stimul specific, este eliberat mesagerul chimic care actioneaza in vecinatate sau la distanta (caracter de celula endocrina). Celula prezinta activitate electrica ce creste in timpul eliminarii produsului (caracter de celula nervoasa).
Celule epiteliale specializate pentru sinteza si secretia de glicoproteineSunt celule endocrine sau celule exocrine. Celulele exocrine sunt celulele mucoase: celula
caliciforma, celulele gastrice, celulele din acinii mucosi. Celulele endocrine sunt celulele din adenohipofiza care secreta hormoni tropi: proopiomelanocortina (POMC), gonadotropina etc.
Prototipul celulelor specializate pentru sinteza si secretia de glicoproteine este celula caliciforma. Ea este localizata intraepitelial. La microscopul optic, in coloratie uzuala (hemalaun-eozina), celula caliciforma are un pol apical dilatat, palid, cu aspect vacuolar (deoarece prin coloratie HE se pierd glucidele) si un pol bazal efilat (ingustat). Prin coloratie PAS, glucidele se pastreaza si poate fi examinat polul apical si platoul striat.
In microscopia electronica, dispozitia organitelor explica procesul de sinteza si secretie:· Reticulul endoplasmic rugos se gaseste la polul bazal· Mitocondriile se gasesc intre cisternele RE rugos
19

· Aparatul Golgi se gaseste supranuclear si are o extindere mare· Granulele de secretie se acumuleaza la polul apical pe care il destind, datorita faptului ca sunt
foarte multe si mari.· Exista si cativa microvili
Procesul de sinteza al glicoproteinelor incepe cu asimilarea aminoacizilor din mediul extern, apoi urmeaza sinteza lantului polipeptidic in ribozomii atasati RE rugos. Lantul trece apoi in lumenul RE rugos unde are loc segregarea, dupa care este transportat prin microvezicule la aparatul Golgi. Aici are loc elaborarea si atasarea componentei glucidice la lantul polipeptidic. De asemenea glicoproteina este apoi sulfatata rezultand o glicoproteina sulfatata. De aici, dupa ambalarea in granule de secretie are loc transportul spre membrana apicala cu ajutorul elemnetelor de citoschelet si apoi exocitoza.
Granulele de secretie din celula caliciforma sau alte celule mucoase sunt granule de mucina. Atunci cand are loc fuzionarea endomembranei granulei de secretie cu membrana celulara are loc o modificare a permeabilitatii pentru apa, care va patrunde in granula de secretie si va hidrata continutul acesteia. Astfel se formeaza mucusul. Mucusul este deci mucina hidratata.
Celulele specializate in sinteza si secretia de lipidePot fi celule endocrine sau exocrine. Celulele exocrine sunt cele din glanda sebacee unde
secretia este o mixtura de lipide cu celule dezintegrate. Celulele endocrine sunt celulele care sintetizeaza steroizi: celulele gonadale (celulele Leydig la barbat si celulele foliculare si luteale la femeie), si celulele din corticosuprarenala.
Prototipul celulelor specializate in sinteza si secretia de lipide sunt celulele care secreta steroizi. La microscopul optic, in coloratie uzuala, ele au forma poligonala, cu nucleul central, eucromatic. Citoplasma este acidofila si cu aspect vacuolar datorat picaturilor de lipide pline cu precursor al hormonilor steroici (colesterol esterificat).
La microscopul electronic se observa un continut mare de reticul endoplasmic neted (REN) si mitocondrii cu criste tubuloveziculare. Nu se cunoaste semnificatia acestor criste mitocondriale speciale. Intre mitocondrii si REN exista stranse relatii functionale si structurale. Relatia structurala este data de faptul ca mitocondriile se afla printre cisternele REN. Relatia functionala este data de cooperarea dintre enzimele REN cu enzimele mitocondriale (unele din enzimele necesare procesului de sinteza apartin REN iar altele apartin mitocondriilor). Astfel, sinteza colesterolului din acetat este catalizata de enzime din REN, transformarea colesterolului in pregnenolona (prima etapa in procesul de sinteza propriu-zisa a hormonilor steroici) este catalizata de enzime mitocondriale iar restul etapelor pana la obtinerea diversilor hormoni steroici se parcurg datorita enzimelor din REN.
Celule epiteliale specializate pentru contractie (celulele mioepiteliale)Sunt localizate intre membrana bazala a unei celule epiteliale si membrana bazala (MB) a
tesutului epitelial. Se gasesc la nivelul adenomerilor (alveole sau acini) si al ductelor din anumite glande exocrine: glandele salivare, lacrimale, sudoripare si mamare.
La microscopul optic apar ca niste prelungiri care incercuiesc alveolele, acinii sau ductele. Prezinta putina citoplasma.
La microscopul electronic, in citoplasma se observa:1. existenta filamentelor intermediare de citocheratina, fapt care atesta originea lor epiteliala2. existenta proteinelor contractile (actina, miozina si tropomiozina), organizate in mod asemanator
cu cele din fibra musculara neteda3. existenta jonctiunilor gap si a desmozomilor. Jonctiunile gap se formeaza intre celulele
mioepiteliale alaturate, fapt care asigura actiunea sincrona a celulelor mioepiteliale. Desmozomii se formeaza intre celulele epiteliale si celulele din unitatile secretorii sau din ducte si astfel se produce o tensiune asupra unitatii secretorii si se ajuta la eliminarea granulelor de secretie.
20

Histologie - curs 4TESUTUL CONJUNCTIV
Este alcatuit din:· celule conjunctive· matricea extracelulara (substanta fundamentala si fibre)
Matricea conjunctiva extracelulara este un ansamblu de molecule proteice si polizaharidice secretate local de celulele conjunctive si asamblate intr-o retea foarte bine organizata. In trecut s-a crezut ca este o masa inerta cu rol de stabilizare. Acum matricea este considerata un mediu extracelular in care celulele migreaza, prolifereaza, cresc, se diferentiaza, capata forma si functie. Ele au nevoie de macromoleculele matricei conjunctive. Pe macromolecule se fixeaza factorii de crestere si hormonii care formeaza un ansamblu masiv de semnale pentru celulele conjunctive.
Toate macromoleculele au un turn-over de reinnoire, deci sunt produse in permanenta si se degradeaza mereu cu ajutorul unor enzime secretate tot de celulele conjunctive. Toate macromoleculele sunt in interactiune cu celulele conjunctive, doar ca unele interactioneaza direct (prezinta receptori specifici pe membrana celulei conjunctive) si altele interactioneaza indirect prin intermediul unor proteine de adezivitate.
21

Dintre receptorii implicati in interactiunea matrice - celula enumeram: integrine, receptori integrine-like si receptorii non-integrine. Proteinele de adezivitate implicate in interactiunea matrice - celula sunt: fibronectina si laminina.
Macromoleculele sunt secretate de majoritatea celulelor conjunctive cu exceptia celulelor sangvine mature. Compozitia moleculelor depinde de tipul de celula, de gradul de diferentiere al celulelor si de statusul metabolic al celulei.
Componentele de baza ale matricei conjunctive sunt proteinele fibroase, glicozaminoglicanii (GAG) si proteoglicanii. Proteinele fibroase alcatuiesc fibrele matricei conjunctive si structurile de adezivitate. Colagenul intra in compozitia fibrelor de colagen si de reticulina iar elastina alcatuieste fibrele elastice. In componenta structurilor de adezivitate intra fibronectina si laminina si aceste structuri mediaza legatura dintre matrice si celule. Proteoglicanii au o structura ramificata pe cand GAG sunt neramificati. GAG sunt structuri polizaharidice cu o incarcatura negativa mare, data de radicalii sulfat si carboxil. Incarcatura negativa este responsabila de atragerea ionilor de Na+ care sunt osmotic activi care vor atrage la randul lor si apa; deci se va realiza o structura de gel puternic hidratat in care faza lichida a gelului este un mediu de vehiculare a hormonilor si metabolitilor pana la nivelul celulei conjunctive. Prin hidratarea matricei de catre GAG si proteoglicani se realizeaza turgescenta tesutului conjunctiv (turgescenta reprezinta umplerea cu apa a unui tesut).
GAG confera matricei rezistenta la compresie iar fibrele conjunctive confera rezistenta la tractiune.
FIBRELE TESUTULUI CONJUNCTIV
Colagenul
Este una dintre cele mai raspandite proteine. Reprezinta 25% din totalul proteinelor structurale si 6% din greutatea corpului (la fel de mult cat cantareste si pielea).
Molecula are un aspect de bastonas cu lungimea de 300 nm, diametrul de 1,5 nm si greutatea moleculara de 28500 daltoni. Molecula este formata din trei lanturi care pot fi de tip a1 sau a2
dispuse intr-un triplu helix. Lantul a are pe fiecare tur de spira cate 3 aminoacizi, deci in total intr-un lant, exista aprox. 1050 aminoacizi. La fiecare al treilea aminoacid se repeta glicina. Deci formula generala este: a = (Y - Gly - X)n, unde Y este hidroxilizina si hidroxiprolina iar X este prolina.
Compozitia procentuala a colagenului este:· glicina - 33%· prolina - 12%· hidroxiprolina - 12%· hidroxilizina - 12%
Mai apar si alti aminoacizi care fac diferenta intre lanturile a1 si a2 si intre tipurile de colagen. Mai pot fi atasate oligozaharide care se fixeaza la nivelul hidroxilizinei. Deci colagenul este o glicoproteina care, in functie de procentul de oligozaharide, poate da o reactie PAS pozitiva mai accentuata sau mai putin accentuata.
Colagenul este de 25 de tipuri care reprezinta combinari ale lanturilor a1 si a2. Dintre acestea, 15 tipuri sunt foarte bine cunoscute d.p.d.v. molecular si functional, iar din acestea, 5 tipuri sunt cele mai raspandite.
Se poate face o clasificare dupa raportul dintre colagen si matrice:· colagen fibrilar, care se organizeaza in fibrile si formeaza fibre de colagen sau reticulina· colagen afibrilar care nu formeaza fibrile deoarece atat la capul amino terminal cat si la cel
carboxi terminal prezinta domenii globulare, care impiedica polimerizarea in fibrile.
22

Tipuri de colagen fibrilarEste colagenul de tip I, II, III, V, IX, X si XI. Colagenul de tip I, II, III, V si XI formeaza
fibrile omogene (contin doar molecule de tip I, doar molecule de tip II etc.). Colagenul de tip IX si X formeaza fibrile heterogene. Colagenul afibrilar se va atasa periodic la fibrilele de colagen si va forma retele bidimensionale in structura laminelor bazale (colagenul de tip IV). Fiecare tip de colagen are in componenta tipuri specifice de lanturi a1 sau a2 (in colagenul de tip I avem lanturi a1(I), in colagenul de tip II avem lanturi a2(II) etc.).
· Colagenul de tip I. Este alcatuit din doua lanturi a1(I) si un lant a2(I) sau din trei lanturi a1(I). Are cea mai larga raspandire (aprox. 90% din tot colagenul). Este localizat in tesutul conjunctiv din piele, tendon, ligament, dentina, sclera, fascia musculara, capsula organului. Este rezistent la intindere si tensiune.
· Colagenul de tip II (colagenul cartilajului). Este alcatuit din trei lanturi a1(II)si se gaseste in cartilajul hialin si elastic, notocord si discurile intervertebrale. El asigura forma si rezistenta la deformare.
· Colagenul de tip III. Este format din trei lanturi a1(III). Se gaseste in stroma conjunctiva a unor organe: uter, ficat, splina, rinichi, plaman, in structura muschiului neted, a vaselor sangvine si in pielea fetala. El intra in componenta fibrelor de reticulina. Are un rol structural si elasticitar.
· Colagen de tip IV. Este format din trei lanturi de tip a1(IV) sau din trei lanturi de tip a2(IV). El reprezinta colagenul amorf din laminele bazale ale epiteliilor, endoteliilor vasculare, epiteliilor din glomerulii renali si din cristalin. Are rol de suport si de bariera de filtrare.
· Colagen de tip V. Este format din doua lanturi de tip a1(V) si un lant de tip a2(V). Este colagenul din laminele bazale ale celulelor musculare striate si netede, laminele bazale ale celulelor Schwann, celulelor gliale si epiteliului placentar. Are rol in principal de suport.
BIosinteza colagenuluiCuprinde doua mari etape, etapa intracelulara si etapa extracelulara.
Etapa intracelulara· Formarea veziculelor de endocitoza prin care isi preia prolina si lizina.· Transcriptia si traducerea
Lanturile a1 si a2 sunt codificate de gene de pe cromozomii 17 (a1) si 7 (a2). Ele sunt copiate pe molecule de ARNm care vor ajunge in citoplasma la nivelul ribozomilor atasati RE. Ribozomii sunt atasati reticulului endoplasmic deoarece se produc proteine de export. Aici are loc traducerea in urma careia rezulta lanturile proa1 si proa2 care sunt foarte lungi si foarte grele (au o greutate moleculara de aprox. 154000 daltoni). Lanturile proa au o portiune centrala, colagenica si doua capete necolagenice (capatul amino terminal si capatul carboxi terminal). Cele doua capete alcatuiesc peptidele de extensie. La capatul amino terminal peptida de extensie se numeste peptida semnal. La capatul carboxi terminal peptida de extensie se numeste peptida extra.
Peptida semnal va fi scindata enzimatic dupa patrunderea in lumenul RE cu ajutorul unei proteaze. Peptida extra are rol in initierea si controlarea spiralizarii lanturilor a in jurul unui ax propriu si in formarea triplului helix.
· Modificari postranslationale· Clivarea secventei semnal cu ajutorul unor proteaze din lumenul RE.· Hidroxilarea reziduurilor de prolina si lizina. Procesul este catalizat de doua enzime:
peptidil-prolin hidroxilaza si peptidil-lizin hidroxilaza. Se formeaza 3-hidroxi-prolina, 4-hidroxi-prolina, 3-hidroxi-lizina si 4-hidroxi-lizina. In afara de aceste enzime mai sunt necesare vitamina C (care are un rol foarte important), ionii Fe, a-ceto-glutaratul si oxigenul.
23

· Glicozilarea polipeptidului, care are loc la hidroxilizina. Se face cu ajutorul enzimelor galactozil transferaza si glucozil transferaza. Se transfera galactoza si glucozil-galactoza la resturile de hidroxilizina.
· Formarea triplului helix de procolagen, proces care este initiat de peptida extra (capatul carboxi terminal al lantului proa. Lanturile a se spiralizeaza in jurul unui ax propriu iar apoi se spiralizeaza unul in raport cu celalalt si rezulta triplul helix. Intre lanturi apar legaturi covalente si de hidrogen care vor stabiliza molecula nou formata, procolagenul. Aceasta molecula are doar partea centrala organizata in triplu helix, partile laterale fiind paralele si spiralizate doar in jurul axei proprii.
· Impachetarea procolagenului in macrovezicule golgiene· Deplasarea macroveziculelor in ectoplasma periferica care se face cu ajutorul
microfilamentelor si al microtubulilor.· Exocitoza procolagenului.
Etapa extracelulara· Formarea tropocolagenului are loc din molecule de procolagen care, ajunse in spatiul
extracelular, sunt clivate sub actiunea unor procolagen peptidaze. Clivarea are loc la capetele nespiralizate in triplu helix, unde se indeparteaza jumatate din capetele nespiralizate si rezulta astfel tropocolagenul.
· In situatia colagenului fibrilar, mai departe va avea loc polimerizarea (asamblarea in fibrile si apoi in fibre). Polimerizarea se desfasoara in felul urmator: la nivelul hidroxilizinei actioneaza o enzima, liziloxidaza care dezamineaza oxidativ hidroxilizina si va rezulta o aldehida reactiva. Apoi se stabilesc legaturi transversale, cross-link-ari, intre moleculele apropiate. Moleculele de colagen se dispun in fibrile cu diametrul intre 20 si 200 nm.La microscopul electronic, fibra de colagen apare ca o miofibrila. Are o periodicitate de
68 nm care curpinde un disc clar si un disc intunecat. In cadrul fibrilei moleculele de colagen se dispun in siruri paralele echidistante. Intr-un sir, intre molecule se gasesc spatii (deschideri, gap-uri) cu largimea de 35 nm. La urmatorul sir apare un defazaj de 68 nm care se va mentine si in sirurile urmatoare. Gapurile se afla la aceeasi distanta o data la cinci siruri.
La microscopul electronic gap-urile se vad intunecate deoarece osmiul sau uraniul se fixeaza la capetele moleculelor de colagen si atunci aceste zone devin dense la fluxul de electroni.
Celule producatoare de colagen:· celulele mezenchimale· fibroblastele· miofibroblastele· celulele perineuronale· cementoblastele (intra in radacina dintelui)· odontoblastele (intra in cavitatea pulpara)· osteoblastele· celulele cartilaginoase· unele celule musculare netede· celulele epiteliale· adipocidele· celulele Schwann· celulele gliale
Fibrele de colagen sunt formate din colagen de tip I, II, V, IX, X, XI. Ele dsunt cele mai groase fibre ale tesutului conjunctiv si au un diametru de aproximativ 20 mm. Ele nu se ramifica si nu se anastomozeaza. Se dispun sub forma de benzi, panglici sau manunchiuri cu fibre paralele
24

care se pot aseza fie in toate directiile formand retele, fie paralel formand tesutul conjunctiv ordonat.
Tehnici de colorare:· coloratie universala - hemelaun-eozina - colagenul apare roz;· coloratii topografice:
· Van Gieson - colagenul apare rosu;· AZAN - colagenul apare albastru;· Mallory - colagenul apare albastru;· Masson - colagenul apare verde
Fibrele de reticulina
Fibrele de reticulina sunt cele mai subtiri, avand un diametru de 1 mm. Se ramifica si se anastomozeaza formand retele. Nu se vad in coloratie HE deoarece exista foarte multe fragmente oligozaharidice care nu fixeaza nici hemalaunul, nici eozina. Fibrele de reticulina se vad in coloratia PAS unde apar rosu violet. Se mai evidentiaza si prin impregnarea argentica deoarece sunt fibre argirofile.
Elastina
Elastina reprezinta 6% din proteinele structurale. Molecula are urmatoarea compozitie:· glicina - 33%:· prolina - 10-13%· putina lizina· 50-60% aminoacizi nepolari de genul dezmozinei si izodezmozinei.
Molecula are o greutate de 72000 daltoni.Dezmozina si izodezmozina sunt responsabile de caracterul puternic hidrofob al moleculei si
de caracterul sau elastic.
Biosinteza elastineiCuprinde doua etape: intracelulara si extracelulara. Sinteza este realizata de aceleasi celule ca
si sinteza colagenului.Etapa intracelulara decurge la fel ca la colagen, doar ca nu are loc hidroxilarea, glicozilarea si
spiralizarea. Molecula rezultata se numeste tropoelastina si are un aspect neregulat, rasucit.In etapa extracelulara tropoelastina va polimeriza. Este nevoie mai intai de o dezaminare a
lizinei realizata cu ajutorul liziloxidazei. Rezulta astfel aldehide reactive intre care se vor forma punti covalente si necovalente. Puntile covalente mentin distantate moleculele de elastina in polimer, atat in intindere cat si in relaxare.
La microscopul electronic are un aspect amorf, astructurat, dens la fluxul de electroni.Sistemul fibrelor elastice este alcatuit din: fibre de elastina, fibre de oxitalan, si fibre de
elaunina. Fibrele de oxitalan si cele de elaunina reprezinta precursori ai fibrelor elastice mature, dar ele se gasesc atat in tesutul embrionar, cat si in cel adult.
Fibrele oxitalanice se gasesc in zone de maxima rezistenta: in ligamentul alveolo-dentar, in zonula lui Zinn, in dermul pielii. Ele sunt alcatuite din numeroase fibrile de fibrilina (glicoproteina care se sintetizeaza in paralel cu tropoelestina).
Fibrele de elaunina se gasesc in dermul pielii, in special in jurul glandelor sudoripare. Sunt alcatuite din microfibrile de fibrilina si cantitati variabile de elastina polimerizata, care se dispune sub forma de spoturi policiclice, disociind microfibrilele de fibrilina. Pe masura ce elastina creste si este eliminata extracelular, microfibrilele de elastina conflueaza si se dispun in
25

mijlocul fibrei. Atunci microfibrilele de fibrilina se dispun radial si atfel se formeaza fibra elastica matura.
Fibrele elastice au un diametru de 5 mm (intermediar intre fibrele de reticulina si cele de colagen). Ele se ramifica si se anastomozeaza. In coloratie HE apar roz. Cu orceina se coloreaza in brun roscat, cu aldehid-fuxina Gömori in negru, iar cu resorcin-fuxina Weigert in rosu.
Glicozaminoglicani (GAG) si proteoglicani
GAG sunt molecule polizaharidice neramificate formate din unitati dizaharidice repetitive care au pe suprafata lor numeroase sarcini negative date de radicalii sulfat si carboxil. O unitate repetitiva este formata dintr-un glucid aminat (N-acetil-galactozamina sau N-acetil-glucozamina) si un acid uronic. Sarcinile negative au urmatoarele roluri:
· Atrag ionii de Na+ care vor atrage la randul lor apa si se va realiza structura de gel puternic hidratat.
· Sarcinile negative se resping intre ele si prin urmare, moleculele de GAG ocupa un volum mare in raport cu greutatea lor moleculara. In spatii se va gasi faza lichida a gelului, fapt care permite mobilitatea celulelor, proliferarea si nutritia lor.
Marea majoritate a GAG, cu exceptia acidului hialuronic, se leaga covalent de miezuri proteice, formand proteoglicani.
Clasificarea GAG se face in 5 clase, in functie de tipul unitatii repetitive, numarul de grupari sulfat si localizarea lor.1. Acidul hialuronic (hialuronan), format din acid glucuronic si N-acetil-glucozamina. Este singurul
nesulfatat, deci sarcinile sale negative vor proveni doar de la gruparile carboxil.2. Dermatan sulfatul, format din acid iduronic si N-acetil-galactozamina3. Heparina (heparan sulfatul), format din acid iduronic si N-acetil-glucozamina4. Condroitin sulfatul, format din acid glucuronic si N-acetil-galactozamina5. Keratan sulfatul este o exceptie, el fiind format din galactoza si N-acetil-glucozamina.
Acidul hialuronicAre cea mai mare molecula (cuprinde aprox. 50.000 unitati dizharidice). Molecula are un
aspect de bastonas cu lungimea de aprox. 1 mm. Acidul hialuronic nu se leaga covalent de un miez proteic.
Este singurul nesulfatat. Prin urmare sinteza sa nu are loc in aparatul Golgi. De asemenea el nu se epimerizeaza in timpul sintezei. Sinteza sa are loc in membrana celulara, alungirea moleculei facandu-se prin adaugarea de oligozaharidice la capetele reduse.
In spatiul extracelular, acidul hialuronic ramane in apropierea plasmalemei, alcatuind glicocalixul. Prin incarcatura negativa mare acidul hialuronic participa la hidratarea mediului extracelular si astfel la migrarea, proliferarea si nutritia celulelor conjunctive.
Acidul hialuronic se gaseste in cantitati mari in tesutul embrionar iar la adult se gaseste in corpul vitros, in lichidul sinovial si in gelatina Wharton.
Alti GAGCeilalti GAG au molecule mai mici (contin aprox. 300 unitati dizaharidice). Ei se
sintetizeaza in aparatul Golgi, unde se sulfateaza, se epimerizeaza si formeaza legaturi covalente cu miezuri proteice, dand nastere la proteoglicani.
Sinteza proteoglicanilor
26

Miezul proteic se sintetizeaza la nivelul ribozomilor atasati reticulului endoplasmic rugos. O data cu patrunderea in lumenul RE, la resturile de serina se ataseaza un trizaharid de legatura format din xiloza-galactoza-galactoza. Apoi lantul peptidic impreuna cu trizharidele de legatura este transportat cu ajutorul veziculeleor de transport la aparatul Golgi. Aici, la nivelul trizaharidelor de legatura, se ataseaza multe unitati dizaharidice la fel sau diferite si astfel ia nastere o molecula foarte ramificata.
Tipuri de proteoglicaniIn matricea cartilaginoasa se gaseste un proteoglican numit agregan. El este alcatuit dintr-un
miez proteic la care se ataseaza mai multe unitati de keratan sulfat si condroitin sulfat. Astfel se formeaza subunitatea A. Mai multe subunitati A se leaga necovalent de o molecula de acid hialuronic prin intermediul unei proteine globulare de link-are.
Proteoglicanii se gasesc in matricea conjnctiva. Ei sunt alcatuiti dintr-un miez proteic pe care se leaga 3-5 molecule de GAG de tipul condroitin sulfat, keratan sulfat sau dermatan sulfat. Acesti proteoglicani se ataseaza periodic la fibrila de colagen intervenind in organizarea matricei conjunctive extracelulare.
Proteoglicanii atasati membranei celulare sunt alcatuiti dintr-un miez proteic reprezentat de proteine dispuse in plasmalema (proteine integrale). Ele ataseaza la domeniul extracelular 5-8 molecule de heparan sulfat.
Rolul proteoglicanilorEste foarte important deoarece moleculele de proteoglicani au specificitate pentru colagenul
individual.· Mediaza adezivitatea celula - celula si celula - matrice.· Mediaza comunicarea intercelulara si comunicarea dintre celula si moleculele matricei
extracelulare.· Controleaza coagularea (la nivelul vaselor)· Controleaza metabolismul lipoproteic (la nivelul vaselor)· Controleaza cresterea musculaturii netede· Controleaza cresterea neuritilor (nervilor)· Controleaza permeabilitatea membranei bazale.
MOLECULELE DE ADEZIVITATE
Sunt impartite in doua categorii: glicoproteine structurale (proteine de ancorare) si receptori de membrana.
Glicoproteine structurale (de ancorare)· fibronectina· tenasceina· laminina· entactina· thrombospondinina· condronectina
FibronectinaEste un dimer proteic cu greutatea moleculara de aprox. 400.000 daltoni, format din doua
subunitati A si B care au fiecare masa moleculara cuprinsa intre 220.000 si 250.000 daltoni. Fiecare subunitate este formata din trei subunitati repetitive (I, II si III).
Unitatea I are intre 45 si 50 de aminoacizi, unitatea II contine 60 de aminoacizi iar in unitatea III se gasesc 90 de aminoacizi.
27

Unitatile I si II alcatuiesc impreuna la capatul amino terminal un locus specific pentru legarea colagenului de tip IV din laminele bazale. Unitatile II si III formeaza impreuna la capatul carboxi terminal un domeniu de legare la proteoglicanii din membrana celulara care au in componenta heparan sulfat.
La capatul carboxi terminal dimerul are subunitatile A si B legate prin punti disulfurice. Partea centrala a moleculei contine mai multe unitati de tip I la nivelul carora vor exista radicali -SH liberi ce vor media polimerizarea dimerilor de fibronectina. Prin polimerizarea dimerilor ia nastere forma multimerica a fibronectinei. Forma dimerica se gaseste doar in plasma iar forma multimerica se gaseste in matricea conjunctiva.
Domeniile de legare ale fibronectinei1. un domeniu de legare pentru fibrinogen sau heparina prin care fibronectina are rol in coagulare;2. un domeniu pentru legarea stafilococului, a gelatinei sau pentru legarea la suprafata celulei;3. doua situsuri pentru cross-link-are la alte proteine ale matricei conjunctive extracelulare;4. situsuri pentru asociere proprie;5. situsuri pentru atasarea de GAG sulfatati.
Histologie - curs 5
LamininaEste un dimer proteic cu masa moleculara de 900.000 daltoni, format din trei subunitati: o
subunitate A cu masa moleculara de aprox 400.000 daltoni si doua subunitati B cu masa moleculara de 200.000 - 250.000 daltoni.
Aceste doua subunitati se asambleaza intr-o molecula cu aspect de cruce. Subunitatea A se dispune la capetele globulare ale fiecarui brat scurt. Ea va forma si bratul scurt vertical in totalitate. Va forma domeniul globular de la capatul bratului lung si axul central al bratului lung.
Cele doua subunitati B realizeaza portiunile filamentoase ale celor doua brate scurte dispuse intre cele doua domenii globulare. Ele se vor dispune sub forma unui dubluhelix in jurul axului central al bratului lung.
Laminina formeaza lamina bazala. Grosimea laminei bazale reprezinta lungimea moleculei de laminina.
Molecula prezinta locusuri speciale. La capatul globular al bratului lung exista un locus pentru legarea colagenului. La domeniile globulare ale bratelor scurte se afla situsuri speciale pentru legarea proteoglicanilor, atat a celor cu heparan sulfat cat si a celor cu molecule mici.
Roluri:· Organizeaza matricea conjunctiva extracelulara.· Intra in structura laminei bazale.
28

CELULELE CONJUNCTIVE
Celulele conjunctive, spre deosebire de cele epiteliale nu prezinta polaritate. Ele intervin activ atat in procesul de secretie al matricei cat si in procesul de degradare a acesteia. Celulele conjunctive secreta enzime lizozomale care vor degrada matricea, care se reinnoieste permanent. Cele mai multe celule conjunctive stabilesc contacte cu matricea conjunctiva: contacte directe sau contacte mediate de receptori sau de glicoproteine de adezivitate.
Clasificarea celulelor conjunctive· D.p.d.v. al provenientei
· celule fixe - stabilesc contacte stranse cu matricea conjunctiva; componentii au proprietatea de interconvertibilitate (un component se poate transforma in altul), proces care se realizeaza pe orizontala sau pe verticala. Procesul realizat pe verticala nu este reversibil.· fibroblast ® fibrocit· condroblast ® condrocit· osteoblast ® ostocit· celula adipoasa· celula musculara neteda (mai ales in peretii vaselor) - este un miofibroblast cu
echipament contractil - microfilamente de actina si miozina.· Celulele libere - nu stabilesc jonctiuni focale cu matricea
· leucocitele granulare (neutrofile, eozinofile, bazofile) - trec prin diapedeza prin peretele capilarelor sau venulelor si ajung in tesutul conjunctiv lax.
· limfocitele B (devin plasmocite)· limfocitele T circulante· monocitele (devin macrofage)· mastocitele· celulele pigmentare
Limfocitele B ajung prin diapedeza in tesutul conjunctiv lax si reticulat si la acest nivel se transforma in plasmocite, celule care intervin in raspunsul imun mediat umoral (secreta anticorpi).
Limfocitele T circulante trec prin diapedeza, ajung in tesutul conjunctiv lax si reticulat. Ele sunt localizate in anumite zone, timodependente, care nu coincid cu zonele ocupate de limfocitele B. La nivelul tesutului conjunctiv lax si reticulat, limfocitele T intervin in imunitatea celulara. Exista mai multe subclase de limfocite T:· Limfocite Th (h de la helper) - limfocite ajutatoare care coopereaza cu
macrofagele si cu limfocitele B in raspunsul imun mediat umoral.· Limfocitele K - intervin in apararea antivirala si antitumorala· Limfocitele T de tip citotoxic - au toxicitate specifica· Limfocitele NC de tip granular care intervin in citotoxicitatea nespecifica
Monocitul trece prin diapedeza in tesutul conjunctiv lax, dens si reticulat si la acest nivel se transforma in macrofag. Macrofagele sunt o populatie heterogena de celule dar cu functii comune: de pinocitoza si fagocitoza. Totalitatea macrofagelor din organismul uman alcatuiesc sistemul fagocitar mononuclear (SFM).
Despre mastocit s-au emis doua teorii. Prima teorie sustine ca mastocitul provine din bazofile circulante, teorie care a aparut ca urmare a numeroaselor similitudini structurale si functionale care exista intre cele doua celule. Cea de-a doua teorie, cea care este acceptata de majoritatea oamenilor de stiinta in zilele noastre,
29

sustine ca originea mastocitului este in celula stem pluripotenta din maduva rosie hematogena, celula care se va diferentia pe linie mastocitara, dand nastere mastoblastului care ulterior se va transforma in mastocit. Rolul mastocitelor este in reactiile de hipersensibilizare imediata: inflamatii, urticarii, alergii si chiar socul anafilactic.
Celula pigmentara are origine nervoasa, in crestele neurale. Este localizata in principal in stroma conjunctiva a irisului (da culoarea lui), in stratul bazal al epidermului si in invelisul epitelial al firului de par. In functie de tipul de pigment, de cantitatea lui si de comportamentul fata de razele luminoase rezulta culoarea ochilor, a pielii si a parului.
· D.p.d.v. functional· Celule implicate direct in producerea si degradarea matricei conjunctive extracelulare -
fibroblastul - celula cea mai activa· Celule cu functie metabolica:
· adipocitul brun· adipocitul alb· adipocitul hepatic - celula Ito, care intervine in sinteza si depozitarea vitaminei A
· Celule reactive (imune)· Celule efectorii:
· celule implicate in fagocitoza:· tip microfag: PMN (neutrofile), bazofile, eozinofile· tip macrofag: celule provenite din monocite care formeaza SFM.
· celule implicate in raspunsul imun mediat umoral - limfocitele B care se vor transforma in plasmocite si care intervin in secretia de anticorpi sau imunoglobuline.
· celule T citotoxice - populatii de limfocite T care secreta perforina ce ataca specific si distruge celulele (intervine in citotoxicitatea specifica).
· celule NK de tip limfocitar - sunt limfocite de tip granular cu origine comuna cu limfocitele B si T in celula stem pluripotenta hematogena, celule care au dimensiuni mult mai mari decat limfocitele B sau T. Ele au in citoplasma un continut granular bogat. Ele vor ataca si distruge nespecific diferite celule.
· Limfocitele K (killer) - sunt o subpopulatii de limfocite T care ataca si distug celulele care au pe suprafata lor anticorpi, ele intervenind in apararea antivirala si antitumorala.
· Celule imune accesorii· macrofagul - incearca sa digere mici cantitati de antigen; activeaza
limfocitele Th si coopereaza cu limfocitele B· celule care prezinta antigen - limfocite Th; ele ajuta macrofagele,
contribuie la elaborarea raspunsului imun umoral.
Originea celulelor conjunctiveExista doua celule ce pot fi originea celulelor conjunctive: celula mezenchimala
nediferentiata si celula stem pluripotenta hematogena.Celule care se formeaza din celula mezenchimala nediferentiata:
· fibroblastul· adipocitul· condroblastul ® condrocitul
30

· osteoblastul ® osteocitul· celulele endoteliale (celule epiteliale)· celulele mezoteliale (celule epiteliale)
Celule care se formeaza din celula stem pluripotenta hematopoietica· mastocitele· PMN (neutrofilele - au un singur nucleu segmentat in mai multi lobi, eozinofilele, bazofilele)· hematiile· megacariocitele (celule care se vor fragmenta si vor da nastere trombocitelor = plachetele
sangvine)· limfocitele care se vor transforma in plasmocite· monocitele care se vor transforma in macrofage si vor alcatui sistemul fagocitar mononuclear,
alcatuit din: · osteoclaste· macrofagele libere din tesutul conjunctiv (histiomacrofage)· macrofagele fixe (histiocitele)
Celula mezenchimala embrionaraEste prezenta in principal in tesutul embrionar dar si in tesuturile conjunctive adulte in
apropierea vaselor de sange. Au originea in mezodermul embrionar sau in mezoectoderm pentru celulele din extremitatea cefalica.
Celula are urmatorul aspect la microscopul optic:· celula are 20-30 mm, forma neregulata cu prelungiri· nucleul ocupa mai mult de jumatate din citoplasma· blastele au citoplasma bazofila din cauza RE si ribozomilor foarte dezvoltati ca urmare a sintezei
intense de proteine· nucleul este rotund sau ovalar si eucromatic (palid colorat)· citoplasma este slab bazofila
La microscopul electronic celula mezenchimala apare astfel:· are prelungiri· raportul nucleoplasmatic este supraunitar· nucleul prezinta 1-2 incizuri (identatii). La nivelul lor se dezvolta organitele celulare: RE si
mitocondrii· nucleul este incarcat cu eucromatina si are un nucleol excentric· in citoplasma apar grupari poliribozomale bine dezvoltate.· mitocondriile, RE rugos si aparatul Golgi din citoplasma sunt slab reprezentate
Celula mezenchimala are urmatoarele roluri· Reprezinta originea celor mai multe celule conjunctive fixe· Secreta matricea conjunctiva extracelulara care este lipsita de fibre in acest stadiu· Induce diferentierea epiteliilor
FibroblastulFormeaza o familie mare de constituienti celulari: fibroblastele, fibrocitele si
miofibroblastele.FibroblasteleSunt cele mai active si mai raspandite celule ale tesutului conjunctiv. Au originea in celula
mezenchimala.Au urmatorul aspect la microscopul optic:
· au forma alungita dar neregulata, cu numeroase prelungiri· au raportul nucleo-plasmatic supraunitar
31

· nucleul are forma ovalar-alungita, este palid colorat si prezinta un nucleol evident localizat in zona mai bombata a celulei
· citoplasma este puternic bazofilaLa microscopul electronic apar urmatoarele particularitati:
· celula stabileste contacte focale cu matricea dar si jonctiuni gap cu alte celule· in citoplasma se evidentiaza foarte multi ribozomi liberi (poliribozomi) si RE rugos· exista foarte multe mitocondrii, aparatul Golgi este bine reprezentat, cu alte cuvinte, intreg
aparatul de sinteza proteica este foarte bine dezvoltat· citoscheletul este de asemenea bine dezvoltat, format din microfilamente de actina si miozina,
microtubuli si filamente intermediare de desmina si timetina.Fibroblastele au urmatoarele roluri:
1. Fibroblastul secreta toate macromoleculele matricei conjunctive: tropocolagen, tropoelastina, fibrilina, glicozaminoglicani si proteoglicani precum si toate cele sase proteine de adezivitate.
2. Fibroblastele secreta proenzimele lizozomale de tipul colagenazei, elastazei, hialuronidazei, glucuronidazei si catepsinei C care intervin in digestia matricei conjunctive.
3. Fibroblastele secreta factorul de proliferare (stimuleaza dezvoltarea microcirculatiei)4. Fibroblastele intervin in secretia de interferon g, care este mult mai activ decat interferonul
secretat de macrofage. El intervine in apararea antivirala si antitumorala.5. Modulatia: fibroblastele sunt interconvertibile atat pe orizontala cat si pe verticala.6. Nonechivalenta: fibroblastele difera intre ele in functie de tipul de tesut conjunctiv caruia ii
apartin si chiar in cadrul aceluiasi tip de tesut conjunctiv, in functie de localizare. O dovada in acest sens avem daca se transplanteaza bucati de derm din pielea palmara sau plantara in pielea de la nivelul trunchiului. Pielea trunchiului se va ingrosa ca urmare a fibroblastelor care sunt diferite si induc diferentele epiteliale
FibrociteleSunt celulele in repaus ale tesutului conjunctiv. Ele se gasesc in majoritatea tesuturilor
conjunctive.La microscopul optic se observa:
· celula are dimensiuni de 15-20 mm si are forma fuziforma, cu capetele bifurcate sau trifurcate· au nucleu alungit, hipercrom· citplasma este acidofila
La microscopul electronic observam in plus:· nucleul are heterocromatina· in citoplasma aparatul de secretie proteica este slab dezvoltat, deoarece celula este in stare de
repaus· sunt bine reprezentate organitele digestiei (lizozomii)· la nivelul membranei plasmatice se observa receptori pentru lipoproteine cu densitate joasa
(colesterol). Fibrocitele intervin astfel in metabolismul colesterolului: controleaza depunerea si sinteza lui.
MiofibroblasteleD.p.d.v. structural se aseamana cu fibroblastele la care se adauga multe filamente de actina si
miozina. Este localizat:· in tesutul de granulatie din apropierea plagilor· in axul vilozitatilor intestinale· in albuginea testiculului· in peretele foliculilor ovarieni· in capsula si septurile splenice
32

Un tip special de miofibroblaste este asa-numitul pericit al lui Rouget, care se afla in peretele capilarelor, intr-o dedublare a membranei.
MastocitulCu privire la originea acestor celule s-au elaborat doua teorii, enuntate mai sus (pag. 2).Ele sunt localizate in dermul pielii sau in mucoasa si submucoasa tubului digestiv sau a cailor
respiratorii.La microscopul optic mastocitul apare in urmatorul fel:
· are forma rotunda sau ovalara cu diametrul de 20-30 mm.· nucleul este rotund, excentric, palid colorat, veziculos si prezinta un nucleol evident.· are putina citoplasma si este slab acidofila. Acidofilia este mascata de prezenta in citoplasma a
numeroase granule mastocitare. Granulele pot fi ortocromatice (se coloreaza in aceeasi culoare pe care o are si colorantul folosit) sau metacromatice (atunci cand se coloreaza prezinta alta culoare decat colorantul folosit - schimba culoarea colorantului). Se folosesc coloranti cationici de anilina: albastru de toluidina - granulatiile ortocromatice se vor colora in albastru iar granulatiile metacromatice se vor colora in rosu-violet.La microscopul electronic mai observam:
· nucleul care aparea palid colorat la microscopul optic aici este incarcat de eucromatina; prezinta un nucleol excentric
· citoplasma are organitele comune slab reprezentate, intreaga citoplasma fiind ocupata de granule opace la fluxul de electroni ce au dimensiuni variate (intre 0,2-2 mm).
· granulele mastocitare au urmatorul continut:· proteoglicani cu heparan sulfat (subpopulatie a heparinei cu rol anticoagulant) sau cu
condroitin sulfat (mastocitele cu condroitin sulfat se numesc mastocite mucoase)· histamina· proteaze neutre· factorul chemotactic al eozinofilelor, responsabil de socul anafilactic.
· membrana celulara prezinta pseudopode cu citoschelet cortical alcatuit din filamente de actina. Acest citoschelet este bine reprezentat.
· la nivelul membranei s-au identificat receptori IgE (reagine). Aceeasi receptori se identifica si in membrana bazofilelor
· la nivelul membranei mastocitelor se evidentiaza sinteza leucotrienelor, care ajuta la formarea complexului antigen-anticorp [Ag-Ac] pe suprafata mastocitului.
Mastocitele au mai multe roluri, in special in reactiile de hipersensibilizare imediata, dintre care cea mai de temut este socul anafilactic, o urgenta medicala. Intr-un soc anafilactic se realizeaza un edem insotit de bronhospasm. Substante care induc socul sunt veninul de viespe, penicilina sau antitoxina antitetanica. In cazul in care un organism vine in contact pentru prima data cu un astfel de alergogen, se va induce secretia de IgE prin plasmocite. IgE au receptori specifici in membrana mastocitelor si a bazofilelor.
La un al doilea contact al organismului cu acelasi antigen, antigenele vor gasi anticorpi deja formati si se vor fixa pe ei. Deci, pe membrana mastocitului vor exista complexe [Ag-Ac], care vor determina doua reactii:1. degranulare mastocitara cu sau fara mastocitoliza2. formare de leucotriene in membrana mastocitului
Degranularea mastocitara cu mastocitolizaEste o urmare a faptului ca nou formatele complexe [Ag-Ac] din membrana mastocitara vor
declansa un lant de mecanisme dupa cum urmeaza: este activata adenilat ciclaza care va duce la fomarea de AMPc. AMPc activeaza o proteinkinaza care va determina deschiderea unor canale de calciu. Ca urmare a deschiderii canalelor de calciu, creste influxul de calciu fapt care determina
33

fuzionarea granulelor mastocitare intre ele si apoi cu membrana celulara. Astfel granulele isi vor varsa continutul la exterior, fapt care poate sa duca sau nu la degradarea celulei in intregime.
Histamina determina cresterea permeabilitatii capilare. Astfel, prin peretele capilar va exista un aflux mare de eozinofile si neutrofile care sunt atrase prin chemostatism pozitiv de catre factorul chemotactic al eozinofilelor care se afla in granulele mastocitare si care acum a fost imprastiat in lichidul interstitial. Neutrofilele si eozinofilele genereaza mastocitoliza in lant care va duce si la eliberarea mai multor granule mastocitare.
Alt efect consta in contractia muschilor netezi din peretii bronhiilor si bronhiolelor mergand pana la bronhospasm si moarte daca nu se intervine cu un bronhodilatator puternic: adrenalina.
Formarea de leucotriene in membrana mastocituluiComplexul [Ag-Ac] stimuleaza fosfolipaza din membrana celulara care va induce sinteza de
acid arahidonic din care se sintetizeaza leucotriene. Leucotrienele pe masura ce se produc se elibereaza in matricea extracelulara. Ele au un rol antagonic histaminei: relaxeaza musculatura neteda. Cam la 1-2 ore dupa reactie, histamina este anihilata prin leucotriene de catre mastocit si prin histaminoxidaza de catre eozinofile.
Mastocitele mai sunt denumite si celule paracrine deoarece actioneaza local prin produsul de secretie eliberat in afara celulei.
MacrofagulAre originea in monocitele circulante.Totalitatea macrofagelor formeaza sistemul fagocitar mononuclear care prezinta trei
compartimente:1. compartimentul medular2. compartimentul sangvin3. compartimentul tisular.
Compartimentul medular este format din celule stem pluripotente care se diferentiaza pe linie monocitara in monoblast ® promonocit ® monocit.
Compartimentul sangvin este format din totalitatea monocitelor circulante.Compartimentul tisular este format din doua tipuri de celule:
1. macrofagul liber (histiomacrofag)2. macrofagul fix (histiocit)
La nivel tisular ele se afla in dermul pielii, in mucoasa digestiva si in cea respi ratorie. Ele se grupeaza de regula in apropierea vaselor de sange.
HistiocitulEste o celula cu dimensiuni intre 10-15 mm, cu nucleu tahicromatic, excentric, cu citoplasma
acidofila, mai mult sau mai putin omogena.
HistomacrofagulEste o celula cu dimensiunile intre 30-40 mm, cu nucleu veziculos excentric, cu nucleol
evident. Citoplasma este acidofila si puternic granulata.La microscopul electronic, histiomacrofagul se prezinta in felul urmator:
· nucleu reniform, situat excentric, incarcat cu eucromatina si cu nucleoli evidenti· citoplasma contine organite comune: mitocondrii, RE rugos, complex Golgi, lizozomi· citoplasma mai contine numeroase vezicule clare (pinozomi) sau opace (fagozomi), in functie de
tipul antigenului (lichid sau solid).· exista si fagolizozomi si pinolizozomi· exista de asemenea si corpi reziduali
Macrofagele au urmatoarele functii:
34

1. Fagocitoza si pinocitoza. Macrofagul fagociteaza particule mari. Fagocitoza poate fi specifica sau nespecifica. Ea este mediata de receptori.
2. Secreta proenzime lizozomale de tipul colagenazei, elastazei, erochinazei, hialuronidazei sau glucuronidazei. Enzimele intervin in digestia matricei conjunctive.
3. Secreta citokine care activeaza la randul lor alte celule (sunt un fel de secretii paracrine). Dintre citokine, interleukina 1 activeaza limfocitele Th (helper), care vor interveni in cooperarea din cadrul raspunsului imun mediat umoral. Interleukina 1 mai activeaza si celulele cu citotoxicitate specifica sau nespecifica.
4. Secreta interferonii a, b, g. Interferonul g este cel mai important, avand rol in apararea antivirala si antitumorala.
5. Secreta factorul de proliferare al fibroblastelor, inducand fibroza in tesuturile in care se gasesc.
Componentii sistemul fagocitar mononuclear1. Macrofagele de tip histiomacrofag si histiocit din tesutul conjunctiv lax si dens aflat in dermul
pielii si mucoasele digestiva si respiratorie.2. Celulele Kupffer din ficat3. Macrofagele alveolare, numite celule cu praf sau prafoase, din plaman.4. Macrofagele din tesutul conjunctiv reticulat din splina, foliculii limfatici, maduva rosie
hematogena si timus5. Macrofagele pleurale si peritoneale din seroasele cavitare6. Osteoclastele din tesutul osos7. Celulele gliale din SNC, numite microglii8. Celulele Langerhans din epiderm.
PlasmocitulAre originea in limfocitele B circulante.Sunt localizate: in dermul pielii, in mucoasele si submucoasele digestive si respiratorii.La microscopul optic plasmocitul apare in felul urmator:
· celula ovalara cu dimensiunea de 30 mm· nucleu rotund sau ovalar dispus excentric, in jurul caruia se observa un aspect de cadran de ceas
sau roata cu spite.· are nucleoli evidenti· citoplasma este abundenta si in cea mai mare parte bazofila; o mica zona din apropierea
nucleului ramane mai palid colorata sau acidofila.· in zonele in care citoplasma este bazofila, exista mici vacuole mai palid colorate sau cu tenta
acidofila. Aceste zone sunt corpusculii lui Russel (lanturi polipeptidice aflate in trecere prin RE rugos)La microscopul electronic observam:
· membrana plasmocitara prezinta microvili· nucleul apare rotund, excentric, cu o alternanta regulata de eucromatina (spatiile dintre spite) si
heterocrmatina (spitele rotii)· in citoplasma abundenta se afla RE rugos responsabil de bazofilie. Unde RE rugos lipseste
citoplasma este acidofila (in jurul nucleului unde se gasesc aparatul Golgi, mitocondrii si elemente de citoschelet).Rolurile plasmocitului sunt:Sintetizeaza anticorpi denumiti imunoglobuline. Imunoglobulinele secretate de plasmocit
sunt:· IgG - imunoglobulina libera in plasma, fixata pe receptorii specifici de pe suprafata
limfocitelor B. Pot interveni in procesul de axonizare a antigenelor.· IgM este prezenta pe suprafata limfocitelor B unde se fixeaza pe receptorii specifici.
35

· IgA este secretata de plasmocite in forma dimerica si apoi este transferata celulelor epiteliale din epiderm unde se fixeaza pe receptorii specifici intervenind in procesele de aparare locala la nivelul pielii.
· IgD - se fixeaza pe receptorii specifici de pe suprafata limfocitelor B.· IgE (reagine) - are receptori specifici pe mastocite si bazofile si intervine in reactiile de
hipersensibilizare imediata.
Raspunsul imun mediat umoralPoate fi de doua feluri: primar sau secundar.
Raspunsul imun umoral primarOrganismul vine in contact cu un anumit antigen pentru prima data. Acum limfocitele B se
vor blastiza (celula adulta redevine celula tanara) si are raportul nucleoplasmatic supraunitar. Nucleul este eucromatic, activ, citoplasma este bazofila, cu aparatul de sinteza proteica foarte bine dezvoltat. Aceste limfoblaste se vor divide intens si se vor diferentia pe doua linii celulare:1. linia limfoblaste ® limfocite cu memorie2. linia limfoblaste ® plasmoblaste ® plasmocite.
Plasmocitele vor sintetiza anticorpi.In raspunsul primar mediat umoral toate aceste evenimente au nevoie de celule imune
accesorii (macrofage, limfocite Th).
Raspunsul imun umoral secundarAre loc la al doilea contact al organismului cu un anumit antigen. In acest caz nu mai este
necesara prezenta celulelor imune accesorii pentru ca exista limfocite B cu memorie care se blastizeaza si rezulta limfoblaste cu memorie care se vor diferentia pe doua linii celulare:1. linia de limfocite B cu memorie2. linia limfoblast ® centrobalst ® centrocit
Centroblastul (centrocitul) este un plasmoblast (plasmocit) dispus in centrul germinativ al foliculului limfatic secundar.
Neutralizarea antigenelelor poate avea loc dupa cum urmeaza:1. Direct - prin aglutinare, precipitare, insolubilizare si liza.2. Prin activarea complementului seric (ansamblu de 15 proteine serice care intervin in distrugerea
antigenelor).3. Prin activarea celulelor care intervin in reactiile de hipersensibilizare imediata.
Celulele pigmentare (melanocitele)Sunt celule care produc pigmenti. Celula melanofora este o celula care depoziteaza pigmentii
produsi de melanocite.La microscopul optic celula pigmentara apare in felul urmator:
· are aspect de fulg de nea sau floare de gheata· nucleul este rotund si dispus central· citoplasma are granulatii brune care nu necesita coloratie speciala. Granulele se numesc
melanozomi.La microscopul electronic mai putem observa in plus:
· sinteza de melanina porneste de la aminoacidul tirozina: tirozina ® 3, 4 dihidroxifenil alanina (DOPA) ® melanina. Prima etapa a sirului de reactii este catalizata de enzima tirozinaza.
· melanozomii cu granule de melanina (pigmentul) pot fi primari, secundari, tertiari sau cuaternari.· melanozomii primari, secundari si tertiari se gasesc in celula pigmentara· melanozomii cuaternari se gasesc in cheratinocite (celulele melanofore)
36

Trecerea pigmentului din celula pigmentara in cheratinocit se face prin secretie citocrina. Dispersia pigmentului este data de ACTH.
Histologie - curs 6
Varietati de tesut conjunctiv
· Tesuturi conjunctive propriu-zise:· tesuturi conjunctive laxe
· propriu-zis· special care este de mai multe feluri:
· lamelar· areolar (fenestrat)· spinocelular
· tesuturi conjunctive fibroase (dense):· semiordonat· ordonat
· Tesuturi conjunctive cu functii speciale· tesut conjunctiv reticulat:
· mieloid· limfoid
· tesut conjunctiv elastic· tesut conjunctiv adipos
· alb· brun
· tesut mucoid· tesuturi conjunctive scheletale:
· tesut cartilaginos:· hialin· elastic· fibros
· tesut osos· lamelar haversian· lamelar trabecular
· articulatii
37

Tesuturi rile conjunctive laxe1. propriu-zise2. speciale:
3. lamelar (fasciile)4. areolar sau fenestrat (epiploon)5. spinocelular (corticala ovarului, endometru)
Tesutul conjunctiv lax propriu-zisEste tesutul cel mai raspandit in organismul uman:
6. Formeaza mucoasele impreuna cu epiteliile de acoperire si cele glandulare. In structura mucoaselor el reprezinta corionul.
7. Il mai gasim in stroma conjunctiva a organelor parenchimatoase (tesutul conjunctiv lax reprezinta scheletul iar umplutura este reprezentata de celulele epiteliale).
8. Se mai gaseste in axul conjunctivo-vascular al seroasei peritoneale.9. in axul conjunctivo-vascular al vilozitatilor intestinale10. in adventicea tubului digestiv si a vaselor de sange11. in regiunea superficiala a dermului (dermul papilar).12. in structura capilarului13. in endotenoniu14. in peritenoniu15. in endomiusium16. in perimisium17. in endonerv18. in perinerv
In structura lui se gasesc in proportii egale fibre, celule si substanta fundamentala.Celulele pot fi fixe sau mobile.
19. celulele fixe sunt fibroblastele si fibrocitele20. celulele mobile sunt celule imuno-reactive (mastocitelele, macrofagele, plasmocitele, celulele cu
efect citotoxic) sau celule accesorii, provenite din sange (limfocitele TH si macrofagele). Prin aceste celule mobile tesutul intervine in imunitate, in apararea locala atat in RI celular cat si umoral.Fibrelele sunt de toate categoriile (colagen, elastina, reticulina) in proportii aproximativ
egale. Fibrele de colagen se dispun in benzi sau panglici. Ele sunt acidofile si se coloreaza in roz. Benzile sunt formate din fibre de colagen dispuse paralel intre ele. Benzile se dispun ulterior in diferite ungiuri fara sa se ramifice sau sa se anastomozeza. Fibrele de elastina se ramifica si se anastomozeaza formand retele cu ochiuri neregulate. Si ele sunt acidofile si se coloreaza in roz. Fibrele de reticulina nu se vizualizeaza in coloratie HE datorita resturilor zaharidice. Ele se pot evidentia prin impregnare argentica sau reactie PAS pozitiva. Aceste fibre sunt ramificate si anastomozate. In ochiurile retelei se dispun celule si substanta fundamentala.
Substanta fundamentala are un procent mare de GAG nesulfatati. De asemenea mai contine si PG mai ales cei cu heparan sulfat si cei cu condroitin sulfat, keratan sulfat si dermatan sulfat. Datorita continutului mare in GAG si PG ea se coloreaza palid in HE. Substanta fundamentala mai contine glicoproteine de adezivitate (fibronectina, entactina si tenasceina).
Vascularizatia este foarte bogata, se face prin vase de calibru minim (capilare, arteriole, venule).
Inervatia este bogata, prin fibre vasomotorii si fibre senzitive, sub forma de terminatii nervoase libere sau terminatii incapsulate.
Tesutul conjunctiv lax lamelar
38

Este bogat in fibre de colagen si elastina dispuse in lamele paralele. Au celule de tip fix (fibroblaste si fibrocite). Se gaseste in fascia superficiala si profunda a muschilor.
Tesutul conjunctiv lax areolar (fenestrat)Este bogat in fibre de colagen care formeaza o retea cu ochiuri foarte largi (de 2-3 ori mai
largi decat in tesutul conjunctiv lax propriu-zis). Celulele sunt fixe sau mobile. Se gaseste in epiploon.
Tesut conjunctiv lax spinocelularSe gaseste in corticala ovarului, in jurul foliculilor ovarieni, in endometru si in jurul
glandelor uterine. Se caracterizeaza printr-o predominata celulara (celule mezenchimale si fibroblaste). Au putina substanta fundamentala si fibre subtiri de reticulina. Celulele se dispun in vartejuri in jurul formatiunilor foliculare.
Tesuturile conjunctiv densTesutul conjunctiv dens semiordonat (fibros)Localizare:
21. in dermul pielii (in partea profunda - dermul reticular)22. in capsulele conjunctive ale organelor23. in structura scleroticii si a corneei24. in epitenoniu25. in epimisium26. in epinerv27. in structura durei mater28. in submucoasa organelor tubului digestiv
Organizare: predomina fibrele de colagen si elastina, putina substanta fundamentala si putine celule.
Fibrele de colagen sunt organizate in benzi (panglici) care contin fibre de colagen paralele si apoi benzile sunt dispuse in toate directiile si chiar sunt ondulate. Fibrele elastice se dispun si ele in toate directiile ramificandu-se. Ele constituie o pasla pentru fibrele de colagen.
Substanta fundamentala este putina si nu se coloreaza in HE datorita prezentei abundente a GAG si PG.
Celulele sunt fibroblaste si fibrocite (celule fixe) dar si macrofage, mastocite, plasmocite, neutrofile.
Vascularizatia este bogata si se face prin vase de calibru minim.Inervatia este bogata si se face prin fibre vasomotorii si fibre senzitive (terminatii nervoase
libere dar mai ales terminatii nervoase incapsulate).Rolul acestui tesut: in general asigura rezistenta la tractiuni.
Tesutul conjunctiv dens ordonatIntra in structura aponevrozelor si tendoanelor. Ambele formatiuni sunt inextensibile si
asigura insertia muschiului pe os. Aponevroza este pentru muschii lati iar tendonul este pentru muschii lungi (ai membrelor).
AponevrozaAponevroza poate fi simpla sau compusa. Cele simple sunt alcatuite din 2 foite conjunctive
suprapuse iar cele compuse sunt alcatuite din 3 foite conjunctive suprapuse.Organizare: predomina fibrele de colagen care sunt paralele intre ele intr-o foita si sunt
perpendiculare pe fibrele din foitele supra si subiacente. Substanta fundamentala este putina si celulele sunt reprezentate de fibroblaste cu forme extrem de variate datorita spatiilor mici dintre
39

fibrele conjunctive. Au o structura avasculara (hranirea se face prin difuziune) iar inervatia este senzitiva si se face prin terminatii nervoase libere.
TendonulTendoanele primare si secundare se unesc si formeaza tendonul tertiar (compus).Tendonul primar - pe sectiune longitudinala se observa ca fibrele de colagen sunt dispuse paralel in benzi care
la randul lor sunt dispuse paralel. Spatiile dintre fibre sunt mici si contin substanta fundamentala si celule. Celulele sunt fibroblaste si fibrocite si se numesc tenocite. Ele au niste prelungiri in forma de aripioare si se mai numesc celule aliforme. Ele apar dispuse metameric, in siruri paralele. Ele sunt extrem de aplatizate si mai au o caracteristica: bigeminismul (doua celule fiice rezultate prin diviziune sunt asezate una langa alta datorita imposibilitatii de deplasara).
- pe sectiune transversala tendonul primar are forme ovalare sau poligonale. Aici se observa aspectul celulelor aliforme. Prelungirile lor imbraca fibrele de colagen marind rezistenta. Fiecare tendon primar este invelit la periferie de endotenonium (tesut conjunctiv lax propriu-zis avascular?).
Mai multe tendoane primare formeaza tendonul secundar care este invelit de peritenonium, care este foarte bine vascularizat. Hranirea tendonului se face pe seama peritenoniului cu rol de asemenea in cresterea in lungime si in grosime. In peritenoniu se mai gasesc corpusculii incapsulati Pacini.
Mai multe tendoane secundate formeaza tendonul compus (tertiar). Epitenoniul inveleste la exterior tendonul compus. Este tesut conjunctiv dens semiordonat. Intre epitenoniu si tendon mai exista o lama de tesut conjunctiv lax iar in afara se afla mezotenonul. Mezotenonul este format din tesut conjunctiv lax si mezoteliul. Mezoteliul este o teaca sinoviala care se dedubleaza din loc in loc si formeaza bursele sinoviale in care se afla lichidul sinovial. Lichidul sinovial contine apa, ioni, proteine, GAG si PG.
Jonctiunea-tendon os: se realizeaza diferit in functie de locul unde se insera tendonul (pe diafiza sau pe epifiza). Diferentele se datoreaza osificarilor diferite ale celor doua straturi osoase.
Jonctiunea tendon-diafiza: fibrele de colagen de la nivelul tendonului strabat periostul diafizei si ajung in corticala diafizara unde se intrepatrund cu fibrele de colagen ale osteoanelor .Acestea sunt fibrele lui Sharpey.
Jonctiunea tendon-epifiza: intre tendon si epifiza se formeaza un tesut conjunctiv cartilaginos hialin de alunecare. Acest tesut se caracterizeaza prin predominanta fibrelor de colagen, substanta fundamentala calcificata si celule cartilaginoase.
Jonctiunea tendon-muschi: Endotenonul se continua cu endomisium, peritenonul cu perimisium si epitenonul cu epimisium. Fibra musculara care ia contact cu tendonul se termina cu prelungiri digitiforme. Fe frontul citoplasmatic al prelungirilor se prind de membrana celulara filamentele subtiri de actina ale ultimului sarcomer. Pe frontul extracelular membrana celulara a prelungirilor digitiforme este dublata de o lamina bazala impreuna cu care formeaza sarcolema. Fibrele de colagen de la nivelul tendonului se prind in portiunile declive ale prelungirilor digitiforme direct pe membrana fibrei musculare in aceste portiuni perforand lamina bazala. Legatura se intareste prin laminina si fibronectina. In jurul fiecarei prelungiri digitiforme se vor localiza circular numeroase fibre de reticulina care vor creste rezistenta jonctiunii miotendinoase. Aceasta jonctiune este foarte activa d.p.d.v. functional. Aici se realizaeaza atat cresterea muschiului cat si a tendonului.
Tesutul conjunctiv elasticAre o raspandire extrem de restransa si anume:
29. ligamentele galbene de la nivelul cefei30. ligamentele articulare31. ligamentul suspensor al penisului.
40

Structura: predomina fibrele elastice dispuse in retea, substanta fundamentala, celule conjunctive (in special celule fixe). Elementele elastice le vom gasi insa larg raspandite in peretele vaselor formate din lamele elastice dispuse concentric. Tot elemente elastice apar si in stroma conjunctiva a plamanului dubland peretele alveolar.
Tesutul mucoidIl gasim in:
32. ordonul ombilical (gelatina Wharton)33. in corpul vitros (in camera vitroasa)
Contine multa substanta fundamentala, bogata in acid hialuronic, cateva celule mezenchimale si fibroblaste si cateva fibre de reticulina.
Tesutul reticulatEste de doua feluri:
1. mieloid (maduva rosie hematogena)2. limfoid (in organele limfoformatoare). Organele limfoformatoare sunt:
3. ggl. limfatici4. pulpa alba a splinei5. placile Peyer din intestin6. amigdaleOrganizare: fibre de reticulina care se dispun sub forma unui fibroreticul, celule conjunctive
(o parte din ele vor forma citoreticul care se va suprapune peste fibroreticul). In ochiurile celor doua retele se gasesc substanta fundamentala si celule conjunctive libere.
Tipuri de celule: 7. celule reticulare de tip fibroblastoid - aceste celule sunt de fapt un fibroblast cu mai multe
prelungiri care se unesc cu prelungirile altor celule prin jontiuni gap. Aceste celule formeaza citoreticulul. Celula are nucleu central eucromatic, citoplasma bazofila si este implicata in secretia matricei conjunctive si a rol structural (formeaza citoreticulul).
8. celule reticulare de tip histioid - au rol macrofag. Pot fi fixe sau libere si se sclerozeaza de regula in mansoane perivasculare. Ca aspect structural celulele pot avea ..................................... Rol de fagocitoza si pinocitoza. De asemenea ele colaboreaza cu limfocitele B si T in realizarea RI umoral si celular. Limfocitele T sunt localizate in zonele timodependente iar limfocitele B sunt localizate in zonele timoindependente.
9. celula reticulara interdigitale10. celule reticulare foliculare dendritice. Ultimele doua clase decelule sunt greu de diferentiat intre
ele (doar prin imunofluorescenta si ME). Aceste doua tipuri de celule colaboreaza cu macrofagele in elaborarea RI mediat umoral si celular.
Tesutul adiposGeneralitatiCelulele se numesc adipocite (lipocite). Tesutul contine pe langa celule si putina substanta
fundamentala si putine fibre, in special de reticulina. Celulele adipoase au tendinta de a se aglomera formand lobuli sau paniculi adiposi separati intre ei prin septuri conjunctive de tesut lax. In aceste septuri se gasesc vase de sange si terminatii nervoase adrenergice si colinergice. In cazul tesutului adipos brun exista doar fibre nervoase adrenergice.
Adipocitele isi au originea in celula mezenchimala nediferentiata care incepand cu S3 pana in S5 I.U. da nastere adipoblastului apoi preadipocitului si in final adipocitului. La nivelul adipoblastului si preadipocitului s-a detectat enzima a-naftil acetat esteraza, care are o actiune crescuta in aceasta celula.
Din preadipocit rezulta:
41

11. adipocitul alb12. adipocitul brun13. adipocitul hepatic (celula Ito) care se dispune intre hepatocite si peretele capilarului sinusoid. Ea
are rol de sinteza si depozitare a vitaminei A.Adipocitele brune nu pot da nastere adipocitelor albe sau celulei Ito.In perioada copilariei are din nou loc sinteza de adipocite sub actiunea STH, a hormonilor
tiroidieni si a insulinei. La pubertate procesul se reia sub actiunea hormonilor sexuali cand apar:14. adipocite ginoide (contin estrogeni)15. lipoide androide care contin androgeni
Marea majoritate a populatiei de adipocite se sintetizeaza la 2-3 ani iar la pubertate aceste celule se hipertrofiaza. Exista doua tipuri de obezitate:
16. hiperplastica (cresterea numarului de adipocite) apare in primii 2-3 ani17. hipertrofica (cresterea volumului adipocitelor) apare la pubertate sau mai tarziu.
Tesutul adipos ocupa:18. in copilarie
19. la barbati 15%20. la femei 25%
21. la pubertate22. la barbati 25%23. la femei 40%In curele de slabire adipocitul sufera o transformare seroasa, reversibila, devenind o celula de
tip fibroblastoid cu prelungiri.Daca se depaseste capacitatea de incarcare a adipocitelor se incarca fibroblastele si
histiocitele. Tesutul adipos albAdipocitul alb are diametru de 100-120 mm. Adipocitele se organizeaza in lobuli separati de
septuri formate din tesut conjunctiv lax ce aduce vase de sange in apropierea adipocitelor. Un capilar corespunde la 2-3 adipocite albe. Tot astfel ajung terminatiunile nervoase adrenergice si colinergice. Celula apare ca un inel cu pecete. Incluziunea lipidica se prezinta ca o picatura unica (tesut unilocular). Citoplasma si nucleul sunt impinse la periferie. Bula se dizolva in parafina si in hidrocarburi benzenice. Citoplasma este acidofila iar nucleul reprezinta pecetea inelului. Tot in tesut se observa numeroase capilare. La ME se observa ca incluziunea nu este invelita de endomembrana si ocupa cea mai mare parte a celulei impingand nucleul la periferie.
Nucleul este alungit, eucromatic. Organitele sunt mitocondrii cu criste tubulare si RE neted. Membrana celulara este dublata la
exterior de o lamina bazala care la randul ei este dublata de o retea fibroreticulara. Aceasta lamina bazala este comuna pentru mai multe adipocite. Intre adipocite exista substanta fundamentala, fibre de reticulina, capilare si filete nervoase adrenergice si colinergice care se opresc la nivelul laminei bazale unde-si varsa mediatorul, acesta ajungand in celula prin difuziune.
Rolul acestor celule este in metabolismul lipidelor, in lipogeneza si lipoliza. Lipogeneza se face din glicerol si acizi grasi (1 molecula de glicerol si 3 molecule de acizi grasi formeaza 1 molecula de triglicerid). Glicerolul se obtine prin aport exogen sau prin glicoliza iar acizii grasi pot fi sintetizat “de novo” sau provin din sursa endohepatica.
Acizii grasi proveniti exogen sunt absorbiti fie direct in circulatia sangvina fie in circulatia limfatica sub forma de chilomicroni. Chilomicronii au diametrul de 3 mm, au un miez central format de triglicerid si colesterol esterificat si o parte periferica formata din colesterol liber, proteine si fosfolipide.
La nivelul capilarelor chilomicronii se desfac sub actiunea lipoproteinlipazei secerata de celulele endoteliale. Acizii grasi rezultati raman legati de proteine si astfel vor traversa lamina
42

bazala a adipocitului alb si apoi membrana celulara si ajung in citoplasma unde formeaza din nou trigliceride. Depozitele adipoase sunt permanent reinnoite. Cel mai lipogenetic hormon este insulina care permeabilizeaza membrana adipocitului pentru glucoza. Lipoliza este controlata de glucagon.
Tesutul adipos brunIn acest tesut exista caroteni (pigmenti) prezenti in citoplasma. Vascularizatia este mai bogata
(1 capilar la 1 adipocit). Citocromii sunt numerosi (exista mitocondrii multe). Astfel se explica culoarea bruna. Acest tesut ocupa 5-6% din greutate la nastere. Este intalnit doar in viata fetala si in primele 5 luni de la nastere. El este localizat:
24. cervical25. retrosternal26. subscapular27. retrosimfizar28. in mediastinul profund29. in regiunea inghinala.
Acest tesut are rol de protectie termica.Organizare: adipocitele brune sunt asezate in paniculi separati de septuri conjunctive formate
din tesut conjunctiv lax cu foarte multe vase de sange (1 capilar la 1 adipocit). Terminatiunile nervoase sunt de tip adrenergic. Exista putina substanta fundamentala si putine fibre de reticulina.
La MO celula are forma poligonala cu nucleul dispus central. In adipocit exista mai multe bule lipidice care nu conflueaza datorita prezentei unor septuri citoplasmatice. Citoplasma este spumoasa cu numeroase gauri pentru ca si aici se dizolva in parafina incluziunile de lipide.
Adipocitul brun este mult mai mic - are un diametru de 30-40 mm.La ME se observa perinuclear numeroase picaturi lipidice nedelimitate de endomembrane ci
doar separate de septuri citoplasmatice. In restul citoplasmei exista mitocondrii cu criste si REN cu rol in sinteza lipidelor. Membrana adipocitului brun este dublata de o lamina bazala inconjurata de o retea fibroreticulara. Lamina bazala este proprie fiecarui adipocit.
Filetele nervoase adrenergice strabat lamina bazala si vin in contact direct cu membrana adipocitului. Cand scade temperatura mediului inconjurator se descarca norepinefrina care va stimula adenilat ciclaza ce duce la crestera cantitatii de cAMP care stimuleaza protein kinaza care va determina ardera acizilor grasi in ciclul Krebs ceea ce duce la formarea de energie. Tot odata este inhibata ATP sintetaza si astfel toata energia formata este disipata sub forma de caldura.
43

TESUTUL OSOSCursul cuprinde:
30. osul ca material31. osul ca tesut32. osul ca organ33. functiile osului34. structura macroscopica a osului35. clasificarea macroscopica a osului:
36. os fibros, imatur37. os lamelar, matur
38. componenetele osoase (2):39. matricea osoasa:
40. este formata din:41. lamele 42. canale vasculare43. periost44. endost
45. organizare biochimica:46. substanta organica (substanta fundamentala si fibre de colagen)47. substanta minerala
48. celulele, care sunt de doua tipuri:49. osteogeneratoare (a, b, g)50. osteoclaste
51. Vascularizatie si inervatie52. Histogeneza
53. osificare intramembranoasa (desmala)54. osificare endocondrala
55. centrii de osificare56. placa epifiara57. cresterea in diametru58. evazarea metafizei59. modelare osoasa60. remodelare osoasa
61. Unitatile multicelulare osoase62. Unitatile de remodelare osoasa (BMU)63. Repararea osoasa, care are trei etape:
64. etapa inflamatorie65. etapa reparatorie
44

66. etapa de remodelare67. Vindecarea prin calus extern68. Vindecarea trabeculara69. Vindecarea osoasa primara
70. Cartilajul71. Clasificare72. Structura (celule si matrice)73. Condron74. Cartilajul hialin
75. Histogeneza76. Cresterea cartilajului77. Condrocitele (la periferie)78. Matricea cartilaginoasa, care este formata din colagen si substanta fundamentala
(acid hialuronic si condronectina). Sinteza ei are loc in condrocit79. Rolul - in cresterea osului, existand cartilaj primar de osificare si cartilaj secundar
de osificare80. Cartilajul elastic
81. Localizare82. Aspect macroscopic83. Organizare84. Origine
85. Fibrocartilajul86. localizare87. Organizare88. Discurile intervertebrale89. Pericondrul
OSULOasele sunt organe vii, supuse unor modificari continue. Are rol in locomotie, in sustinere, in
protectie, in metabolism. De asemenea, el raspunde la stimuli interni si externi prin remodelare.
Osul ca materialOsul este un material complex, foarte rezistent la compresiune, similar betonului armat. El
are prin urmare urmatoarele caracteristici:90. rezistent dar usor91. rezistent la indoire ca stejarul92. rezistent la compresiune ca fierul, avand doar 2/3 din greutatea acestuia.
Acestea se datoresc urmatorilor factori:1. constructia lamelara a osului2. designul tubular3. intaririi prin reteaua trabeculara
Osul ca tesutCelulele vii sunt inglobate intr-o matrice bogat vascularizata. Din greutatea osului 92% este
substanta solida (uscata) iar 8% este apa. Din greutatea uscata, 65% este substanta minerala iar 35% este substanta organica, care reprezinta insa 95% din volum. Celulele reprezinta 3% din volum.
Osul ca organ
45

Osul contine si alte tesuturi: cartilaje, tesut fibros, tesut adipos, tesut vascular, maduva, tesut nervos. Oasele se modifica in functie de varsta: oasele tinere sunt flexibile, iar oasele batrane sunt rigide si au maduva fibroasa. Forma globala aosului este determinata de factorii genetici. Factorii epigenetici (nutritionali, greutatea, activitatea musculara) au rol in remodelarea osoasa.
Functiile osului1. Sustinere a trunchiului si extremitatilor, alaturi de muschi, ligamente2. Protectie a creierului, maduvei spinarii, viscerelor toracice3. Reprezinta mediul specific tesutului hematopoietic4. Asigura homeostazia ionilor minerali, continand
5. 99% din totalul de Ca2+ din organism6. 85% din totalul de fosfor7. 66% din totalul de magneziu8. 60% din totslul de sodiu
Structura macroscopica a oaselorOsul este fie compact (80% din masa osoasa), care se gaseste in oasele lungi si late, fie
spongios sau trabecular, care reprezinta 20% din masa osoasa. Epifiza este alcatuita din tesut osos compact si din tesut osos spongios.Diafiza este alcatuita din tesut osos compact.La nivelul craniului straturile se succed in felul urmator: dura mater - tabla interna - diploe -
tabla externa.Periostul lipseste la nivelul urmatoarelor structuri;
9. zona de insertie a ligamentelor, tendoanelor10. suprafata patelei11. colul femural, subcapsular12. astragalul13. oasele sesamoide.
Clasificarea microscopica a oaselor:14. os fibros, imatur, primar, in care fibrele de colagen sunt asezate la intamplare. Nu are o
organizare structurala. El poate persista la locul de atasare a ligamentelor si tendoanelor. In acest tip de os numarul de celule este de 4 ori mai mare decat numarul de lamele. Persistenta sa patologica se traduce prin boala Paget (osteozis imperfecta) sau prin sarcom osteogenic
15. os lamelar care se formeaza pe o formatiune preexistenta, osoasa, mineralizata in prealabil. Este format din osteoane cu diametrul de 250mm. Osteoanele reprezinta 2/3 din osul adult, intre ele existand sisteme de lamele interstitiale.
Componentele osoaseSunt in numar de doua: matricea osoasa si celulele.Matricea osoasaEste alcatuita din lamele care sunt in general in numar de 3-7, dar pot ajunge pana la 20 si
din lacune , in care se gasesc osteocite care au prelungiri ce patrund in canalicule ce se anastomozeaza.
Lamelele (4-20) sunt dispuse concentric in jurul unui canal vascular longitudinal cu diametrul de 50-90 mm. Acest canal vascular se mai numeste si canal haversian. Osteonul are un diametru de 200-300 mm. Osteoanele pot fi primare, secundare etc. Lamelele sunt si ele de mai multe feluri: lamele T (intunecate sau lamele A (alternanate), alternanad lamelele longitudinale (L) cu cele transversale (T). La periferia osteoanelor secundare exista linia de ciment, care contine mai putin colagen, mai putin fosfor si mai mult sulf.
Intre osteoane exista sistemul lamelar interstitial, angular. De asemenea, la periferie mai exista un sistem circumferential intern si extern, in corticala oaselor lungi. Lamelele din sistemul
46

trabecular nu prezinta canale vasculare (haversiene) si se hranesc prin difuziune din spatiul inconjurator.
Canalele vasculare sunt de doua feluri: longitudinale si transversale. Canalele longitudinale se mai numesc si haversiene si reprezinta capilare fenestrate sau chiar venule. Ele sunt marginite de celule turtite (mezenchimala, osteoblast in repaus, osteocit in activitate). Aceste celule sun asezate probabil in strat continuu, unite prin jonctiuni gap cu osteocitele din lacune. Aceste celule alcatuiesc bariera os - celule. Canalele transversale sau oblice se mai numesc si Volkmann si nu sunt inconjurate de lamele.
Periostul este alcatuit din doua straturi: extern, alcatuit din fibre (Sharpey) si bogat vascularizat si un strat intern, osteogenic, alcatuit din celule.
Endostul este alcatuit din celule aplatizate si acopera peretii tuturor cavitatilor osoase din osul spongios precum si canalele vasculare haversiene si Volkmann din osul compact. La osul spongios endostul reprezinta stratul periferic al stromei medulare.
Organizarea biochimica a matricei osoase este din substante orgenice si anorganice. Substanta organica intra in alcatuirea fibrelor si a substantei fundamentale. Ea da reactii PAS pozitive, metacromazie (pozitiva sau negativa), este acidofila si se poate evidentia prin incorporare de sulf.
Substanta fundamentala este amorfa. Ea contine proteine necolagenice specifice tesutului conjunctiv dar si proteine necolagenice specifice tesutului osos.:osteocalcina, osteopontina, sialoproteina osoasa. Osteocalcina si osteopontina se leaga de hidroxiapatita. Sialoproteina se leaga de celula. Sunt importante 1, 25-dihidroxi vitamina D, cu rol in sinteza proteinelor necolagenice si vitamina K (osteocalcina).
Fibrele (colagenul ) reprezinta 90% din totalul de substanta organica. Colagenul este de tip I, fibrele au diametru de 50-70 nm, si o periodicitate de 67 nm, fiind bogat in lizina hidroxilata. Prezinta numeroase legaturi transversale, avand o mare rezistenta mecanica si la agenti acii. Fibrele au un aranjament bine orientat. Ele alcatuiesc lamelele intunecate sau luminoase, existand diferente de orientare si/sau cantitative. Fibrele sunt dispuse helicoidal in raport cu substanta minerala. Pasul helixului se modifica de la o lamela la alta.
Substanta minerala reprezinta 65% din totalul de substanta uscata. Ea apara ca depozite submicroscopice de fosfat de calciu care se transforma in hidroxiapatita. La ora actuala exista o disputa cu privire la starea substantei anorganice in os. Nu se stie daca substanta este de la inceput sub forma de cristale sau fosfatul de calciu este amorf in timp ce hidroxiapatita este cristalina. Cristalele de hidroxiapatita sunt de forma unor bastonase cu lungimea de 40 nm si diametrul de 1,5-3 nm, distribuite pe o lungime de 60-70 nm in lungul fibrelor de colagen. Aranjarea colagenului este astfel facuta incat sa dea nastere unor goluri in interiorul unei retele ordonate. Golurile au dimensiuni de 40 nm / 2,5 nm si aici sunt dispuse cristalele. Substanta anorganica mai contine ioni citrat (la suprafata cristalului), ioni carbonat, substante fosfatice, fluor (inlocuieste OH). Izotopii radioactivi ai Ca si P pot inlocui ..........stabil. Mai este prezent plumbul, strontiul, radiul. In ostomalacie sau rahitism substanta anorganica poate scadea pana la o proportie de 35%. Hidroxiapatita are formula Ca10(PO4)6(OH)2.
CeluleleCelulele sunt de doua feluri: celule osteogeneratoare (celule osteoprogenitoare, osteoblaste,
osteocite) si osteoclastele.Celulele osteogeneratoare sunt de fapt aceeasi tip celula dar in faze functionale diferite.
16. Celulele osteoprogenitoare (a) provin din celula mezenchimala primitiva si dau nastere condroblastilor si osteoblastilor. Ele se gasesc la nivelul suprafetelor osoase libere (endost, periost, trabecule, cartilaj calcifiat, metafiza osului in crestere). Osteoblastele in repaus se aseamana morfologic cu aceste celule, fiind cel putin o parte din celulele ce marginesc osul adult.
47

17. Osteoblastele (b) depun osteoidul (matricea osoasa). Ele sunt celule columnare sau epiteloide care se gasesc la suprafata osului si cand sunt inactive au un aspect scuamos. Sunt celule sintetizatoare de proteine, produc factori de crestere care actioneaza autocrin sau paracrin, contin fosfataza acida. Se leaga de osteoblastele vecine prin jonctiuni gap, prezinta receptori pentru hormoni, vitamine, citokine. Spre exemplu, parathormonul actioneaza asupra osteoblastelor care produc factori de stimulare a osteoclastelor. Secreeta procolagenoza din care rezzulta colagenoza care lizeaza osteoidul potentand activitatea osteoclastelor. Au un mic rol in resorbtia osoasa, resorbind o portiune mica de osteoid, dupa care intervin osteocitele.
18. Osteocitele (g) se gasesc in lacune, prezinta prelungiri care patrund in niste canalicule si se anastomozeaza cu prelungirile osteocitelor vecine prin jonctiuni gap. Aceste prelungiri formeaza un fel de retea. Osteocitele prezinta o activitate dar redusa. Ele au organite “in regresie”. Se stie ca osteocitele provin din osteoblaste dar transformarea osteocitelor inapoi in osteoblaste este controversata.Osteoclastele au un diametru de 150 mm, prezinta cate 50 de nuclei in fiecare celula, si se
gasesc in niste lacune speciale, numite Howship. Celulele sunt polarizate: nucleii se afla la polul opus frontului osos iar inspre frontul osos
prezinta o margine in perie (margine dantelata) care in repaus dispare. Prezinta numerosi lizozomi primari, cu diametrul de 0,5-3 mm care contin fosfataza acida rezistenta la tartrat si divarese enzime proteolitice. In osteoclaste exista o zona numita compartiment subosteoclastic, in care mediul este puternic acid si care este echivalenta unui lizozom secundar. Mediul acid din acest compartiment este obtinut cu ajutorul unei pompe protonice ATP dependenta care pompeaza in zona subosteoclastica protonii obtinuti cu ajurorul anhidrazei carbonice. Acest compartiment este inconjurat de o zona de inchidere. Spre periferie mai exista si o zona “clara” deasupra careia se afla marginea dantelata (in perie) care este bogata in actina si mai este numita si zona filamentoasa.
Originea osteoclastelor se afla in precursori comuni cu monocitele si granulocitele, in maduva osoasa. Osteoclastele se obtin prin fuzionarea precursorilor, proces care este dependent de un factor necunoscut. Insa monocitele fuzionate nu au proprietati osteoclastice, desi provin din aceeasi precursori ca si osteoclastele.
Receptorii pentru calcitonina inhiba activitatea osteoclastelor dar ele sunt sensibile la factorul de stimulare a osteoclastelor, secretat de osteoblasti la actiunea parathormonului.
Vascularizatie si inervatieOsul primeste 10 din debitul cardiac. Osul este vascularizat cortical, prin arteriole ce provin
din artera nutritiva a osului respectiv, in timp ce vascularizatia venoasa se face centripet, venulele adunandu-se in venele medulare. Venele medulare au o capacitate de 6-8 ori mai mare decat capacitatea arteriala. Sistemul vascular periostal este de rezerva. Hemodinamica este necunoscuta. vascularizatia limfatica are loc la nivelul periostului.
Inervatia are loc prin prelungiri nemielinizate ale nervilor in canalele haversiene. Sinapsele sunt de tip adrenergic. Astfel sunt inervate: canalul haversian, periostul si vasele medulare. In maduva se gasesc corpusculi Pacini.
Modelarea osoasaFormarea osoasa are loc la nivel endosteal iar resorbtia osoasa are loc la nivel subperiosteal.
De asemenea are loc mentinerea formei conice a epifizelor. La nivelul boltii craniene osteogeneza are loc la nivelul partii externe iar osteoresorbtia are loc in regiunea interna. Aceasta distributir a proceselor asigura forma de bolta si mentinerea ei in concordanta cu cresterea organelor din cavitati.
48

Modelarea osoasa (organogeneza osoasa) consta in cresterea in lungime, cresterea in grosime (in diametru) si asigurarea formei conice a epifizelor. Acestea au loc prin procese complexe, coordonate, neintelese inca pe deplin, si care mentin toata viata forma oaselor. Prin remodelare osoasa 10% din intreaga masa osoasa a organismului se inlocuieste in fiecare an la individul tanar.
Remodelarea osoasa consta in inlocuirea osului primar, fibros, cu os secundar, lamelar organizat in sisteme haversiene (osteoane secundare). Mai intai are loc depunerea osteoidului si apoi mineralizarea acestuia. Un sistem haversian este depus in 4-5 saptamani, in primele zile are loc mineralizarea in proportie de 70% iar apoi, in cateva luni, are loc mineralizarea completa. Mineralizarea este influentata de factori locali sau generali si rezulta osul embrionar.
Placa epifizeala (cartilajul de crestere) reprezinta confluenta centrilor primar si secundar. La acest nivel se asigura cresterea in lungime a osului care este adaugat in sens diafizar si erodat in sens epifizar).
Histogeneza (osificarea)Osificarea este de doua tipuri: osificare de membrana (desmala) sau osificare de cartilaj
(encondrala). Mai intai are loc depunerea de matrice ideatica(in osificarea encondrala). Apoi are loc indepartarea cartilajului calcificat. Osul nou format este organizat intr-o retea trabeculara spongioasa primara. In zona corticala osul trebuie compactat prin umplerea golurilor si rezulta astfel osul primar, fibros, care nu este lamelar.
Osificarea intramembranoasa (desmala)Areloc la nivelul oaselor frontal, parietal si partial pentru oasele temporale, occipital,
mandibular, maxilare, corpii claviculelor.Osificarea se face din tesut conjunctiv primitiv, bine vascularizat ce contine numeroase celule
mezenchimale. Aceste celule se transforma in osteoblasti care depun osteoidul care apoi se va mineraliza. Osteoblastele se transforma apoi in osteocite si se obtine osul fibros care este neregulat si contine numeroase fibre de colagen. Osul fibros mai cuprinde si vase de sange, si nu e lamelar. El realizeaza spongioasa primara. Prin condensarea tesutului conjunctiv aflat la periferie se formeaza periostul si endostul. Mai departe are loc compasctarea (pentru osul compact) sau ................ umplerea spatiilor libere (osul trabecular).
Osificarea encondralaAre loc in centrii primari, secundari, la baza craniului, la nivelul coloanei vertebrale, in
oasele lungi si la extremitatile oaselor. Centrii de osificare reprezinta modificari ale tesutului cartilaginos prin aparitia unor muguri periostali vasculari.
Cartilajul sufera modificari zonale dinspre epifiza spre diafiza. La acest nivel celulele vor fi dispuse pe mai multe zone:
19. cartilaj de rezerva20. zona de proliferare21. zona de maturare (celulele cresc)22. zona de generare - unde are loc calcifierea matricei in sens longitudinal si nu la nivelul septurilor
transversale. 23. linia de eroziune (frontul de osificare) unde se gasesc vase (anse capilare) si celule mezenchimale
nediferentiate care vor da nastere diferitelor celule osoase precum si celule stem hematopoietice. Intr-o etapa ulterioara are loc distrugerea septurilor transversale, care nu sunt mineralizate.In procesul de osificare mai inatai se formeaza osteoblastii care depun osteoidul pe cartilajul
calcificat si se formeaza astfel spongioasa primara. Apoi actioneaza osteoclastele si se formeaza spongioasa secundara (lame obtinute prin resorbtie care formeaza osul).
Procesul actioneaza de la diafiza inspre epifiza. Apare gulerasul periostal epifizar.
49

Centrii secundari apar dupa nastere la nivelul epifizelor. Sub pericondru nu se depune osteoid.
Unitatile multicelulare ale osului - unitati de remodelare osoasa (BMU)Aceste unitati realizeaza remodelarea osoasa. De asemenea, dupa varsta de un an, ele asigura
transformarea osului primar in os secundar. Tot BMU realizeaza lamelele interstitiale.
Repararea osoasa - vindecarea fracturilor1. Etapa inflamatorie - Hemsitomul formarea coagului fibros, dureaza 3-4 zile2. Etapa reparatorie - se poate forma un calus moale, fibros sau un calus dur, fibrocartilaginos si se
formeaza osul primitiv. Etapa dureaza 3-6 luni.3. etapa de remodelare - osul lamelar inlocuieste osul primitiv. Remodelarea dureaza ani, uneori
pana la 7 ani.Vindecarea se poate face in mai multe feluri:
1. Vindecare prin calus extern - la oasele aparut eprin osificare encondrala, calusul este cartilaginos (dur) iar la oasele aparute prin osificare de membrana, calusul este fibros, care da nastere osului primar.
2. Vindecarea trabeculara - se formeaza os nou pe trabeculele deja existente, atunci cand leziunea este mica.
3. Vindecarea osoasa primara - prin BMU. “Oboseala osoasa” are ca substrat microfisuri la nivelul osului.
Cresterea in diametruConsta in adaugare de os (osificare) la nivel periosteal concomitent cu o resorbtie osoasa la
nivel endosteal. Prin acest proces se asigura si cresterea cavitatii medulare. Cresterea in grosime este influentata si de factorii biomecanici.
TESUTUL CARTILAGINOS
Este forma inalt specializata a tesutului conjunctiv. Este un tesut foarte bogat in matrice extracelulara. El formeaza scheletul embrionar axial si al membrelor. Tot tesutul cartilaginos asigura cresterea in lungime a oaselor. Nu contine vase sangvine, limfatice sau nervi, deci se hraneste prin difuziune.
Clasificari:· tesut cartilaginos embrionar· tesut cartilaginos permanent care poate fi de mai multe feluri:
· tesut calcifiat· tesut necalcifiat care poate fi scheletic sau nescheletic
In functie de raportul matrice/celule, tesutul cartilaginos se clasifica in:· tesut cartilaginos hialin· tesut cartilaginos eleastic · tesut cartilaginos fibros
Din punct de vedere structural tesutul are doua componente: celulele si matricea extracelulara. Celulele pot fi condrocite (se gasesc in lacune) sau condroblaste. Clasificarea este arbitrara, in functie de cantitatea de matrice din jurul lor. Din punct de vedere fenotipic condrocitul poate aparea ca o precondrocit (celula................. angajata), condrocit diferentiat sau condrocit hipertrofiat. Matricea celulara este si ea de doua feluri: teritoriala (in jurul lacunei) sau interteritoriala.
Condronul este unitatea functionala atesutului cartilaginos si este format dintr-o celula impreuna cu matricea teritoriala aferenta.
50

Cartilajul hialinSe gaseste in inelele traheale, in laringe, capetele ventrale ale coastelor, pe suprafetele
articulare. Este translucid (semitransparent) si de culoare albastrui-gri.HistogenezaCelulele mezenchimale isi pierd prelungirile si se apozitioneaza. Centrele de condrificare
secreta matricea, care este o substanta amorfa si fibrele care sunt mascate de matrice. Celulele se vor izola in lacune. La periferie, mezenchimul se condenseaza si se formeaza pericondrul.
Cresterea cartilajuluiEste apozitionala (pericondrul) sau interstitiala, formandu-se grupurile izogene. De asemenea
mai este si cresterea cartilajului epifizal, la oasele lungi.Aspectul la microscopul optic - vezi L.P.CondrociteleAu forma elipsoidala, avand membrana celulara paralela cu marginea lacunei, sau, daca se
gaseste mai in profunzimea lacunei, are forma hemisferica sau rotunjita. Celula (celulele) ocupa intreaga lacuna. Prezinta incluzzii de glicogen si lipide, are aparat Golgi si reticul endoplasmic rugos, care sunt mai dezvoltate in perioadele de sinteza. Ele sintetizeaza toate componentele matricei simultan: colagen, proteoglicani, acid hialuronic.
La periferie prezinta un invelis matricial pericelular format din proteoglicani, cu aspect de pasla si cu grosimea de 1-3 mm. Urmatorul strat este inelul lacunar, format din fibrile de colagen, ca un cosulet. Apoi urmeaza matricea teritoriala formata din condroitin sulfat, avand o grosime de 50 mm si cu o structura intens bazofila. Condrocitele embrionare pot avea un mic................. Pe suprafata au caveole, microvili. Condrocitul hipertrofic are o sinteteza scazuta de colagen II, proteoglicani si inhibitori de proteaze.
Matricea cartilaginoasaAre o structura omogena la microscopul optic. Este bazofila. Se coloreaza metacromatic cu
coloranti cationici, Azur A, albastru de toluidina, care formeaza complexe impreuna cu glicozaminoglicanii (GAG). Nu se evidentiaza fibrilele colagenice. Ele se pot evidentia dupa digestia GAG cu .................. Matricea teritoriala este bazofila datorita condroitin sulfatului.
Componentele matricei sunt colagenul si substanta fundamentala care este amorfa.Colagenul reprezinta 40% din greutatea uscata a cartilajului. El este de tip II, iar diametrul
fibrilelor este de 15-45 nm. Ele nu se asambleaza in fascicule groase, si formeaza o retea in matrice. Colagenul nu exista in matricea pericelulara. Arhitectura fibrilelor este dictata de conditiile mecanice si in general sunt tangentiale la pericondru. In cartilajul articular, la suprafata, fibrilele sunt paralele cu ......................... si tangentiale la pericondru. Mai exista si alte tipuri de colagen: IX, X, XI. Colagenul de tip IX sau XI se gaseste asociat cu fibre de colagen de tip II iar colagenul de tip X se gaseste numai in zona de cartilaj hipertrofic. De asemenea mai exista si colagen de tip I si fibronectina care sunt localizate numai in zona hipertrofa, in portiunea inferioara.
Substanta fundamentala este un gel cu proprietati speciale. Ea contine proteoglicani (PG) formati dintr-un miez proteic numit “agrecan” si aproximativ 50 GAG (condroitin sulfat, keratan sulfat) atasati agrecanului. Acidul hialuronic leaga apoi aproximativ 100 PG prin proteine de legatura, intre PG fiind o distana de 30 nm. PG se gasesc la distante regulate de-a lungul fibrei de colagen. Cu ajutorul microscopului electronic se poate vizualiza reteaua de PG care apare in elul urmator: fibrile fine, discrete, corespunzatoare lanturilor de GAG si fibrile groase, evidente, corespunzand miezurilor proteice. Agregatele de PG realizeaza un gel puternic hidratat. Apa reprezinta 70-80% din greutatea cartilajului. Ea se depune structurat, in staturi, fiind atasata gruparilor COO- si SO4
2-. Aceasta structura explica rezistenta la compresie.
51

Condronectina este o glicoproteina fibronectin-like. Ea este incorporata pe suprafata condrocitelor si se leaga de colagenul de tip II si de GAG. Aceasta proteina modifica atasarea celulelor la matricea cartilaginoasa.
Cartilajul elasticLocalizare: in urechea externa, peretii cailor auditive, trompa lui Eustache, in epiglota,
cartilajele corniculate si cuneiforme.Aspectul sau macroscopic este mai opac si galbui, flexibil.Este alcatuit din condrocite similare celor de la tesutul hialin, asezate in lacune izolate sau
grupate cate 2-4 in grupe izogene. Matricea este mai putin bogata si prezinta numeroase fibre elastice ramificate.
Originea lor este in ariile primare ale tesutului conjunctiv care contin celule conjunctive ramificate si fibre care nu sunt nici colagen nici elastina. Se dezvolta ca in ariile tesutului hialin dar dobandesc caracteristicile tesutului cartilaginos elastic. Tesitul conjunctiv de la periferie da nastere pericondrului.
Rolul importantei matricei este demonstrat de experimentul cu papaina.
FibrocartilajulSe aseamna foarte mult cu tesutul conjunctiv dens regulat. Cele doua tesuturi sunt adesea
unul in contnuarea celuilalt, alcatuind ligamentele si tendoanele ce se insera pe os.Localizare: discuri intervertebrale, simfiza pubiana, unele cartilaje articulare.Sinteza in condrocit a componentelor matriciale:In RER dureaza 10 minute, in aparatul Golgi dureaza 30 minute, iar in granulele secretorii
stau 2-3 ore. Condroitin sulfatul si GAG sunt adaugati in aparatul Golgi iar in granulele secretorii exista un material filamentos (colagen) si un material granular (PG). PG sunt secretati ca monomeri si asamblati pe acid hialuronic extracelular.
Aceeasi celula cartilaginoasa secreta concomitent toate componentele matricei. Organizarea microscopica se face pe baza unor condrocite dispuse in siruri intre manunchiuri
groase de fibre de colagen de tip I. Nu are pericondru si este acidofil datorita colagenului. Condrocitele sunt situate in lacune inconjurate de matrice extracelulara bazofila datorita condroitin sulfatului si keratan sulfatului.
Discul intervertebralVertebrele sunt acoperite de cartilaj hialin iar discul se afla in centru si e format din
fibrocartilaj. Nucleul pulpos este derivat din notocord si este format din acid hialuronic si putine celule. Fibrocartilajul este alcatuit din lamele formate din manunchiuri de fibre de colagen de tip I si care se termina in cartilajul hialin al vertebrelor. Lamelele adiacente au directii oblice dar opuse, de aici aparand aspectul penat.
Pericondrul este prezent in toate cartilajele mai putin in cartilajul articular. Este un tesut conjunctiv dens, mai bogat in celule in partea indreptata spre cartilaj. Celulele se diferentiaza in condroblaste . Contine colagen de tip I.
Histofiziologia cartilajului· cartilajul are rezistenta crescuta la compresiune· cartilajul ajuta la diminuarea la minim a frecarii in articulatii· cartilajul este responsabil de crestera in lungime a oaselor· cartilajul contine numeroase proteine, vitamina C, vitamina D, calciu, fosfat. Absenta acestor
principii din alimentatie duce la rahitism si osteomalacie. Acumularea de matrice fara sa se osifice duce la aparitia unor oase mai usor deformabile sub actiunea diferitelor forte mecanice.
· Hipofiza produce STH care este responsabil de aparitia nanismului, gigantismului si acromegaliei.
52

· Imbatranirea ....................· daca celulele cartilaginoase mor atunci cartilajul este invadat de vase sangvine si fagocite si are
loc fibrozarea.· extractia cartilajului impiedica angiogeneza· transplantul de cartilaj NU este urmat de rejectie deoarece este nevascularizat si prin urmare nu
permite patrunderea limfocitelor si a imunoglobulinelor.·
Rolul cartilajului in cresterea in lungime a osuluiCentrul primar de osificareIn centrul tijei modelului cartilaginos, un grup de condrocite se hipertrofiaza , avand
modificari ccelulare precis programate:· sinteza de colagen de tip X· sinteza de fibronectina· sinteza de anhidraza carbonica· sinteza de fosfataza alcalina· scaderea sintezei de inhibitori de proteaza· scaderea sintezei de colagen tip II· Scaderea sintezei de PG specifici cartilajului.
Ulterior are loc largirea lacunelor si apar modificari ale PG care favorizeaza depunera calciului. Apoi patrund vase sangvine si celule formatoare de osteoid in cartilajul mineralizat (calcificat). Apoi are loc o proliferare a condrocitelor cu dispunerea in coloane paralele.
Centrul secundar de osificareApare la nivelul epifizelor ca urmare a acelorasi modificari in structura tesutului cartilaginos.
Centrii avanseaza unul spre celalalt pana nu mai ramane decat placa epifizeala din tesutul cartilaginos (cartilajul de crestere) care va sigura cresterea in lungime a osului.
53

SANGELE
Are origine mezenchimala. Este alcatuit din organite (elemente figurate) si plasma. Elementele anucleate sunt hematia si trombocitul. Elementele nucleate sunt leucocitele care
pot fi granulare (PMN) - neutrofile, eozinofile sau bazofile - sau pot fi agranulare - limfocitele si monocitele.
Plasma sangvina contine 90% apa si 10% substante uscate. Substantele organice sunt albumine, globuline, fibrinogen, hormoni si vitamine in cantitati mici. Substante anorganice sunt sarurile minerale si ioni aflati in diverse concentratii. De asemenea mai se gasesc metaboliti: urea, acidul uric, creatinina, glucoza.
La fiecare element figurat se vor studia:· numarul· diametrul· forma· organizarea structurala, observata la MO· organizarea ultrastructurala, observata la ME· durata de viata in sangele circulant· originea· functiile indeplinite
Hematia
NumarulSe gaseste in numar de 4 mil. / mmc la femeie si 5 mil. / mmc la barbat. Aceste cifre insa nu
sunt fixe. Exista cresteri ale acestor valori (poliglobulii) care pot fi fiziologice sau patologice. Poliglobulie fiziologica se poate intalni:
· la altitudini ridicate· la sportivii de performanta· la femeia gravida in ultima perioada de sarcina· la nou nascut (7-8 mil.) dar care scade insa repede.
Poliglobulii patologice se intalnesc in:· boli ale sangelui· boli generale
Scaderea numarului de hematii se numeste anemie si apare in boli ale sangelui sau in boli generalizate.
Dimensiuni· 8,6 mm la hematia circulanta· 7,5 mm la hematia uscata
Dimensiunile mari ale hematiilor: macrocite (10 mm) sau megalocite (12 mm) insotesc anemiile sau alte boli ale sangelui. Diametre scazute (microcite) se intalnesc in anemii. Poate exista si anizocitoza (diametre diferite).
Forma· discocit (forma clasica, fiziologica) - in plan frontal are forma rotunjita iar in plan sagital are
forma de disc biconcav (piscot). · sferocit - hemati rotunjite, cu forma de balon
54

· echinocit - au forma de sfera dar membrana celulara emite prelungiri de lungime, forma si grosime egala si situate la distante egale unele de altele. Aceasta forma apare atunci cand concentratia de ATP din celula sau din exterior scade.
· stomatocite - sunt celule rotunjite care prezinta o infundatura. Aceasta forma apare cand in mediul extern apare o modificare a pH-ului.
· ovalocite - hematii de forma ovalara· acantocite - celulele au forma sferica dar membrana emite prelungiri de lungimi si grosimi
diferite asezate la distante inegale. Aceasta forma poate sa devina si ireversibila in unele boli, spre exemplu in ciroza hepatica.De asemenea exista si hematii cu forma ireversibila, patologice:
· drepanocitele - hematii in forma de secera· leptocite - hematii turtite
Poikilocitoza apare atunci cand hematiile au forme diferite. In mod fiziologic, atunci cand capilarele sangvine au un diametru mai mic de 7 mm, pentru ca sa poata trece, hematiile se alungesc foarte mult, deformandu-se iar apoi revin la forma initiala.
Structura si ultrastructura hematieiHematia circulanta este anucleata (la om).Citoplasma: dispar toate organitele celulare si raman doar microfilamentele si microtubulii
care formeaza citoscheletul si mentin forma celulei. Este posibil sa ramana membrana nucleara care va forma inelele Cabat. Tot din nucleu raman resturi de cromatina sub forma de granule bazofile - granulele Jolly.
Proteinele sunt proteine structurale si protein enzime. Proteinele structurale pot fi fibroase, alungite si alcatuiesc o pseudoretea cu ochiuri diferite in care se gasesc proteine structurale de forma globulara. Protein enzimele sunt:
· lactat dehidrogenaza (enzima a glicolizei anaerobe)· enzime din suntul pentozic - glucozo-6-fosfat dehidrogenaza, 6-fosfogluconat dehidrogenaza· ATPaza· urme de catalaza· anhidraza carbonica - intervine in legarea CO2
· hemoglobina - ocupa 33% din greutatea hematiei care are 40% substanta uscata in total. Este formata dintr-o parte proteica - globina si partea de hem care este alcatuita din protoporfirina IX si Fe2+.
In formarea Hb (hemoglobinei) exista mai multe etape:1. Se formeaza globina in RER. 2. Se formeaza protoporfirina IX3. Se leaga Fe la protoporfirina si se formeaza hemul4. Se unesc globina si hemul
Hemoglobina este de mai multe feluri:· HbA1 - 97% din totalul de Hb. Globina este formata din 2 lanturi a si 2 lanturi b.· HbA2 - 2% din totalul de Hb. Globina este formata din 2 lanturi a si 2 lanturi d.· HbF (fetala) - 1% din totalul de Hb. Globina este formata din 2 lanturi a si 2 lanturi g. La nastere
ea reprezinta 80% din total.· HbAlc - globina este format adin 2 lanturi a si 2 lanturi b dar la valina terminala se adauga o
molecula de glucoza.Exista boli ereditare determinate de defecte genetice in formarea Hb - secvente gresite de
aminoacizi in lanturile b ale globinei.
Hemoglobinopatii sunt determinate de defecte genetice in lanturile b cel mai frecvent, mai rar in lanturile a si foarte rar in amandoua simultan.
55

Hemoglobinopatii determinate de greseli in lanturile b:· HbS - inlocuirea acidului glutamic din pozitia 7 cu valina· HbC - inlocuirea acidului glutamic din pozitia 7 cu lizina· HbG - inlocuirea acidului glutamic din pozitia 7 cu glicina· HbE - inlocuirea acidului glutamic din pozitia 26 cu lizina
Hemoglobinopatii determinate de defecte in lanturile a· HbI - inlocuirea lizinei din pozitia 17 cu acid glutamic
Hemoglobinopatiile sunt raspandite diferit: cele cu HbS, HbC sunt mai frecvente in tarile mediteraneene iar cea cu HbG este mai raspandita in Tailanda si Asia de Est.
Talasemia este un defect genetic in care celulele (hematiile tinere) au o cantitate redusa de RER, deci se sintetizeaza mai putina globina dar in acelasi timp formarea de hem este normala si astfel Hb va fi mult alterata si redusa in hematii. Talasemia majora este o forma foarte grava cu rata de supravietuire redusa. Talasemia minora permite supravietuirea pana la varsta adulta sau chiar mai mult.
Membrana celulara este de natura lipoproteica, sub forma de mozaic. La exterior prezinta glicocalix. A fost prima membrana izolata si cea mai mult studiata pentru ca hematia in mediu hipoton se umfla si se dezintegreaza si rezulta o hematie fantoma, ramanand doar membrana. In mediu hiperton se ratatineaza si emite tepi.
Structura ei este: 30 % lipide, 50-52% proteine, 18% glucide. Pe fata exterioara a membranei se gasesc acetilcolinesteraza si nicotin-amid-dinucleotid. Pe fata interna a membranei se gasesc ATPaza, 3-gliceraldehid dehidrogenaza si proteinkinaza. De asemenea pe fata externa prezinta numerosi antigeni de suprafata, de ordinul trilioanelor, grupati in: antigen A, B.A (antigen A1 / A2) B (antigen B) AB (antigene A, B) 0 (nici un antigen)aglutinina antiB aglutinina antiA nici o aglutinina aglutinina antiA, antiB
In functie de prezenta sau absenta factorului Rh, indivizii pot fi Rh pozitivi sau Rh negativi. La nou nascut poate aparea icterul hemolitic al nou nascutului. Aceasta se intampla la al doilea copil provenit de la un tata Rh pozitiv si o mama Rh negativ. In timpul sarcinii el primeste de la mama anticorpi anti Rh dezvoltati de mama in timpul primei sarcini, ceea ce determina liza hematiilor.
Durata de viata120 zile. Hematiile imbatranite ajung in ficat sau splina unde sunt fagocitate de macrofage si
sunt apoi atacate de lizozomi care contin hidrolaze acide. Hematia se descompune si se elibereaza Fe care reintra in circulatie si ajunge in maduva rosie hematogena ajutand la formarea de noi hematii. Astfel se realizeaza circuitul Fe in organism.
OrigineaEste in maduva rosie hematogena, hematopoietica sau hematoformatoare (MRH) care da
nastere elementelor figurate ale sangelui. Aceasta se gaseste in oasele late (stern, coxal) si in corpurile vertebrelor.
FunctiileEste transportoare de gaze, factorul principal fiin Hb care leaga si dezleaga rapid O2 sub
actiunea methemoglobin reductazei si CO2 sub actiunea carboanhidrazei. Se poate forma si o legatura ireversibila in carboxihemoglobina determinata de legarea monoxidului de carbon - CO.
Trombocitul
Numarul
56

200.000 - 400.000 / mmc. Cresterea numarului se numeste trombocitoza iar scaderea numarului se numeste trombocitopenie si insotesc boli ale sangelui sau boli generale.
DiametrulEste cel mai mic element figurat al sangelui avand un diametru de 3-4 mm.
FormaEste ovalara, in stare proaspata sau cu margine crenelata in stare uscata.
Structura si ultrastructura trombocituluiEste anucleat.Citoplasma prezinta doua zone: centrala, intunecata - granulomer, si una periferica, clara -
hialomer.GranulomerulEste format din vezicule de diferite marimi si opacitati, inconjurate fiecare de o
endomembrana. Unele vezicule sunt organite celulare, foarte putine la numar insa; 2-3 mitocondrii cu rare criste, cativa ribozomi liberi, cateva vezicule golgiene, putin RE, numar mic de lizozomi (granulele l).
Veziculele dense au diametrul de 250-300 nm si contin serotonina, epinefrina, norepinefrina, ATP, ADP, ioni Ca2+. Veziculele a sunt implicate direct in coagulare si contin: tromboxan A2, trombospondina, prostaglandine G2, H2, factorul VIII (Willebrand).
HialomerulPrezinta 3 sisteme canaliculare, de la exterior spre interior:
1. Sistem canalicular deschis - tubi care comunica cu invaginarile membranei plachetare, favorizand schimbul dintre mediul intern si mediul extern.
2. Sistem tubular dens alcatuit din tubi anastomozati asezati ca un inel in hialuromer. Pot fi tubi ai RE.
3. Sistem marginal reprezentat de un inel de microtubuli care mentin forma celulei.Exista proteine speciale cu rol in coagulare:
· trombostenina· factorul III plachetar· factorul IV plachetar
Toate aceste structuri stau asezate pe citoscheletul format din microfilamente de actina si miozina si microtubuli. Citoscheletul are rol de suport, mentine forma celulei, asigura deplasarea sau contractia in formarea cheagului sangvin.
Membrana plachetara este lipoproteica, mozaicata, cu glicocalix la exterior. Prezinta frecvente invaginari care comunica cu sistemul canalicular deschis prin intermediul carora se fac schimburi bidimensionale. De asemenea ea poate emite prelungiri sub forma unor valuri ondulate cu ajutorul carora se deplaseaza. La exterior se gasesc receptori pentru factorii plasmatici cu rol in coagularea sangelui.
Durata de viataVariaza de la cateva ore pana la 3-4 zile. Ele parasesc vasul si patrund in tesuturile
conjunctive unde sunt fagocitate de macrofage.
OrigineaSe formeaza in MRH pe linie megacariocitara, provenind din megacariocit care are 60 nuclei.
Trombocitele sunt astfel fragmente din citoplasma megacariocitelor.
57

FunctiiPlachetele intervin direct in coagularea sangelui. Aderarea trombocitelor are loc in momentul
in care se lizeaza peretele vasului. Ele intra in actiune in trei etape:1. Aderarea la peretele lezat2. Agregarea la locul leziunii3. Metamorfoza vascoasa
AderareaPlachetele se lipesc de endoteliul vaselor langa locul lezat si lipirea este favorizata de
colagenul din peretele vasului cu care stabilesc legaturi.AgregareaReprezinta formarea de grupuri de plachete la locul de lezare. Aceasta este favorizata de
colagen si alti componenti din plachete: serotonina, ADP, ATP, tromboxanul A2, Ca2+.Metamorfoza vascoasaTrombocitele isi modifica forma si structura, acumuleaza apa, endomembranele se rup si
continutul veziculelor se raspandeste in toata citoplasma dandu-i un aspect vascos, astructurat. Apoi membrana plachetara se rupe si intreg continutul citoplasmic se raspandeste incepand procesul de coagulare.
Procesul de coagulareExista mai multe tipuri de factori:
1. tisulari - tromboplastina - factorul tisular principal2. plasmatici - protrombina si fibrinogenul3. plachetari - serotonina, ADP/ATP4. prezenti atat in trombocit cat si in plasma - Ca2+, trombostenina
Procesul de coagulare are mai multe etape:1. Intr-o prima etapa tromboplastina va actiona supra protrombinei si o va transforma in trombina.2. Trombina actioneaza asupra fibrinogenului care se transforma in fibrina care este o masa de fibre
cu directii diferite ce formeaza o pseudoretea.3. Apoi factorii plachetari cu rol contractil (serotonina, Ca2+) contracta masa de fibrina si se
formeaza cheagul sangvin4. Complexul plasminogenul - plasmina asigura hemoliza (trombocitoliza) cheagului sangvin.
Acest complex este format din 4 tipuri de molecule:1. plasminogen2. plasmina3. inhibitori ai complexului4. stimulatori ai complexului
Plasminogenul se transforma enzimatic in plasmina si aceasta este influentata de inhibitorii si stimulatorii complexului.
Hemofilia este un defect enzimatic, determinat genetic si care impiedica transformarea fibrinogenului in fibrina. Astfel, orice rana poate fi mortala.
Plachetele mai au si alte functii decat cele din coagularea sangvina:· Intervin in procesele inflamatorii si elibereaza substante care stimuleaza chemostatismul
neutrofilelor la locul de invazie microbiana, neutrofilele fiind primele care actioneaza.· Pot endocita bacterii si virusuri dar nu le pot distruge deoarece au un continut redus de hidrolaze
acide.· Pot interveni in procesul imun prin emiterea de substante care stimuleaza producerea de
imunoglobuline (in special IgG)· Pot interveni in vindecarea ranilor eliberand substante care stimuleaza miogeneza si angiogeneza
(formarea de noi capilare)
58

Leucocitele
Sangele are matricea extracelulara fluida. El are doua componente: plasma si componenta celulara. Hematiile si plachetele sunt elemente care isi desfasoara functiile fiziologice in spatiul vascular. Leucocitele se afla doar in trecere prin spatiul vascular si au rol doar in tesuturi.
59

Tehnici de studiere a morfologiei si fiziologiei celulelor sangvine:· Tehnica examinarii pe frotiu a sangelui periferic - se vad forma, dimensiunile si aspectele
caracteristice ale celulelor, numarul acestora si se pot stabili raporturile intre unele elemente figurate (formula leucocitara).
Importanta utilizarii frotiului:Cu ajutorul lui se poate stabili un diagnostic ipotetic. Diagnosticul cu ajutorul
sangelui periferic se face prin hemoleucograma. Frotiul este relativ usor de realizat, sangele periferic fiind usor de prelevat iar un esantion mic da informatii multe.
Se pot obtine date orientative pentru diagnostic, chiar cand nu exista nici un semn de boala asa cum este cazul in leucemii sau reactii inflamatorii. Astfel se permite orientarea planului de investigatii: anemia cu hematii mici poate fi determinata insuficienta Fe - anemie feripriva. Dupa tratament se face din nou investigatia prin frotiu.
Frotiul este necesar pentru pacientii ce suporta cure agresive de radioterapie si chimioterapie in cazul neoplaziilor (exista aprox. 200 de tipuri de cancer). Exista elemente comune tuturor cancerelor: proliferarea anarhica a unor celule si ............................................................................ metastaze.
In cazul diagnosticului precoce are loc interventia chirurgicala si tumora este eliminata. In cazul diagnosticului tardiv, cand a avut loc si o metastazare se incearca iradierea pacientului sau chimioterapie. Prin chimioterapie se incearca de fapt intarzierea proliferarii celulelor tumorale, insa aceasta terapie nu este selectiva, actionand si pe celulele indemne. Tesutul hematopoietic din MRH, avand un turn-over crescut, poate fi afectat, ajungandu-se uneori pana la aplazie medulara. De aceea este obligatorie transfuzia de sange periodica la bolnavii aflati sub chimioterapie.
· Citometria in flux - foloseste instrumente scumpe, necesita proteine marcate, sau anticorpi marcati. Aceasta tehnica este folosita frecvent in centre de hematologie si transplant de organe. Ajuta la stabilirea compatibilitatii intre receptor si donator. Se ofera informatii despre dimensiuni, granularitatea si fosforescenta celulelor.
Tehnica este urmatoarea:Celulele trec prin fata unui fascicul si se inregistreaza fluorescenta lor. Initial celulele
sunt puse in contact cu anticorpi marcati pentru anumite proteine. Avand in vedere faptul ca celulele cu acelasi rol functional prezinta pe suprafata acelasi set de proteine, anticorpii marcati se vor atasa numai la anumite tipuri de celule.
Aceasta tehnica poate fi folosita in vindecarea unor leucemii folosindu-se autotransplantul de maduva. Se preleveaza maduva de la pacient (care de obicei prolifereaza anarhic pe o singura linie hematopoietica, mai rar multiclonal) si apoi, cu ajutorul proteinelor marcate si a citometriei in flux se separa celulele stem sanatoase de cele anormale, acestea prezentand antigene de suprafata diferite. Apoi maduva purificata este introdusa inapoi in organismul pacientului. De altfel, cancerele constau de obicei in proliferare monoclonala. In cazul leucemiei linia celulara proliferata creste si sufoca celelalte linii impiedicand dezvoltarea lor. Prin citometria in flux se reactiveaza maduva hematopoietica. In viata oricarui individ se formeaza cateva zeci de celule canceroase, care sunt insa distruse de sistemul imun si pacientii nu se imbolnavesc de cancer.
· Exista si unele aparate care au sortatoare de celule si se obtin grupuri de celule identice.
Neutrofilele
Au originea in MRH.Dupa raportul cu peretele vascular exista doua populatii de neutrofile: circulante si marginale
(au tendinta de aderare).Numarul 3.000 - 6.000 / mmc.
60

Durata de viata este de 6 h in sange dupa care trec in tesuturi unde au o durata medie de viata de 4 zile.
Au rol in apararea antiinfectioasa, apartinand sistemului fagocitar.In sange ele au stadiul de evolutie final, diferentiat. Ele nu se mai divid si se pot autoelimina,
activandu-si programul genetic de autodistrugere. La microscopul optic se calculeaza dimensiunea lor 10 -12 mm, raportul nucleu / citoplasma = 1 : 1. Celulele au un singur nucleu care este segmentat. Celulele sunt PMN (polimorfonucleare) avand nucleul lobat. La celulele tinere nucleul este nesegmentat, in banda. Nucleul se va segmenta in 2, 3, 4, sau 5 lobi.
Citoplasma prezinta granulatii mici, colorate roz sau rosu-portocaliu si unele granulatii colorate in albastru-violet. Ele sunt raspandite uniform in citoplasma. Se poate vedea un apendice, ca un bat de toba - corpusculul Barr.
Distributia granulelor la microscopul electronic arata ca in vecinatatea membranei exista mai putine granule si mai multe structuri ale citoscheletului. In momentul aderarii la endoteliul vascular miscarea neutrofilului este de tip amoeboidal care se face ca urmare a rearanjarii citoscheletului.
Continutul granulelor a fost studiat prin histochimie. Granulele pot fi de doua feluri primare (comune) si secundare (specifice), clasificare care s-a facut dupa afinitatile tinctoriale. Granulele primare sunt azurofile si dau reactii pozitive pentru coloranti azur. Ele contin un set de enzime lizozomale. Granulele specifice nu au aceleasi enzime specifice si au proteine bazice numite fagocitone. Aceste granule asigura capacitatea neutrofilului de a fagocita bacterii.
D.p.d.v. functional neutrofilele se clasifica in:1. celule inactive2. celule activate.
Celulele activate prezinta pe suprafata lor anumite molecule de adeziune cu ajutorul carora se ataseaza la celulele ce captusesc vasele, migreaza apoi in tesuturi unde ingera particulele straine pe care apoi le digera.
Etapele distrugerii microorganismelor:1. Identificarea microorganismelor prin factori chemotactici care sunt niste compusi chimici pe care
celula ii detecteaza ca fiind prezenti in mediu iar apoi are loc raspunsul celular, neutrofilul migrand spre zona din organism unde concentratia factorilor detectati este maxima.
2. Apoi celulele se ataseaza de vas, strabat peretele endotelial la locul de jonctiune intre doua celule endoteliale, distrugand membrana celulara cu enzima numita colagenaza. Neutrofilele trec apoi in tesutul conjunctiv prin miscari amoeboidale si ajung la locul unde concentratia de microorganisme este maxima.
3. Distrugerea, liza microorganismelor.4. Prelucrarea materialului rezultat din liza.
Atasarea neutrofilelor atat la celulele endoteliale cat si la tesutul conjunctiv se face prin molecule de adeziune care se gasesc fie pe leucocite fie pe celulele epiteliale sau pe celulele tesutului conjunctiv. Exemple de molecule de adeziune:· ICAM 1 (molecula de adeziune intercelulara - intercelular adhesion molecule)· ELAM 1 (endothelial-leukocyte adhesion molecule)
Exemple de factori chemotactici:· peptide scurte, cu radicali N-formil· diferite tipuri de componente ale sistemului complement· mesageri ai mastocitelor?: metaboliti ai acidului arahidonic.
Ingestia este precedata de procesul de opsonizare (bacteria este acoperita din loc in loc de proteine de tip Ig sau elemente componente ale sistemului complement). Opsonizarea reprezinta acoperirea unei molecule (particule) bacteriene cu proteine plasmatice recunoscute de receptorii neutrofilului ca semnal pentru ingestie. Rolul opsonizarii este de a reduce mobilitatea microorganismului si de a recunoaste particula antigenica.
61

In momentul recunoasterii, citoscheletul neutrofilului se distruge, si veziculele de fagocitoza sunt internalizate. Ulterior veziculele de fagocitoza fuzioneaza cu granulele neutrofilului si are loc o degranulare (eliberarea continutului granular spre vezicula de fagocitoza). In acest moment se activeaza productia de radicali liberi care ulterior desfac elementele componente ale microorganismelor ingerate. Pentru neutrofile, degranularea are loc in citoplasma celulei, spre deosebire de bazofile si eozinofile unde degranulare are loc extracelular, uneori cu efecte toxice asupra organismului propriu.
Eozinofile
Se gasesc atat in sange cat si in unele tesuturi. Sunt bine reprezentate in tesuturile ce au contact direct cu mediul extern: aparatul respirator, digestiv, genito-urinar.
Numarul este de 450 / ml (mmc). Raportul de eozinofile in sange / tesuturi este 1 / 300. Numarul lor creste in infectiile parazitare.
Durata de viata este mai mare dacat la neutrofile, fiind de cateva saptamani.La MO se stabilesc dimensiunile de 10-12 mm, raportul nucleu / citoplasma este 1 : 1.
Nucleul poate fi nesegmentat sau, mai frecvent, are doi lobi iar nucleul are un aspect caracteristic, in forma de desaga. Citoplasma se coloreaza in rosu portocaliu cu eozina iar granulele se coloreaza eozinofil.
La ME, granulele au in centru un miez cristaloid caracteristic, diferit de la o specie la alta. D.p.d.v. histochimicgranulele contin:
· un set de hidrolaze lizozomale· un set de proteine cationice
Rolul functional:· Fagocitoza si liza bacteriana, putin importanta in vivo. S-a experimentat acest lucru pe sobolani,
modificandu-li-se formula leucocitara. Li se scot neutrofilele si li se introduce un numar egal de eozinofile. Ulterior, soarecii mor din cauza unor infectii cu germeni obisnuiti. Concluzia este ca unele eozinofile sunt capabile de fagocitoza dar ele nu pot suplini lipsa neutrofilelor.
· Apararea in infectiile cu paraziti multicelulari. In infectiile parazitare, celulele sistemului imun detecteaza celulele straine, secreta o serie de molecule implicate in comunicarea intercelulara, numite limfokine care determina elaborarea unui raspuns mai specific fata de acest tip de infectie. Ca urmare, creste numarul de eozinofile, creste concentratia unui anume tip de anticorpi, din familia IgE.
Moleculele de anticopri se ataseaza de parazit, recunoscand proteinele de suprafata. Eozinofilele au receptori ce recunosc celalalt domeniu al anticorpului si prin intermediul lui se ataseaza de celula parazitara. Apoi are loc procesul de degranulare care este citotoxic pentru celulele parazitare. Procesul de degranulare se face in spatiul extracelular si prin urmare el poate provoca un raspuns si din partea celulelor proprii ale organismului. Pot aparea astfel asa numitele boli alergice, caracterizate printr-o hipersensibilitate a propriilor celule la actiunea eozinofilelor asupra parazitilor. Aceasta hipersensibilitate se poate manifesta chiar in absenta parazitului si prezenta doar a antigenului.
Bazofilele
Sunt celule cu proprietati asemanatoare cu mastocitele.Numarul lor este intre 15-50 / mmc, aproximativ 100 milioane in total in organism.Au durata scurta de viata. In sange se gasesc sub forma de celule diferentiate, ce nu se divid.
62

La MO se stabilesc diametrul - 12-25 mm, raportul nucleu / citoplasma - 1 : 1, forma nucleului, de potcoava la cele tinere sau bilobat la cele adulte. Citoplasma este abundenta, si adesea tinde sa aiba o tenta bazofila. Exista granule foarte neuniforme d.p.d.v. al conturului care se coloreaza cu un colorant bazic si apar in albastru inchis pana la negru. Granulele se suprapun si peste nucleu. Continutul granulelor este hidrosolubil, si de aceea, pe frotiu, la spalare intensa, continutul granulelor se poate dizolva.
La ME se vad organitele: aparatul Golgi, RE, mitocondrii si incluziunile - granule de glicogen.
Granulele sunt rotunde sau ovale si specifice. Au un continut de histamina, heparina si alti mediatori. Nu au enzime lizozomale.
Rolul lor functional este incert. Datorita faptului ca reprezinta o proportie mica din totalul de leucocite adesea exista tendinta de a se considera neimportante. Degranularea lor rapida poate avea insa consecinte grave. In aceste cazuri au loc pusee hipersensibile, cand se elibereaza anticorpi IgE.
Bazofilele au pe suprafata lor receptori pentru IgE. Cand o persoana hiperreactiva produce IgE acestea pe de o parte circula in sange, pe de alta parte se leaga de bazofile. La reexpunerea acelei persoane la acelasi antigen, bazofilele cu anticoprii atasati fixeaza antigenul respectiv, se degranuleaza si astfel se pot produce diferite tipuri de reactii alergice, de la astm bronsic pana la urticarie acuta si chiar soc anafilactic, care este cel mai grav dintre bolile alergice.
Limfocitele
Exista atat in sange cat si in organele limfoide. Ca numar sunt 5-10 miliarde in sange si 2*1012 in diferite organe limfoide.Durata de viata este lunga deoarece cea mai mare parte sunt recirculate: sange ® organe
limfoide ® sange ®organe limfoide ® sange ® ..........O particularitate a limfocitelor consta in afinitatea proprie fiecarui limfocit pentru un anume
organ limfoid. Pe suprafata limfocitului se exprima un receptor pentru care exista un contrareceptor potrivit in endoteliul organului limfoid.
La MO, dupa dimensiuni, se pot distinge 3 categorii de limfocite: mici (cu diametrul 6-8 mm), mari (diametrul de 12-18 mm) si intermediare cu dimensiuni intermediare. Raportul nucleu / citplasama este de 4 : 1 sau 3 : 1. Forma nucleului este rotunda sau indentata, pozitia lui este excentrica, nucleolul este absent. Nucleul se coloreaza cu colorant bazic si apare in “pata de cerneala”. Citoplasma este ca un inel subtire perinuclear si este albastra ca “cerul senin”.
Conform teoriei clasice limfocitele apartin agranulocitelor. Insa azi se considera ca unele limfocite au granule in citoplasma la fel ca unele monocite.
S-a stabilit ca exista mai multe populatii celulare: unele se maturizeaza in timus si sunt deci timodependente - limfocitele T. Altele se maturizeaza in maduva osoasa la om - limfocitele B (bone marrow). Acestea din urma, la pasare, se dezvolta in bursa lui Fabricius.
Dupa studierea la ME s-a stabilit ca nu exista diferente intre limfocitele B si T in stare inactiva. In stare activata apar mici diferente:
· limfocitele T activate au diametrul mai mare decat al celulei inactive, nucleul mai putin condensat, iar in citoplasma apar cateva cisterne ale RE si cativa ribozomi liberi.
· limfocitele B activate au dimensiuni mult mai mari decat ale celulei inactive, iar citoplasma este plina cu RER. Aceste celule sunt specializate in sinteza anticorpilor care se face in stadiul final de diferentiere - plasmocitul.Sistemul imun cuprinde milioane de clone (familii) de limfocite. Limfocitele din fiecare
familie poseda un receptor de suprafata unic - apartine doar acestei familii. Acest receptor permite legarea unui determinant antigenic specific.
63

Cele doua mari populatii de limfocite sunt limfocitele B, specializate in sinteza de anticorpi, si limfocitele T care participa la raspunsul imun. Diferentierea la MO intre cele doua clase nu se poate face iar la ME exista diferente doar intre formele activate ale limfocitelor, diferente care se evidentiaza prin metode imunocitochimice (identificarea unor markeri de suprafata).
Fenomene de memorie imunologica si toleranta imunaIn evolutia limfocitelor exista 3 stadii potentiale in functie de intalnirea cu antigenul pe care
il recunoaste.Limfocitele naive nu au avut contact cu antigenul. Prima intalnire cu antigenul poate
determina comportament diferit din partea limfoctelor: unele determina raspuns imun specific iar altele nu determina raspunsul imun ci doar prolifereaza intens aparand un set de limfocite denumite limfocite cu memorie biologica.
Limfocitele cu memorie, la o alta intalnire cu antigenul respectiv, elaboreaza un raspuns imun prompt, rapid. In cursul elaborarii raspunsului imun pot lua nastere familii de limfocite care recunosc drept straine celulele proprii ale organismului. Lipsa de raspuns a sistemului fata de structurile proprii se numeste toleranta imuna. Mecanismul de instalare e legat fie de distrugerea clonelor autoreactive fie de inactivarea lor. Exista un grup de boli denumite boli autoimune, in care apar defecte ale raspunsului imun, sistemul imun reactionand asupra propriilor structuri.
Limfocitele B sintetizeaza anticorpi. Caracteristicile anticorpilor:· Au unul sau doua locuri de legare pentru proteinele straine.· Au domenii ce pot activa sistemul complementului sau pot fi recunoscute de receptorii de pe
suprafata fagocitelor. Reprezentantii unei singure familii de limfocite elaboreaza un singur tip de anticorp.
· Are forma literei Y, avand doua lanturi polipeptidice lungi (cu masa moleculara mare - heavy H) si doua lanturi polipeptidice scurte (cu masa moleculara mai mica - light L). Zona centrala este o zona balama.
Exista 5 clase de anticorpi: IgA, IgD, IgE, IgG, IgM, care difera intre ele prin structura lantului greu.
Moleculele de anticorpi sunt fixate pe membrana limfocitului dar sunt si eliberate in circulatie.
Orice vaccin induce proliferarea familiei de limfocite care produce anticorpul ce recunoaste antigenul introdus.
Limfocitele T cuprind doua subpopulatii: limfocitele T citotoxice si limfocitele T helper, care au pe suprafata receptori ce se aseamana dar cele doua subpopulatii difera d.p.d.v. functional. Limfocitele T citotoxice lizeaza direct celulele infectate (mai ales viral) iar limfocitele T helper faciliteaza producerea de anticorpi de catre limfocitele B si ingerarea si distrugerea celulelor infectate de catre macrofage. Limfocitele T detecteaza si celulele proprii ca fiind nonself.
Receptorii limfocitelor T recunosc proteinele straine atunci cand sunt legate de un set de proteine denumite moleculele sistemului major de histocompatibilitate (CMH). Exista 2 clase de complexe de histocompatibilitate: clasa I si clasa II. Cele din clasa I se gasesc pe membrana aproape a tuturor celulelor nucleate. Moleculele CMH II se gasesc pe membrana celulelor ce coopereaza cu limfocitele T. Acest sistem s-a descoperit in cursul cercetarilor pentru refuzul transplantelor. Acest sistem arata ca celulele organismului sunt sanatoase.
Limfocitele T citotoxice provoaca moartea celulelor ce poarta pe suprafata o proteina straina legata de moleculele CMH. Moartea celulei este provocata prin doua mecanisme:1. mecanismul lung2. mecanismul scurt: limfocitul T elibereaza o proteina care se numeste perforina care
polimerizeaza si formeaza canale transmembranare in membrana celulei tinta. Apoi din
64

citoplasma, prin pori trec cateva enzime ce activeaza verigile finale ale cascadei biochimice. Proteozomul este o “masinarie ce taie”celulele infectate.
Limfocitele T helper au mecanism de actiune asemanator ca etape iar rezultatul final consta in eliberarea de molecule semnal de catre limfocitul T helper cat si de catre celula cu proteina straina. Diferenta la acest mecanism este legata de tipul de celule implicate: celule prezentatoare de antigen si de tipul de molecule ce prezinta moleculele straine, care in acest caz apartin CMH II.
Dupa natura lor, semnalele chimice se impart in:· substante chemotactice (“cheama” si alte celule)· interleukine (determina proliferarea unor seturi de celule albe)
In infectia cu HIV, unele celule infectate pot exprima pe suprafata lor proteine-semnal ce intorc semnalul primit de la limfocitele T citotoxice (ex.:moarte celulara). In cazul camerei anterioare a globului ocular sau a testiculului prezinta continuu un semnal de apoptoza, astfel incat orice celula ajunsa aici moare (“moartea intrusilor”), lucru care este folositor in transplanturi.
Monocitele
Sunt celule aflate in trecere prin sange unde stau 1/2-1 zi. Ele ajung in tesuturi unde isi schimba morfologia (se transforma in macrofage). Exemple: macrofagele din tesutul conjunctiv din ficat, din plaman.
Sunt in numar de 300-500 / mmc.La MO apar ca fiind celule mari, cu diametrul pana la 20 mm. Raportul nucleu / citoplasma
este 2 : 1 iar nucleul are forma de potcoava. In nucleu eucromatina si heterocromatina alterneaza, dand aspectul de tabla de sah. Citoplasma este bazofila si apare colorata in albastru intens. Uneori exista granulatii mici ce apar roz palid.
La ME se observa RE, ribozomi liberi si mitocondrii. Granulele au enzime lizozomale.Rolul fagocitelor mononucleare:
1. secreta diversi compusi (proteici sau neproteici)2. ingera diferite particule de dimensiuni mari (celule intregi sau resturi celulare)3. coopereaza cu limfocitele mai ales4. fagocitoza (functia principala) - ingera celulele imbatranite (vezi splina pentru eritrocite).
Eritrocitele exprima semnale de degenerescenta care sunt recunoscute si fagocitate. De asemenea fagocitele pot inghiti proteine straine, resturi celulare, produsi fiziologici formati in exces (factori de coagulare). Mecanismul de transport poate fi pinocitoza (pentru faza fluida), endocitoza (pentru particulele solide) sau fagocitoza (vezi neutrofilul).
Diferente intre neutrofil si macrofag:· macrofagul utilizeaza un set amplu de molecule de adeziune· macrofagul poate fagocita particule dupa ce au fost opsonizate cu mai multe tipuri de anticorpi.· macrofagele se pot deplasa sub actiunea factorilor chemotactici sintetizati de limfocite· macrofagele au receptori pentru manoza (se gaseste in capsula bacteriana) si astfel bacteria poate
fi fagocitata fara opsonizare.
HEMATOPOIEZA
Celulele, inainte de a se definitiva au o serie de etape intermediare. Exista multe etape descrise. Celulele sangvine provin din celulele stem din maduva hematopoietica. Celulele stem
65

nu au pe suprafata lor determinanti antigenici. De aceea celulele stem sunt folosite pentru a introduce in organism anumite gene care sunt absente in diverse maladii.
Hematopoieza este procesul care asigura formarea si reinnoirea continua a celulelor sangelui periferic. In general se pastreaza un numar relativ constant de celule circulante iar hematopieza permite si ajustarea necesitatilor organismului de anumite tipuri celulare la un anumit moment dat. Astfel, in hemoragii este crescuta sinteza de hematii, in infectii este crecuta sinteza de granulocite etc.
Localizarea tesuturilor hematopoietice - la adult este maduva rosie osoasa = maduva rosie hematopoietica (MRH). Hematopoieza este realizata mai intai prin procese de multiplicare a celulelor stem si a celulelor precursoare, apoi urmeaza procesele de diferntiere, cand se intrerup diviziunile iar celulele exprima proteine specifice pe membrana iar in final are loc citodiaboza care consta in eliberarea celulelor mature in circulatie.
Hematopoieza este un proces autointretinut. Mecanismele de reglare a hematopoiezei sunt multe si complexe, fiind implicate molecule semnal, factori de crestere etc.
Etapele hematopoiezei
Etapa prenatalaExista mai multe locuri:
· etapa mezoblastica (mezenchimala, prehepatica) care dureaza in perioada L1-L3 de viata intrauterina (I.U.). Hematopoieza are loc in insulele sangvine din peretele sacului vitelin. Apar primii precursori ai celulelor liniei rosii, care au doua particularitati: sunt nucleate si contin Hb Gower - embrinara (a2e2).
· etapa hepatosplenica: se suprapune partial peste prima etapa, durand L2-L9 I.U. La un anumit moment este cea mai importanta (in L4-L5 I.U.) dupa care scade ca importanta datorita etapei mieloide. In L3-L4 apar granulocitele iar in L5 apar limfocitele. Precursorii liniei rosii au HbF (fetala) - a2g2. Productia de HbF scade treptat, la nastere fiind de 2% din totalul de Hb pentru ca mai tarziu sa dispara.
· etapa mieloida - debuteza semnificativ la sfarsitul L3 - inceputul L4 I.U. Ea creste ca importanta in a doua jumatate a sarcinii. Precursorii liniei rosii au HbA (adult - a2b2).
Conceptia actuala asupra hematopoiezei1. Celula stem are capacitate migratorie asemenea limfocitelor, care au origine in MRH si se gasesc
in tranzit in sangele periferic. Dintr-o celula stem rezulta 1000 de celule din sangele periferic.2. Celula stem pluripotenta are morfologia necunoscuta. Ar fi asemanatoare monocitului sau
limfocitului. 3. Celulele stem circulante pot coloniza: ajung din maduva osoasa in splina sau ficat care devin
locuri de hematopoieza extramedulara.4. Celula stem sta la originea tuturor liniilor hematopoietice. Celula stem pluripotenta reprezinta o
populatie de celule nediferentiate, capabile de diferentiere in toate liniile hematopoietice.5. Celulele stem au capacitatea de autoreinnoire. Daca o celula stem se divide, se formeaza doua
celule identice. Una o inlocuiste pe cea de provenienta, inactiva functional iar cealalta prolifereaza dand nastere elementelor figurate. Cu cat sunt mai nediferentiate, cu atat celulele au capacitate mai mare de autoreinnoire.
Descoperirea celulelor stem s-a facut la soareci, folosind celule marcate. S-a descoperit ca existau celule ce repopulau ficatul sau splina la soarecii iradiati. S-a observat ca exitau colonii de celule ce erau formate de o singura celula de origine. Aceste celule ce determinau colonii au fost numite unitati formatoare de colonii (celulele stem sunt deci colony forming units - CFU).
Exista multi factori de reglaj ce orienteaza hematopoieza spre o linie celulara. Spre exemplu eritropoietina orienteaza spre linia rosie, granulopoietina duce la formarea de granulocite iar trombopoietina duce la formarea de plachete. Cantitati crescute de eritropoietina duc la stimularea
66

puternica a productiei de eritrocite. Deci, mecanismele de control al diviziunii si diferentierii celulelor stem sunt multe si complexe.
Nomenclatura celulelor stem
Celulele stem ce determina liniile au grad de potentialitate diferit. Spre exemplu unele celule stem pot determina toate liniile (sunt totipotente) si au viata mai lunga si capacitate de autoreinnoire crescuta. Alte celule sunt pluripotente - formeaza mai multe linii dar nu toate, altele sunt bipotente formeaza 2 linii iar altele sunt unipotente, formand o singura linie.
Se considera ca prin diviziunea celulei totipotente scade potentialitatea celulei, rezultand o celula stem pluripotenta. Dincolo de maduva spinarii exista doar celula precursoare care si-au pierdut capacitatea de reinnoire, fiind capabile doar de diferentiere.
Criterii morfologice generale pentru liniile celulareSe permite astfel incadrarea unei celule intr-o linie celulara si plasarea intr-un anumit stadiu.
Aceste criterii sunt:1. marimea celulalor: cu cat celula este mai tanara, cu atat este mai mare.2. cu cat nucleul este mai mare cu atat celula este mai tanara3. cu cat raportul nucleu / citoplasma este mai mare cu atat celula este mai tanara4. cu cat materialul nuclear este mai eucromatic (mai putin condensat) cu atat celula este mai
tanara. Aceasta se explica prin faptul ca o celula tanara transcrie cat mai multe gene pentru a le traduce in proteine.
5. Cu cat in citoplasma sunt mai multi ribozomi si RE cu atat celula este mai tanara. La MO citoplasma este cu atat mai bazofila cu cat celula este mai tanara, datorita prezentei unui numar mare de poliribozomi)
6. Cu cat celula are mai multi nucleoli cu ata ea este mai tanara. Numarul acestora poate varia: 1,2-5.
7. Existenta produsilor caracteristici unei linii celulare: granule (dupa forma, dimensiunea si afinitatea tinctoriala aacestora se poate stabili carei linii granulocitare ii apartine celula). In linia rosie, acumularea Hb in citoplasma determina aparitia acidofiliei.
Pe masura inaintarii pe linia celulara scade volumul celulei, scade volumul nucleului care devine mai compact (se transcrie un numar mai redus de gene pentru proteinele specifice), scade numarul de organite, bazofilia citoplasmei se atenueaza.
Astfel:· blast = celula tanara· cit = celula evoluata· meta = dupa· pro = inainte
Exemplu: In linia rosie, initial citoplasma este bazofila iar in final este acidofila. In etapa intermediara coexista acidofilia si bazofilia celula fiind policromatofila.
In maduva trebuie sa existe un raport fix intre precursorii unei anumite linii.
Seria rosie - eritropoieza
Celula stem ® proeritroblast ® eritroblast bazofil ® eritroblast policromatofil ® eritroblast acidofil ® eritrocit. In linia rosie nucleul este rotund. Cantitatea de ARN scade pe parcursul evolutiei datorita scaderii sintezei proteice.
Proeritroblastul
67

· Diametru 20-25 mm· Nucleu ocupa 80% din volumul celulr, este eucromatic, prezinta mai multi nucleoli· Citoplasma este intens bazofila
Eritroblastul bazofil· Diametru 16-18 mm· Nucleu necondensat, uneori se vad nucleoli, alteori nu; uneori prezinta halou perinuclear· Citoplasma mai putin intens bazofila
Eritroblastul policromatofil· Diametru 15 mm· Nucleu destul de condensat (heterocromatina abundenta) · Citoplasma isi modifica nuanta: exista putini ribozomi si apare Hb in citoplasma, deci coexista
acidofilia si bazofilia. La ME se observa ca numarul de organite scade foarte mult.
Eritroblast ortocromatofil (aceeasi nuanta cu celula adulta - acidofil)· Diametru 10-12mm· Nucleu condensat la maxim (exista doar heterocromatina), nu are nucleoli· Citoplasma contine o cantitate crescuta de Hb care ii determina acidofilia. La ME se observa
eliminarea celor mai multe dintre organite.In stadiul final celulele expulzeaza nucleul, membrana celulara se reface si se formeaza
reticulocitul.
Reticulocitul· Diametru 7mm· Nucleu - nu exista· Citoplasma este diferita de cea a hematiilor adulte prin prezenta catorva resturi de organite
Ele raman 12-48 de ore in circulatie dupa care sunt eliminate. Se evidentiaza cu o coloratie speciala - albastru de Crecil stralucitor. Reticulocitele reprezinta 1% din elementele circulante. Un numar crescut de reticulocite arata o anemie.
Pentru seria rosie, maduva nu este un compartiment de depozit, toatee elementele liniei rosii fiind eliberate in circulatie, spre deosebire de celulele granulocitare ce pot stocate in maduva atunci cand organismul le necesita.
In maduva, raportul intre precursorii liniei rosii si precursorii linie granulocitarea este invers fata de formula leucocitara, in maduva existande de 3-4 ori mai multi precursori ai granulocitelor decat ai eritrocitelor.
Nucleii eliminati de eritroblasti sunt fagocitati de macrofagele din MRH. De aceea exista o asociere intre hematopoieza si aceste macrofage.
Timpul necesar este de 20 de ore pentru primele etape, pentru ca ultimele etape sa se desfasoare lent.
Seria granulocitara
Celula stem ® mieloblast ® promielocit ® mielocit (neutrofil, eozinofil sau bazofil) ® metamielocit (neutrofil, eozinofil sau bazofil) ® granulocit nesegmentat ® granulocit segmentat.
Mieloblast· Diametru 20 mm· Nucleu mare, usor indentat sau oval, isi modifica conturul, are cromatina laxa si multi nucleoli
raportul nucleu/citoplasma = 4 : 1,
68

· Citoplasma este putina, are tenta bazofila si nu are granule.Proeritroblastul seamana cu mieloblastul.
Promielocitul· Diametru 10-20 mm· Nucleu scade in volum, este compact, se observa cativa nucleoli· Citoplasma este mai multa (volumul celular fiind constant) si se detecteaza primele granulatii
(granulele primare, nespecifice, azurofile)
De la acest punct mai departe cele trei linii granulocitare se diferentiaza, dupa cum urmeaza:
Mielocitul neutrofil· Diametru 18 mm (celula isi reduce volumul)· Nucleul se compactizeaza mult, are forma ovala, uneori rotund; materialul nuclear este granulat,
heterocromatic, uneori prezinta nucleoli, alteori nu; raportul nucleu / citoplasma este 2 : 1 pana la 1 : 1.
· Citoplasma are nuanta tranzitorie intre bazofil si acidofil (albastru - rosu); prezinta ambele tipuri de granulatii
Metamielocitul neutrofil· Diametru 12-18 mm (volum relativ constant)· Nucleul - conturul sau prefigureaza forma nucleului la neutrofilul segemntat (potcoava), are
heterocromatina, nu exista nucleoli, raportul nucleu / citoplasma este 1 :1· Citoplasma isi modifica afinitatea tinctorial (unele sunt tranzitorii iar altele au citoplasam
acidofila); apar granulele specifice, secundare.
Neutrofilele nesegmentate· Diametru este mai mic, pana la 16 mm· Nucleu in banda sau sub forma de potcoava, nesegmentat· Citoplasma este la fel ca la neutrofilul adult
Din neutrofilul nesegmentat se formeaza neutrofilul segmentat si uneori neutrofilul hipersegmentat.
Mielocitul eozinofil· Diametru 18 mm· Nucleu - forma sa prefigureaza forma nucleului viitoarei celule; sinteza proteinelor incepe sa
scada, cromatin este mai agregata, uneori are nucleoli, alteori nu; raportul nucleu / citoplasma este 2 : 1 sau 1 : 1.
· Citoplasma are culoare tranzitorie (albastru-rosiatic); granulele sunt eozinofile
Metamielocitul eozinofil· Diametru 18 mm· Nucleu are forma precis conturata (potcava, rinichi), cromatina este mai compacta; scade
raportul nucleu / citoplasma pana la 1 : 1· Citoplasma este acidofila, granulele sunt numeroase cantitativ
Eozinofilul nesegmentat (tanar)· Diametru 16 mm· Nucleu cu forma de potcoava, raportul nucleu / citoplasma este 1 : 1
69

· Citoplasma este identica cu cea a eozinofilului matur
Mielocitul bazofil· Diametru 18 mm· Nucleu oval sau indentat, cromatina usor impachetata, raportul nucleu / citoplasma este 2 : 1
pana la 1 : 1· Citoplasma are tenta albastruie, moderata cantitativ si apar granule specifice, intens colorate
Metamielocitul bazofil· Diametru 18 mm· Nucleu in forma de potcoava, cromatina intens colorata, nucleolii nu se vad; scade usor raportul
nucleu / citoplasma· Citoplasma are tenta albastruie si granule specifice in numar mare
Bazofilul tanar nesegmentatAre toate caaracteristicile bazofilului adult mai putin forma nucleului care este alungita,
nesegmentata.
Seria monocitara
Celula stem ® monoblast ® monocit tanar ® monocit adult
Monoblastul· Diametru 12-20 mm (se pastreaza pe toata seria)· Nucleu este rotund, oval, usor plicaturat, compactat, volumul sau scazand de-a lungul liniei
monocitare; raportul nucleu / citoplasma este 4 : 1.· Citoplasma este bazofila, nu se vad granulatii
Monocitul tanar· Diametru 12-20 mm· Nucleu cu forma si aspect apropiat de cel adult (plicaturat cu cromatina dispusa in tabla de sah),
nucleolul finnd uneori prezent; raportul nucleu / citoplasma 3 : 1 pana la 2 : 1· Citoplasma devine abundenta datorita compactarii nucleului, are tenta bazofila, exista granule
fine, azurofile, vizibile la limita de rezolutie a MO.
Seria limfocitara
Celula stem ® limfoblast ® limfocit tanar ® limfocit adultNu toate etapele de diferentiere au loc in MRH, unele limfocite diferentiindu-se si in timus.
Limfoblastele· Diametru 20 mm (celula mare)· Nucleu este mai putin compactat, prezinta 1-2 nucleoli; se pastreaza raportul nucleu / citoplasma
de 4 : 1 pe toata linia (caracteristica a limfocitelor)· Citoplasma este putina si bazofila; nu se vad granule citoplasmatice
Limfocitul tanar
70

· Diametru 9-18 mm (dupa populatia careia ii apartin d.p.d.v morfologic - mari, intermediare, mici)
· Nucleu este dens, fixeaza o cantitate mare de colorant bazic, are rar nucleoli· Citoplasma - nu se identifica prezenta granulelor
Seria trombocitara (plachetara)
Celula stem ® megacarioblast ® promegacariocit ® megacariocit trombocitogen ® trombocite.
Precursorii au talie foarte foarte mare, iar diametrul lor creste pe masura ce se apropie de faza finala. Nucleul este foarte mare si creste in dimensiuni de-a lungul liniei deoarece precursorii isi multiplica numarul de cromozomi fara diviziune. Se obtin celule cu grad inalt de ploidie.
Megacarioblastul· Diametru 50 mm· Nucleu cu mai multi nucleoli, cromatina este putin compactata; raportul nucleu / citoplasma este
de 4 : 1· Citoplasma este bazofila
Promegacariocitul· Diametru 80 mm· Nucleu cu cromatina mai compacata dar oricum laxa, raportul nucleu / citoplasma variaza de la 4
: 1 pana la 1 : 1· Citoplasma cu mici granule azurofile
Megacariocitul trombocitogen· Diametru pana la 160 mm· Nucleu lobulat, intens impachetat, colorat bazic, fara nucleoli; raportul nucleu / citoplasma
variaza de la 1 : 1 pan la 1 : 12· Citoplasma are o tenta palid albastruie cu granule azurofile distribite uniform
La sfarsitul perioadei de diferentiere megacarioblastele se ataseaza la peretele vascular, membrana lor emite prelungiri plicaturate, trec prin peretele vascular iar fluxul snagvin detaseaza din citoplasma megacariocitului plachete. Daca cea mai mare parte a megacariocitului.................. de autodistrugere...........
Prezenta acestor celule este evidentiata prin frotiu de MRH.Exista un raport considerat normal intre precursori care constituie mielograma
(medulograma).· 1 mmc MRH contine 70.000 - 100.000 de cariocite.· granulocite: neutrofile 50-93%
eozinofile 1-7%bazofile - rare
· eritrocite - 13-25,5% - la 100 de granulocite· limfocite 1-10 % - la 100 de granulocite· monocite, plasmocite, histiocite - 3-10% - la 100 de granulocite· megacariocite 1-8‰ - la 100 de granulocite
Aceste valori sunt variabile.
71

TESUTUL MUSCULAR
Mecanismele de contractie-relaxare se realizeaza prin interactia intre doua tipuri de proteine. Aceasta interactiune este controlata de oscilatia concentratiei ionilor de Ca2+ care este mesagerul secund.
Exista trei tipuri de muschi:1. striat scheletic2. striat cardiac3. neted
Muschiul striat scheletic
Localizare:· toti muschii somatici· la nivelul organelor interne:
· m. cavitatii· m. faringelui
72

· m. din 1/3 superioara a esofagului· sfincterul anal extern· sfincterul uretral extern· laringe· m. extrinseci ai globului ocular· m. urechii mediiCelulele si fibrele musculare au lungimi foarte mari, de ordinul mm sau cm. Raprtul
lungime / latime este unic in fiecare organ. Grosimea este de 25-100 mm. Celulele musculare din componenta m. scheletic sunt asociate cu tesut conjunctiv. Celulele musculare sunt asezate paralel cu axul lung orientat de-a lungul directiei de miscare a m. respectiv. In sectiune transvaersala au forma poligonala.
Fiecare celula musculara este imbracata de o structura conjunctiva numita endomisium. Aceast tesut este format din fibre de reticulina, colagen, vase sangvine si fibre nervoase. Mai multe fibre musculare alcatuiesc un fascicul primar. Mai multe fascicule primare alcatuiesc un fascicul secundar. Fiecare fascicul este imbracat de perimisium care este format din vase sangvine si fibre nervoase de calibru mai mare. Intreg muschiul este imbracat de epimisium prin care se ataseaza de structurile invecinate. Rolul acestor structuri conjunctive este de a pune in valoare capacitatea contractila a celulei musculare.
Celulele sunt de forma cilindrica, neramificate si asezate paralel una cu alta. Celulele sunt multinucleate iar nucleii sunt asezati in imediata vecinatate a membranei si au o dispozitie foarte ordonata, alternanata. Cea mai mare parte a citoplasmei este ocupata de o structura ce apare sub forma de striatii. Aspectul striat al citoplasmei este dat de organitele specifice, miofibrilele care sunt striate.
In sectiune transversala celulele au aspect poligonal. In functie de locul unde s-a efectuat sectiunea, se pot vedea 1, 2 sau chiar 3 nuclei. Citoplasma contine miofibrilele sectionate transversal ce apar sub forma unor puncte separate de fante citoplasmatice. Se mai obsrva raportul cu celulele conjunctive si vasele sangvine.
Celulele musculare se deosebesc de celulele obisnuite prin aspecte ultrastructurale observate cu ajutorul ME. Astfel celula contine:
· membrana - sarcolema· citoplasma - sarcoplasma· organite comune si organite specifice (miofibrilele). (Sarcos - gr. - carne)
Sarcolema are acelasi model structural ca membrana celulara in general. Ea este alcatuita dintr-o plasmalema cu glicocalix. La exterior prezinta o membrana bazala formata de matricea celulara condensata. Caracteristic sarcolemei este faptul ca mambrana bazala poseda un sistem tubular, sistemul tubilor T (transversi) care reprezinta o prelungire a spatiului extracelular in interiorul celulei pana in vecinatatea miofibrilelor. Din loc in loc exista structuri sub forma unor invaginari ale sarcolemei cu diametru de 100 nm care poarta numele de caveole. La acest nivel sunt adunate toate componentele unor cai de semnalizare transmembranara.
Organite specificeMiofibrilele care prezinta doua tipuri de benzi.
· banda A - banda anizotropa, banda intunecata. · banda H - o regiune mai palida aflata la jumatatea benzii A. · banda M (de mijloc) se afla la mijlocul benzii H.· banda I - banda izotropa de lumina polarizata (clara).· banda Z este o regiune mai intunecata aflata la mijlocul benzii I.
D.p.d.v. functional, intre doua benzi Z consecutive se delimiteaza SARCOMERUL cu lungimea de 25 mm. Acesta este alcatuit din o banda A intunecata intraga si cate o jumatate din cele doua benzi I adiacente.
73

In timpul contractiei se scurteaza banda I.
MiofilamenteleSunt subtiri, formate din actina sau groase formate din miozina. Miofilamentele reprezinta
agregate complexe supramoleculare formate din proteine contractile si reglatoare si alte structuri cu rol de suport.
Filamentele subtiri sunt formate din actina si proteine reglatoare (tropomiozina si troponina).Actina este alcatuita din actina F (fibrilara) al carui lant formeaza un dublu helix. Prin
polimerizarea actinei F se formeaza actina G (globulara). Fiecare monomer de actin poseda un loc de interactiune cu miozina.
Tropomiozina este o proteina fibrilara. Ea este infasurata si asezata in santul pe care il formeaza actina F. De moleculele de tropomiozina sunt atasate la distanta de 400 Å (7 monomeri de actina G) complexe de troponina.
Troponina prezinta 3 subunitati: · troponina C care leaga ionii Ca2+.· troponina T care se ataseaza de tropomiozina· troponina I (inhibitoare) care previne interactiunea actina-miozina.
Atunci cand celula este in repaus concentratia Ca2+ din celula este atat de mica incat nu este suficienta pentru a ocupa locurile de legare de pe troponina C. Ca urmare configuratia spatiala a troponinei este impusa de troponina I. Aceasta subunitate face ca ansamblul troponina-tropomiozina sa acopere toate locurile de interactiune actina-miozina.
Filamentele groase contin doar molecule de miozina. Meromiozina are doua lanturi unul usor (L) si unul greu (H). Miozina este o molecula complexa formata din mai multe lanturi grele si usoare. Molecula de miozina are o parte orizontala si o proeminenta globulara la unul din capete. In componenta ei lanturile grele se intind pe toata lungimea moleculei (atat in portiunea orizontala cat si in portiunea globulara). Pe lanturile grele, la nivelul proeminentei globulare se afla sediul ATPazei. La nivelul proeminentelor se afla si lanturile usoare.
Moleculele de miozina sunt asamblate pentru a da nastere filamentelor groase in seturi antiparalele. Astfel, pe o jumatate a filamentelor groase proeminenta la terala este orientata intr-o directie iar pe cealalta jumatate este orientata in directia opusa, partea centrala fiind lipsita de proeminente.
Reticulul sarcoplasmic (RS)Ii este caracteristica capacitatea de a stoca o cantitate mare de Ca2+. La ME se observa ca RS
este de doua feluri:· jonctional - RS prezinta o regiune cu cisterne mai dilatate care fac jonctiunea cu sistemul tubilor
T.· liber - partea din RS ce nu vine in contact cu sistemul tubilor T.
In interior Ca2+este legat de o proteina care se numeste calsechestrina. Aceasta asigura o concentratie de Ca in RS de 10.000 - 100.000 de ori mai mare decat in citosol. Acumularea Ca in RS se face cu ajutorul unei proteine din membrana RS (calciuATPaza) care preia Ca din citoplasma in RS. In membrana RS exista o serie de canale ionice ce permit eliberarea rapida a Ca in virtutea gradientului de concentratie dinspre RS spre citoplasma atunci cand se declanseaza contractia.
Organite comuneAu foarte mult mitocondrii care asigura sinteza de ATP sursa de energie. Ele sunt asezate in
siruri printre miofibrile.
74

Pentru m. scheletic s-a descris la MO triada care este formata din doua terminatii de RS jonctional si un segment de tubi T. Triada reprezinta o deosebire ultrastructurala intre m. scheletic si cel cardiac.
Teoria contractiei musculare (1957) - mecanismul “puntilor transversale” elaborat de Huxley. Mecanismul contractiei se bazeaza pe cicluri de asocieri si disocieri intre monomerii de actina si proeminentele laterale ale miozinei. In timpul contractiei nici unul dintre cele doua filamente nu se scurteaza ci gliseaza, aluneca unele printre altele. Se scurteaza insa distanta dintre doua benzi Z. Filamentele subtiri aluneca spre centrul sarcomerului. La fiecare ciclu de atasare-dezatasare actina-miozina filamentele se deplaseaza cu 5-10 nm.
Relaxarea musculara inseamna revenirea la starea initiala.In timpul contractiei, prin intermediul tubilor t se transmite o unda de depolarizare. In RS
exista un sistem care detecteaza schimbarea de potential si ca uramre se deschid canalele de Ca si Ca este eliberat in citoplasma. Atunci creste concentratia de Ca in citoplasma si creste astfel si probabilitatea ca subunitatile C ale troponinei sa ataseze Ca. In acest moment se modifica asezarea in spatiu a complexului troponinei, ridicandu-se actiunea inhibitorie a troponinei I. Complexul troponina-tropomiozina elibereaza atunci locul de interactiun de actina si se realizeaza interactiunea actina-miozina. In acest moment se da posibilitetea de actiune a ATPazei care se gaseste pe proeminentele laterale ale miozinei. Actina (filamentele subtiri) este trasa (gliseaza).
Apoi intra in actiune mecanismele de reacumulare a Ca in RS, concentratia Ca in citoplasma scazand pana se atinge “concentratia critica”. In acest moment Ca se desprinde de pe subunitatea C a troponinei si complexul troponina-tropomiozina revine la starea initiala, de repaus.
Regenerarea celulei musculare striateDesi celulele musculare sunt ultraspecializate ele au posibilitatea de a se regenera dupa un
traumatism cu pierdere musculara, lucru care se petrece prin intermediul celulelor satelite, observabile doar la ME. Celulele satelite sunt mici, putine, nediferentiate si situate in jurul celulelor adulte. Ele se aseamana cu mioblastii embrionari si au capacitate regenerativa limitata. In pierderi musculare masive, regenerarea se face prin tesut cicatricial, de natura conjunctiva.
Muschiul striat cardiac
Este localizat in miocard. Sunt in total aprox. 2 miliarde de celule. HIPERTROFIE - cresterea volumului organului prin cresterea volumului elementelor
componente (celulelor).HIPERPLAZIE - cresterea volumului organului prin cresterea numarului elementelor
componente (celule).
Lungimea fibrei 100 mm, latimea (diametrul) - 15 mm. Isi pot dubla volumul, diametrul ajungand pan la 30 mm (in hipertrofie).
Celulele au un singur nucleu, asezat central. Prezinta striatii transversale care insa lipsesc in jurul nucleului. Pe masura inaintarii in varsta se acumuleaza pigmenti de imbatranire (hipofuxina). Celulel pot fi ramificate dar nu se anastomozzeaza. atasearea cap la cap a celulelor se face prin discuri intercalare care se obsrva mai bine pe preparate colorate cu hematoxilina fosfotoxica. Discurile apar ca niste ingrosari transversale in scara la capetele celulei. Ele se mai numesc striurile scalariforme Eberth. Printre celule se gasesc structuri fine conjunctivale.
75

Caracteristic acestui tip de tesut este vascularizatia de tip terminal. Astfel, celulele dintr-un anumit teritoriu sunt toate dependente de un anume vas, ceea ce explica procesele ce au loc in infarctul miocardic acut.
La ME discurile intercalare apar ca niste formatiuni alcatuite din complexe jonctionale, prezentand portiuni transversale si longitudinale. Portiunile transversale sunt alcatuite din jonctiuni cu rol strict de adeziune (dezmozomi si jonctiuni intermediare). Portiunile longitudinale sunt alcatuite din jonctiuni comunicante (gap, nexus). Acestea din urma asigura continuitatea electrica.
Si celulele cardiace au sistem de tubi T. Ele sunt in numar mai mare decat in fibrele musculare striate scheletic si stabilesc contacte doar cu o terminatie a RS formand diadele.
Dintre organitele comune cel mai bine reprezentate sunt mitocondriile care ocupa 35% din volumul celulei, restul fiind in cantitate mica.
Materialul contractil este asemanator ca organizare cu cel de la m. striat scheletic. El are dispozitie ordonata dar nu o respecta in totalitate. Contractia este controlata de oscilatiile concentratiei de Ca2+. De asememnea exista o calciuATPaza care acumuleaza Ca2+ in RS, canale de calciu in membrana RS care sunt sensibile la mediatori chimici (IP3 - inozitol trifosfat). Acesti mediatori pot deschide canalele care duc la trecerea Ca din RS in citoplasmic.
Miocardiocitele atriale sunt asemanatoare cu cele ventriculare dar au in plus granule atriale. Aceste granule contin un precursor al factorului natriuretic. Factorul natriuretic este de natura endogena ajuta la eliminarea Na+ din organism. El are efect antagonic fata de ADH si ACTH. Eliberarea granulelor se face atunci cand celulele sunt tensionate datorita cresterii volumului sangvin.
Celulele tesutului excitoconductor sunt de 3 tipuri:· Celule P (pacemaker) care genereaza impulsuri electrice in nodulul sinoatrial si nodulul
atrioventricular. Au forma rotunda sau ovala, nucleul mare este situat central. Aparatul contractil este slab reprezentat si au putine mitocondrii si cisterne de RS. Intre aceste celule se stabilesc jonctiuni desmozomale. de asemenea ele mai sunt legate si de celulele T (de tranzitie) prin discuri intercalare.
· Celulele T (de tranzitie) prezinta mai multe mitocondrii si cisterne de RS. Ele au rol de preluare si raspandire a impulsurilor electrice. Stabilesc jonctiuni cu celulele P si cu miocardiocitele de lucru.
· Celulele Purkinje sunt mai mari, au citoplasma clara. La ME se observa in vecinatatea membranei materiale contractile cu orientare paralela cu axul longitudinal al celulei. Ele au mai multe mitocondrii si lipseste sistemul tubilor T. Stabilesc jonctiuni de tip gap sau desmozomi. Nu prezinta disc intercalar.
76

Muschiul neted
IstoricObservat prima data la MO in 1841 de catre Henle si dacatre Schwann in 1847, care a
elaborat si teoria celulara. La MO celula este fusiforma, nucleu mic, situat central, citoplasma aparent omogena. La ME Csapo a observat m. neted visceral (1955) iar Fawcett (1959) si Sonebyo (1970) au observat m. neted vascular.
Inervatia a fost studiata intre anii 1950-1970. Muschiul are inervatie exclusiv vegetativa: simpatica (descoperita de Falck in 1962) si parasimpatica (descoperita de Ruska in 1967).
Fiziologia a fost studiata de catre Hidaka, Bozler (1947), Bullbring (1970) si Hidata. S-a stabilit ca celulele musculare netede au proprietatea de excitabilitate, adica sunt capabile sa declanseze un potential de actiune ce are la baza intrarea Ca in celula prin canalele de Ca. De asemenea, ele genereaza spontan (automat) potentiale de actiune, ceea ce asigura caracterul miogenic al tesutului muscular neted. Generarea potentialului de actiune se face in interiorul celulei, in absenta stimulilor exteriori. Prin aceste doua proprietati celulele musculare netede se aseamana atat cu celulele nervoase cat si cu cele musculare.
Localizare:
77

· in organele cavitare (cu perete si lumen). Peretele este format din mai multe tunici, una fiind m. neted. Exemple:· arborele respirator - de la trahee pana la ductul alveolar· arborele vascular - tot cu exceptia capilarelor si venulelor, ce au peretele subtire, fara
tunica musculara neteda dar totusi celule musculare speciale (pericite) care sunt de fapt celule contractile si ajuta la progresia coloanei sangvine
· tubul digestiv - din 1/3 mijlocie a esofagului pana la rect· aparatul urinar - de la ureter pana la uretra· aparatul genital feminin - tube uterine pana la vagin· aparatul genital masculin - epididim, canalul deferent, veziculele seminale, prostata,
penisul.· glandele exocrine:
· ductele excretoare (glandele salivare, caile biliare etc)· portiunea secretorie (acinii glandelor salivare etc) - este inconjurata de celule
mioepiteliale care ajuta la eliminarea produsului de secretie.· organele de simt:
· in piele - m. piloerectori· globii oculari - m. irisului, m. intrinseci (ciliari)
· celule izolate sau fascicule foarte mici raspandite in tesutul conjunctiv. Acestea se gasesc in:· capsulele organelor (ggl. limfatici, splina etc)· pleura viscerala· ligamentele largi ale uterului· tesutul conjunctiv subcutanat, in unele zone speciale (zona mamelonara, perineu, scrot,
orbita)
Celulele musculare netedeAspect: forma fuziforma dar exista si unele “ramoase” care sunt ramificate cu prelungiri ( in
iris, tunica medie aorta). Dimensiuni: lungimea 20 mm - 1 mm. Au lungimea de 1 mm la stomac si uterul gravid.
Diametrul este de 5 mm. Raportul diametru / lungime variaza de la 1/5 pana la 1/100, fiind mai mic fata de cel de la fibra scheletica.
Arhitectura tisulara:Exista cel putin doua tipuri de arhitectura tisulara in m. netezi:
· adaptata pentru executarea miscarilor peristaltice (la tubul digestiv). Aici exista doua straturi de muschi: exterior (longitudinal) si interior (circular). Aceste doua paturi musculare cu directie perpendiculara una pe alta se contracta automat concomitent sau consecutiv pentru a asigura progresia continutului. La fel exista la nivelul ureterului. In caile urinare m, exterior este circular iar cel interior este longitudinal.
· adaptata pentru golirea continutului in timp scurt (vezica urinara, uterul gravid). Aici fibrele sunt spiralate in jurul axului principal al organului.Structura matricei extracelulareEste alcatuita din:
· o structura asemanatoare membranei bazale epiteliale care constituie un invelis celular. Aceasta structura este alcatuita din straturi succesive, in ordine, dinspre membrana catre celula:· lamina rara -- de natura glicoproteinica· lamina densa - alcatuita din colagen V· lamina fibroreticularis - alcatuita din f. reticulina, colagen III si GAG.
· o structura cu aspect fibrilar care ocupa restul matricei extracelulare. Este alcatuita din colagen III (reticulina) predominant si colagen I. De asemenea exista uneori f. elastice abundente (tunica medie a vaselor mari, aorta). In afara de fibre, peste tot exista proteoglicani.
78

Spre deosebire de m. striat (scheletic si cardiac), m. neted are multa matrice extracelulara. Ea ocupa 50% din voulmul celular fata de 15% cat ocupa la muschiul striat. M. beted cu cea mai multa matrice extracelulara este m. neted vascular.
Matricea celulara este secretata si sintetizata de celulele musculare netede. Exista doua tipuri de cel. musc. netede: pentru contractie sau pentru secretia matricei extracelulare. Prin matricea extracelulara vin spre celule terminatii nervoase simpatice si parasimpatice in functie de muschi. Aceste terminatii sunt la o oarecare distanta de sarcolema fibrelor musculare netede, deci nu se formeaza sinapse. Astfel mediatorul chimic difuzeaza in spatiul intercelular.
Structura celulei musculare netedeSarcolemaAre aceeasi structura ca si membrana celulara obisnuita cu mici particularitati: jonctiunile
celulare - de tip gap (nexus), adherens (fascia adherens) sau digitiforme.Jonctiunea gap realizeaza comunicarea directa intre celule prin canale cu diametrul de
1500..., ceea ce permite trecerea ionilor de Ca, acesta fiind implicat in generarea potentialului de actiune (PA) si in mecanismul de contractie-excitatie.
Fibra musculara neteda are automatism (genereaza impulsuri). Exista zone pacemaker care genereaza PA ce se raspandesc in masa cu ajutorului jonctiunilor gap ce permit raspandirea instantanee a Ca in celule. Are loc o cuplare electrotonica. Exista si o cuplare metabolica.
Fasciile adherens si jonctiunile digitiforme solidarizeaza mecanic celulele. Astfel are loc sumarea contractiilor celulelor alaturate care se manifesta ca o contractie uniforma.
La nivelul sarcolemei exista formatiuni specifice, in cantitate crescuta. Acestea sunt caveolele si densificariile (ariile dense).
Caveolele reprezinta niste invaginatii ale membranei. A nu se confunda cu alte invaginari. Adancimea este de 120 nm, latimea maxima este de 70 nm iar deschiderea este de 35 nm. Ele sunt cvasipermanente, stabilitatea lor fiind data de citoschelet. Ele reprezinta prelungiri ale spatiului extracelular spre interiorul celulei. Cu toate acestea mediul din interiorul caveolei este diferit fata de ccel din spatiul extracelular, mai ales in ceea ce privesta Ca, concentratia sa fiind de 2-3 ori mai concentrat decat in mediul extracelular. In concluzie, caveolele participa la traficul ionilor de Ca, ceea ce le aseamana cu tubii T.
Ariile dense (densificarile membranei) au dimensiuni de 100-400 nm. Atat caveolele cat si ariile dense sunt asezate alternativ pe meridianele acestui fus. Compozitia chimica a ariilor este asigurata de 2 proteine (vinculina si talina) care apartin clasei de proteine de legare a actinei (ABP - actine binding protein). Ele sunt proteine de ancorare a actinei la mambrana celulara, realizand legatura intre actina si citoscheletul membranie celulare.
Pe preparatele de ME se observa ca actina se insera in unghi ascutit la nivelul ariilor dense.
SarcoplasmaCuprinde doua regiuni distincte: una situata central (mica d.p.d.v. al volumului) care contine
nucleul si unele organite comune si o regiune periferica, mai intinsa. Regiunea periferica contine aparatul contractil care contine organite asemanatoare celor din m. striat dar cu dispozitie diferita, fapt care indreptateste denumirea lor ca organite specifice. Acest aparat contractil contine mai multe elemente:
· filamente contractile: de actina (subtiri) sau de miozina (groase). · filamente intermediare, specifice acesti muschi, alcatuite din proteina numita desmina.· filamente de vimentina (in m. netezi vasculari) care sunt echivalente cu filamentele din
fibroblasti· filamente intermediare de simedina (in m. netezi viscerali)
Alte componente ale sarcoplasmei:· microtubulii - componenta mica
79

· proteine asociate actinei. Acestea sunt multe si unele au expresie morfologica sub forma corpilor densi (aglomerari de proteine). Corpii densi sunt raspanditi in toata citoplasma, au dimensiuni de sute de nm si sunt alcatuiti din doua proteine de legare a actinei: a-actinina si filamina. Ele au un rol special: solidarizeaza capetele filamentelor de actina prin punti transversale pe care aceste proteine le formeaza.
· RE este sursa de eliberare a ionilor de Ca pentru a creste concentratia de Ca in momentul contractiei m. neted. El reprezinta o alta sursa de intrare a Ca in citosol in afara de spatiul extracelular. Actina se insera cu un capat pe ariile dense iar celalalt capat este liber. Uneori acest capat
liber se insera pe corpii densi. De asemenea. filamentele de actina se pot insera cu un capat pe corpii densi, celalalt ramanand liber. Mecanismul contractiei consta in glisarea fibrelor de miozina printre cele de actina. “Motorul” contractiei este miozina. In contractie au loc scurtari ale fibrelor in directii diferite si ca urmare areloc o scurtare a celulei care este asociata si cu o scurtare a membranei celulare. La ME in contractie se observa scurtarea foarte mare a fibrei musculare. La ME de baleiaj, pe sectiune transversala se vad puncte ce reprezinta filamentele (citoplasma este plina de filamente). Recunoasterea filamentelor pe sectiune se face pe baza diametrului pe sectiune transversala:
· filamentele de actina au 6-8 nm· filamentele de miozina au 12-15 nm· filamentele intermediare au 11 nm
De asemenea apar densificari din loc in loc (corpii densi). Pentru reconstructia aparatului contractil s-ar folosi sectiuni seriate din aceeasi celula pentru ca m.neted este dificil de preparat pentru ME. Nu s-a reusit inca stabilirea unui model convenabil.
Structura moleculara a filamentelor contractileMolecula de miozina este asemanatoare cu miozina muschiului striat (este izoenzima cu
miozina din m. striat). Cele doua proteine su aceeasi structura secundara, structura tertiara, aceeasi morfologie, aceeasi functie, difera insa foarte putin la nivelul structurii primare, avand 2-3 aminoacizi diferiti. Aceste diferente minime determina consecinte importante d.p.d.v. functional. Aceasta enzima este formata din 2 lanturi: unul greu (H) si unul usor (L). Cele doua lanturi se infasoara unul in jurul celuilalt, apoi se despart avand o structura complicata si formand capul miozinei. Molecula are situsuri importante:
· situsul de interactiune cu actina· situsuri reglatoare care se gasesc pe lantul usor ce trebuie fosforilat de kinaza· situsul ATPazic ce elibereaza energie pentru contractie
Deosebirile intre cele doua tipuri de miozina apar la lanturile usoare (vezi situs reglator si ATPazic). Astfel, la m. neted situsul reglator trebuie fosforilat pentru producera contractiei, spre deosebire de m. striat unde contractia nu depinde in mod obligatoriu de fosforilare. De asemenea, situsul ATPazic la m. neted are afinitate mult mai mica pentru ATP decat situsul de la m. striat si deci hidroliza ATPului e mai inceata si se elibereaza energie mai incet si deci contractia este mai lenta.
De-a lungul portiunii liniare a miozinei exista situsuri de proteoliza limitata in care proteaza taie molecula in fragmente mai mici. Astfel, sub actiunea tripsinei, molecula de miozina este scindata in meromiozina usoara (LMM) si meromiozina grea (HMM). Acest situs are o mobilitate crescuta (este zona de balama) unde molecula de miozina se deformeaza. Meromiozina grea este legata la actina. Meromiozina grea basculeaza in procesul contractiei, capul miozinei fiind legat de actina. Bascularea meromiozinei grele trage de actina.
In contractie creste concentratia de Ca (10-7 M - 10-6 M). Efectul este activarea proteinei calmodulina. Calmodulina prezinta 4 situsuri de legare a Ca si este activata atunci cand toate situsurile sunt ocupate. Calmodulina activata actioneaza asupra kinazei lantului usor (MLCK)
80

care prin fosforilare se declanseaza activitatea ATPazica a lantului usor, producandu-se energie. Energia este folosita pentru deformarea miozinei in cadrul contractiei.
Filamentele de miozina sunt formate din multe molecule de miozina. M. striat nu are punti transversale pe toata lungimea sa (in portiunea benzii H nu are punti transversale) in timp ce m. neted are punti transversale pe toata lungimea filamentului gros. Lungimea filamentului gros la m. neted este de 3 ori mai mare decat la m. striat. De aceea interactiunea actina-miozina este pe o distanta mai mare si prin urmare m. neted se scurteaza mai mult.
In m. striat, unui filament gros ii corespund 2 fialmente subtiri. In m. neted, unui fialment gros ii coerspund 8-15 filamente subtiri, de aceea o punte transversala in m. neted trage mai multe filamente de actina si deci forta de contractie este mai mica.
Actina - la baza filamentelor sta actina F care este alcatuita din doua lanturi proteice filamentoase. Fiecare dintre cele doua lanturi este alcatuit din mai multi monomeri de actina G. Aceste lanturi de actina sunt stabilizate de tropomiozina care are forma de bastonas ce subintinde 7 monomeri de actina. Moleculele tropomiozinei sunt situate in santul dintre cele doua lanturi ale actinei F. Tropomiozina mascheaza situsul de interactiune intre actina si miozina in relaxare. La acest nivel exista diferente fata de m. striat. La m. striat, starea tropomiozinei este controlata de troponina care prin subunitatea C leaga Ca si se activeaza modificand si conformatia tropomiozinei. La m. neted rolul troponinei C este prelluat de catre calmodulina care este o proteina solubila in citosol. Prin legarea a 4 ioni de Ca ea se activeaza si actioneaza asupra caldesmonei care la randul ei actioneaza asupra tropomiozinei si determinand modificarea ei conformationala, ce are ca efect expunerea situsului de interactiune actina-miozina. Exista 3 tipuri de actina, in functie de tipul de m. neted.
Histofiziologia muschiului netedMecanismul contractieiConsta in alunecarea filamentelor de actina in raport cu cele de miozina datorita puntilor
transversale. Are loc o dubla reglare a contractiei: · una actioneaza asupra filamentelor groase prin MLCK si se produce fosforilarea acestui lant care
are drept consecinta declansarea activitatii situsului ATPazic· alta actioneaza asupra tropomiozinei prin CD (caldesmona) si astfel se modifica conformatia
tropomiozinei cu expunerea situsului de interactiune actina-miozina.De asememnea un rol important in mecanismul de contractie il au proteinele de legare a
actinei (ABP) care controleaza polimerizarea si pozitionarea moleculelor de actina.
Particularitatile contractiei· forta de contractie e scazuta (datorita nr. mic de punti transversale)· viteza de contractie e scazuta (datorita activitatii ATPazice scazute)· capacitatea de scurtare e crescuta (datorita existentei puntilor transversale pe toata lungimea
filamentului gros)
Particularitati functionale· Activitatea contractila ritmica spontana - au loc contractii cu frecventa si amplitudine constanta
dependenta de muschi. - acestia fiind m. fazici. Exista insa si m. tonici care au un tonus spontan, o stare de contractie sustinuta, tradusa printr-o miograma in platou
· Contractia e modulata de urmatorii liganzi, prin legare la receptorii de pe membrana celulei musculare netede:
· neurotransmitatori· hormoni
81

· prostaglandine· Prezinta dubla inervatie vegetativa, excitatorie si inhibitorie; sinapsele sunt absente. Acetilcolina
poate influenta procesul contractil la nivelul placii motorii. Aceste molecule (toti liganzii) actioneaza la nivelul receptorilor. Ei actiuni de tip elastic: intr-un loc poate fi inhibitor iar in alt loc poate fi stimulator al contractiei. Actiunea lor depinde de tipul de receptor exprimat in membrana celulara si de tipul de mecanism de transductie ce are loc in celula.
· Fibrele sunt cuplate electric prin jonctiuni gap si cuplate mecanic prin jonctiuni adherens. Aceste jonctiuni asigura functionalitatea tesutului.
Clasificarea functionala a muschilor netezi· Mm. netezi unitari (viscerali, vasculari) - au activitate ritmica spontana, initiata in arii
pacemaker. Se contracta ca o unitate. U multe jonctiuni gap care asigura raspandirea PA si au o inervatie slaba (o terminatie nervoasa inerveaza un fascicul de fibre: 100-300 fibre).
· Mm. netezi multiunitari (iris, mm. ciliari, mm. piloerectori, tunica medie a vaselor mari). Fiecare fibra se contracta independent. Au putine jonctiuni gap si nu au contractii spontane. de asememnea fiecare fibra este inervata de o terminatie nervoasa libera.
· Mm. netezi intermediari (vezicula seminala, canalul deferent) Nu au actiune spontana, au numar moderat de jonctiuni gap si o terminatie nervoasa libera inerveaza un fascicul mic de fibre.
Etapele desfasurarii procesului de contractie sunt:· semnalizarea transmembranara (prin intermediul complexului ligand-receptor). Se activeaza
sistemul fosfolipazei C care produce contractie sau se activeaza sistemul AMPc sau GMPc care duc la relaxare.
· cuplarea excitatiei cu contractia se face cu ajutorul ionilor de Ca· contractia depinde de:
· canalele de Ca din membrana RS care sunt comandate electric sau chimic· eliberarea Ca din RS· proteinele contractile actina si miozina
· relaxarea depinde de:· pompa de extruzie a Ca· pompele de ca din membrana RS care reintroduc Ca din citosol in RS· proteinele de contractie actina si miozina
PIP2 (fosfatidil inozitol bis fosfat) sub actiunea PLC (fosfolipaza C) se transforma in DAG (diacil glicerol) si IP3 (inozitol trifosfat). DAG determina activarea PKC (protein kinaza C).
Fenotipuri ale celulelor musculare netede adulte:· Starea contractila - celula este oprita in G1. Aceasta stare este mentinuta de inhibitori ai
proliferarii, celula avand receptori pentru acesti factori. Celula nu are receptori pentru factorii mitogeni (de stimulare a cresterii), ea este refractara la acestia. De asemenea, in acest stadiu celula raspunde la agentii care contracta. Spre exemplu, m. neted vascular raspunde la angiotensina II. In aceasta etapa celula nu sintetizeaza matrice extracelulara.
· Starea secretorie - celula iese din G1. Ea raspunde la factorii mitogeni (spre exemplu PDGF). Nu se contracta dar sintetizeaza matrice extracelulara (colagen, fibre elastice, proteoglicani).Aceste doua fenotipuri sunt foarte bine exprimate la m. neted vascular. Atfel se explica
ateroscleorza, boala a celulelor musculare netede, in care creste numarul celulelor secretoare. De-a lungul timpului s-au elaborat mai multe modele experimentale ale aterosclerozei.
Initial, dintr-un motiv oarecare, are loc lezarea endoteliului vascular, ceea ce determina expunerea matricei extracelulare aflate sub endoteliu la fluxul sangvin. Are loc aderarea plachetelor si se formeaza un trombus plachetar, ceea ce duce la eliberarea PDGF si a altor factori existenti in granule. Daca creste concentratia PDGF sunt stimulate celulele secretorii ale
82

m. neted. Acestea devin capabile sa secrete PDGF si concentratia acestuia creste si mai mult. Concentratia PDGF crescand din ce in ce mai mult are loc cooptarea celulelor din tipul contractil in cel secretor. Celulele secretorii migreaza in intima vasului lezat unde secreeta matrice extracelulara ce determina placa de aterom. La toate acestea se adauga si faptul ca celulele secretorii au si receptori pentru LDL (“carausii colesterolului”). Atunci LDL sunt internalizate in celula si acestea se blocheaza, ceea ce duce la crestera volumului placii de aterom.
Consecintele aterosclerozei pot fi :· accindet vascular cerebral· infarct miocardic· arterita obliteranta membranara
Tratamentul aterosclerozei este paleativ.
TESUTUL NERVOS
Sistemul nervos clasic este impartit in SNC (sistemul nervos central) si SNP (sistemul nervos periferic). Sistemul nervos este alcatuit din organele nervoase:
· SNC:· maduva spinarii · creier alcatuit din:
· trunchi cerebral· cerebel· diencefal· scoarta cerebrala
· SNP:· ganglioni:
· senzitivi· spinali· cranieni
· vegetativi· intramurali· laterovertebrali· prevertebrali
· nervi· senzitivi· motori
83

Aceste organe sunt alcatuite din tesut nervos la care se adauga capilare sangvine (foarte multe). De asemenea se mai adauga paratul auxiliar (de protectie), de facilitare a functiilor organului.
SNC (creierul si maduva spinarii) este invelit de meninge care mediaza contactul cu cavitatea osoasa. Meninigele este alcatuit din duramater, piamater si arahnoida (de la exterior spre interior). De asemenea in SNC mai exista sistemul de apeducte, canalul ependimar. Sistemul de cavitati si apeducte comunica cu spatiul de sub arahnoida si este plin cu LCR (lichid cefalorahidian).
La SNP aparatul de sustinere este format de tecile nervoase:· epinerv· perinerv· endonerv
In categoria organelor auxiliare sunt incluse si aparatele receptoare asociate fibrelor nervoase. Acestea sunt alcatuite din tesut epitelial senzorial. Deci organele auxiliare sunt formate din:
· tesut nervos (sistemele canaliculare)· tesut conjunctiv (tecile)· tesut epitelial (organele receptoare)· vase sangvine
Teutul nervos este alcatuit din celule si matrice extracelulara aflata in cantitate mica. Celulele sunt de doua tipuri: neuroni si celule gliale.
Neuronii genereaza o forma de energie numita impuls nervos si comunica unii cu altii, impulsul nervos circuland de la un neuron la altul. Dupa functie, neuronii se impart in:
· senzitivi· motori· integratori (mediaza relatia intre neuronii senzitivi si motori)
La vertebrate si om nr. de neuroni integratori este de aprox. 99% din totalul de neuroni din organism.
Celulele gliale sunt de mai multe feluri:· in SNC se numesc nevroglii (neuroglii):
· astrocite· oligodendrocite· microglii· celule ependimare
· in SNP:· celulele Schwann· celulele satelite
Alcatuirea celulei nervoaseNeuronul este alcatuit din corp celular (pericarion) si prelungiri (dendrite si axon).
Pericarionul si dendritele au rol de receptie a impulsului nervos de la alt neuron prin sinapse si rol de sumatie a informatiilor primite de la alti neuroni.
Neuronul are potentialul mambranar de repaus de -90 - -70 mV. Impulsul poate fi de excitatie ce are ca urmare cresterea PR (potentialul membranar de repaus) = depolarizare sau poate fi de inhibitie si are ca rezultat scaderea PR = hiperpolarizare.
Integrarea reprezinta sumarea acestor informatii ce vin la celula si determina un raspuns final. Zona ce raspunde la impuls este cea cu PR crescut (se atinge mai usor pragul de excitatie). Scaderea PR pana la valoarea prag determina declansarea PA care se produce in axon, in partea initiala a acestuia (zona conului axonal urmat de segmentul initial). In zona conului axonal exista
84

canale ionice de Na cu cel mai mic prag de excitabilitate. Declansarea raspunsului este asociata si cu deschiderea canalelor de Ca.
Axonul este zona declansatoare a PA. De asemenea el are o portiune terminala prin care transmite impulsul prin intermediul sinapsei. Axonul are si o zona de conducere a impulsului nervos care poate fi mielinizata sau nu. Mielina creste viteza de transmitere a fluxului nervos.
Celulele de sustinere (nevrogliile) au rol ajutator, auxiliar (au rolul “valetului”). Ele mentin homeostazia mediului si reprezinta complementul structural pe toata lungimea neuronului. Tipurile de neuroglii sunt: astrocite, oligodendrocite, microglii, celule ependimare. Acestea, impreuna cu celulele Schwann au urmatoarele roluri:
· mediaza raportul dintre neuron si vasele de sange (astrocitele)· elaboreaza teaca de mielina in SNC (oligodendrocitele)· macrofage (microgliile)· secreta lichid cerebrospinal (LCR secretat de celulele ependimare)· formeaza teaca de mielina in SNP (celulele Schwann)
Proprietatile tesutului nervos:1. automatism endogen2. excitabilitate3. conductibilitate4. comunicare
Automatismul endogenReprezinta generarea de impulsuri nervoase in interiorul sistemului nervos insusi, in absenta
oricarui stimul extern. Acest fenomen apare atat in interiorul SN in totalitate cat si la nivelul fiecarui neuron. Nu se cunoaste mecanismul acestui proces care mai este prezent si la celulele musculare netede, celulele tesutului excitoconductor nodal sau celulele adenohipofizei. Astfel, prin izolarea indivizilor in intuneric, liniste, cu hrana putina, s-a observat o scadere a stimularii receptorilor dar functionarea SN este normala, singura alterare este cea legata de perceperea timpului.
Excitabilitatea si conductibilitateaReprezinta capacitatea de a genera impulsuri nervoase atunci cand se aplica un stimul si
respectiv transmiterea impulsului de-a lungul celulei.Stimulul actioneaza asupra receptorului care transmite impulsul prin intermediul nn. senzitivi
periferici care ajunge in SNC unde are loc integrarea si declansarea senzatiei sau se da comanda motorie care ajunge prin nn. motori periferici la organul efector. Exceptie de la aceasta regula fac reflexele viscerale.
ComunicareaReprezinta transmiterea impulsului nervos de la un neuron la altul. Se face prin sinapse sau prin contiguitate (continuitate - jonctiuni interneuronale). Sinapselel
pot fi electrice (gap) sau chimice. Transmiterea in sinapsele chimice se face prin neurotransmitatori sau neuromodulatori (moduleaza activitatea sinapsei)
Comunicarea se mai poate face si la distanta, pe cale endocrina, prin neurohormoni. Unele categorii de neuroni pot elibera substante in curentul sangvin (neurohormoni).
Traseul impulsului nervos este urmatorul:· declansarea impulsului nervos se face la nivelul conului axonal si a portiunii initiale a axonului· conducera impulsului nervos se face in segmentele invelite sau nu de mielina· partea terminala a axonului (partea ramificata - telodendronul) participa la sinapse.
Exista o polarizare dinamica a neuronului: impulsul nervos circula doar de la dendrita si pericarion catre axon. Aceasta specializare se datoreaza membranei neuronale.
Clasificarea neuronilor se poate face dupa mai multe criterii:1. dupa numarul de prelungiri:
85

· neuroni unipolari: la adult nu exista, acestia existand doar la nevertebrate si in viata intrauterina. In viata I.U. neuronii migreaza dintr-un loc in altul iar axonul este indreptat spre directia miscarii si e folosit la deplasare
· neuroni bipolari - foarte rari, de natura senzitiva:· in retina· neuronii olfactivi· neuronii cu sediul in ggl. acusticovestibulari VIII
· neuroni pseudounipolari - au o singura prelungire la adult dar in viata I.U. au fost neuroni bipolari. Ulterior aceste doua prelungiri se apropie una de alta si se unesc formand un trunchi comun care se ramifica apoi in “T”. Aceasta prelungire in forma de T este axon dar culege si informatii. Astfel sunt neuronii din ggl, spinali senzitivi, situati pe radacina posterioara a n. spinal.
· neuroni multipolari - sunt foarte complicati. Exista numeroase forme de neuroni multipolari. Ei prezinta un corp stelat. De acest tip sunt:· neuronii din coarnele anterioare ale maduvei spinarii· neuronii din scoarta cerebrala care sunt:
· stelati in scoarta cerebrala· fusiformi in stratul polimorf· piramidali in stratul piramidal
· celulele Purkinje in scarta cerebeloasa, cu forma de para, dendritele fiind foarte ramificate, in forma de “corn de cerb”
· celulele mitrale din bulbul olfactiv· exista si celule multipolare fara axon. Prelungirile acestor celule sunt dendrite dar care
pot functiona si ca axon. Exemple:· celulele amacrine din retina (celule de asociatie· celulele granulare din bulbul olfactiv
2. Dupa forma axonului:· celule cu cosulet - axonul este scurt si prezinta multe ramificatii ce infasoara corpul celulei ca un
cos. Acestea sunt celulele Purkinje din scoarta cerebeloasa· celulele in panza de paianjen - in scoarta cerbrala· celulele in dublu buchet - in scoarta cerebrala
3. Dupa lungimea axonului:· neuroni de proiectie, cu axonul foarte lung. Pot fi motorii sau senzitivi. Sunt Golgi tip I.
Realizeaza proiectia pe scoarta cerebrala a tuturor regiunilor corpului.· neuroni de asociatie (Golgi tip II) - neuroni cu axonul scurt, de tip integrator
4. Dupa invelisul axonului:· neuroni mielinici - au viteza mare de conducere, proportionala cu grosimea tecii si grosimea
fibrei· neuroni amielinici - au viteza mica de conducere
Numarul neuronilor este de ordinul 109. Numarul de neuroni senzitivi si motori e constant filogenetic. Numarul de neuroni integratori creste pe scara filogenetica si este maxim la om.
Marimea celulei nervoase este de 3-5 mm (minim - la celulele granulare din stratul granular al scoartei cerebeloase) pana la 100-120 mm (maxim la celulele piramidale gigantice Betz din cortexul motor, girul precentral). Aceste celule Betz pot avea axonul lung de pana la 1 m si un diametru ce variaza intre 20-40 mm. Exista o relatie de proportionalitate intre marimea corpului neuronal si lungimea axonului deoarece proteinele axonale sunt sintetizate de catre corpul celular, axonul neavand RER. Deci corpul intretine acest axon.
Soma (pericarionul) are rol trofic (intretine axonul), de receptie si de integrare a semnalelor. El contine nucleul care este mare, eucromatic, cu nucleol evident datorita sintezei crescute de proteine. Exista insa si exceptii - neuroni cu nucleu mic tahicromatic. Nucleul este central sau
86

periferic. Fiind eucromaatic, la femei se vede corpusculul Barr atasat de nucleol sau membrana nucleara.
In citoplasma contine:· corpi Nissl care se pot observa cu coloratie speciala la baza dendritelor si lipsind in conul axonal
si fiind alcatuit din RER si poliribozomi liberi· REN foarte abundent, cu rol in eliberarea Ca pentru cresterea concentratiei sale citosolice,
fenomen ce participa la transmiterea impulsului nervos.· Aparat Golgi - foarte bine dezvoltat· Vezicule sinaptice, care se formeaza la acest nivel pentru ca mai apoi sa migreze pana la
terminatia axonala. Aceste sunt putine. In neuronii vegetativi simpatici ele sunt umplute cu catecolamine si dense la fluxul de electroni. Au diametrul de 60-80 nm.
· Vezicule de neurosecretie - contin peptide si neurofizine care sunt eliberate in capilare. Ele au dimensiuni de 100-300 nm, se gasesc in hipotalamus si sunt dense la fluxul de electroni. Complexele formate din peptide si neurofizine este depozitat in neurohipofiza de unde este eliberat in circulatie.
· Mitocondrii· Rezerve de picaturi lipidice· Lizozomi - de-a lungul timpului acestia se incarca cu un material - lipofuscina - pigment cu
semnificatie fiziologica necunoscuta ce se acumuleaza cu varsta.· Melanina (la unii neuroni - neuronii din substanta neagra din diencefal - locus coeruleus)· Centrioli - prezenti in toti neuronii, avand rol in organizarea fusului nuclear si a microtubulilor.· 1 cil cu semnificatie necunoscuta
Citoscheletul este alcatuit din:· microtubuli, cu diametrul de 25 nm. Microtubulii sunt complexe alcatuite din tubuli cu diametrul
amintit impreuna cu MAP (proteine asociate microtubulilor) si proteinele motorii care mediaza interactiunea microtubulilor cu organitele celulare care se deplaseaza de-a lungul microtubulilor.
· neurofilamente, cu diametrul 10 nm, fiind filamente intermediare. Ele au un rol neclarificat si sunt alcatuite din citokeratine specifice care sunt implicate in boala Alzheimer. In mod normal neurofilamentele sunt asezate paralel. La examenul anatomopatologic al unui bolnav Alzheimer s-a constatat dispozitia anarhica a acestora - incalcite, rupte, ondulate. Boala Alzheimer (dementa senila) este o tulburare neurologica generalizata.
· microfilamente cu diametrul de 6 nm, alcatuite din actina F. Actina F este ancorata la membrana neuronala prin fodrina care este analoaga spectrinei din hematii.Conform teoriei actuale o parte din neuroni (nu toti) se divid. Neuronii sunt celule stationate
in faza G1 (postmitotica). Exista zone importante din sistemul nervos in care are loc o reinnoire a neuronilor:
· regiunea paraventriculara (ventriculul III)· neuronii din bulbul olfactiv· neuronii din mucoasa olfactiva
Acest proces de reinnoire se numeste neuropoieza si are numeroase similitudini cu hematopoieza.
Dendritele reprezinta prelungiri ale somei care contin corpi Nissl si toate organitele celulare, abundente la baza dendritei si din ce in ce mai putine la varf, unde exista mai multe elemente de citoschelet. Contin multi microtubuli si neurofilamente. Transportul dendritic poate fi anterograd (se face cu viteza de 3 mm/h si se transporta proteine si organite, ex: mitocondria) si retrograd.
Principala functie a denritelor este receptia. Dendritele prezinta specializari cvasipermanente - muguri (spini) dedritici care primesc sinapsele. Acestia au un material dens la fluxul de electroni, prezinta microtubuli si aparatul spinului, alcatuit din cisterne paralele unite prin
87

material electronodens. Numarul spinilor de pe ficare dendrita variaza. Daca se taie aferentele numarul spinilor scade.
Axonul este unic. El are mai multe parti:· con si segment initial. Aici lipsesc corpusculii Nissl, insa prezinta numerosi microtubuli,
microfilamente si neurofilamente care se vad cu impregnare argentica. Rolul acestei regiuni este de a declansa impulsul nervos deoarece este zona cu cel mai mic prag de excitabilitate.
· segmentul de conducere care poate fi mielinic sau amielinic· terminatiile axonale formate din butonii terminali (sinaptici, de pasaj)
Transportul axonal poate fi rapid sau lent.· transportul rapid se poate face anterograd (cu viteza 100-400 mm/zi) sau retrograd (100-200
mm/zi). Se transporta organitele cu endomembrane (mitocondrii), vezicule cu mediatori si componentele membranei. Transportul se face de-a lungul microtubulilor cu ajutorul proteinelor motorii care sunt de doua feluri: kinezina, efectueaza transportul anterograd si syneina care efectueaza transportul retrograd. Aceste proteine mediaza interactiunea microtubulului cu organitele. De asemenea exista si o reciclare locala, rapida a veziculelor cu mediatori.
· transportul lent se face numai anterograd cu viteza de 1-3 mm/zi, avand un mecanism necunoscut. Se transporta proteine solubile (enzime, monomeri)
Fibrele nervoaseSunt formate din axoni si din microglii.
· fibrele nervoase periferice - formeaza nervii care reprezinta fascicule de fibre nervoase. Fibrele nervoase pot fi amielinice (inconjurate doar de celule Scwwann) si fibre mielinice (inconjurate de celule Schwann ce au format taeca de mielina). O celula Schwann poate acoperi mai multi axoni saau un singur axon.
· fibrele nervoase centrale pot fi amielinice (“libere” in componenta neuropilului) sau mielinice, invelite de prelungiri ale oligodendrogliilor care formeaza teaca de mielina. Acestea intra in componenta substantei albe. Teaca de mielina se observa cu impregnare argentica. Ea se formeaza prin succesiunea
celulelor Schwann. Intre doua noduri Ranvier, se afla o celula Schwann care acopera cu citoplasma ei axonul. De asemenea, in teaca de mielina exista din loc in loc linii clare numite incizuri Schmidt-Landermann.
Formarea tecii de mielinaInitial fibra este amielinica iar celula Schwann formeaza doua prelungiri in jurul fibrei, in
sensuri opuse (formeaza un mezoaxon sau mezaxon). Aceste prelungiri cresc in mod spiralat in jurul axonului. Straturile succesive ale membranei celulei Schwann fuzioneaza intre ele si deci teaca de mielina reprezinta suprapuneri ale membranei celulei Schwann. La exterior are mezaxon extern si fibrele se rotesc ajungand la mezaxon intern.
Clasificarea fibrelor nervoase periferice dupa viteza de conducereDaca diametrul axonului este mare, teaca de mielina este groasa, portiunea internodala este mai lunga si astfel viteza de conducere este mai mare deoarece in fibrele mielinice, transmiterea (depolarizarea) are loc doar la nivelul nodului Ranvier (transmiterea este saltatorie).
Tipul fibrei Viteza de conducere Functia acestor fibremielinice A 15-120 m/s fibrele motorii scheletice
fibrele somatosenzitive proprioceptive (Ia)fibrele somatosenzitive tactile (Ib)fibrele somatosenzitive termice (II)fibrele somatosenzitive pentru durerea rapida (III)
88

mielinice B 3-15 m/s fibre viscerosenzitiveAmielinice C (2/3 din
fibrele periferice)0,5-2 m/s fibre somatosenzitive pentru durerea lenta (IV)
fibre visceromotoriiIn substanta alba (in SNC) fibrelele mielinice sunt determinate de oligodendroglie.
Internodurile sunt formate de mai multe oligodendroglii.Varstele mielinizarii
· la nastere: sunt mielinizate doar radacinile motorii ale nn. spinali, ceea ce explica miscarile necoordonate ale copiilor.
· la 3-4 luni se mielinizeaza si nn. optici si radacinile senzitive ale nn. spinali, deci la aceasta varsta copiii vad bine si au senzatii precise.
· la 1 an se mielinizeaza tracturile corticospinale deci copii merg bine.· la 7 ani se mielinizzeaza si axonii fibrelor comisurale (de asociatie) ale emisferelor cerebrale ce
apartin corpului calos.Functiile mielinice:
· creste viteza de conducere a impulsului nervos de la 1 m/s la 120 m/s deoarece la nivelul internodului este o zona cu rezistenta mare si capacitanta mica. Aceasta se datoreaza faptului ca la acest nivel membrana axonala are o compozitie moleculara diferita. La nivelul internodului canalele ionice sunt rare, acestea fiind adunate la nivelul nodurilor Ranvier.
· asigura nutritia axonului deoarece mediaza raporturile cu exteriorul. De asemenea, celulele Scwann au si rol in regenerara axonilor dupa traumatisme.
Sinapsa
Este o jonctiune intercelulara specifica neuronilor. Prin intermediul sinapsei se faciliteaza eliberarea neurotransmitatorilor care asigura transferul de informatie de la o celula la alta. Neurotransmitatorul influenteaza conductanta celulei receptoare actionand asupra canalelor ionice si schimba potentialul membranar al celulei receptoare.
Structura sinapsei:· element presinaptic (axon)· element postsinaptic care poate fi dendrita, soma sau axon· spatiu sinaptic
Butonul presinaptic contine: REN, mitocondrii, vezicule sinaptice, microfilamente, microtubuli, neurofilamente. Spatiul sinaptic are 20-30 nm. Elementul postsinaptic este un spin dendritic care contine microfilamente, elemente de citoschelet, aparatul spinului (optional).
Membrana postsinaptica are pe versantul intern o depunere de material electronodens ce trimite prelungiri ininterior. La fel se intampla si la membrana presinaptica. Dupa natura materialului electronodens s-a facut clasificarea sinapselor de catre Gray:
· sinapse asimetrice (materialul electronodens presinaptic este mai mare decat cel postsinaptic)· sinapse simetrice (materalele electronodense au dimensiuni aproximativ egale)
Aceasta clasificare este legata de aspecte functionale.Veziculele sinaptice pot fi si ele clasificate dupa cum urmeaza:
· vezicule clare, sferice - au diametrul de 40-60 nm; frecvent au neurotransmitatori excitatori, uneori si inhibitori; ex.: veziculele de acetilcolina
· vezicule clare, turtite - au diametrul de 20-60 nm; contin doar neurotransmitatori inhibitori· vezicule sferice cu miez dens - au diametrul de 40-60 nm; contin catecolamine· vezicule mari, sferice cu miez dens si halou clar - au diametrul de 80-100 nm. Aceste sinapse se
gasesc in corpul si butonul axonal in SNC si au un continut necunoscut.
89

Clasificarea morfofunctionala a sinapselor:· excitatorii - sunt asimetrice, au vezicule clare, sferice, efectul pe membrana postsinaptica este de
depolarizare (cresterea PR) - se declanseaza un potential postsinaptic excitator (EPSP)· inhibitorii - sunt simetrice, au vezicule clare sferice sau turtite, efectul fiind de hiperpolarizare
(scaderea PR) - se declanseaza un potential postsinaptic inhibitor (IPSP).Unul si acelasi neurotransmitator poate avea efecte diferite in functie de receptori (tipul de
receptor determina efectul).Neurotransmitatorii Isi exercita efectul pe membrana postsinaptica in timp scurt (de ordinul ms). Clasificarea lor
dupa efectele realizate in majoritatea cazurilor:· neurotransmitatori inhibitori:
· GABA (acid g-amino-butiric) - folosit ca inhibitor de celulele Purkinje din cerebel· glicina - are efect pe celulele din maduva spinarii· purine (adenozina, AMP, ADP, ATP)· NO (monoxid de azot)
· neurotransmitatori excitatori· glutamat (actioneaza in SNC)· aspartat (actioneaza in SNC)· taurina (actioneaza in SNC)
· neurotransmitatori cu efecte inhibitorii sau excitatorii in functie de receptor:· acetilcolina· noradrenalina· serotonina· dopaminaNeuromodulatoriiIsi exercita efectul un timp mai indelungat, au durata mai mare a efectului, latenta este mai
mare, se cumuleaza in timp si pot avea efecte ce dureaza zile, ani. Orice substanta poate actiona ca neurotransmitator, neuromodulator sau neurohormon
(uneori).Neuromodulatorii sunt de obicei peptide comune cu sistemul APUD (sistemul neuroendocrin
difuz). Ei au efecte atat pe membrana presinaptica cat si pe membrana postsinaptica:· pe membrana presinaptica au efecte de accelerare sau incetinire a eliberarii veziculelor· pe membrana postsinaptica influenteaza legarea mediatorilor de receptori
NeurohormoniiSunt peptide secretate in hipotalamus care sunt eliberati si apoi ajung in neurohipofiza (in
lobul posterior sau in emineta mediana).Clasificara topografica a sinapselorTipul de sinapsa depinde de partile neuronului ce participa la sinapsa:
· tipurile clasice de sinapse (cele mai frecvente):· axospinoase (sinapsa se face cu spinul dendritic) - sunt numai excitatorii· axodendritice - sunt excitatoare sau inhibitoare· axosomatice - sunt excitatoare sau inhibitoare
axoaxonale - sunt numai inhibitorii; din axon ca element postsinaptic este exclusa partea mielinizata (conducatoare) deci participa la formarea sinapsei axoaxonale doar conul cu segmentul initial si butonii terminali.
· tipurile noi de sinapse (de regula inhibitorii):· dendrodendritice· dendrosomatice· dendroaxonale · soamtodendritice
90

· somatosomatice· somatoaxonale
· speciale (la ele participa concomitent mai multe prelungiri):· reciproce - sunt dendrodendritice; reprezinta un complex de 2 sinapse aflate una langa
alta si care conduc impulsul nervos in sens invers· panglica (in retina) - este axodendritica; participa 1 axon si 2 dendrite foarte apropiate
intre ele; axonul face sinapsa cu ele (1 sinapsa cu 1 dendrita). Axonul prezinta pe linie mediana o densificare in forma de panglica
· seriala - este axoaxonica; reprezinta o insiruire de sinapse axoaxonice , axonii facand sinapsa unul cu altul
Clasificarea sinapselor dupa mecanismul de transmitere· sinapse chimice (cu neurotransmitatori)· sinapse electrice (prin jonctiuni gap) - foarte putine in SN· sinapse mixte (cuplaj intre sinapsa electrica si cea chimica). Sunt rare. Ex.: nucleii vestibulari
laterali, nucleii tractului mezencefalic al trigemenului.O celula Purkinje primeste aprox. 300.000 de sinapse
Principii de functionare a sinapselorUn neuron nu foloseste un singur neurotransmitator, intotdeauna exista mai multe substante
neuroactive. Aceste substante exercita efecte diferite in situatii diferite, efecte mediate de tipul receptorilor de care se leaga.
Un exemplu este dopamina care coexista in neuronii trunchiului cerebral cu peptide precum colecistokinina, neurolentina. Ea functioneaza ca:
· neurotransmitator cand este eliberata de neuroni din substanta neagra· neurohormon cand este eliberata de neuronii hipotalamici din nucleul arcuat din adenohipofiza
(inhiba secretia de prolactina este PIF - prolactin inhibiting factor)Ea excita, inhiba sau moduleaza activitatea celulei tinta, in functie de natura receptorului din
membrana postsinaptica.Substante neuroactive
· gaze: NO (oxid nitric), CO· monoamine:
· catecolamine: adrenalina, noradrenalina, dopamina· amine din triptofan: serotonina, triptamina· aspartat· glutamat· GABA· glicina· taurina
· peptide:· actioneaza ca neuromodulatori: colecistokinina, endorfine, encefaline, neurotensina,
substanta P, VIP· actioneaza ca neurohormoni: LH-RH, TSH-RH, somatostatina, vasopresina
· purine: adenozina, AMP, ADP, ATPIn unele celule se formeaza NO sau CO. No se formeaza sub actiunea NO sintetazei, din
arginina iar CO se formeaza sub actiunea hemoxigenazei din hem. NO actioneaza ca neurotransmitator inhibitor, activeaza GMPc, activeaza pompa de extruzie a Ca, ceea ce duce la scaderea Ca intracelular (efect inhibitor). De asemenea NO mai este folosit in unele terminatii nervoase vegetative din m. netezi cum ar fi cei din corpii cavernosi (vezi erectia, vezi aplicarea
91

Viagra) dar si pe m. netezi vasculari. De aceea NO poate determina accidente vasculare majore. Injecarea NO in SNC determina cascatul.
La nivelul sinapselor are loc permanent reciclarea materialelor. veziculele sinaptice se formeaza la nivelul corpului celular dupa care sunt transportate pana la nivelul butonilor terminali. La aparitia impulsului nervos veziculele fuzioneaza cu membrana sinaptica si se elibereaza continutul in spatiul sinaptic. Fuzionarea veziculelor cu membrana presinaptica este limitata deoarece aria membranei presinaptice creste foarte foarte mult. Dupa eliberarea veziculelor apar vezicule de clatrina care contin proteine importate din RER, se formeza alte vezicule cu mediatori si ciclul se reia.
Aceeasi reciclare are loc si la nivelul mediatorului. Exemplu pentru sinapsa colinergica (cu acetilcolina). Materia prima este colina care se gaseste in cantitate crescuta in spatiul sinaptic. Ea patrunde prin membrana butonului sinaptic prin transportorul specific simultan cu Na (are loc un simport). In citoplasma, colina este acetilata luand radicalul acetil de la acetilcoenzima A prezenta in mitocondrii. Acetilarea se face in prezenta enzimei CAT (colin acetil transferaza). Acetilcolina formata nu stationeaza ci este preluata de un transportor existent in membrana veziculei sinaptice care o introduce in vezicula, scotand pe de alta parte H+. Se realizeaza astfel un antiport. Excesul de acetilcolina este hidrolizat de catre acetilcolinesteraza.
Vezicula sinaptica contine:acetilcolina, ATP si VIP care sunt asezate intr-un material bogat in PG. Transmiterea impulsului nervose datorata deschiderii canalelor de Na si a celor de Ca care sunt dependente de potential. Are loc cresterea concentratiei Ca in citoplasma. Introducerea Ca determina exocitoza veziculelor sinaptice (fuzionarea cu membrana presinaptica). Toate substantele sunt eliberate in fanta sinaptica si determina efectele.
Excesul de acetilcolina nelegata de receptori si aflat in spatiul sinaptic este hidrolizat de acetilcolinesteraza si se formeaza radicali acetil si colina, care va relua ciclul.
Substante ce blocheaza cu transmiterea sinaptica· Transportorul ce efectueaza simportul Na-colina poate fi blocat de unele medicamente ce sunt
analoage colinei (hemicolina). Efectul este de paralizie. · Exocitoza poate fi si ea blocata, spre exemplu de endotoxina botulinica ce provoaca paralizia,
inclusiv a m. respiratori.· Exista si inhibitori ai acetilcolinesterazei, ceea ce duce la acumularea acetilcolinei in spatiul
sinaptic si la blocarea receptorilor de catre aceasta. Aceste substante sunt folosite pentru tratamentul bolii miastenia gravis, determinata de modificari la nivelul placii motorii.Controlul exocitozei veziculelorExocitoza este declansata de cresterea Ca intracelular. Ca activeaza calmodulina, care se
transforma in kinaza si fosforileaza sinapsinele I si II. Sinapsinele sunt proteine de legare a actinei: au un capat pe actina si un capat pe veziculele de mediatori. In repaus veziculele se gasesc in asteptare. Cand sinapsina se desprinde este determinata deplasarea veziculelor catre membrana presinaptica. Interactiunea veziculelor cu membrana este mediata de doua proteine: sinaptotagmina si sinaptofizina.
Distributia componentelor celulelor nervoase si nevrogliilor in SNC si SNP
Componentele SNPSunt ganglionii si nervii.Ganglionii contin corpuri neuronale, dendrite, axoni mielinici sau amielinici. Unii ganglioni
contin sinapse, altii nu, cum este cazul ggl. senzitivi. GGl. senzitivi contin fibre nervoase mielinice iar ggl. vegetativi contin fibre preggl. mielinice si fibre poasggl. amielinice. In ggl. mai exista un tip de celule Schwann numite celule satelite care se gasesc in jurul pericarionului.
In nervi exista axoni inconjurati de celule Schwann.
92

Organizare / elemente celulare Ganglioni Nervineuroni exista -soma exista -
dendrite exista -axoni mielinizati / nemielinizati exista exista
celule Schwann exista exista
Celulele sateliteSunt celule Schwann ce se gasesc in ggl. spinali, cranieni sau vegetativi. Ele se gasesc in mai
multe locuri:· in ggl. spinali si cranieni - formeaza o capsula ce inconjoara complet pericarionul neuronilor
pseudounipolari, care nu primesc sinapse· in ggl. vegetativi - formeaza o capsula discontinua in jurul perikarionului neuronilor multipolari
care primesc sinapse.· tot in ggl. spinali si cranieni formeaza o teaca de mielina in jurul segmentului convolut al
axonului neuronului pseudounipolar.· in ggl. vestibulocohlear - formeaza teaca de mielina in jurul axonilor neuronilor bipolari
Constitutia SNCElemente celulare Substanta
cenusieSubstanta alba Invelisul
ventricularInvelisul subpial
Soma exista - - -Dendrite exista - - -Axoni amielinici exista - - -Sinapse exista - - -Axoni mielinici - exista - -Astrocite protoplasmatice exista in nr.
mareexista - exista
Astrocite fibroase exista exista in nr. mare
- exista
Oligodendrocite satelite exista - - -Oligodendrocite interfasciculare - exista - -Microglii exista exista - -Celule ependimare - - exista exista sau nu
Astrocitele si celulele ependimare alcatuiesc un invelis in jurul SNC numit invelis subpial, care se gaseste sub piamater. Astrocitele se interpun intre neuroni si orice altceva (neuroni si vase, neuroni si piamater etc.).
Bariera hematoencefalica (sange - creier)Capilarele cerebrale sunt continue, celulele fiind unite prin jonctiuni stranse (ocludens) care
sunt foarte selective. Transportul se face prin transcitoza redusa, fiind foarte selectiv. Permeabilitatea difera in functie de substanta:· mare - pentru H2O, CO2 si O2
· mica - pentru glucoza, Na+, K+, Mg2+, Cl-, HCO3-, HPO3
2-, uree· foarte mica: saruri biliare, catecolamine circulante· extrem de mica (aprox. zero) - pentru proteine
93

Functiile barierei hematoencefalice:1. mentine constanta mediului extracelular2. protejeaza creierul de toxinele circulante3. previne patrunderea neurotransmitatorilor (care ar putea actiona ca neurohormoni) in circulatia
generala.Oligodendroglie Microglie
Aspect la MO · forma ovala· nucleu rotund· prelungiri putine (10-50), groase,
putin ramificate
· forma eliptica· nucleu eliptic· prelungiri multe, cu ramificatii in
forma de spini· Aspect la
ME· Citoplasma cu aspect electronodens· RER, ribozomi, aparat Golgi,
mitocondrii microtubuli· Nu are lamina bazala
· Are lizozomi numerosi
· Localizare · in substanta cenusie sunt satelite ale pericarionilor
· in substanta alba sunt interfasciculare - intre axonii mielinici
· in substanta cenusie· in substanta alba
Oligodendrocitele formeaza teaca de mielina in SNC iar microgliile sunt macrofage ale SNC.
Astrocit fibros Astrocit protoplasmaticAspect la MO
· forma stelata· nucleu sferic, eucromatic, central· prelungiri subtiri, putin ramificate· citoplasma cu gliofibrile (se vad cu
impregnare argentica)
· prelungiri groase si ramificate· citoplasma omogena
· Aspect la ME
· contine filamente intermediare (gliofilamente) avand un diametru de 8 nm si alcatuite din GFAP (proteina acidafibrilara gliala cu greutatea moleculara de 51 kD).
· granule de glicogen· prezinta lamina bazala
· contine rare filamente intermediare (gliofilamente), izolate, avand un diametru de 8 nm si alcatuite din GFAP.
· granule de glicogen· prezinta lamina bazala
· Locali-zare si tipuri
· predomina in substanta alba· glia limitans - la periferia SNC, subpial
(celule in candelabru, celule penate)· perivasculare in jurul capilarelor
sangvine pe care aplica “picioruse”
· predomina in substanta cenusie· astrocite velate - in jurul sinapselor
(nevroglia Bergmann - cerebel, celula Muller - in retina)
Functiile astrocitelor:1. Constituie o bariera morfologica ce se interpune intre neuroni si vasele sangvine sau tesutul
conjunctiv.2. Participa indirect la formarea barierei hematoencefalice (reprezentata de celulele endoteliale ale
capilarelor cerebrale). Ele participa la geneza celulelor endoteliale, caci existenta lor alaturi de capilare ofera celulelor endoteliale cerebrale fenotipul caracteristic ce asigura competenta de bariera hematoencefalica (nu permit trecerea oricarei substante).
94

3. Au PR ce variaza in functie de K extracelular dar nu genereaza PA.4. Contribuie la mentinerea constanta a concentratiei extracelulare de K sau de neurotransmitatori,
pe care ii preia din mediul extracelular.5. reprezinta un suport structural:
6. in cursul dezvoltarii SNC ghideeaza prin prelungirile lor migrarea neuronilor7. prolifereaza si umplu spatiile ramase in urma distrugerii neuronilor. Neuronii distrusi
sunt mancati de macrofage (microglii)
Celulele ependimareLocalizare: captusesc ventricule si apeducte, acopera plexul coroid.Structura: sunt asezate pe un singur rand (aranjament epitelial - epiteliul ependimar), deci
celulele au polaritate. Domeniul apical prezinta: microvilei si cili (cu exceptia celulelor de pe podeaua ventriculului III. Domeniul laterobazal este turtit, (cu exceptia celulelor de pe podeaua ventriculului III, care se numesc fanicite si a caror membrana bazala trimite o prelungire spre eminenta mediana sau alte zone).
La ME se observa: multe mitocondrii, aparat Golgi situat apical, RER, jonctiuni gap si adherens, lipsesc jonctiunile ocludens (cu exceptia celulelor plexului coroid).
Exista zone speciale cu celule ependimare mici, turtite - zonele circumventriculare (organul subcomisural, OVLT, area postrema)
Functii:8. la nivelul plexului coroid secreta LCR si formeaza bariera sange-LCR9. restul formeaza bariera creier-LCR (bariera permeabila ce permite accesul mai multor substante).
Aceasta proprietate este folosita pentru administrarea unor medicamente ce nu depasesc bariera hematoencefalica: administrarea se face prin punctie. In caz de meningita, encefalita, se administreaza antivirale in acest mod.
Plexurile coroideSecreta LCR (14-36 ml/h). Volumul total al LCR este 150 ml. Aceste plexuri sunt formate
prin apozitia dintre piamater cu vas sangvine si epiteliul ependimar ce captuseste ventriculii. Suprafata este cutata si are o suprafata totala de 200 cmp. Ea are doua straturi:1. piamater cu vase sangvine2. celulele ependimare
Pliurile se numesc vilozitati coroide. O vilozitate coroida este formata din:· epiteliul ependimar· vase piale aferente· retea capilara· vase piale eferente
Celulele ependimare coroidiene sunt cuboidale, cu nucleu sferic, mitocondrii multe si RE abundent. Domeniul apical prezinta marginea in perie formata de microvili bulbosi.
Lichidul cefalorahidian (LCR)Este produs de catre plexurile coroide in cantitate de 0,35 ml/min (500 ml/zi). De asemenea
LCR mai este produs si prin curgerea fluidului extracelular prin epiteliul ependimar din ventriculi (ex.: area postrema din ventriculul IV)
LCR este in cantitate de 150 ml dintre care 30 ml se gasesc in ventriculi iar 120 se gasesc in spatiul subarahnoidian.
LCR curge din ventriculii I, II, III si IV in spatiul subarahnoidian. Din spatiul sub arahnoidian o mica parte se scurge prin spatiile perineurale din jurul radacinilor nervilor spinali
95

si cranieni iar cea mai mare parte se scurge prin specializari ale arahnoidei (vilozitati arahnoidiene) in sinusurile venoase mari din grosimea durei mater (ex.: sinusul sagital)
Functii:· mentine mediul fluid necesar creierului· protectie mecanica· comunicausor cu fluidul extracelular pentru ca bariera LCR-creier e permeabila· este locul de excretie a metabolitilor tesutului nervos· asigura preluarea de catre creier din LCR a substantelor necesare
SISTEMUL VASCULAR SANGVIN
Este un sistem inchis de tuburi (vase) prin care curge continuu sange. Peretele vasului are trei straturi concentrice numite tunici:
· la interior - intima· la mijloc - media· la exterior - adventitia
Arterele au un perete gros si un lumen rotund. Venele au perete subtire si lumen ovalar, turtit si mult mai mare.
Diagnosticul de vas sangvin· se observa structura tubulara· peretele este alcaatuit din mai multe straturi· lumenul este delimitat de celule endoteliale· in lumen se observa elementele figurate.
Structura (stratigrafia) peretelui vascular
Intima· endoteliu (structura mereu prezenta)· membrana bazal (structura mereu prezenta)· strat subendotelial· limitanta elastica interna (LEI)
Dispozitia elementelor este longitudinala. Functii: asigura schimbul si este netrombogenica (impiedica aderarea plachetelor prin: incarcarea negativa a endoteliului si secretia unor factori anti trombogeni).
Media· fibre musculare netede si/sau· lamele elastice· limitanta elastica externa.
Dispozitia elementelor este circulara (spiralata) si are functie de reglare a fluxului sangvin.
Adventitia· tesut conjunctiv (fibre si celule)
96

· fibre musculare netede (la vasele mari)· vasa vasorum (la vasele mari)· vase limfatice· nervi ai vaselor
Dispozitia elementelor este in general longitudinala dar poate fi si circulara. Functii: are rol mecanic (de ancorare), de protectie si de nutritie.
Vasele sangvine snt alcatuite din mai multe tipuri de tesut:· Epiteliu simplu pavimentos (asezat pe membrana bazala) - endoteliul. Membrana bazala este o
componenta amorfa. Ea are o componenta fibrilara alcatuita din colagen de tip IV sau V sintetizat de celulele endoteliale sau de fibrele musculare netede din stratul subendotelial.
· Tesut conjunctiv care contine:· fibre de colagen si reticulina: colagen III in medie si colagen I in adventitie. · fibre elastice izolate sau dispuse in lamele groase, ondulate.· fibroblasti - in adventitie
· Tesut muscular neted alcatuit din mai multe randuri de celule dispuse concentric. Intre celule exista jonctiuni de tip gap. Fibra musculara neteda vasculara se comporta fenotipic ca un fibroblast si in caz de agressssiune la nivelul peretelui vascular va raspunde ca un fibroblast, putand prolifera si capatand proprietati de fagocitare.
· Tesutul nervos - este alcatuit din:· fibre nemielinizate - apartin SNV si sunt sub forma de terminatii butonate· fibre mielinizate - se gasesc in zonele baroreceptoare si chemoreceptoare.· microganglioni nervosi
Clasificarea morfofunctionala a vaselor:· vase pentru conduce re - arterele mari, elastice· vase pentru distributie - arterele musculare· vase pentru reglarea presiunii - arteriole si metaarteriole· vase pentru schimb - capilare si venule post capilare (venule pericitare)· vase pentru intoarcere - vene mici, mijlocii si mari
Vasele de transport, diostributie si intoarcere fac parte din macrovasculatura. Sunt vizibile macroscopic si au vasa vasorum.
Vasele pentru reglarea presiunii si schimb sunt microvasculatura au un diamtru intre 10 mm si 300-500 mm si nu au vasa vasorum.
Artere Artere Arteriole Capilareelastice musculare continui fenestrat
esinusoide venule
pericitareVen
eEndoteliu c c c c fenestrate +/-
diafragmediscontinui c c
MB + + + + + discontinua sau absenta
+ +
Strat sub-endotelial
+ + exista numai la cele mari
p p +/- p p +/-
LEI + absenta la cele <50 mm cu exceptia celor renale
+/-
F.m.n. + +++ + (1-3 r) +L.el.
fenestrate+++ (>60 lame)
+ - rar +
LEE +
97

Tes. conj. + + + +F.m.n. + + + +Vasa
vasorum+ + +
Nervii vaselor
+ + +++
c = continuud = discontinuup = pericitr = randuri+++ = componenta principala a stratului.
Arterele elasticeArterele elastice cuprind lame elastice (L. el) dispuse pe mai mult de 60 de randuri intre care
exista fibre musculare netede (f.m.n.). Lamelele elastice sunt fenestrate, prezinta orificii mari ce asigura comunicarea intra straturile de lame elastice.
Venele, in functie de predominanta tesutului conjunctiv sau a f.m.n. se impart in vene conjunctive si musculare.
La arterele mari, vasele din vasa vasorum patrund doar pana la jumatatea externa a mediei, iar restul (jumatatea interna si intima) se hranesc din lumen. La vene, vasele din vasa vasorum ajung pana in stratul subendotelial (deoarece sangele venos este sarac in substante nutritive. Limfaticele urmaresc traseul vasa vasorum.
Microganglionii nervosi sunt in general rari dar se gasesc din abundenta in:· aa. coronare· aa. mezenterice· trunchiul celiac
Sunt artere elastice:· aorta· aa. pulmonare· a. carotida comuna· trunchiul brahiocefalic drept· a. subclavie· a. iliaca comuna
Exista vase cu perete mai gros (aa. coronare, aa. membrului inferior) si vase cu perete mai subtire (vasele intracraniene si pulmonare). Exista si vase cu ingrosari subintimale: arterele coronare, intracraniene si renale.
ArteriolaDiametrul lumenului este de 2 ori diametrul peretelui. Are 1-3 randuri de f.m.n. Unele
arteriole au LEI dar nici unele nu au LEE. Nu au vasa vasorum.
Metaarteriola (arteriola precapilara, aria sfincterului precapilar)Metaarteriola regleaza fluxul sangvin dar fara a intrerupe complet fluxul. Doar sfincterul
precapilar poate opri complet fluxul. Prezinta endoteliu si membrana bazala. Au un singur rand de f.m.n. discontinuu care
organizeaza la emergenta capilarului sfincterul precapilar - inel circular ce ralizeaza contractii ritmice la interval de 2-8 secunde. Ea are o forma conica. Intreg peretele vascular este inglobat
98

intr-un gel de proteoglicani si glicozaminoglicani care este hidratat in proportii variabile. Astfel turgiditatea vasului este variabila si are proprietati vasco-elastice.
Limfaticele urmaresc traiectul vasa vasorum. Jumatatea interna a peretelui vascular se harneste prin sangele din lumenul vasului, prin difuziune.
Intre celulele endoteliale si celulele musculare exista jonctiuni comunicante gap deoarece membrana baala prezzinta discontinuitati, fapt pentru care celulele musculare vin in contact direct din cand in cand cu celulele endoteliale.
CapilareleD.p.d.v. histollogic nu se deosebeste capilarele arteriale si cele venoase.Sistemul de capilarizare este de mai multe feluri:
· Normal: metaarteriola ® capilar ® venula postcapilara. Acest sistem se gaseste pretutindeni in organism.
· Sunt arterio-venos: intr-un sistem de capilarizare normal apare o comunicare intre metaarteriola si venula postcapilara care evita capilarele. Astfel se depaseste sistemul de capilare, fara a avea loc schimburile nutritive. Acest sistem de capilarizare se intalneste in mm. scheleltici si la nivelul pielii, in zona mainilor si picioarelor, la degete, unghii si urechi. La acest sistem, componenta arteriolara este foarte bin e inervata, si are rolul de a regla fluxul sangvin pentru a mentine constanta temperatura in aceste teritorii.
· Sistemul venos port: capilare venoase ® venula postcapilara ® capilare venoase. Acest sistem se intalneste in ficat si la nivelul tijei pituitare.
· Sistemul arterial port: capilare arteriale ® arteriola ® capilare arteriale. Acest sistem se intalneste in glomerulul renal care este constituit din arteriola aferenta ce se capilarizeaza la nivelul capsulei Bowman, de unde pleaca mai apoi arteriola eferenta ce se recapilarizeaza la nivelul tubilor uriniferi formand capilarele peritubulare. De asemenea, acest sistem se mai intalneste si la nivelul pancreasului, unde arteriolele se capilarizeaza mai intai la nivelul pancreasului endocrin (insulele Langerhans) si apoi se recapilarizeaza la nivelul pancreasului exocrin (acinii pancreatici).Capilarele sunt alcatuite din endoteliu, membrana bazala si pericite (celule Rouget).
Pericitele (celulele Rouget)Se pare ca aceste celule sunt celule musculare modificate. Ele au rol contractil si fagocitar.
Ca aspect, aceste celule au multe prelungiri cu ramificatii ce inconjura capilarul. Celula este cuprinsa in intregime intr-o clivare a membranei bazale (atat nucleul cat si prelungirile).
Pericitele exista si in capilarele din mm. scheletici dar si in capilarele sinusoide, fiind reprezentate de calulele parasinusoidale. Acesste celule au mare capacitate de a se diferentia in:
· osteoblaste· condroblaste· fibroblaste· fibre musculare netede· preadipocite, din care vor rezulta adipocitele albe.· microglii
Pericitele intervin si in procesul de cicatrizare a ranilor.
Organizarea morfologica generala a peretelui capilar:
Capilare Continui Fenestrate SinusoideCelulele endoteliale Sunt prezente in strat
continuuCelulele au fenestratii
(orificii)Stratul este
discontinuu, au spatii
99

largi. Celulele pot fi si fusiforme
Membrana bazala Este continua Este continua (exista si sub fenestratii)
Este discontinua sau poate chiar lipsi. Nu
exista in spatii.Pericitele Exista Sunt rare Exista sau nu. Ele sunt
prezente sub forma de celule parasinusoidale.
Diametrul 5-9 mm 5-9 mm 20-25 mmAlte caracterisitici Vezicule plasmalemale
Au jonctiuni stranseAu jonctiuni gap
Au jonctiuni stranseAu jonctiuni gap
Fenestrele au diametrul de 50-80 nm si sunt in numar de 20-60/mm2. Sub ele exista sau nu
diafragme cu grosimea de 5-6 nm.
Localizare In SNC, in nervi, in muschi, in glandele
exocrine, in plaman, in timus, in ganglionii
limfatici, in testicul si in retina
In glandele endocrine, in tractul gastro-
intestinal, in plexurile coroidesi in rinichi
(unde nu au diafragma)
In ficat, in splina, in maduva osoasa si in hipofiza anterioara
Retea abundenta de capilare se gaseste in ficat, rinichi, m. striat iar retea mai redusa se gaseste in m. neted sau tesutul conjunctiv dens.
Functiile capilarelor:· barierea selective:
· sange tesut (sange-lichid intestitial). Se considera ca distanta maxima a unei celule fata de capilarul sangvin este de 20 mm. Exceptii se gasesc in tendon, aponevroza, os sau in jumatatea interna a vaselor de calibru mai mare.
· sange-aer: la niveli; campului dehematoza, exista bariera alveolo-capilara.· de ultrafiltrare: la nivelul glomerulului renal
· functia metabolica· secretie de factrori inhibitori sau de crestere· functie antitrombogenica
Bariera este asigurata doar de capilarele continui si fenestrate.Bariera sange-tesut este foarte selectiva in unele organe:
· SNC (bariera hemato-encefalica)· timus· testicul· ochi (retina, umoare apoasa, cristalin)
La nivelul barierei hemato-encefalice se gasesc capilare fenestrate. Aceasta bariera nu opereaza la nivelul capilarelor din plexurile coroide, din aria postrema, de la nivelul eminentei mediane, din neurohipofiza, din organul pineal si din organele subcomisural si subfornical.
Selectivitatea mare a barierelor se datoreaza existentei jonctiunilor stranse , numarului scazut de vezicule plasmalemale si membranei bazale continui (in capilarele continui).
100

Bariera sange-tesut functioneaza in ambele sensuri. Ea exista la nivelul capilarelor continui si fenestrate dar nu exista la nivelul capilarelor sinusoide din ficat.
Stratigrafia generala a barierei sange-tesut:1. Invelisul mobil si fix al fetei luminale a celulelor endoteliale: electronegativitatea asigura
respingerea elementelor figurate din sange.2. Celula endoteliala prezinta canale de transcitoa si vezicule plasmalemale care faciliteaza
transportul substantelor prin peretele capilar. Celula propriu-zisa si jonctiunile intercelulare impiedica trecerea nutrientilor si gazelor. De asemenea, fenestratiile cu sau fara diafragme, permit sau nu trecerea dintr-o parte in alta a substantelor. La nivelul celulelor endoteliale are loc endocitoza mediata sau nu de receptori care se poate face cu sau fara ocolirea lizozomilor.
3. Membrana bazala impiedica traversarea peretelui capilar4. Pericitele
Factori care participa la schimb:· presiunea hidrostatica, oncotica, si gradientul de concentratie· activitatea specifica a celulei endoteliale· marimea si incarcarea electrica a moleculei
Astfel:· jonctiunile intercelulare permit trecerea moleculelor cu diametru mai mic de 2 nm.· Diafragmele fenestratiilor sunt puternic incarcate negativ si asigura selectivitatea transportului
d.p.d.v. electric. De asemenea, diafragmele fenestratiilor permit trecerea moleculelor cu diamtru mai mic de 10 nm.
· Veziculele plasmalemale si canalele de transcitoza asigura trecerea particulelor cu diametru mai mare de 10 nm, fara selectivitate electrica. Acest proces se numeste transcitoza sau diacitoza.
Venulele pericitareLa nivelul lor, pericitelel formeaza un strat continuu in membrana bazala a andoteliului,
avand prelungiri multe si ramificate. Au un diamtru mare, de 50-700 mm. La venulele cu diametru mai mare de 200 mm apar 1-2 straturi de fibre musculare netede. Prezinta jonctiuni slabe intre celulele endoteliale, fiind mai permeabile. Celulele endoteliale se pot contracta si atunci jonctiunile se desfac. Aceste procese au loc sub actiunea histaminei, serotoninei, bradikininei si prostaglandinelor.
Aceste vase reprezinta locul desfasurarii “dramei inflamatorii”. In jurul lor exista numeroase mastocite, eozinofile, macrofage si fibroblasti.
Venulele musculareContinua venulele pericitare. Ele au fibre musculare netede in loc de pericite.
101

GLANDE ENDOCRINE
Clasificarea mesagerilor chimiciDupa criteriul functional
· secretii autocrine: celulele elaboreaza un mesager chimic ce actioneaza pe receptori aflati chiar pe celula secretorie. Ele asigura controlul local al cresterii celulare. Ex.: factorii de crestere.
· secretii paracrine: mesagerul actioneaza pe celule adiacente celulei secretorii. Si acestea asigura controlul local al cresterii celulare. Ex.: majoritatea celulelor din sistemul neuroendocrin difuz (SND).
· secretii endocrine: realizate de un grup de celule specializate in sinteza si secretia mesagerului chimic, hormon, descarcat in mediul intern. El actioneaza pe tesutul tinta unde celulele au receptori specifici. Actiunea este la distanta.
Secretia de mesageri chimici poate fi continua sau numai cand organul o cere. Acelasi mesager poate fi eliberat de mai multe tipuri de tesut. Ex.: hormonii din mucoasa digestiva sunt peptide si pot fi secretate si de neuroni din nevrax. Un mesager chimic pote actiona pe mai multe tesuturi, avand mai multe efecte. O celula poate sa raspunda la actiunea mai multor mesageri chimici. Orice mesager poate actiona endo, para si autocrin.
· secretie sinaptica: mod de comunicare specific in tesutul nervos in care mesagerul interactioneaza cu receptori in membrana post sinaptica.Dupa criteriul chimic
· peptide si proteine:· peptide mici (10 aminoacizi): in hipotalamus, SND· peptide mari (10-50 aminoacizi): calcitonina (tiroida)· proteine (.50 aminoacizi): parathormonul si cea mai mare parte a factorilor de crestere
· glicoproteine (componenta glucidica poate ajunge la 30% din greutate): hormonii tropi hipofizari· steroizi (derivati ai colesterolului) hormonii corticosuprarenalieni, gonadali· derivati de aminoacizi:
· amine - medulosuprarenala· iodtironine - tiroida
· eicosanoide - derivati de acid arahidonic: · prostaglandine· tromboxani· leucotriene
· oxid nitric (NO)Dupa modul (mecanismul de actiune) al mesagerilor chimiciModul de actiune depinde de solubilitatea in apa.
· hidrosolubile: hormonii peptidici, proteinele. Difuzeaza liber prin mediul intern insa nu pot traversa membrana celulara. Interactioneaza cu proteinele mebranare de suprafata (receptorii membranar). Numarul de receptori membranari variaza in functie de gradul de diferentiere al celulelor dar si cu concentratia plasmatica a mesagerului chimic. Cand concentratia plasmatica creste, numarul de receptori se reduce - mecanism de autoreglare. Celula se
102

protejeaza de concentratia plasmatica prea mare a hormonului. Cresterea concentratiei poate determina insa si cresterea numarului de receptori membranari: prolactina si angiotensina II.
· hidrofobe: h. corticosuprarenalieni, gonadali. Nu poate circula in stare libera in mediul intern ci se combina cu proteina specifica de transport care il face hidrosolubil si astfel e adus la celula tinta unde se disociaza de proteina, difuzeaza prin membrana celulara, ajunge in citoplasma si interactioneaza cu proteinele citoplasmatice (receptorii intracelulari)Celula endocrina este celula care sintetizeaza si secreta substante cu rol de mesager chimic.
Originea celulelor endocrine· origine epiteliala - elaboreaza mesagerul chimic cu actiune la distanta· neuronii - elaboreaza neurohormoni si neuromodulatori· paraneuronii - au aspectul si morfologia unei celule receptoare, au capacitatea de a raspunde la
stimul adecvat prin secretia mesagerului chimic pe care il elibereaza in sange sau lichidul interstitial. Ei au mai multe caracteristici morfologice si functionale:· tipul de secretie: sintetizeaza si secreta peptide si/sau amine· la polul apical au microcriste (microvili) lungi cu rol receptor· citoplasma: prezinta granule de secretie (stocheaza produsul sintetizat)· toti paraneuronii au origine in crestele neurale
Distributia celulelor endocrine· pot forma organe specializate (glande endocrine):
· pineala (epifiza)· pituitara (hipofiza)· tiroida· paratiroida· suprarenala
Aceste structuri respecta acelasi plan de organizare care are mai multe caracteristici:· au capsula conjunctiva fibroasa care trimite spre parenchimul glandelor niste septuri ce pot
compartimenta complet sau incomplet glanda in lobi sau lobuli.· parenchimul e format din celule endocrine ce vin in contact cu cpilare ce functioneaza
favorizand schimburi repetate, de obicei capilare fenestrate dar si sinusoide.· stroma e formata din retea alcatuita din fibre de reticulina si celule conjunctive (fibroblasti si
mastocite)· fibrele nervoase vegetative se distribuie la vasele din bogata retea vasculara· au o bogata retea vasculara
Structura glandei pote veni uneori in contact cu celulele endocrine· celulele endocrine exista sub forma de grupuri celulare (cuiburi sau insule celulare) incluse in
parenchimul unor alte glande specializate in alta functie; pancreasul, gonadele.· celulele endocrine sunt dispersate in structura diferitelor tesuturi si organe si formeaza sistemul
neuroendocrin difuz (SND).
Sistemul neuroendocrin difuz (SND)SND este alcatuit dintr-o varietate mare de celule ce sunt capabile sa sintetizeze anumite
peptide si amine produse pe care le depoziteza in citoplasma sub forma granulelor de secretie eliberate de celula prin exocitoza. Aceste substante au actiune autocrina, paracrina, endocrina, de neurohormoni sau neuromodulatori.
IstoricPrima descriere a acestor celule endocrine din afara glandelor a fost facuta de Heidenheim.
El a evidentiat celule endocrine in mucoasa gastrica si intestinala cu dicromat de potasiu. Acestea se numesc celule cromafine.
103

Bayless si Starlin au evidentiat la nivelul mucoasei duodenale celule implicate in sinteza hormonilor. Secretia ajunge in mediul intern actionand pe acinii pancreatici, stimuland secretia pancreatica.
Pearce introduce conceptul de sistem APUD format din toate celulele endocrine dispersate dar cu origine embriologica comuna, similare d.p.d.v. structural si functional. Celulele incorporeaza precursori (aminoacizi) din care sintetizeaza mesagerul chimic si pe care apoi il stocheaza.
Feyrtec introduce conceptul de SND format din celule endocrine ce au origine nervoasa (din crestele neurale).
A avut loc evidentierea unei enolaze neuronale specifice (NSE) cu localizare perinucleara la toate celulele endocrine dispersate. Ea este ........... netural al unei enzime din glicoliza.
Din 40 de celule ale SND, 7 se gasesc in organe nervoase iar restul in organe endodermale. SND este alcatuit din
· celule endocrine ce au asemanare functionala cu neuronii (isi cresc activitatea electrica sub actiunea stimulului)
· celule endocrine clasice (sub actiunea stimulului sintetizeaza mesager chimici si ii stocheaza)Celulele SND au raspandire ubicuitara, de obicei intraepitelial.Clasificarea celulelor din SND
· celule ale SND ce fac parte din glandele endocrine clasice (adenohipofiza, tiroida, paratiroida, MSR, pineala). Aceste celule reprezinta o componenta structurala importanta a glandei.
· celule ale SND se pot intalni in tesuturi si organe ce fac parte din alte sisteme:· aparatul digestiv (de la esofag pana la canalul anal). Aceste celule alcatuiesc axa
gastroenteropancreatica. Ele se mai gasesc si iin peretii canalelor de excretie ce se deschid in duoden (coledoc, pancreatic). Ele se afla in mucoasa si submucoasa digestiva.
· piele· aparatul respirator· tractul urogenital· glanda mamara
· celule ale SND pot fi si neuroni peptidergici.Orice celula din SND se caracterizeaza prin:
· sintetizeaza si secreta amine si peptide care se inrudesc fiecare intre ele.· dau reactie pozitiva pentru enolaza neuronala specifica· au antigene comune
Relatia APUD-SNDExista si celule care nu au capacitatea de a incorpora si stoca precursori deci nu sintetizeaza
amine. Spre exemplu celule ce secreta calcitonina (nu amine). Proprieteatea de a incorpora precursori si a stoca amine nu este specifica numai celulelor endocrine ci si glandelor exocrine: acinii pancreatici incorporeaza precursori - aminoacizi si stocheaza amine.
Hipotalamusul endocrinEste un organ ce apartine SNC. Se gaseste in partea ventrala a diencefalului si la baza
ventriculului III. Delimitare:· superior - e separat de talamus prin santul hipotalamic· anterior - comisura alba anterioara si in jos lamina terminalis si chiasma optica· posterior - fosa interpedunculara· inferior eminenta mediana (componenta a infundibulului)......................................
El are o arhitectura de organ nervos, fiind alcatuit din neuroni (corpi neuronali si prelungiri). Corpii neuronali formeaza nucleii iar substanta alba este alcatuita din prelungiri (aferente si eferente) si celule gliale.
104

Hipotalamusul endocrin nu este diferit structural de hipotalamusul nervos. O parte din neuroniii din structura hipotalamusului isi pastreaza proprietatea de celule neuronale dar capata si proprietati de sinteza si secretie de mesageri chimici. Acesti neuroni secreta neurohormoni si neuromodulatori. Clasificarea neuronilor din hipotalamusul endocrin s-a facut prin imunocitochimie si dupa un criteriu functional. Totalitatea neuronilor ce partin hipotalamusului endocrin au organizare ca a oricarui neuron si in plus au un aparat de sinteza bine dezvoltat (RER, aparat Golgi, microtubuli si granule de secretie citoplasmatice).
D.p.d.v. functional exista doua tipuri de neuroni:1. neuroni peptidergici2. neuroni secretori de peptide hipofizare
Neuronii peptidergici sunt neuroni secretori de peptide ce se stocheaza in hipofiza posterioara. Sunt neuroni de talie mare (magnocelulari). Acestia sunt localizati aproape in totatlitate in doi nuclei hipotalamici: paraventricular si supraoptic. Ei secreta ADH (arginin vasopresina) si ocitocina.
La MO si la ME, prin imunocitochimie, s-a observat ca neuronii au talie mare (25 mm in diamtru), in citoplasma prezinta granule de secretie de talie mare (160-200 nm). Axonii au traseu descendent spre lobul posterior al hipofizei si sunt nemielinizati. Conexiunea intre cei doi nuclei si hipofiza posterioara se face individual, dela nucleul supraoptic la hipofiza si de la nucleul paraventricular la hipofiza.
O parte din axonii neuronilor din nucleul supraoptic nu ajung la lobul posterior ci se termina la nivelul eminentei mediane unde exista un plex capilar la nivelul caruia elibereaza continutul. Pe traseul axonillor exista dilatatii unde se stocheaza sub forma de granule de secretie peptidul sintetizat. Dilatatiile sunt numite corpi Hering.
Sinteza peptidelor in hipofiza posterioara are loc in pericarion si se face sub forma unor molecule precursoare ce contin alaturi de peptid si o proteina de transport care se numeste neurofizina. Sinteza peptidelor si a neurofizinelor are loc concomitent, cele doua molecule legandu-se intre ele prin legaturi necovalente.
In vecinatatea nucleilor hipotalamici exista capilare. Peptidele nu pot fi insa secretate local, aici existand barieira hematoencefalica ce nu poate fi depasita de produsii de secretie. Substantele vor fi vehiculate prin axoni, cu ajutorul microtubulilor, prin polimerizari si depolimerizari si vor ajunge in hipofiza posterioara sau in corpii Hering. La unele granule, in timpul transportului, sub actiunea proteinazelor, membrana se rupe si peptidul se separa de neurofizina si se vor forma granule se parate de peptid si neurofizina.
Eliberarea peptidului se face prin crearea de potentiale electrice prin stimularea receptorului specific din sitemul vascular. Se creeaza impulsurri care ajung la neuronul secretor din hipotalamus care declanseaza la cpatul proximal al axonului un potential de actiune ce se propaga pana la nivelul secventei terminale. Ca urmare se vor deschide canalele de Ca2+ iar peptidul va fi eliminat in capilarele sangvine cu care vine in raport axonul, prin exocitoza dependenta de Ca2+.
Axonii neuronilor secretori nu se termina sinaptic la nivelul lobului posterior (nu formeaza sinapse) ci se termina asinaptic, prin conexiune neurovasculara, cu capilarul sangvin.
Neuronii secretori de peptide hipofizare sunt de talie mica (parvocelulari) si se afla in nucleii hipotalamici din aria numita aria hipofizotropa:· nucleul arcuat, unde se secreta GH-RH, GH-IH, dopamina (pentru prolactina), Gn-RH· nucleii ventromediali unde se secreta GnRH si TRH· nucleii periventriculari, unde se secreta CRH, GH-IH· nucleii paraventriculari, unde se secreta TRH, CRH, Gn-RH, GH-IH
In aceasta arie se secreta peptide ce influenteaza activitatea secretorie a celulelor din adenohipofiza. Ei sunt neurohormoni. Aceste substante au influente:
· secretorii (GH-RH) si inhibitorii (GH-IH=somatostatina)· inhibitorii - pentru prolactina
105

· secretorii - pentru Gn-RH, TRH, CRHAcesti neuroni au talie mica (15 mm) si granule de secretie mici (100-110 nm). Axonii
neuronilor din aria hipofizotropa se temina si ei asinaptic, prin conexiune neurovasculara. Ei formeaza tractul tuberoinfundibular, care este o conexiune nervoasa intre aria hipofizotropa si eminenta mediana.
Mecanismul eliberarii este prin feed-back negativ asigurat de concentratia plasmatica a hormonului secretat de glanda endocrina aflata sub controlul trop hipofizar, controlat la randul lui de un neuron din aria hipofizotropa.
Glanda hipofizaAre rol integrativ pentru ca are legaturi cu hipotalamusul si secreta hormoni tropi.Ea este acoperita de o capsula conjunctiva ce provine din duramater si compartimenteaza
zone din parnechim in lobi. Ea ajunge pana la depresiunea osului sfenoid (seaua turceasca) sii adera la periostul seii turcesti.
Hipofiza are doua componente:· hipofiza anterioara = adenohipofiza - ea deriva dintr-o evaginare a stomodeumului primitiv
(punga Rathke). Are structura glandulara si este formata din mai multi lobi:· lobul anterior (75% din adenohipofiza)· lob intermediar, aflat in raport cu lobul anterior· lob tuberal
· hipofiza posterioara = neurohipofiza - ea are origine nervoasa, fiind determinata de o extensie (evaginare) a pereetelui ventriculului III pana la nivelul fosei pituitare. Si ea are 3 componente:· lobul posterior, situat lateral de adenohipofiza, in seaua turceasca· se continua cu tija pituitara (infundibulara)· baza ventriculului III retrochiasmatic = eminenta medianaHipofiza are fibre nervoase vegetative vasomotorii. Nu s-a observat distributia fibrelor la
celulele secretorii.Vascularizatia este bogata, asigurata de ramuri ale a. carotide interne, aa. hipofizare
superioara si inferioara.A. hipofizara superioara patrunde in tija pituitara si vascularizeaza lobul lateral, tija pituitara
si eminenta mediana. Din ea se desprinde un plex capilar primar in care se termina tractul tuberoinfundibular si o parte din axonii neuronilor din nucleii supraoptici. Acest plex se prelungeste in tija si determina niste vase de calibru mare, vase portale ce se capilarizeaza la nivelul lobului anterior hipofizar unde formeaza un plex capilar secundar. Toate aceste trei elemente alcatuiesc sistemul port hipofizar. Plexul secundar se extinde si la nivelul lobului intermediar, denumirea de lama avasculara fiind improprie pentru lobul intermediar, aici avand loc anastomoze cu vasele din lobul posterior.
Dispozitivul vasculaar ce uneste hipotalamusul cu lobul anterior al hipofizei se numeste Popa-Fielding. In acest dispozitiv, fluxul sangvin are dublu sens:
· sens descendent: sangele transporta de la plexul capilar primar la adenohipofiza hormoni produsi in aria hipofizotropa si peptidul ADH produs in nucleul supraoptic. ADH are efect specific asupra celulelor din adenohipohiza, stimuland secretia de CRH.
· Sensul asecendent se face in proportie redusa. Prin el se realizeaza un feedback scurt, prin concentratia hormonilor adenohipofizari . Astfel este controlata secretia de hormoni tropi la nivelul ariei hipofizotrope.A. hipofizara inferioara vascularizeaza lobul posterior si formeaza aici un plex capilar. Acest
plex capilar primar are dispozitie in palisada (capilarele sunt paralele).Elemente de structura a hipofizeiHipofiza anterioara
106

Are o structura predominant celulara.
Lobul anterior (distal)Celulele se organizeaza in cuiburi sau cordoane celulare, inconjurate de o retea fina de fibre
de reticulina. Celulele se afla in raport cu capilarele sinusoide.`Clasificarea celulelor din lobul anterior se face dupa metode imunocitochimice, in celule
granulare si celule agranulare.· celulele granulare sunt endocrine si elaboreaza hormonii. Denumirea este data de hormonul
sintetizat:· somatotrofe (acidofile, PAS negative) - sintetizeaza somatotropul (STH=GH)· lactotrofe (acidofile, PAS negative) - sintetizeaza prolactina (PRL)· corticotrofe (bazofile, PAS pozitive) - sintetizeaza proopiomelanocortina (POMC)· tireotrofe (bazofile. PAS pozitive) - sintetizeaza h. tireotrop (TRH)· gonadotrofe (bazofile, PAS pozitive) - sintetizeaza h. foliculostimulant (FSH) si h.
luteinizant (LH) sau h. de stimulare a celulelor interstitiale (ICSH)Aceste celule pot fi :
· cromofile (fixeaza colorantul) acidofile sau bazofile)· cromofobe (nu fixeaza colorantul)
Aceste celule pot fi PAS pozitive sau negative in functie de natura hormonului secretat: STH si PRL sunt peptide si deci PAS negative iar POMC, TSH si GnRH sunt glicoproteine si deci PAS pozitive.
· celulele agranulare pot fi foliculostelare (bazofile) sau celule STEM (cromofobe)· celulele foliculostlare au la mijloc cordoane celulare. La polul apical au microvili si
ocazional cili. Ele nu au rol secretor ci rol metabolic, trofic pentru celulele adiacente.· despre celulele cromofobe exista doua supozitii:
· sunt precursoare ale celulelor granulare, de unde si denumirea de STEM (de origine)
· reprezinta un stadiu final de evolutie a celulelor granulare (o celule granulara care si-a pierdut granulatiile)
Dispozitia celulelor in lobul anterior hipofizar:· celulele somatotrofe sunt distribuite pe partile laterale ale lobului anterior. tumorile secretante de
STH duc la modificari de volum ale seii turcesti.· celulele corticotrofe sunt situate in portiunea mediana si spre posterior. In stadii tardive de
tumori ale acestor celule apar modificari de glanda pituitara.· celulele tirotrofe sunt situate in portiunea mediana si anterioara.· celulele lactotrofe, gonadotrofe si celulele agranulare sunt distribuite difuz.
Fiecare tip celular, in conditii fiziologice dar si patologice poate elibera mai multi hormoni:· celulele corticotrofe elibereaza POMC care se scindeaza in: corticotropina (ACTH),
melanotropina (MSH), b lipotrofina si b endorfiiina· celulele gonadotrofe secreta FSH si LH.· in conditii patologice apar celulele somatomamotrofe (celule acidofile) care secreta STH si PRL.
Prin ME de transmisie s-a descoperit ca celulele lactotrofe au granulatiile cele mai mari (in sarcina ajung pana la 900 nm) iar celulele tireotrope au granulatiile cele mai mici.
Lobul intermediarEste separat de lobul anterior prin fanta Rathke si se alipeste de lobul posterior. Celulele
secretorii se organizeaza in cordoane celulare ce contin celule in cea mai mare parte bazofile. mai rar aparand celule cromofobe. Langa cordoanele celulare, in lobul intermediar pot apare si
107

foliculi cu coloid care reprezinta resturi din punge Rathke. Celulele din folicul sunt diferite structural de cele din cordoane.:
· in folicul, celulele au putine organite de sinteza, au un continut citoplasmatic format din numeroase vacuole clare si prezinta un microvil sau cil la polul dinspre......................
· in cordoane, celulele au granule de secretie fiind celule impliccate in sinteza si secretia de POMC, adica b-lipotrofina si a-MSH in special.Exista un fenomen de migrare a unor celule din lobul intermediar spre cel posterior. Cum
celulele sunt in majoritate bazofile, are loc infiltrarea bazofila a lobului posterior.
Lobul tuberalAre un numar redus de celule granulare. El incojura incomplet tija pituitera si are o structura
predominant celulara. Celulele secretorii sunt organizate ca in lobul anterior, in cordoane si foliculi. Celulele sunt de talie mica si bazofile, fiind implicate in secretia de gonadotropi. Acest lob este foarte bine vascularizat, prin vase de calibru mare ce intra in componenta sistemului port.
Hipofiza posterioaraAre o structura predominant fibrilara. Are neuroni secretori in nucleii supraoptic si
paraventricular si in rest prezinta axonii nemielinizati ai acestor neuroni. Mai prezinta de asemenea si cateva fibre catecolaminergice ce controleaza activitatea de descarcare a peptidelor hipofuzare. Mai apar in plus urmatoarele elemente:
· retea formata din fibre de reticulina· multe capilare fenestrate· celule (pituicite). Ele sunt asemantoare unor strate protoplasmatice cu rol trofic pentru fibrele
nervoase si cu continut mare de lizozomi, deci cu rol de fagocitoza.
Tija pituitaraPe langa elementele fibrilare mai apar si artere mici si capilare de tip glomerular, ce iriga
formatiunile de la acest nivel.
Eminenta medianaEste organizata in 3 straturi:
1. stratul ependimar, alcatuit din tanicite (un tip particular de celule)2. stratul mijlociu, alcatuit din fibre nervoase3. stratul palisadic, alcatuit din capilar ce apartin plexul capilar primar din sistemul port.
Tanicitele au la polu apical (spre ventriculul III) un microvil. La polul opus au o prelungire ce vine in contact cu plexul capilar primar. In citoplasma sunt vizibile granule ce nu sunt vezicule ce stocheaza produs de secretie ci contin celule endocitate din LCR din ventriculul III si care prin prelungiri ajung in plexul capilar primar. Exista si un sens invers pus in dicutie: descarcarea in LCR a unor hormoni preluati din plexul capilar primar.
Glanda pineala (epifiza)
108

Epifiza a fost incadrat in sistemul neuroendocrin difuz de catre C.I. Paulescu care, in perioada 1936-1940, a demonstrat ca glanda are actiune secretorie.
Ea se gaseste in SNC, localizata intre corpul calos si coliculii cvadrigemeni superiori. Ea este legata de tavanul ventriculului III printr-o formatiune de substanta alba numita habenula. Ca si celelalte glande endocrine, epifiza are si ea o capsula conjunctiva formata de piamater care trimite septuri in parenchim ce compartimenteaza incomplet glanda in lobuli. Parenchimul este organizat in cuiburi (gramezi) sau cordoane.
Glanda este bogat vascularizata, aici existand cel mai mare flux sangvin din sistemul endocrin.
Inervatia provine din SNV prin fibre amielinice ce reprezinta axoni ai neuronilor din ganglionul cervical superior din lantul paravartebral simpatic. Aceste fibre detin controlul asupra secretiei hormonale a glandei. Mai exista de asemenea si fibre mielinice cu originea in SNC, fibre aferente venind din comisura alba superioara si din nucleul talamic numit habenular.
La MO, glanda apare ca o formatiune eterogena alcatuita din celule si fibre nervoase. Pe masura inaintarii in varsta apar modificari: · in parenchim se dezvolta formatiuni chistice care inlocuiesc anumite zone din el.· creste cantitatea de tesut conjunctiv· lobulatia este mai completa· apar concretiunile calcare care se mai numeste si nisip cerebral, corpora arenacee, corpii
Psamona sau acervulus cerebri. Aceste formatiuni sunt depozite de fosfat de calciu si carbonat de calciu si se formeaza in timp, motiv pentru care ele au structura lamelara, aspect coraliform sau muriform. Ele pot fi localizate in spatiile perivasculare sau in parenchim.Glanda pineala este de origine nervoasa. Parenchimul sau este alcatuit din pinealocite si din
celule gliale (interstitiale).
PinealocitelePinealocitele sunt paraneuroni. Paraneuronii sunt celule care d.pd.v. morfologic sunt la fel
cu celulele receptoare, prezentand la polul apical microvili cu rol receptor. Ei au capacitatea de a raspunde prin sinteza si secretie de mesageri chimici la actiunea unui stimul. Din aceasta cauza ca si din cauza faptului ca mesagerii chimici secretati sunt amine si polipeptide mici paraneuronii apartin sistemului neuroendocrin difuz.
Corpul lor are doua sau mai multe prelungiri care se termina in apropierea capilarelor sangvine in mod butonat. Citoplasma este slab bazofila si prezinta incluziuni: de glicogen, lipidice, pigmentare iar dupa puberatate apar si incluziunile de lipofuscina. Citoplasma acestor celule est bogat in enzime implicate in sinteza de amine biogene. De asemenea aparatul de sinteza este bine dezvoltat si contin multe granule de secretie mici cu diametrul de 100 nm. Nucleul este mare si eucromatic.
Intre pinealocite exista un sistem de canalicule ce comunica intre ele si formeaza sitemul labirint ce comunica cu LCR printr-o evaginare la nivelul ventriculului III. Acest sistem are doua roluri:
· transport de molecule catre pinealocite· armonizarea functionala a populatiei de pinealocite
Activitatea secretorie are ca rezultat doua categorii de mesageri chimici:1. derivati de benzopirol (indol amina): melatonina2. peptide: argininvasotocina si angiotensina II.
Celulele glialeD.p.d.v. structural seamana cu astrocitele fibroase. Inconjoara incomplet capilarul sangvin
sau pinealocitul.
109

Rolul glandei pinealeGlanda pineala este o componenta a sistemului neuroendocrin difuz, fiind un traductor
neuroendocrin: are capacitatea de a transforma un impuls nervos in secretie de mesageri chimici.La nevertebrate are functie de fotoreceptie, fiind un organ senzorial. La mamifere si om
aceasta functie nu se pastreaza dar se pastreaza fotoperidiocitatea acestei functii, activitatea secretorie fiind coordonata cu ritmul circadian.
La om glanda isi dezvolta inervatia pinealopeta (aferenta) formata de fibre simpatice provenind din ggl. cervical superior. Aceste fibre sunt cuplate prin hipotalamus cu retina, fapt care asigura fotoperiodicitatea functiei secretorii.
In retina, populatia celulelor fotoreceptoare este neomogena existand unele care transmit impulsul nervos pana la cortex unde este elaborata senzatia constienta de vaz si altele care mediaza secretia de melatonina.
La intuneric celulele care mediaza secretia de melatonina iniotiaza impulsuri nervoase care ajung la neuronii bipolari, la cei multipolari si apoi iau calela nervului optic. De aici se desprind si merg prin tractul retinohipotalamic ajungand la nucleul suprachiasmatic din hipotalamus. Acest nucleu are legaturi cu maduva spinarii din regiunea toracala, coarnele laterale. Aici are loc o sinapsa. Axonii acestor neuroni sunt fibre preganglionare ce fac sinapsa in ggl. cervical superior. Axonii neuronilor de la acest nivel sunt fibre postganaglionare ce se termina la nivelul pinealocitelor din gl. epifiza. Aceasta ultima sinapsa este mediata de noradrenalina care stimuleaza sinteza de melatonina care va fi eliminata prin difuzie in mediul intern.
La lumina celulele care mediaza secretia de melatonina isi hiperpolarizeaza membrana celulara, fapt care le face mai greu excitabile deci se inhiba eliberarea de noradrenalina la nivelul fibrelor postganglionare simpatice. Ca urmare a acestor succesiuni de evenimente secretia de melatonina are un ritm circadian, cu un maxim de secretie noaptea. Acest ritm se sincronizeaza cu ritmul endogen al neuronilor din nucleul suprachiasmatic din hipotalamus.
Efectele melatoninei:· inhiba tonusul SNC, avand un efect hipnotic. Astfel melatonina este folosita in tratamentul unor
insomnii cronice.· antigonadal si prin urmare antiovulator datorita inhibitiei secretiei de LH. Ea poate fi deci
implicata in etiopatogenitatea unor forme de sterilitate.· stimuleaza sistemul imun, fiind corelata cu modificarea ritmului de crestere a unor celule
tumorale (inhiba cresterea tumorilor)· este corelata cu procesul de senescenta (imbatranire): o scadere a secretiei de melatonina duce la
o accelerare a procesului de imbatranire.
Glanda tiroida
Este cea mai mare glanda la adult. Ea este alcatuita de obicei din doi lobi uniti printr-un istm.Are o dubla origine embriologica: endodermala si ectodermala. Partea de origine
endodermala deriva dintr-o prelungire de la baza limbii care este canalul tireoglos. In mod obisnuit acesta se oblitereaza si se atrofiaza dar pot exista si exceptii:
· aparitia lobului piramidal, formatiune fara implicatii patologice.· chisturi in canal, care trebuie eliminate chirurgical· tesut tiroidian ectopic dintr care cea mai frecventa este tiroida liguala (portiunea initiala a
canalului tireoglos)De origine ectodermala sunt celulele parafoliculare care deriva din crestele neurale si sunt o
componenta a sistemului neuroendocrin difuz.
110

Glanda depoziteaza mari cantitati de hormoni, extracelular, la nivelul unei substante omogene numita coloid. Depozitul la adult este suficient pentru acoperirea necesarului timp de 2-3 luni.
Capsula este de natura conjunctiva si compartimenteaza glanda in lobuli. In lobuli celulele sunt organizate in foliculi. Intre foliculi exista stroma interfoliculara, element de diagnostic diferential intre glanda mamara in lactatie si tiroida. Glanda mamara (tubuloacinoasa), in lactatie, are un aspect asemanator cu foliculii tiroidieni numai ca nu prezinta stroma. La glanda tiroida, dupa varsta de 50 de ani stroma interfoliculara se umple cu adipocite (sufera o transformare grasoasa).
Este bogat vascularizata, avand multe capilare de tip fenestrat ce vin in raport cu peretele folicular.
Inervatia tiroidei este de natura vegetativa dar exista o deosebire fata de celelalte glande endocrine. De obicei, la alte glande endocrine terminatiile nervoase actioneaza la nivelul capilarelor sangvine modificand fluxul de sange si implicit si secretia glandei. La nivelul tiroidei exista terminatii nervoase ce ajung pana la capilare, influentand indirect secretia dar si terminatii care ajung pana la nivelul peretelui folicular, influentand direct secretia de hormon.
In mod normal, foliculii tiroidieni au forme si dimensiuni variate. Foliculul este o cavitate inchisa, delimitata de un epiteliu simplu cubic asezat pe o membrana bazala. In cavitate se gaseste coloidul. La periferia coloidului, pe preparatele obisnuite cu HE se pot vedea unele spatii, uneori asemanatoare cu vacuolele. Multa vreme s-a crezut ca acestea sunt vacuole de resorbtie dar apoi s-a stabilit ca ele sunt artefacte de fixare deoarece ele nu apar deloc in produsele fixate prin difuzie.
Coloidul are caracter amfoter, colorandu-se cu coloranti acizi cat si cu coloranti bazici. De asemenea coloidul este PAS pozitiv deoarece tiroglobulina, depozitul si precursorul hormonilor tiroidieni este o glicoproteina.
In functie de aspectul foliculilor se poate stabili starea de actiune a glandei: daca sunt mari si cu epiteliu turtit avem hipotiroidie iar daca foliculii sunt mici si cu epiteliu inalt, columnar, observandu-se si celule in mitoza si coloid putin, avem de a face cu hipertiroidie.
Parenchimul glandular este alcatuit din doua tipuri de celule: celulele foliculare (tirocite) si celulele parafoliculare C (“C” de la “clar”, celulele avand citoplasma palida, clara sau de la “calcitonina”, hormonul secretat de aceste celule).
TirociteleTirocitele au o polaritate diferita d.p.d.v. al distributiei organitelor in citoplasma.La MO se observa ca celulele au citoplasma bazofila, pozitia nucleului variaza in functie de
inaltimea celulei foliculare, dar de obicei este situat inspre polul bazal.Tirocitul fiind specializat in sinteza, secretia, reabsorbtia si digestia de proteine se poate
deduce tipul de organite celulare, cantitatea lor si asezarea in celula:· Au un continut mare de RER. In perioadele de sinteza RER apare ca cisterne dilatate si
dispersate prin citoplasma.· Aparat Golgi bine reprezentat. In repaus secretor este situat spre polul bazal fata de nucleu iar in
activitate este situat spre polul apical (supranuclear). Aceasta schimbare de pozitie continua a fost denumita “valsul golgian”.
· Prezinta multe mitocondrii cu criste filamentoase dispersate in citoplasma.· La polul apical, spre coloid prezinta microvili, ocazional se observa si unele pseudopode. La
acest nivel se formeaza veziculele de pinocitoza.· Prezinta multi lizozomi sau fagolizozomi (atunci cand au fuzionat cu veziculele de endocitoza)
in special in zona apicala.Citoplasma prezinta doua tipuri de vezicule:
· mici, care contin tiroglobulina care va fi exocitata in coloid.
111

· mari, care sunt picaturi endocitate de coloid.
Biosinteza hormonilor tiroidieni (triiodotironina si tetraiodotironina) are loc in mai multe etape:1. sinteza, secretia si depozitarea tiroglobulinei, etapa care se desfasoara concomitent cu cea de-a
doua etapa;2. captarea din circulatie si oxidarea iodului;3. iodarea tiroglobulinei si formarea hormonilor tiroidieni;4. secretia hormonilor tiroidieni.
Preluarea precursorilor (aminoacizii) din circulatie are loc prin membrana bazala printr-un transport activ. In ribozomi are loc sinteza componentei proteice. In lumenul RE are apoi loc prima etapa a glicozilarii: incorporarea manozei dupa care prin vezicule mici proteina ajunge in aparatul Golgi unde are loc a doua etapa de glicozilare, incorporarea de galactoza. Apoi tiroglobulina este impachetata in vazicule de secretie de origine golgiana si este exocitata in coloid.
Preluarea iodului anorganic din circulatie se face tot pe la polul bazal, activ, impotriva gradientului de concentratie si a gradientului electric. Preluarea se face cu ajutorul pompei de iod care este stimulata de tirotropina (TSH). Iodul este concentrat, atingand o concentratie de cateva zeci de ori mai mare decat cea din plasma, uneori chiar de 40 de ori. Iodul este transportat la polul apical unde este oxidat cu ajutorul peroxidazei lizozomale si transformat in iod organic, molecular. Tiroidita autoimuna (Hashimoto), care reprezinta 80% din cazurile de hipotiroidie, se datoreaza aparitiei anticorpilor fata de structurile proprii ale glandei tiroide. Exista astfel o concentratie foarte mare de anticorpi anti peroxidaza lizozomala, ceea ce determina imposibilitatea oxidarii iodului.
Iodarea tiroglobulinei se face la resturile de tirozina care sunt foarte multe (aprox. 140 de resturi de tirozina in fiecare molecula de tiroglobulina). Iodarea tirozinei se face in molecula de tiroglobulina, aflata la interfata coloid - pol apical tirocite, tiroglobulina aflandu-se in coloid. Tirozina iodata ramane in tiroglobulina. Dupa iodare rezulta monoiodtirozina (MIT) si diiodotirozina. Acest proces este catalizat de iodaze (iodinaze). Apoi, prin condensarea unei molecule de DIT cu una de MIT se obtine T3 (triiodotironina) iar prin condensarea a doua molecule de DIT se obtine o molecula de T4 (tetraiodotironina). T3 si T4 raman inglobate in molecula de tiroglobulina care se afla in coloid.
In cazul stimularii tiroidei, in scopul secretiei de hormon, are loc resorbtia tiroglobulinei prin endocitoza la nivelul microvililor sau pseudopodelor. Sub actiunea protezelor sunt indepartate apoi acestea difuzeaza prin membrana bazala in circulatie, fiind liposolubile.
Deci, in procesul de sinteza a h. tiroidieni apar doua faze:1. faza exocrina (este exocitata tiroglobulina in coloid)2. faza endocrina (difuzia hormonilor in circulatie)
Actiunile h. tiroidieni sunt de stimulare a proceselor metabolice mai ales metabolismul oxidativ.
Celulele CFac parte din sistemul enuroendocrin difuz (SND).Ele sunt mai mari decat celulele foliculare putand sa apara izolate sau in grupuri mici. Ele
sunt prezente in numaar mai mare la copil decat la adult. Ele sunt localizate in:· stroma interfoliculara· peretele foliculului tiroidian, fiind situata in grosimea membranei bazale a foliculului.
Celula C nu vine niciodata in contact cu coloidul. Are un nucleu eucromatic si citoplasma bazofila. La ME se observa ca celula C nu are
polaritate d.p.d.v. al dispozitiei organitelor in citoplasma (ca celula foliculara), organitele fiind dispersate in toata citoplasma. Ea prezinta granule de secretie cu diametrul de 150 nm care contin calcitonina (tirocalctonina), hormon care inhiba mobilizarea sarurilor fosfocalcice.
112

Glandele paratiroide
Sunt glande vitale, prin rolul lor in homeostazia calciului. Ele deriva din endoderm si adesea variaza ca forma, dimensiuni, numar si localizare. In mod
normal se gasesc posterior de lobii tiroidieni, cuprinse in capsula glandei tiroide si deci apare riscul lezarii lor in timpul operatiei pe glanda tiroida. Sunt in numar de 4 (doua superioare si doua inferioare).
Celulele secretorii sunt organizate in cordoane, gramezi sau foliculi mici. Sunt bogat vascularizate. Parenchimul cuprinde doua tipuri de celule:
· celulele principale· celulele oxifile (au afinitate pentru colorantii acizi)
S-a elaborat teoria conform careia aceste celule sunt stadii diferite de dezvolare a aceleiasi celule.
Celulele principaleAu forma poligonala si talie mica. Citoplasma este palida, vacuolara uneori. Cuprinde
incluziuni de glicogen, lipide ce depind de activitatea celulei: daca celula este inactiva sunt mai multe incluziuni care reprezinta substratul energetic. La ME se observa, ca la orice celula ce sintetizeaza si secreta proteine, granulatii mici si mari care au un continut identic (spre deosebire de tirocite) atat la periferie cat si la centru, fiind forme de stocare a produsului de sinteza. Prin fixare clasica si colorare cu HE celulele principale apar ca o populatie heterogena de celule clare si intunecate. Daca insa se face fixare prin perfuzie nu mai apar aceste doua populatii, populatia fiind de fapt omogena. Deci avem de-a face cu niste artefacte in cazul fixarii clasice.
Intre foliculi, ca si la glanda tiroida, pot aparea adipocite care apar dupa pubertate si cresc in numar o data cu inaintarea in varsta astfel incat dupa varsta de 40 de ani reprezinta mai multde 50% din tesutul glandular.
Celulele oxifileAu un rol necunoscut. Sunt mai mari decat celulele principale si au doua caracteristici prin
care se deosebesc de ele:· la MO prezinta o citoplasma intens acidofila· la ME se observa ca au un continut foarte mare de mitocondrii.
Aceste celule sunt un tip aparte de celule. Ele apar doar dupa varsta de 5-7 ani, neexistand la nastere. apoi cresc numeric pana la pubertate si mai tarziu astfel incat la senescenta realizeaza ca niste formatiuni tumorale care insa nu sunt patologice.
le lipsesc granulele de secretie iar organitele de sinteza sunt slab reprezentate. Deci aceste celule sunt inactive d.p.d.v. endocrin. Au insa rol in desfasurarea anumitor procese metabolice datorita numarului mare de mitocondrii.
Glanda suprarenala
Sunt glande pereche, situate la polul superior al rinichiului, in grasimea perirenala, retroperitoneal. Au o capsula conjunctiva ce trimite septuri finecare nu compartimenteaza glanda in lobuli. Glanda are doua componente diferite d.p.d.v. embriologic, structural si functional. Corticosuprarenala (CSR) se gaseste la periferie si are origine mezodermala in timp ce modulosuprarenala (MSR) se gaseste in centru si are origine endodermala, din crestele neurale.
Corticosuprarenala (CSR)Este o componenta vitala, prin efectele hormonilor sai. Celulele se organizeaza in diverse
modalitati. Este foarte bogat vascularizata. CSR are trei zone (de la capsula spre centru):
113

1. zona glomerulara2. zona fascculata3. zona reticulata
Raportul intre cele trie zone difera in functie de varsta individului si in ffunctie de conditiile de viata ale individului (stress sau liniste):· in viata intrauterina si la nastere nu exista zona glomerulara iar zona reticulata este bine
reprezentata· pana la pubertate apare zona glomerulara iar zona reticulata diminua. In acesta perioada domina
zona fasciculata.· la adult domina tot zona fasciculata dar zonele glomerulara si reticulata sunt mai bine dezvoltate.· la senescenta are loc o transformare regresiva a CSR, involuand zonele glomerulara si reticulata.· in conditii de stress cea mai dezvoltata este zona fasciculata pentru ca secreta cortizol. In aceste
perioade are loc o transformare progresiva: zonele reticulata si glomerulara se pliaza iar zona fasciculata creste.Zona glomerularaCelulele secretori sunt celule mici, cu citoplasma acidofila, cu rare picaaturi lipidice. Ele se
organizeaza fie in cercuri fie in gramezi sau cercuri rasucite cu capilare intre ele. Rolul lor este secretia de mineralocorticoizi, dintre care cei mai importanti sunt aldosteronul si dezoxicorticosteronul.
Zona gloemrulara este o zona discontinua, fapt care este o caracteristica de specie. De asemenea, aceasta zona nu depinde de ACTH, activitatea celulelor sale fiind controlata in special de sistemul renina-angiotensina. Dovezi in acest sens au fosta aduse prin hippofizectomii sau administrare de substante care inhiba secretia de ACTH, care au dus la atrofierea zonelor fasciculata si reticulata, zona glomerulara ramanand nemodificata.
Zona fasciaulataZona fasciculata apare pe preparat ca fiind zona cea mai palida, alcatuita din cordoane de
celule paralele care sunt separate prin capilare sangvine. In mod obisnuit, aceste cordoane sunt formate din doua siruri de celule paralele. Celulele sunt mai mari decat celulele din zona glomerulara, au citoplasma usor bazofila si un continut mare de picaturi lipidice care nu se vad in coloratie HE si deci aspectul lor este vacuolar, de unde si denumirea de spongiocite. Celulele au un aspect spongios, buretos, cu gaurele. Rolul acestor celule este de a secreta glucocorticoizi dintre care fac parte cortizolul si hormonii androgeni.
Zona reticulataApare pe preparat de MO ca fiind zona cea mai intunecata. Celulele sunt organizate in
cordoane scurte ce se anastomozeaza intre ele si formeaza un reticul (retea ) in ochiurile careia se afla capilarele sangvine. Celulele sunt asemanatoare ca talie cu celulele zonei glomerulare, fiind mai mici decat cele din zona fasciculata. Ele au ciroplasma intunecata, acidofila, continand cantitati mari de lipofuscina. Rolul lor este secretia de hormoni androgeni slabi si putini glucocorticoizi. Ele secreta DHEA (dehidroepiandrosteron) si androsteron.
La ME, indiferent de zona in care se afla, celula are aceleasi caaracteristici, cele ale unei celule secretoare de steroizi: continut mare de mitocondrii cu criste tubuloveziculare, mult REN. De asemenea, nu exista granule de secretie deoarece acesste celule au o secretie continua. Descarcarea produsilor de secretie are loc la nivelul membranei care ia contact cu capilarul sangvin prin difuziune.
Medulosuprarenala (MSR)Este derivata din crestelee neurale. Ea apartine sistemului neuroendocrin difuz. Ca extindere,
MSR ocupa 8-10% din volumul suprarenalei. Ea face parte din paraganglioni, fiind un paraganglion
114

simpatic. Paraganglionul este o mica glanda endocrina care se asociaza cu SNV. Aceste formatiuni sunt formate din celule secretorii (cromafine) care sintetizeaza si secreta peptide si amine. Paraganglionii pot fi simpatici si parasimpatici. MSR este un paraganglion simpatic.
MSR este un traductor neuroendocrin, ca si pineala. Dupa stimularea nervoasa specifica, prin intermediul fibrelor preganglionare celulele sunt stimulate si sintetizeaza si secreta catecolamine.
Parenchimul MSR este organizat in gramezi sau in cordoane care se ramifica si formeaza o retea cu ochiuri largi in care se gasesc capilare si venule postcapilare. Parenchimul cuprinde doua tipuri de celule. Cea mai mare parte sunt celule cromafine (feocromocite) iar restul sunt celule ganglionare. Toate celulele sunt neuroni simpatici postganglionari dar celulele ganglionare mai prezinta prelugiri nervoase, fiind niste neuroni simpatici postganglionari, in timp ce feocromocitele nu mai au prelungiri.
Celulele cromafine dau reactia cromafina: continutul granulelor de secretie, in prezenta unui agent oxidant, se oxideaza si polimerizeaza dand nastere unui compus colorat brun numit adrenocrom. Celulele cromafine au activitate de sinteza si prin urmare au citoplasma bazofila si aparatul de sinteza bine dezvoltat. De asemenea, in citoplasma exista si multe granule de secretie care contin in mod obligatoriu catecolamine si, pe langa acestea este posibil sa existe si cromogranine (care tin catecolaminele in vezicule), Ca2+ si ATP din citoplasma, encefaline (substante opioide) si dopamin-b-hidroxilaza.
Exista doua tipuri de celule cromafine: adrenergice (>95%) si noradrenergice (putine). Aceste doua tipuri se deosebesc prin imunocitochimie, prin ME, diferentele existand la nivelule granulelor de secretie. La celulele adrenergice, granulele de secretie au un continut electronodens care ocupa toata granula de secretie. La celulele noradrenergice, continutul electronodens nu ocupa toata granula de secretie ci este situat excentric.
OVARUL
115

Este un organ cu functie mixta (endo- si exocrina). Este un organ pereche situat in pelvis, inapoia ligamentelor largi. El nu este invelit de peritoneu, care se prinde doar la nivelul hilului ovarului unde continua epiteliul de acoperire de pe suprafata ovarului.
Forma, dimensiunile si greutatea difera in functie de varsta femeii. In perioada fertila ovarul are 6-8 g, lungime 3 cm, latime 2 cm si grosime 1 cm.
In structura ovarului intre 4 componente importante:1. epiteliul de acoperire2. albugineea3. corticala4. medulara
Epiteliul de acoperireSe mai numeste si epiteliul germinativ Waldayer. Se numeste epiteliu germinativ pentru ca
s-a considerat ca la nivelul lui isi au originea celulele germinative primordiale care dau nastere ovogoniilor. Dar, de fapt, originea lor nu este la nivelul epiteliului ovarului ci la nivelul epiteliului celomic, in tesut extragonadal.
Forma epiteliului variaza in functie de varsta:· pana la pubertate epiteliul este simplu cilindric· de la pubertate pana la menopauza este simplu cubic· la menopauza este simplu pavimentos.
Aceste celule nu sunt sensibile la actiunea hormonilor ovarieni, deci aspectul lor morfologic nu se modifica in functie de fazele ciclulul ovarian.
Ca aspect, au citoplasma putina iar la ME s-au obsrvat numeroase formatiuni veziculare cu mucina, aglomerate catre polul bazal si multe granule dense la fluxul de electroni. Initial s-a emis ipoteza ca aceste granule sunt lizozomi care isi permeabilizeaza membrana cu 2-3 ore inainte de ovulatie si isi elibereaza enzimele lizozomale la nivelul albugineei, iar aceste enzime vor slabi peretele folicular al foliculului matur de Graaf, in zona in care acesta intra in contact cu albugineea. Ulterior, dupa perfectionarea tehnicilor citoenzimatice, s-a constata ca aceste granule nu sunt lizozomi deoarece ele dau rezultat negativ la reactia Gömöri de evidentiere a fosfatazei acide.
Forma nucleului poate varia, in functei de forma celulei: celula cilindrica are nucleu alungit, celula cubica are nucleu rotund iar celula scuamoasa are nucleu turtit, paralel cu membrana bazala. La ME s-a observat ca nucleul prezinta numeroase indentatii, care au tendinta sa fragmenteze nucleul in mai multi lobi, fapt pentru care celula seamana cu un PMN, de unde a aparut si ipoteza ca celula din epiteliul de acoperire al ovarului ar avea rol in apararea locala a folicului matur de agentii patogeni din peritoneu.
La polul apical al celulei se evidentiaza o retea de microvili si printre ei cativa cili. Printre celulele epiteliului de acoperire se evidentiaza insule celulare care pot sa degenereze
chistic, formand microchiste ce se pot detasa de suprafata ovarului, formand microchisturi paraovariene, care sunt anormale dar nu au semnificatie patologica. Ele pot ramane ca atare sau pot sa degenereze calcar, formand concretiuni calcare.
AlbugineeaEste formata din tesut conjunctiv dens neorientat si incapsuleaza aproape complet organul, cu
exceptia hilului, fiind responsabila de culoarea albicioasa a ovarului. Nici ea nu sufera influente din partea hormonilor ovarieni, fapt pentru care nu isi modifica in timpul ciclului ovarian.
Uneori ea poate suferi ingrosari pasagere care pot fi responsabile de ciclurile anovulatorii si poate degenera chistic. Cand albugineea se ingroasa excesiv, apare sindromul ovarelor de portelan - Stein-Löwental care nu vor ovula. Este necesara rezectia albugineei pentru vindecare.
116

Albugineea are un rol foarte important in ovulatie. La zona de contact intre foliculul matur si albuginee, cu cateva ore inainte de ovulatie, se formeaza stigma. Stigma reprezinta locul unde se va rupe albugineea. La acest nivel scade fluxul sangvin, deoarece vasele isi schimba cursul, iar zona devine palida si transparenta si se poate vedea vezicula cu foliculul matur, prin celioscopie, cu 2-3 ore inainte de ovulatie.
Medulara ovaruluiSe situeaza in centrul ovarului, inconjurata aproape complet de corticala, cu exceptia hilului.Este formata din tesut conjunctiv lax, bogat vascularizat, in care se gasesc:
· aa. helicine, care au un traseu spiralizat si care pe sectiune pot fi surprinse de mai multe ori· vene de calibru mare care formeaza plexuri· vase limfatice· fascicule de fibre musculare netede· fibre nervoase· celule ganglinare· celule deciduale· celule interstitiale Berger· rete ovari (vestigiu embrionar ce apare uneori) care reprezinta canalul Wolf si care se prezinta ca
niste formatiuni tubulare acoperite de epiteliu simplu cubic.
Corticala ovaruluiEste componenta cea mai dezvoltata a ovarului. Ea variaza ca dezvoltare in functie de varsta
femeii. La pubertate are un maxim de dezvoltare, se gaseste un numar maxim de foliculi ovarieni. Corticala scade apoi in grosime spre menopauza, iar la menopauza apare ca un tesut conjunctiv fibros fara parenchim.
D.p.d.v. structural, corticala are doua componente:· stroma conjunctiva· formatiunile parenchimatoase
Stroma conjunctivaEste formata din tesut conjunctiv lax cu functii speciale: tesut conjunctiv lax spinocelular in
care predomina fibroblastele, in principal cele de forma fusiforma, care sunt organizate in fascicule ce formeaza vartejuri in jurul vaselor de sange si in jurul formatiunilor parenchimatoase. De asemenea, exista putina substanta fundamentale si putine fibre de reticulina.
Fibroblastele au particularitati structurale si functionale diferite de fibroblastele cu alte localizari. Aceste fibroblaste sunt foarte sensibile la actiunea hormonilor ovarieni. Prin urmare:
· unele fibroblaste pot capata caractere de celula endocrina si vor edifica teaca interna foliculara care va produce hormoni androgeni: androstendionul.
· alte fibroblaste se vor transforma in miofibroblaste sau chiar fibre musculare netede si vor forma teaca externa foliculara.
· alte fibrobaste pot edifica in stroma conjunctiva glanda interstitiala a ovarului care secreta hormoni ovarieni: estrogen si progesteron. Insa acest lucru nu este posibil la om.
Formatiunile parenchimatoaseAcestea sunt:
· foliculii ovarieni· corpul galben· corpii albicans
117

Foliculii ovarieni pot fi si ei de mai multe feluri:· evolutivi:
· primordiali· primari:
· unilamelari· multilamelari, preantrali sau plini
· secundari (veziculari) dintre care unii pot fi antrali· matur (de Graaf) - antrali
· involutivi (atretici)
OvogenezaCelulele germinative primordiale iau nastere in L1 de viata intrauterina din epiteliul celomic.
Acestea se vor divide intens mitotic la acest nivel iar apoi vor migra spre crestele genitale. In crestele genitale o parte din celulele germinative primordiale se vor transforma in ovogonii
prin mitoza. Apoi aceste celule migreaza in corticala viitorului ovar unde sufera diviziuni mitotice (de 4, 5 ori) care au loc pana in L5 de viata intrauterina cand exista aproximativ 3 milioane de ovogonii pentru fiecare ovar (deci 6 milioane de ovogonii in total).
In paralel, din L3, unele ovogonii trec in ovocite de ordinul I, proces care se definitiveaza in L7. Ovocitul I intra imediat in prima diviziune de maturatie si se opreste in profaza. Apoi el se inconjoara de un strat de celule epiteliale (numite celule foliculare) care vor forma, impreuna cu ovocitul aflat in meioza, foliculul primordial care va ramane in acest fel pana la pubertate.
In aceeasi perioada au loc procese de atrezie foliculara (apoptoza), in special in L5-L7 cand incepe moartea celulara. Astfel, la nastere exista doar aprox. 400.000 foliculi primordiali in ambele ovare.
In perioada pubertate procesul de ovogeneza se continua. Cu putin timp inainte de ovulatie (2-3 ore inainte) ovocitul I isi termina prima diviziune meiotica si rezulta ovocitul II si primul globul polar. Aceste celule sunt fiecare haploide. Globulul polar preia foarte putina citoplasma si este foarte mic. El involueaza si este fagocitat de macrofagele locale.
Apoi ovocitul II, aflat tot in corticala, incepe a doua diviziune de maturatie dar se opreste in metafaza si va fi expulzat asa in trompa uterina. Aici exista doua posibilitati:1. ovocitul II este fertilizat in zona infundibulara si atunci acesta isi termina a doua diviziune
meiotica din care se elimina al doilea globul polar iar celula care ramane este preovulul. Preovulul se uneste cu spermatozoidul si se reface garnitura diploida luand nastere zigotul. Zigotul, timp de 4-5 zile se divide in trompa uterina apoi el patrunde prin ostium in uter unde va nida si se va implanta, astfel incepand o sarcina uterina. Exista totusi si sarcini tubare care, in functie de localizarea lor pot evolua pana la 3 luni.
2. ovocitul II nu este fecundat. In acest caz nu se definitiveaza a doua diviziune meiotica si ovocitul II se autolizeaza (apoptoza) sau este fagocitat de macrofagele locale.
Organizarea structurala a foliculilor ovarieni· componentele de origine epiteliala:
· ovocitul I· celulele foliculare· lichidul folicular· zona pellucida· corpii Call-Exner
· membrana Slavianski ce separa elementele epiteliale de cele conjunctive· elementele de origine conjunctiva:
· tecile foliculare· vasele sangvine
118

In procesul de crestere foliculara, cele doua tipuri de elemente cresc concomitent, intre ele existand fenomene de inductie morfogenetica.
Procesul de crestere foliculara este un proces dinamic, ce poate avea loc pe toata durata vietii. Initierea acestui proces este independenta de secretia de hormoni gonadotropi hipofizari (FSH si LH). Initierea are loc si in copilarie, sarcina, ovulatie, anovulatie sau in apropierea menopauzei. Cresterea foliculara insa este strict dependenta de hormonii gonadotropi hipofizari, mai ales de FSH.
Cresterea foliculara are loc alternativ intr-un ovar si in celalalt. Ovarul care ovuleaza este de 2-3 ori mai mare decat cel aflat in repaus. Ovarul in activitate este turgescent, hiperemic, primeste un flux sangvin crescut, putand fi examinat celioscopic pentru a se vedea foliculul matur.
Foliculul primordialFoliculii primordiali sunt cei mai numerosi. La nastere exista 400.000 foliculi primordiali in
cele doua ovare. In viata intrauterina au loc fenomene de atrezie foliculara prin apoptoza. Aceste fenomene pot exista si in momentul nasterii, si in timpul sarcinii si la pubertate si la intrarea la menopauza, adica in toate cazurile in care se schimba profilul hormonal al organismului. In timpul sarcinii, corpul galben si din L5 placenta secreta hormoni ovarieni, iar la nastere aceasta secretie dispare.
Foliculii primordiali sunt formati din ovocitul I si un singur rand de celule foliculare.Ovocitul I are un diametru de 25-30 mm, un nucleu excentric, eucromatic, cu nucleol vizibil
(deoarece cromozomii nu sunt spiralizati inca). Citoplasma este usor acidofila si are un aspect fin granular. La ME se observa mitocondrii mici, RE, cateva zone Golgi, toate organitele fiind dispuse in jurul nucleului. Membrana prezinta rari microvili.
In jurul ovocitului I sunt dispuse celulele foliculare care sunt celule epiteliale turtite, cu citoplasma putina si care stabilesc intre ele jonctiuni de tip desmozom. Aceste celule sunt asezate pe MB ce separa foliculul avascular de stroma conjunctiva.
Pe parcursul cresterii foliculare creste ovoictul, cresc elementele epiteliale, cresc elementele conjunctive. La inceput creste mai mult ovocitul iar in a doua jumatate cresc mai mult celulele foliculare si tecile conjunctive. In final, ovocitul va ajunge pana la 130 mm.
Foliculul primar unilamelarFoliculul primar unilamelar este un pic mai mare decat foliculul primordial. Ovocitul are un
diametru de aprox. 50 mm. Ovocitul se incarca de citoplasma acidofila si cu granulatii (incluziuni de glicogen). La ME se observa ca organitele celulare cresc ca numar si se hipertrofiaza. Mitocondriile se disperseaza in toata citoplasma, aparatul Golgi se dispune in ectoplasma periferica si creste numarul de microvili de pe membrana.
Celulele foliculare sunt de forma cubica, sunt atasate unele de altele prin desmozomi si sunt asezate pe MB care separa foliculul de stroma conjunctiva. Foliculul primar unilamelar este si el avascular.
Foliculul primar multilamelar (plin, preantral)Ovocitul I creste foarte mult in dimensiuni. El ajunge la un diametru de 125-135 mm.
Prezinta o citoplasma abundenta, cu multe incluziuni de glicogen. Mitocondriile sunt hipertrofiate si dispuse in toata citoplasma. REN si RER este si el hipertrofiat. Aparatul Golgi este hipertrofiat si dispus in ectoplasma. Nucleul creste si el si de acum se va numi vezicula germinativa iar nucleolul se va numi pata germinativa. Membrana prezinta multi microvili.
In jurul ovocitului I apare o zona foarte acidofila numita zona pellucida. Ea este alcatuita din glicoproteine secretate de ovocit se de celulele foliculare. La ME, zona pellucida are un aspect
119

striat, striurile fiind formate de unirea microvililor ovocitului cu prelungirile celulelor foliculare din primul rand.
In afara zonei pellucida, celulele foliculare se inmultesc si se dispun pe mai multe straturi. Se constituie astfel granuloasa foliculara. Granuloasa foliculara este alcatuita din celule mari, poligonale, cu caractere morfologice de celule secretoare de hormoni steroizi. Ele contin picaturi lipidice, mitocondrii cu criste tubuloveziculare si cantitati importante de REN. Intre celulele granuloasei se stabilesc jonctiuni desmozomale si jonctiuni gap. Granuloasa foliculara nu este niciodata vascularizata.
In membrana unor celule ale granuloasei se exprima receptori pentru FSH. Numarul receptorilor creste progresiv ajungand la 1.500 receptori / celula. Dar nu toate celulele prezinta receptori pentru FSH. O data ajuns mesajul la o celula cu receptori, acesta se transmite prin intermediul jonctiunilor gap la toate celulele din granuloasa. FSH induce procesul de aromatizare a androstendionului la estriol, proces care are loc numai in celulele granuloasei cu ajutorul unei enzime, aromataza, care este activata de FSH.
In afara granuloasei foliculare incepe diferentierea tecilor. Prima teaca, interna, este formata din cateva straturi concentrice de fibroblaste fusiforme cu
caracter de celule secretorii de hormoni steroizi. Ele secreta androstendionul. Ele au receptori pentru LH care stimuleaza secretia de androstendion.
A doua teaca, externa, este alcatuita din miofibroblaste. Ea contine numeroase vase de sange, in special capilare, care penetreaza si teaca interna, fara sa o depaseasca si formeaza plexuri capilare in jurul folicului.
In aceasta perioada se diferentiza si membrana Slavianski care in acest stadiu este discontinua. Ea este de fapt MB a epiteliului folicular devenit acum granuloasa foliculara.
La MO, limita dintre teci este greu vizibila, limita intre teaca interna si stroma este de asemenea dificila dar limita dintre teaca interna si granuloasa este usor vizibila.
Cand granuloasa foliculara ajunge la 6-12 randuri celulare, ea incepe sa secrete estrogeni. Androstendionul, precursor al hormonilor estrogeni, este sintetizat in celulele tecii interne. El treverseaza membrana Slavianski si patrunde in celulele granuloasei foliculare. La acest nivel este activata aromataza in prealabil, de catre FSH. Daca activarea aromatazei de catre FSH nu a avut loc, androstendionul insusi va activa aromataza, insa doara atunci cand este in cantitati mici. Prin activarea aromatazei, androstendionul este transformat in estriol.
Daca exista insa cantitati mari de androstendion, acesta va inhiba aromataza, activarea ei facandu-se doar de catre cantitati mici de androgeni. Prin inhibarea aromatazei se acumuleaza 5’androgeni care nu mai pot fi transformati in estriol si foliculul devine atretic. Succesul unui folicul de a se matura depinde de capacitatea lui de a converti mediul androgenic in mediu estrogenic. Din 20 de foliculi care intra in procesul de maturare la fiecare ciclu ovarian, unul singur ajunge la maturare.
Foliculul primar multilamelar secreta estrogeni si lichid folicular. Ambele sunt secretate de catre granuloasa foliculara. Lichidul folicular se va acumula intre celulele granuloasei, se disociaza granuloasa, si se formeaza niste vacuole, cavitati cu lichid folicular, acidofile in jurul carora celulele foliculare se dispun radiar. Acestea, impreuna cu vacuola de lichid folicular, alcatuiesc corpii Call-Exner. Pe masura ce lichidul folicular se secreta, cavitatile cresc, unele vor conflua, devin din ce in ce mai mari, iar prin confluenta tuturor vacuolelor se formeaza o cavitate unica, antrul folicular. In acest stadiu avem de a face cu in folicul secundar, vezicular, antral.
Foliculul secundar (vezicular, antral)Granuloasa foliculara prezinta caracteristicile descrise mai inainte. Ovocitul I are un diametru de 130 mm, nucleu este vezicula germinativa iar nucleolul este
pata germinativa. Ovocitul I prezinta toate celelalte caracteristici enumerate mai inainte. In jurul
120

ovocitului I se afla zona pellucida. Pe masura ce se formeaza antrul folicular se va forma si corona radiata, care va ramane legata de granuloasa foliculara prin discul proliger. Cumulus oophorus este o formatiune formata din ovocit, impreuna cu zona pellucida, corona radiata, granuloasa foliculara si discul proliger.
Lichidul folicular contine: estrogeni, androgeni, FSH, LH si putin progesteron (apare inainte de ovulatie). De asemenea, el mai contine glicozaminoglicani si proteine de reglare a sintezei de hormoni steroizi.
Granuloasa foliculara este separata de teaca interna prin membrana Slavanski. La exterior se gasesc teaca externa si teaca interna foliculara.
Foliculul matur de GraafDintre foliculii anteriori, la fiecare ciclu ovarian se selecteaza un singur folicul care se va
matura. Acest folicul este cel mai mare dintre toti foliculii. El are un diametru de 5 mm. El are o structura polara: la un pol se afla ovocitul iar la celalalt pol se afla antrul folicular.
Ovocitul este cuprins in cumulus oophorus, printre celulele granuloasei foliculare. Ovocitul are un diametru de 130 mm, este inconjurat de zona pellucida si de corona radiata si are toate caracteristicile morfologice ale ovocitului din stadiul anterior.
Granuloasa foliculara se subtiaza, ajungand la 5-6 randuri de celule, deoarece inceteaza diviziunile celulare si, prin cresterea cantitatii de lichid folicular si implicit cresterea antrului folicular, are loc alunecarea celulelor granuloasei foliculare unele pe langa altele si astfel are loc micsorarea numarului de straturi.
Este prezenta membrana Slavianski si teaca interna si cea externa.Intr-o etapa urmatoare, foliculul migreaza aproape de suprafata ovarului, ia contact cu
albugineea si se formeaza stigma, proces care are loc atunci cand ovocitul impreuna cu zona pellucida si corona radiata se desprind de granuloasa foliculara, cu 2-3 ore inainte de ovulatie.
Ovulatia reprezinta procesul de eliminare a ovocitului II, in fiecare luna, alternanad de la un ovar la altul, in trompa uterina.
Pot avea loc si ovulatii spontane. Ele pot fi produse de medicamente precum Clomifen, de stress, poluare, anticonceptionale oprite brusc. In caz ca intr-o zi femeia a uitat sa ia anticonceptionalele, a doua zi ea trebuie sa ia doza dubla.
Ovulatia are loc in ziua a 14-a a ciclului ovarian. Dupa ovulatie nu mai pot avea loc ovulatii spontane, deoarece apare corpul galben care secreta progesteron care impiedica maturarea unui alt folicul.
La ovulatie, la nivelul ovarului au loc descarcari de histamina, vasopresina si acumulari de progesteron in lichidul folicular. Progesteronul va activa plasminogenul care se va transforma in plasmina care va duce la permeabilizarea membranei lizozmale. Aceasta permeabilizare va permite eliberarea colagenazei, enzima foarte importanta in slabirea peretelui folicular.
Concomitent, din interiorul foliculului se exercita o presiune datorita contractiei miofibroblastelor (care are loc sub actiunea histaminei) si care, impreuna cu actiunea colagenazei, va duce la ruperea foliculului la zona de contact cu albugineea. Ovocitul II va fi captat de trompa uterina.
Ceilalti foliculi, din totalul de 18-20 foliculi ce incep procesul de maturare la fiecare ciclu ovarian, vor involua prin atrezie foliculara. Atrezia foliculara se face diferit in functie de tipul de folicul:
· pentru foliculii mici (primordiali, primari unilamelari si primari multilamelari mici) aceasta se face in doua etape:· are loc involutia ovocitului pana dispare· are loc apoi degenerescenta granuloasei pana dispare si ea complet
121

· pentru foliculii mari (foliculi primordiali multilamelari cu mai mult de 6 randuri ale granuloasei si foliculi secundari), involutia cuprinde mai multe etape:· involutia pana la disparitie a ovocitului I· in locul lasat liber de disparitia ovocitului apare zona pellucida care se ingroasa si se
pliaza.· in afara zonei pellucida, granuloasa degenereaza, se detaseaza de MB, iar celulele dispar
asincron. Prin urmare exista spatii libere in granuloasa degenerata.· teaca interna se hipertrofiaza. Unele din celulele tecii interne se pot detasa de la nivelul
foliculului si vor forma in stroma conjunctiva insule ce reprezinta glandele interstitiale ale ovarului ce secreta doar hormoni androgeni.
Structura corpului galbenStructurile epiteliale si conjunctive ramase in corticala ovarului dupa expulzarea ovocitului se
organieaza intr-o formatiune cu rol endocrin numita corp galben (luteinic), denumire ce se datoreaza pigmentului ce se acumuleaza in celulele sale si care este de culoare galbena.
La ovulatie se expulzeaza ovocitul si lichidul folicular. Prin urmare, peretii colabeaza, se pliaza, si dau un aspect policiclic, cu multe indentatii.
Evolutia corpului galben cuprinde 3 perioade importante:1. perioada de luteinizare2. perioada de vascularizatie3. perioada de regresie
Perioada de luteinizzareIn momentul in care foliculul matur a expulzat ovocitul, se deschid vasele de sange care fac
sangele sa extravazeze si va patrunde in antrul folicular. Sangele se va coagula si se va forma un cheag. Cheagul va fi invadat de tesut conjunctiv si se va forma un cheag invadat de tesut conjunctiv.
Granuloasa foliculara si si teaca interna sufera procese de luteinizare. Granuloasa luteinica va reprezenta 80% din parenchimul corpului galben iar celulele tecale luteinice vor reprezenta 20% din parenchimul corpului galben. Diferentele intre cele doua tipuri de celule luteinice constau in aceea ca celulele granuloase sunt de 2,3 ori mai mari decat celelalte, avand un diametru de 50-60 mm fata de 15-20 mm cat au celulele tecale. De asemenea, citoplasma celulelor granuloase este mai palida, in timp ce citoplasma celulelor tecale este mai densa, mai intens acidofila.
Luteinizarea este un proces ce cuprinde mai multe etape. Mai intai are loc hipertrofierea celulelelor (cresc in dimensiuni), apoi are loc hipertrofierea organitelor, aparand multe mitocondrii cu criste tubulo-veziculare, REN, RER, aparat Golgi si numeroase picaturi lipidice. Apoi celulele se incarca cu un pigment de culoare galbena (luteina).
Perioada de vascularizatieVasele de sange de la nivelul tecii interne patrund in granuloasa luteinica si vor apare lacuri
sangvine in granuloasa luteinica.
Perioada de regresieRegresia se face diferentiat, in functie de aparitia sau nu a unei sarcini.Daca nu apre o sarcina, corpul galben se va numi corp galben menstrual, catamoenial si va
incepe sa involueze in ziua a 10-14-a de la formare.Daca apare o sarcina, corpul galben va exista in corticala ovarului pana in L6, cand va incepe
sa involueze dar nu va disparea de tot. El are dimensiunile de 2, 3 ori mai mari decat un folicul matur si secreta estrogeni, progesteron (care este miorelaxant) si relaxina (care se secreta doar in sarcina si are rol de a facilita imbibitia apoasa a articulatiilor bazinului, in special a simfizei pubiene). Corpul galben de sarcina involueaza dinspre cheagul din centru catre granuloasa luteinica
122

si teaca, inlocuind celulele granuloasei si celulele tecale cu bandelete de tesut conjunctiv bogat in fibre de colagen. Inainte de inlocuire, nucleii celulelor luteinice devin picnotici si se incarca in exces cu pigment luteinic.
Treptat se va forma in locul corpului galben, corpul albicans (alb de la fibrele de colagen) care apoi sufera o degenerescenta hialina si in final rezulta o structura amorfa care nu fixeaza nici un coloarant (nu are tinctorialitate). El formeaza o cicatrice la suprafata ovarului.
Ciclul ovarianAre o durata de 28 de zile.
· faza foliculinica (din ziua 0 pana in ziua a 14-a) - are loc cresterea foliculara sub actiunea FSH.· ovulatia - are loc in ziua a 14-a sub actiunea LH.· faza luteinica- are loc formarea corpului galben care va secreta estrogeni si progesteron.
ORGANELE LIMFATICE
Sistemul imun este alcatuit din celule si structuri distribuite la nivelul intregului organism cu rol de aparare impotriva agresorilor exteriori: bacterii, paraiti, fungi, virusuri dar si fata de structurile proprii ce au suferit modificari.
Sistemul imun poate face deosebirea intre doua categorii de structuri, molecule, fragmente antigenice: self si nonself. Nonself poate fi considerata orice structura (molecula, celula) capabile de a fi recunoscute de sistemul imun si impotriva carora organismulelaboreaza un raspuns imun (ex.: o celula tumorala provine dintr-o celula normala a organismului si capata caracteristici ce vor fi recunoscute de sistemul imunitar.
Imunogenele sunt substante nonself capabile de a produce raspuns imun.
123

Antigenele = orice molecula ce poate fi recunoscuta specific de anticorpi. Ex.: bacteriile sunt imunogene pentru ca determina un raspuns imun. Ele prezinta mai multe antigene - proteinele de suprafata.
Sistemul imun este alcatuit din celule si structuri. Celulele sunt:· limfocitele· celulele ce apartin sistemului fagocitar mononuclear· granulocitele· celulele prezentatoare de antigen (Antigen Presenting Cells - APC)
Celulele sistemului imun sunt raspandite in intreg organismul: in organe, la nivelul tesutului conjunctiv (plasmocitele) sau sunt grupate in tesutul limfoid.
Tesutul limfoid este o varietate de tesut conjunctiv alcatuit dintr-o stroma cu fibre si celulel reticulare care alcatuiesc o retea in ochiurile careia se afla limfocite, macrofage si alte APC. La randul lui, tesutul limfoid alcatuieste cea mai mare parte a organelor limfatice sau se gaseste asociat mucoaselor tracturilor digestiv, respirator sau genito-urinar. Acest tesut se numeste tesut limfoid asociat mucoaselor (Mucosa Associated Lymphid Tissue - MALT). Deci tesutul limfoid este:
· la nivelul organelor limfoide· asociat mucoaselor
Organele limfatice sunt organe parenchimatoase alcatuite predominant din tesut limfoid. De aceea aceste organe se mai numesc si tesut limfoid incapsulat.
Limfocitele T si BIsi au originea in celula stem din maduva rosie hematogena. Caile de maturare ale celor doua
tipuri de limfocite sunt diferite:· precursorii limfocitelor T parasesc maduva si populeaza in perioada prenatala timusul. Aici are
loc maturarea iar limfocitele parasesc organul ca limfocite T mature (imunocompetente).· limfocitelel B se matureaza in maduva rosie hematogena pe care o parasesc ca limfocite B
mature.Maduva rosie hematogena si timusul sunt organe limfoide primare (centrale) denumite astfel
deoarece la acest nivel limfocitele se matureaza fara a veni in contact cu antigene nonself. Restul organelor limfoide sunt secundare sau preiferice (splina, ganglionii limfatici si tesutul limfatic asociat mucoaselor). Aceastea se numesc astfel pentru ca la acest nivel limfocitele vin in contact cu anatigenele nonself, se activeaza, prolifereaza si se diferentiaza.
Structuri ale MALT:· amigdalele· placile Peyer de la nivelul ileonului· apendicele ileocecal
Maduva rosie hematogena este atat organ limfoid primar cat si secundar (aici exista si limfocite B, T si plasmocite mature).
Organele limfatice secundare sunt sediul de desfasurare a raspunsului imun (primele etape ale raspunsului imun au loc la acest nivel - recunoasterea si diferentierea).
TimusulEste situat in medistiul anterior. Este organ limfoid primar. Are o structura limfoepiteliala
spre deosebire de celelalte organe limfoide. El are dubla origine embriologica: exista un primordiu de origine epiteliala care este invadat de celule mezenchimale. Stroma are deci origine epiteliala iar capsula este de origine conjunctiva. Capsula prezinta septuri care impart organul in lobuli.
Un lobul timic are doua regiuni:· periferica - corticala
124

· centrala - medularaCorticala, in coloratie HE, are aspect intunecat (inchis la culoare) iar medulara are aspect
clar. Corticala are aspect intunecat datorita prezentei unui numar mare de celule tinere, precursoare ale limfoctelor T numite timocite. Timocitele au nuclei rotunzi, tahicromatici, mici. Ele prolifereaza intens. Din tote timocitele corticale (5% mor prin apoptoza. Celelalte 5% isi continua maturarea apoi migreaza pana ajung in regiunea medulara.
Regiunea edulara are o structura caracteristica numita corpusculul Hassal. Acestia sunt alcatuiti din celule de natura epiteliala deoarece contin in citoplasma filamente de cheratina. Celulele sunt dispuse in structuri concentrice. Au nuclei picnotici, intens tahicromatici. Corpii Hassal au o functie necunoscuta. Se presupune ca ar secreta factori umorali timici.
La nivelul lobulului timic exita timocite, precursori ai limfocitelor T insa nu exista precursori ai limfocitelor B sau ai limfocitelor killer. Timocitele sufera un proces de maturare. Celulele mai tinere se gasesc in regiunea subcapsulara si de aici, concomitent cu maturarea, ele migreza spre medulara. La nivelul timusului, procesul de maturare a limfocitelor T are doua etape:1. selectia pozitiva2. selectia negativa
Selectia pozitiva are ca rezultat supravietuirea acelor celule ce sunt capabile de recunoasterea prin intermediul receptorilor pentru antigen propriile antigene HLA (CMH) ale organismului.
Selectia negativa permite supravietuirea acelor timocite ce nu reactioneaza impotriva propriilor antigene ale organismului.
CMH este de doua feluri CMH I si CMH II. Alte celule din corticala lobulului timic:
· reticuloepiteliale - au forma stelata si nuclei ovalari. Ele au prelungiri care se anastomozeaza, jonctiunile realizandu-se prin desmozomi si formand astfel o retea in ochiurile careia exista timocite. Ele intervin in selectia pozitiva.
· celula doica - se gasesc in regiunea subcapsulara, au natura epiteliala si rol in maturarea limfocitelor T. Ele secreta ca si celulel reticuloepiteliale hormoni timici cu rol important in dezvoltarea si maturarea limfocitelor T:· timulina· timozina· timopoietina
· celulele dendritice interdigitale - se gasesc la limita dintre corticala si medulara. Au origine in maduva rosie hematogena si rol in selectia negativa.
· macrofage - atat in corticala cat si in medulara. Ele au rol de APC si sunt importante in procesul de selectie. Ele au si functia de a fagocita timocitele moarte prin apoptoza.Vascularizatia:Arterele sunt ramuri pentru septurile conjunctive si dau ramuri pentru parenchim la nivelul
jonctiunii cortico-medulare. Acestea sunt ramuri care vascularizeaza corticala, dupa care se arcuiesc si vascularizeaza si medulara. In medulara este originea circulatiei venoase a timusului. Nu are circulatie limfatica.
Circulatia timica are si o particularitate: la nivelul corticalei exista bariera sange-timus. Aceasta este alcatuita din endoteliul nefenestrat al capilarelor care este insotit de o lamina bazala groasa dar si din prelungiri ale celulelor reticulo epiteliale ce inconjura capilarele. Ea impiedica antigenele nonself sa ia contact cu timocitele corticale.
Timusul involueaza la varsta adulta, dupa pubertate. cea mai afectata este regiunea corticala unde timocitele sunt inlocuite cu adipocite albe - are loc degenerescenta grasoasa. Dar timusul pune limfocite T in circulatie chiar si la varsta adulte, mai putin insa.
Limfocitele care parasesc timusul sau maduva rosie se numesc limfocite naive pentru ca nu au venit in contact cu antigenul.
125

La nivelul organelor limfoide secundare are loc initierea raspunsului imun impotriva antigenelor venite pe cale limfatica sau sangvina de la nivelul tesuturilor. Splina asigura apararea impotriva antigenelor din sange, ganglionii limfatici asigura apararea imptriva antigenelor din circulatia limfatica iasr MALT asigura apararea la nivelul mucoaselor.
Ganglionii limfaticiSe gasesc raspanditi in tot organismul. Sunt o statie de filtrare a limfei, aflandu-se pe traseul
vaselor limfatice. Sunt organe incapsulate, avand o capsula conlucntiva care trimite septuri (trabecule) ce nu compartimenteaza complet organul.
Un ganglion are o fata convexa si un hil. La nivelul fetei convexe intra vasele aferente caare ies pe la nivelul hilului sub forma unui singur vas eferent. La nivelul hilului se afla si arterele si venele ce vascularizeaza ganglionul.
Ganglionul limfatic are o regiune periferica (corticala) si una centrala (medulara). Intre cele doua regiuni exista zona paracorticala. Corticala contine tesut limfoid organizat in foliculi limfoizi. Acesti foliculi limfoizi sunt formatiuni rotund-ovalare cu aspect caracteristic. La nivelul lor exista limfocite T in diferite stadii de diferentiere.
Foliculii pot fi primari (areactivi) si secundari. Foliculii primari sunt omogeni si mai inchisi la culoare. Foliculii secundari prezinta doua regiuni: una periferica, mai inchisa la culoare care se numeste manta si una centrala, mai palida, care se numeste centru germinativ. De asemenea, la nivelul centrului germinativ exista doua zone: una intunecata si una clara. Zona intunecata este alcatuita din centroblaste, care au dimensiuni mai mari, citoplasma bazofila si nuclei palizi, eucromatici. In zona clara se gasesc centrocite, care au un aspect caracteristic, cu nuclei clivati.
Foliculii limfoizi se gasesc in toate organele limfoide secundare. Foliculii limfoizi conti in majoritate limfocite B. In zona paracorticala se gasesc predominant
lifocite T. In medulara, celulele se organizeaza in cordoane ce delimiteaza sinusurile medulare.Circulatia sangelui si limfei din ganglionLimfa patrunde prin vasele aferente care prezinta valve ce asigura curgerea unidirectionala
catre sinusul subcapsular ® sinusurile intermediare (strabat corticala) ® sinusurile medulare si iese din ganglion prin vasul eferent, prin hil.
Circulatia sangelui se face printr-o artera care patrunde prin hil, da ramuri pentru septuri care vor da ramuri pentru foliculi. Sangele se aduna apoi in venule care conflueaza si formeaza venele de la nivelul septurilor, ce formeaza prin unire venele ce parasesc ganglionul pe la nivelul hilului.
La nivelul venulelor postcapilare, celulele endoteliale au o particularitate: au forma inalta, sunt cubice sau chiar cilindrice. Ele se numesc venule cu endoteliu inalt (HEV - high endothelium venules)
Alte tipuri de celule in ganglionul limfatic:· in foliculi: celule dendritice foliculare - APC· regiunea paracorticala - celule interdigitate· in sinusuri, in cordoanelel celulare ale medularei si la nivelul tesutului limfoid propriu-zis din
corticala exista si macrofage.· in sinusuri sunt mai putine limfocite decat in restul parendchimului si prin urmare aici pot fi
observate mai usor celulele reticulate prin MO. Rolul ganglionilor limfatici: produc doua tipuri de limfocite B efectorii:
1. plasmocite2. limfocite B cu memorie
Splina
126

Splina este cel mai mare organ limfoid secundar din organism. Prezinta o capsula in care exista fibre musculare netede si din care pornesc septuri ce nu compartimenteaza organul. Parenchimul splinei se numeste pulpa. Macroscopic, daca sectionam orgaanul fara sa il fixam, el apare rosu cu puncte albe. Acestea reprezinta pulpa alba iar partea rosie se numeste pulpa rosie.
Circulatia la nivelul splineiArterele dau ramuri trabeculare ce dau la randul lor ramuri ce se distribuie parenchimului.
Aceste ramuri se inconjoara de tesut limfoid si formeaza arterele centrale ale pulpei albe. Din acestea se desprind ramuri - arteriolele foliculilor limfoizi din pulpa alba splenica. Ramurile terminale ale a. centrale sunt arteriolele penicilate (au forma de pensula - scurte si drepte) care conduc sangele spre sinusurile venoase ale pulpei rosii.
Exista doua ipoteze privind trecerea sangelui din arteriolele penicilate in sinusurile venoase. Prima sustine ca exista continuitate intre arteriolele penicilate si sinusurile venoase, circulatia fiind de tip inchis si sangele neparasind arborele circulator. A doua ipoteza sustine ca arteriolele penicilate se termina printre cordoanele de celule ale pulpei rosii. Sangele strabate spatiul dintre aceste celule pana la capilarele sinusoide ale pulpei rosii. Aceaasta circulatie este deschisa deoarece sangele paraseste arborele circulator. Din sinusoidele pulpei rosii iau nastere venele pulpei rosii care se unesc si formeaza venele trabeculare ce nu prezinta un perete muscular propriu. Acestea vor forma in final venele splenice ce parasesc splina prin hil.
Splina nu are circulatie limfatica.Pulpa alba a splinei este alcatuita din:
· mansonul de tesut limfoid care inconjoara arteriola si se numeste teaca limfoida periarteriala. Mansonul contine predominant limfocite T.
· foliculii limfoizi splenici care sunt alcatuiti din manta si centru germinativ, Ei semana cu foliculii secundari insa se asociaza cu o arteriola pozitionata excentric, acest ansamblu (folicul si arteriola) alcatuind corpusculul Malpighi. Foliculii contin prediominant limfocite B.La limita dintre pulpa alba si pulpa rosie exista zona marginala ce contine sinusuri, numite
sinusuri marginale. Aici se deschid ramurile arteriolelor foliculare si ramuri din artera centrala a pulpei albe. Aceasta zona este importanta d.p.d.v. functional deoarece este locul prin care patrund anumite tipuri de limfocite circulante iar antigenele vin pe cale sangvina.
Pulpa rosie este alcatuita din cordoane celulare ce delimiteaza sinusurile venoase. Sinusurile pulpei rosii au in structura lor celulel epiteliale cu propritati contractile. Aceste celule au forma alungita cu axul lung paralel cu axul sinusului iar in jurul sinusului sunt dispuse circular fobre de reticulina. Apar estfel niste spatii prin care patrund elementele figurate lae sangelui.
Tipuri de celule:In foliculii limfoizi se gasesc celulel foliculare dendritice, celule interdigitate, macrofage si
limfocite B. In mansonul de tesut limfoid se gasesc de asemenea celule foliculare dendritice, celule interdigitate si macrofage dar exista limfocite T in loc de B.
Siusurile marginale contin macrofage distincte: imobile, cu capacitate crescuta de a fagocita antigene aduse prin sange pana la nivelul sinusurilor marginale. Apoi aceste macrofage prezinta antigenul limfocitelor B.
Cordoanele celulare al pulpei rosii (cordoanele Billroth) sunt alcatuitte dintr-o stroma, un citoreticul, in ochiurile caruia exista multe limfocite B, limfocite T, macrofage, plasmocite ca si celelalte elemente figurate ale angelui. Macrofagele de aici sunt cele ce fagociteaza eritrocitele imbatranite.
Tesutul limfoid asociat mucoaselor (MALT - Mucosa Associated Lymphoid Tissue)Este alcatuit din zone de tesut limfatic: foliculi secundari agregati, situati in mucoasa si
subnucoasa tracturilor digestiv, respirator sau urogenital. Fac parte din MALT:· amigdalele
127

· placile Peyer· apendicele
MALT cuprinde majoritatea tesutului limfoid din organism.La acest nivel se produc IgA (care sunt cele mai multe). ACeste Ig au rol in apararea zonei
luminale.AmigdaleleAmigdalele sunt:
· amigdala palatina· amigdala linguala· amigdala faringiana
Toate alcatuiesc inelul limfatic Waldayer.Amigdala palatina se gaseste in peretele lateral al orofaringelui.
TUBUL DIGESTIV
Sistemul digestiv este format din tub digestiv si glande anexe.
Structura generala a tubului digestiv
Tubul digestiv, cu exceptia cavitatii bucofaringiene, este alcatuit din 4 tunici:1. mucoasa2. submucoasa3. musculara4. tunica externa
Mucoasa
128

Are trei componente:1. epiteliul de acoperire2. corion3. musculara mucoasei
Epiteliul de acoperireAre doua aspecte morfologice, in functie de segmentul de tub digestiv in care se afla. El
poate fi:· epiteliu pluristratificat pavimentos nekeratinizat· epiteliu simplu cilindric
Tranzitia intre cele doua zone se face de obicei brusc dar uneori se poate face si treptat.
CorionulEste format din tesut conjunctiv lax ce cuprinde:
· celule imunocompetente, semn ca aici exista o intensa activitate imunologica· capilare sangvine si limfatice, ce reprezinta locul unde vor trece moleculele absorbite atunci cand
se realizeaza absorbtia.· terminatii nervoase vegetative cu rol de control a activitatii de la acest nivel· glande - foarte dese cu exceptia esofagului
Musculara mucoaseiEste formata din tesut muscular neted ce are aceeasi organizare ca la tunica musculara: un
strat circular intern si unul longitudinal extern. De la acest nivel se desprind benzi musculare spre corion care se grupeaza in jurul glandelor si au rol de a ajuta la eliminarea produsilor de secretie.
Exista modificari ale mucoasei in diverse segmente:· evaginari· invaginari
Evaginarile pot sa intereseze doar mucoasa si atunci se numesc vilozitati sau pot sa antreneze si submucoasa si atunci se numesc pliuri. Scopul este de a mari suprafata de absorbtie.
Invaginarile mucoasei spre corion, fara sa depaseasca musculara mucoasei - alcatuiesc glandele mucoase, iar daca depasesc musculara mucoasei si ajung pana in submucoasa se numesc glande submucoase. Acestea au rol de a mari suprafata de secretie a glandelor. Invaginarile care strabat tot peretele tubului digestiv sunt canalele glandelor anexe.
Cele mai multe modificari, de-a lungul tubului digestiv, apar la nivelul epiteliului de acoperire (la diverse etaje ale tubului). Musculara mucoasei asigura miscari independente ale mucoasei si reprezinta si o bariera ce impiedica migrarea celulelor. Acest lucru este foarte important in conditii patologice deoarece musculara mucoasei impiedica migrarea celulelor neoplazice.
SubmucoasaEste alcatuita din tesut conjunctiv moderat dens, iar comparativ cu corionul are mai multe
fibre de colagen, fibre elastice si mai putine celule. De asemenea submucoasa mai contine:· vase limfatice si sangvine mari· tesut limfoid (difuz sau nodular)· plezul nervos Meissner· glande (doar la nivelul esofagului si duodenului).
MuscularaCu exceptia partii superioare a esofagului si a partii terminale a canalului anal, unde exista
fibre musculare striate, in rest musculara contine fibre musculare netede. Ele se organizeaza in doua
129

straturi: circular intern si longitudinal extern. De fapt, ambele straturi au dispozitie helicoidala, insa stratul circular are buclele orizontale si foarte stranse, in timp ce stratul longitudinal are buclele oblice, aproape verticale, si cu pasul larg. Intre cele doua paturi exista tesut conjunctiv lax cu vase de sange si plexul nervos Auerbach.
Stratul circular prezinta din loc in loc ingrosari - sfinctere. Prin contractia stratului intern are loc comprimarea si amestecarea continutului digestiv, iar prin contractia paturii externe se scurteaza segmentul de tub digestiv si, prin intermediul undelor peristaltice, are loc deplasarea continutului digestiv.
Tunica externaDupa caz, poate fi adventice sau seroasa.Adventicea este formata din tesut conjunctiv lax care adera si ancoreaza segmentul de tub
digestiv de organele din jur: esofagul toracic, rectul si celelalte segmente ale tubului digestiv care adera la peretele abdominal posterior (duodenul, colonul ascendent si descendent).
Tunica externa din restul tubului digestiv este seroasa, fiind alcatuita din tesut conjunctiv lax dar care este acoperit de mezoteliul ce reprezinta foita viscerala a peritoneului.
Tunica externa contine vase sangvine si nervi.
Tesutul limfoid asociat mucoasei tubului digestiv
Este de doua feluri:1. tesut limfoid difuz - localizat de obicei in mucoasa2. tesut limfoid nodular, localizat in mucoasa cu posibilatea de a se extinde si in submucoasa
In tesutul limfoid difuz exista numeroase limfocite, macrofage, plasmocite (in special cele care secreta IgA), eozinofile si mastocite. Acest tesut este sediul major in activitatea imunologica la nivel local.
Nodulii limfatici pot fie solitari, fie sub forma de agregate nodulare. Nodulii solitari se gasesc in esofag, la nivelul jonctiunii esogastrice si in corionul din jurul canalelor de secretie ale esofagului. De asemenea ei se mai gasesc in stomac, in regiunea pilorica si in general la orice nivel al intestinului subtire si gros. Agregatele nodulare se gasesc la nivelul ileonului si al apendicelui vermiform, sub forma placilor Peyer.
In tesutul limfoid difuz predomina limfocitele T. In tesutul limfoid nodular avem atat limfocite T cat si B.
In initierea raspunsului imun un rol important il are celula M (M de la microfald). Ea se localizeaza la nivelul epiteliului tubului digestiv, aflandu-se in raport cu nodulii limfatici ce fac parte din agregatele nodulare si care se gasesc la nivelul apendicelui si ileonului.
In ME de baleiaj, celula M prezinta la suprafata microfalduri si microvili care sunt mai lungi si mai grosi dar mai putini decat la nivelul polului apical al enterocitelor. Corpul celulei este scobit de catre limfocite si de pseudopodele macrofagelor.
Celule M are rol in initierea raspunsului imun local. Celula M endociteaza antigenul si il elibereaza apoi in zona in care detecteaza prezenta limfocitelor si macrofagelor. Macrofagele vor fagocita antigenul si il vor prezenta apoi pe suprafata. Limfocitele vor citi antigenul prezentat si vor sintetiza anticorpi. Acestia vor migra prin circulatia sangvina pana la nivelul corionului unde vor stimula limfocitele B din tesutul limfoid difuz sa secreta noi cantitati, mult mai mari, de anticorpi IgA. Enterocitele, la polul bazal, au receptori pentru IgA si deci moleculele de IgA vor traversa enterocitul si se vor fixa pe versantul extracitoplasmatic al moleculei proteinei J care se afla la polul apical al enterocitului. Fixarea la nivelul glicocalixului de proteina J a IgA se face pentru a proteja molecula de anticorp de actiunea proteazelor intestinale. Se tapeteaza astfel mucoasa intestinala cu molecule de IgA si ea va fi aparata de agresiunea antigenului.
130

Inervatia tubului digestiv
Exista fibre nervoase de la nivelul sistemului nervos central si fibre nervoase locale, ce intra in componenta plexurilor intramurale de la toate nivelele tubului digestiv, de la esofag pana la sfarsitul canalului anal. La acest nivel se gasesc foarte multi neuroni, aproape tot atati de multi cati se gasesc la nivelul maduvei spinarii: 100 mil. de neuroni. Acesti neuroni au caractere functionale asemanatoare cu cele ale neuronilor din nevrax: exista neuroni care pot prelua informatia din mediu si neuroni care pot elabora comanda. De aceea, aceste plexuri sunt denumite “creierul tubului digestiv”. Initial, aceste fibre nervoase au fost considerate partea terminala a inervatiei eferente a tubului digestiv.
Inervatia simpatica a tubului digestiv se face de la nivelul maduvei spinarii, iar inervatia parasimpatica se face prin parasimpaticul cranian (prin n. vag) si prin parasimpaticul sacrat (S2-S4). Distributia eferentei vegetative: fibrele simpatice preganglionare fac sinaspsa in ganglioni vegetativi paraviscerali iar fibrele parasimpatice fac sinapsa in ganglionii intramurali.
Plexurile nervoase locale contin mai multe elemente:· mici ganglioni nervosi care contin un numar variabil de neuroni (3-50)· fibre nervoase amielinice ce se grupeaza intr-o retea in jurul ganglionilor intramurali
parasimpatici si care sunt conectate si cu neuronii din ganglionii intramurali· celule gliale, care sunt mai multe decat neuronii si sunt asemanatoare cu astrocitele din nevrax.
O parte din celulele gliale secreta interleukine si pot exprima la suprafata membranei moleculele CMH II. Aceste celule gliale au rol in desfasurarea raspunsului imun.Rolurile plexurilor intramurale:
· asigura controlul activitatii motorii a tubului digestiv· asigura controlul secretiei exo si endocrine digestive· controleaza microcirculatia sangvina· intervin in raspunsul imun si in raspunsul inflamator.
Se considera ca plexurile intramurale sunt o parte a nevraxului migrata la nivelul tubului digestiv. Astfel se explica existenta neuronilor motori si senzitivi care pot inchide arcuri reflexe scurte. Aceste plexuri nu sunt total independente, fapt dovedit de legatura cu neuronii din nevrax. Prin urmare controlul local este influentat de nevrax prin conexiunile sale cu plexurile.
Neuronii din ganglionii intramurali sunt de trei feluri: senzitivi, motori si interneuroni. Prelungirile neuronilor senzitivi pot forma aferentele arcurilor reflexe, interneuronii asigura legatura cu motoneuronii iar neuronii motori exercita influente inhibitorii sau excitatorii asupra elementelor componente ale tubului digestiv. Cand exista un stimul se inchid doua tipuri de arcuri reflexe: un arc lung, ce implica si neuronii din nevrax si un arc reflex scurt, ce se realizeaza la nivel local.
Neuronii motori isi pot exercita efectul in doua moduri: fie axonii lor ajung direct pe celula musculara si ii modifica activitatea, fie axonul sau se opreste pe o celula intermediara unde poate avea efect sau nu. Aceste celule intermediare pot fi celule endocrine, celule ale sistemului imun (mastocite) sau celule interstitiale Cajal care sunt niste celule dotate si cu capacitate de pace-maker.
De obicei, fibrele parsimpatice sunt conectate cu neuronii excitatori iar fibrele simpatice sunt conectate cu neuronii secretori.
Plexul Meissner contine fibre pentru celulele secretorii din mucoasa digestiva, controleaza activitatea muscularei mucoasei si activitatea fibrelor musculare de la nivelul vaselor sangvine din submucoasa. Plexul Auerbach controleaza activitatea motorie a tubului digestiv controland tunica musculara. Rareori fibrele plexului Auerbach pot ajunge si in submucoasa.
Afectarea acestor plexuri nervoase conduce la tulburari de motilitate a tubului digestiv. Dintre aceste tulburari citam:
· achalazia
131

· megacolonul congenital: boala Hirschprung.In ambele cazuri are loc distrugerea selectiva a neuronilor motori inhibitori din plexurile
intramurale la nivelul esofagului in achalazie si la nivelul colonului distal si rectului in boala Hirschprung.
In achalasie nu are loc relaxarea musculaturii esofagului si a sfincterului cardia si prin urmare nu se face pasajul din esofag in stomac. Au loc spasme continui la acest nivel, fapt care duce la hipertrofierea musculaturii esofagului. Achalazia se manifesta prin varsaturi de alimente nedigerate.
In boala Hirschprung au loc spasme ale colonului distal si ale rectului si astfel se acumuleaza gaze in colonul proximal. Pacientul va avea abdomenul destins, dureri colice si constipatie care poate dura uneori de la cateva zile pana la cateva saptamani.
Tratamentul in ambele cazuri este dificil.
Esofagul
La nivelul toracelui se prezinta ca un tub rectiliniu cu lungimea de 25 cm. Este acoperit de adventitie. Dupa ce strabate diafragma devine esofag abdominal care are o lungime de 2-4 cm si care este acoperit de seroasa, fiind fixat la peretele posterior al abdomenului.
MucoasaEste alcatuita dintr-un epiteliu pluristratificat pavimentos nekeratinizat care are mai multe
straturi celulare:· celulele bazale, care se dispun in 1-3 randuri si care au citoplasma bazofila si prezinta numeroase
mitoze.· celulele poliedrice - unele au un continut bogat in glicogen.· celulele scuamoase. Unele prezinta putine granule de keratohialina in citoplasma. Ele nu sunt
keratinizate in mod normal, dar in cazul unor traume mecanice provocate de bolul alimentar aceste zone se keratinizeaza.Mai pot exista si alte tipuri de celule:
· 25% din indivizi prezinta in epiteliu celule endocrine ce apartin sistemului neuroendocrin difuz. In acest caz exista riscul crescut de aparitie a carcinoamelor primare cu celule mici, endocrine.
· 4-8% din indivizi pot contine in epiteliul bazal melanocite. Si in acest caz exista riscul de aparitie a melanoamelor primare la nivelul esofagului.
· celule Langerhans care se gasesc in stratul bazal si sunt niste celule prezentatoare de antigen.Tranzitia de la epiteliul pluristratificat pavimentos la cel simplu cilindric de la nivelul
stomacului se face brusc, dar deasupra jonctiunii esogastrice cu 0,5-1,5 cm. Deci 0,5-1,5 cm din esofag vor fi captusiti de epiteliu simplu cilindric. De aceea, cand se produc refluxuri gastroesofagiene repetate apar leziuni la nivelul epiteliului simplu cilindric (ulceratii, inflamatii etc.).
Cand, in timpul vietii intrauterine, epiteliul simplu cilindric al esofagului nu se transforma complet in epiteliu pluristratificat pavimentos apar zone ale esofagului cu epiteliu simplu cilindric. Acest esofag se numeste esofag Barret, la care incidenta carcinoamelor este mult crescuta.
Corionul mucoasei formeaza la nivelul epiteliului pluristratificat pavimentos papile conjunctive inalte. Acest corion este mai sarac celular comparativ cu cel de la nivelul altor organe. La nivelul corionului pot aprea glandele esofagiene superficiale, care se gasesc doar la nivelul extremitatilor esofagului (inferioara si superioara). Ele sunt glande simple, tubulare ramificate si secreta un mucus neutru. Sunt deci glande mucoase.
132

Musculara mucoasei, in 2/3 superioare ale esofagului contine numai fibre cu orientare longitudinala, doar in 1/3 inferioara aparand si stratul circular intern.
SubmucoasaIn submucoasa creste numarul fibrelor elastice ce asigura distensia elastica a esofagului la
trecerea bolului alimentar. Are un continut ridicat de vase sangvine mari, sistemul venos fiind foarte bine dezvoltat, acesta fiind sediul uneia din anastomozele portocave.
Glandele de la nivelul submucoasei sunt glandele esofagiene propriu-zise. Morfologic difera de glandele esofagiene superficiale, fiind simple, tubuloacinoase ramificate. D.p.d.v. functional sunt glande mucoase si secreta un mucus acid. Canalul lor de excretie se deschide la suprafata epiteliului, la varsare fiind mai larg (cisterna Schaffer). Ocazional acest orificiu se poate obstrua.
Ocazional, in structura glandelor se pot gasi si alte celule: · celule secretoare de lizozim· celule pesinogene· peste varsta de 50 de ani pot aparea oncocitele (celule mari - oncos = mare). Nu se cunoaste
functia lor. Ele sunt celule epiteliale alterate. Au citoplasma acidofila si granulata. Contin de asemenea si multe mitocondrii, si ele alterate.
MuscularaEste alcatuita din tesut muscular neted si striat. In treimea superioara a esofagului se gasesc
exclusiv fibre musculare striate, in treimea mijlocie se gasesc si fibre musculare striate si netede iar in treimea inferioara se gasesc numai fibre musculare netede.
Ca la toate nivelele tubului digestiv, si la nivelul esofagului musculara este impartita in doua straturi: unul circular intern si unul longitudinal extern.
Stomacul
Stomacul prezinta mai multe regiuni anatomice:· regiunea cardiala· regiunea fundica· regiunea corpului gastric· regiunea antrului piloric· regiunea canalului piloric
MuscularaLa nivelul stomacului se afla cea mai groasa zona a tunicii musculare. Fibrele musculare
netede sunt situate in trei straturi: · intern - oblic· mijlociu - circular· extern - longitudinal
Fibrele musculare oblice se gasesc doar la nivelul cardiei si corpului gastric.Fibrele circulare sunt dispuse intr-un strat continuu, in toate regiunile stomacului. Ele
formeaza sfincterele.Fibrele longitudinale sunt intr-un strat discontiinuu si se concentraza de-a lungul celor doua
curburi ale stomacului.
MucoasaMucoasa prezinta numeroase pliuri ce dispar la distensia stomacului. De asemenea, suprafata este brazdata de numeroase santuri putin adanci ce delimiteaza ariile
gastrice cu dimensiuni de 1-4 mm. La nivelul unei arii se deschid la suprafata epiteliului mai
133

multe milioane (3,5 mil.) de invaginari ale epiteliului gastric ce se numesc foveole gastrice = cripte gastrice. Acestea nu sunt glande ci la nivelul fundului lor se deschid canalele de secretie ale glandelor gastrice.
Epiteliul de acoperire este epiteliu simplu cilindric, alcatuit dintr-un singur tip de celule: celulele mucoase de suprafata care secreta mucus. Aceasta este o celula cillindrica, cu un nucleu oval, situat spre polul bazal, polul apical al celulei fiind clar care contine granule de secretie (cu mucus). Acelasi epiteliu exista si in cripte, numai ca exista mai putine granule de secretie si mai mici. La ME, aceste granule sunt dense la fluxul de electroni. Rolul acestor celule mucoase este de a secreta un mucus neutru. O parte din acest mucus se varsa in cavitatea gastrica si intra in componenta sucului gastric iar alta parte are rol de protectie impotriva acidului clorhidric.
Corionul permite diferentierea celor trei regiuni histologice ale stomacului:· regiunea cardiala - contine cripte putin adanci (1/4 din grosimea corionului) si glande lungi,
sinuoase, cu lumen larg. Mai multe glande se deschid in aceeasi cripta.· regiunea corpului si fundului stomacului - contine cripte lungi (1/2 din grosimea corionului) si
glande mai scurte, sinuoase si cu lumen larg.· regiunea pilorica (antrul si canalul piloric) - contine cripte scurte si glande lungi, drepte, cu
lumen stramt, care se pot bifurca la capete.Glandele cardiale si pilorice contin in principal celule mucoase. Rareori mai pot contine si:
· celule parietale (oxintice)· celule care secreta lizozim· celule nediferentiate· celule endocrine ce apartin sistemului neuroendocrin difuz si care se gasesc numai in glandele
pilorice.Glandele de la nivelul fundului si corpului gastric sunt glandele gastrice propriu-zise. Ele
sintetizeaza HCl si enzimele digestive. Ele contin urmatoarele tipuri celulare:· celule mucoase ale colului· celule parietale (oxintice)· celule principale (pepsinogene)· celule nediferentiate· celule ce apartin sistemului neuroendocrin difuz.
Celulele mucoase ale coluluiColul glandelor gastrice reprezinta portiunea din canalul de excretie al glandei aflata in
apropierea fundului criptei gastrice. Glandele gastrice mai au un corp si o baza.Celulele mucoase ale colului se afla in regiunea colului si a corpului glandei gastrice. Ele
seamana cu celulele mucoase de suprafata doar ca ele sunt mai mici, au nucleul rotund, au mai multe organite si sunt mai putin dense la fluxul de electroni. Ele au rolul de a secreta mucus acid care va crea mediul acid necesar activarii enzimelor digestive din sucul gastric.
134

Stomacul (continuare)
Tipuri de celule ce alcatuiesc glandele gastrice propriu-zise:
Celula parietala (oxintica)Celulele parietale (oxintice) apartin glandelor gastrice propriu-zise si se gasesc pe tot
cuprinsul lor dar se aglomereaza insa in partea superioara a lor, in zona colului si corpului glandei.
D.p.d.v. morfologic aceste celule sunt sferice sau piramidale, au un nucleu mare, situat central dar este posibil uneori ca aceste celule sa fie binucleate.
In MO se observa citoplasma intens acidofila iar la ME se observa ca polul bazal intra in raport cu MB a epiteliului glandular iar polul apical nu ajunge pana la suprafata epiteliului glandular. Dar polul apical comunica cu lumenul. De asemenea se observa continutul mare in organite cu membrane netede: mitocondrii, REN. Celula oxintica prezinta si un sistem canalicular intracelular si un sistem tubulovezicular.
135

Sistemul canalicular intracelular se formeaza prin invaginarea membranei apicale care se face spre polul bazal, uneori chiar pana in apropierea lui. La nivelul acestor canalicule exista microvili. Aceste canalicule comunica cu lumenul, fiind invaginari ale membranei apicale.
Sistemul tubulovezicular se gaseste in apropierea sistemului canalicular intracelular si este reprezentat de un sistem de membrane netede. Reprezinta fragmente de plasmalema invaginata. Acest sistem tubulovezicular este considerat un rezervor de membrana care poate fi transferata sistemului canalicular intracelular cand se sintetizeaza HCl (pentru a se forma microvilii).
Deci aspectul ultrastructural al celulei parietale variaza in functie de starea de activitate a celulei: in repaus sistemul canalicular intracelular este slab dezvoltat si are putini microvili iar sistemul tubulovezicular este abundent; in momentul intrarii in activitate, sistemul canalicular se dezvolta brusc, ajungand pana la polul bazal si prezinta multi microvili in timp ce sistemul tubulovezicular se reduce.
Rolul celulei oxintice este in sinteza si secretia de HCl si factor intrinsec Castle. Factorul intrinsec este o glicoproteina cu greutate moleculara mare care asigura absorbtia vitaminei B12 la nivelul ileonului. In caz de aclorhidrie, care se datoreaza absentei celulelor parietale, urmeaza absenta absorbtiei vitaminei B12 care este urmata de aparitia anemiei pernicioase Biermer, vitamina B12 fiind implicata in maturarea eritroblastelor.
Sinteza HCl are loc la nivelul sistemul canalicular. Aici exista un continut crescut de anhidraza carbonica si se genereaza protonii. La nivelul polului bazal are loc preluarea din capilarele sangvine a CO2 care se va combina cu H2O sub actiunea anhidrazei carbonice si se va forma H2CO3 care va disocia rapid in H+ si HCO3
-. Ionul bicarbonic se va reintoarce in mediul intern iar H+ va fi expulzat la polul apical prin pompa de H+/K+ care introduce un ion K+ si elimina in lumen un H+. Clorul este preluat din sange si este transportat activ in lumenul sistemului canalicular intracitoplasmatic. HCl se va forma la suprafata membranei sistemului canalicular.
Helycobacter pylori este o bacterie ce este prezenta in mucusul din stomacul a doua treimi din populatia globului. Aceasta bacterie este toxica pentru ca produce enzima ureaza care descompune ureea si formeaza NH3. Amoniacul va neutraliza HCl si astfel va scadea aciditatea gastrica. Prin urmare se stimuleaza secretia de HCl a celulelor paritale. Cand aceasta stimulare se cronicizeaza, creste numarul de celule oxintice pana la de 2, 3 ori si se favorizeaza aparitia gastritelor si aulcerului gastric.
Celulele principale (pepsinogene)Sunt situate de-a lungul intregii glande gastrice propriu-zise dar predomina in partea bazala.
Ele au caracteristicile unor celule ce sintetizeaza proteine de export. La MO apar ca niste celule inalte, piramidale, cu nucleul oval sau rotund, situat la polul
bazal. La polul bazal au citoplasma bazofila iar la polul apical au citoplasma acidofila. Partea acidofila de la nivelul polului apical poate lipsi atunci cand in urma prepararii se pierd granulele de secretie (prin fixare).
La ME se observa ca celula principala are contact cu MB a epiteliului glandular iar polul apical ajunge pana la suprafata epiteliului. Organitele sunt reprezentate de RER situat in zona bazala si laterobazala, mitocondrii, aflate printre cisternele RER, aparat Golgi situat supranuclear si granule de secretie aflate la polul apical.
Aceste celule au mai multe roluri:· sintetizeaza si secreta pepsinogen, proenzima proteazica ce se activeaza in lumenul glandei· sintetizeaza si secreta lipaza (putina) care ajuta la digestia lipidelor· sintetizeaza si secreta renina - functie recent dovedita
136

Celulele nediferentiateSunt localizate printre celulele mucoase ale colului, in partea apicala a glandei si nu la baza
cum ar fi normal. Sunt singurele celule din componenta glandelor gastrice care prezinta mitoze, fapt care permite diferentierea acestor celule la MO.
Sunt celule columnare, cu nucleu oval si citoplasma bazofila. Sunt celulele care se pot diferentia in oricare din celulele componente ale glandei gastrice. In mod obisnuit numarul lor este redus dar ele cresc ca numar atunci cand mucoasa este agresata.
Celulele endocrine ce apartin sistemului neuroendocrin difuzSe gasesc in regiunea glandelor gastrice propriu-zise. Ele prezinta granule de secretie care se
aglomereaza la polul apical.
Intestinul subtire
Are o lungime medie de 6 m dar in timpul vietii are o lungime de numai 3-4 m datorita tonusului permanent la care este supus. El are trei segmente:1. duoden - 25 cm2. jejun 2,5 m3. ileon 3,5 m
Delimitarea intre jejun si ileon se poate face numai cu microscopul.Peretele este format din 4 tunici. Modificarile semnificative fata de celelalte segmente ale
tubului digestiv apar la nivelul mucoasei si submucoasei. Aceste modificari au in principal scopul de a mari suprafata de absorbtie si secretie. Mucoasa si submucoasa prezinta urmatoarele modificari:
Plicile circulare (valvele conivente Kerckring)Reprezinta proiectii transversale, permanente (spre deosebire de stomac), in care este
antrenata si submucoasa. Sunt vizibile cu ochiul liber, avand o inaltime de 1 cm. Ele proemina in lumen si pot ocupa pana la 2/3 din lumen.
Ele nu exista peste tot. Apar la o anumita distanta de orificiul pilori (maxim 5 cm), sunt evidente in jejun si diminua spre ileon, disparand in partea distala a ileonului (ultimul metru de ileon nu mai prezinta plici circulare).
Ele au rol de a mari suprafata de aproximativ 3 ori.
Vilozitatile intestinaleReprezinta proiectii permanente ale mucoasei. Ele exista doar in intestinul subtire si dispar in
apropierea valvulei ileocecale.Vilozitatile difera intre ele ca forma si ca numar / unitatea de suprafata.Ca forma - lungimea vilozitatilor diminua de la duoden spre ileon. Ca aspect variaza in
functie de segmentul intestinului subtire si in functie de varsta individului:· La nivelul duodenului exista vilozitati largi, asemanatoare cu o frunza· In jejun vilozitatile sunt cilindrice, digitiforme· In ileon gasim vilozitati mai scurte, ca o maciuca· Numarul este mai mare in duoden si jejun (40 vilozitati / mm2) fata de ileon (10 vilozitati /
mm2).Vilozitatile au rolul de a mari suprafata mucoasei de 8 ori.Structura vilozitatii la MO: prezinta un ax conjunctiv care este o extensie din corion care este
acoperit de epiteliu simplu cilindric. El contine 2 tipuri principale de celule: celule absorbante (enterocite) si celule caliciforme. De asemenea mai exista limfocite migrate de la nivelul tesutul limfoid asociat mucoasei intestinale.
Axul conjunctiv este bogat in celule:
137

· celule conjunctive (predomina fibroblastele si miofibroblastele)· celule imunocompetente
Miofibroblastele au axul lung perpendicular pe axul longitudinal al vilozitatii, fiind orientate orizontal. Dintre celulele imunocompetente, predomina limfocitele, macrofagele, plasmocitele.
La nivelul axului conjunctiv exista o retea vasculara formata dintr-o arteriola, capilare fenestrate care se aduna in final in una sau mai multe venule. Exista si un capilar limfatic (chiliferul limfatic central) care se vede doar cand este dilatat.
De asemenea pot sa apara fibre musculare netede care se desprind din musculara mucoasei. Ele reprezinta a doua populatie de elemente contractile. Ele au axul longitudinal paralel cu axul vilozitatii. Ansamblul fibrelor musculare netede alcatuiesc m. Brucke.
La nivelul axului conjunctiv exista si fibre nervoase vegetative pentru vasele din vilozitate si pentru fibrele musculare netede. Aceste fibre provin din plexul submucos Meissner.
Vilozitatea prezinta contractii rimice cu frecventa de 6 contractii / min. Aceste contractii intervin in absorbtie, facilitandu-se trecerea susbstantelor nutritive din enterocit in capilarul sangvin sau limfatic. Contractia este asigurata de miofibroblaste care asigura ingustarea si alungirea vilozitatii si de fibrele musculare netede care determina scurtarea si latirea vilozitatii.
Exista o situatie patologica cu caracter genetic - boala celiaca sau enteropatia glutenica. Ea apare in jurul varstei de 1 an si este determinata de modificari ale vilozitatii intestinale de la nivelul jejunului. Aceste vilozitati vor fi mici si turtite, cu celule epiteliale cubice cu un numar scazut de microvili. Modificarea suprafetei de absorbtie duce la malabsorbtie care se manifesta prin diaree, scaderea in greutae, fatigabilitate, astenie pronuntata. Cauza acestei afectiuni este de natura genetica si consta in intoleranta la o fractiune proteica din gluten. Glutenul este un complex ce se gaseste in grau, secara, iar fractiunea proteica este gliadina. In cazul acestor pacienti lipseste enzima proteolitica care determina scindarea gliadinei iar gliadina nedigerata are un efect toxic ce determina modificarea celulelor epiteliale intestinale. Leziunile mucoase sunt reversibile daca se scoate din alimentatie glutenul.
MicroviliiSe gasesc la nivelul enterocitului, la polul apical. In MO microvilii de la acest nivel se
observa sub forma platoului striat si nu ca margine in perie. Microvilii reprezinta forme de organizare stabila a microfilamentelor de actina care se ancoreaza la citoplasma prin placa terminala.
Ei au rolul de a mari suprafata mucoase intestinale de 30 de ori.
Criptele LieberkuhnReprezinta invaginari simple, tubulare, ale epiteliului de acoperire ce nu depasesc musculara
mucoasei. Ele se deschid in lumen intre bazele a doua vilozitati intestinale. La nivelul lor se gasesc celule nediferentiate din care se formeaza toate celulele din mucoasa intestinala. O glanda Lieberkuhn da nastere tuturor celulelor din 3-6 vilozitati intestinale.
Suprafata totala a mucoasei intestinale este de 200 m2.Epiteliul intestinal este de tip simplu cilindric si are un turn-over crescut. Acesta variaza in
functie de specie, la om fiind de 6 zile. In 6 zile se inlocuieste tot epiteliul de suprafata si epiteliul glandelor Lieberkuhn.
Corionul mucoasei intestinale este subtire. Sub epiteliu apare o teaca formata din fibroblaste stelate care se numeste teaca subepiteliala (pericriptala). Fibroblastele stabilesc intre ele jonctiuni gap iar in citoplasma prezinta microfilamente de actina si miozina care le confera proprietati contractile. Ele au si proprietati secretorii, realizand sinteza si secretia de colagen. Exista un singur strat de colagen care se organizeaza in fibre de reticulina si proteoglicani. Acest strat este situat sub MB si ele regleaza ritmul de difuzie a moleculelor absorbite.
138

Fiind o mucoasa expusa la actiunea agentilor agresori, corionul contine o cantitate mare de tesut limfoid. Acesta poate fi nodular (in partea distala a ileonului se grupeaza in placile Peyer) sau difuz. Placile Peyer sunt situate in grosimea peretelui intestinal, in partea opusa insertiei mezenterului. In total exista 30-40 placi Peyer, fiecare cu cate 10-70 de noduli limfoizi. Nodulii din corion pot ajunge pana in submucoasa. In dreptul placii Peyer criptele Lieberkuhn sunt scurte sau lipsesc. In dreptul nodulilor, epiteliul contine celule M.
Tipuri de celule la nivelul intestinului subtire· celule de suprafata - enterocite· celule caliciforme· celule Paneth· celule ce apartin sistemului neuroendocrin difuz· celule M (doar la nivelul epiteliului din dreptul placilor Peyer)· celule nediferentiate· celule intermediare· celule migrate, sangvine - limfocite T ce se gasesc intre doua celule absorbante
EnterociteleSunt principale celule epiteliale de suprafata. Ele apar in numar mic si in criptele Lieberkuhn.La polul apical prezinta platoul striat. Au citoplasma bazofila. Nucleul este oval, situat la
polul bazal. Prezinta un numar mare de microvili (aprox. 3.000 microvili / celula) care sunt acoperiti de un glicocalix gros.
La ME se observa ca au foarte multe organite. Exista REN abundent, iar REN si lizozomii se gasesc in zona apicala, imediat sub placa terminala. Mitocondriile sunt raspandite in toata citoplasma, sunt alungite, iar in zona apicala au un ax paralel cu axul celulei. Ele au de asemenea o tendinta de aglomerare in zona bazala a celulei. RER este situat in zona bazala a celulei, aparatul Golgi este supranuclear. Prezinta putine granule de secretie care sunt mici, situate la polul apical si care contin enzime sintetizate de enterocit.
Celule sunt unite la nivelul apical prin complexe jonctionale care impiedica trecerea continutului intestinal din lumen in spatiul intercelular. Aceste complexe jontionale sunt responsabile si pentru mentinerea celor doua zone ale membranei eritrocitare. In zona bazala exista spatii intercelulare care se maresc atunci cand se transporta apa, ioni, lipide.
Rolul enterocitelor este in procesul de digestie. Ele sintetizeaza enzime care termina digestia glucidelor si proteinelor. Ele secreta aceste enzime prin polul apical dar enzimele nu ajung in lumen ci se fixeaza la glicocalix prin legaturi hidrofobe (sunt adsorbite). Enterocitele intervin in absorbtie, aici avand loc absorbtia tuturor principiilor alimentare, a ionilor si a apei.
Enterocitele sintetizeeaza trigliceride la nivelul REN din acizii grasi absorbiti. Se formeaza chilomicroni in aparatul Golgi.
Celulele caliciformeSe localizeaza in epiteliu si in cripte. Numarul lor creste dinspre duoden spre ileon. Ele
secreta mucus care protejeaa mucoasa intestinala. Ele protejeazi impotriva invaziei bacteriilor din lumen.
Celulele PanethSe gasesc doar la baza glandelor intestinale Lieberkuhn. Ele sunt mai numeroase in ileon. Ele
se mentin la baza glandei Lieberkuhn aprox. 20 de zile.La MO se observa o polaritate morfologica, citoplasma fiind bazofila la polul bazal si
acidofila la polul apical unde exista granule de secretie cu proteine bazice cu arginina.La ME apar ca niste celule producatoare de proteine de export. Ele prezinta si multi
lizozomi. Aceste celule au mai multe roluri:
139

· secreta lizozim, substanta ce distruge peretele unor bacterii· secreta peptide antibacteriene - criptidine· au rol fagocitar asupra bacteriilor si protozoarelor prin care regleaza flora microbiana intestinala.
Recent s-a descoperit ca o parte din celulele Paneth contin foarte multe IgA si astfel ele coopereaza cu plasmocitele din corion.
Celulele endocrineLa nivelul tubului digestiv exista cea mai mare varietate de celule endocrine. Ele sunt
localizate la baza glandelor Lieberkuhn dar nu lipsesc nici in epiteliul vilozitatii.
Celulele stem, nediferentiateSe afla la baza glandelor Lieberkuhn. La MO se observa usor daca apar elemente de mitoza. La ME nu se diferentiaza decat daca
se foloseste o tehnica speciala - injectarea de timidina tritiata. Timidina tritiata este inglobata de celulele nediferentiate si de celulele intermediare. Dar celulele stem raman la baza glandelor Lieberkuhn timp de 3 saptamani, in timp ce celulele intermediare migreaza spre lumen.
Celulele imatureEle sunt o parte din celulele nediferentiate, care apoi se vor diferentia in celulele epiteliului.
Celulele imature migreaza mereu spre lumen, cu exceptia cazurilor in care se vor diferentia in celule Paneth.
Celulele intermediareAu aspect intermediar intre celula caliciforma si enterocit. La polul apical prezinta microvili
si cateva granule cu mucina. Ele se localizeaza in cripte, in 2/3 inferioare ale criptei. Ele se pot divide si se matureaza dand nastere celulei absorbante sau celulei caliciforme.
Ele au fost descrise mai recent si au rol important in stabilirea originii cancerului. Daca in tumora exista celule intermediare, atunci tumora are origine intestinala si nu este metastaza.
Elemente particulare pentru submucoasaGlandele din partea superioara a duodenului sunt particulare intestinului subtire. Acestea sunt
glandele Brunner. D.p.d.v. morfologic ele sunt glande tubulo-alveolare ramificate. Ele au canale excretoare care se deschid la baza glandelor Lieberkuhn sau direct in lumen (mai rar). D.p.d.v. functional sunt glande mucoase dar contin si celule de tip endocrin care produc urogastron (factorul de crestere al epiteliului - EGF). Urogastronul inhiba secretia de HCl si stimuleaza diviziunea celulara.
Diagnosticul histologic pozitiv de duoden se pune atunci cand se gasesc vilozitati si glande in submucoasa.
Intestinul gros
Este format din:· cec· apendice· colon· rect
140

El este o structura adaptata pentru absorbtia de apa si ioni si pentru secretia de cantitati mari de mucus pentru lubrifiere. El este format din 4 tunici. Nu contine valve Kerckring si nu contine nici vilozitati. Cecul si colonul au acelasi aspect histologic.
Cecul si colonulMucoasa este acoperita de epiteliu simplu cilindric cu celule absorbante si celule caliciforme.
Corionul are un continut mare de glande Lieberkuhn, mai lungi si care sunt inconjurate de o teaca subepiteliala de fibroblaste. Aceste glande au aceleasi tipuri de celule cu glandele de la nivelul intestinului subtire cu exceptia:· au mai putine celule absorbante· au mai multe celule caliciforme· au mai putine tipuri de celule endocrine· prezinta numai ocazional celule Paneth, la nivelul glandelor din cec si colonul proximal iar in
restul intestinului gros lipsesc.Corionul este bogat in tesut limfoid. Prezinta noduli limfoizi mari care se extind pana in
submucoasa.Celulele absorbante si cele caliciforme se deoasebesc de cele din intestinul subtire:
Celulele absorbanteCelulele absorbante au un platou striat mai subtire (microvilii sunt mai subtiri), au placa
terminala mai putin dezvoltata, glicocalixul nu are enzime digestive. Ele se gasesc in epiteliul de suprafata si in cripte.
Celulele absorbante sunt cele mai numeroase iar raportul celule absorbante / celule caliciforme este 4 / 1. Acest raport se modifica langa rect unde este 1 / 1. Celulele caliciforme apar insa ca fiind mai multe deoarece ele sunt mai voluminoase si comprima celulele absorbante. In cripte exista mai multe celule caliciforme decat absorbante.
Rolul celulelor absorbante este de absorbi apa si ioni si de a sintetiza glicoproteine care vor fi eliberate la nivelul glicocalixului sau in lumen. La nivelul colonului proximal se pot absorbi si principii - acizi grasi, din care se vor sintetiza trigliceride si apoi chilomicroni dar cu intensitate mai mica decat in celula absorbanta intestinala.
Celulele caliciformeSunt mai multe in intestinul gros decat in intestinul subtire si mai multe in cripte decat in
epiteliul de suprafata. In 2/3 inferiare ale criptelor Lieberkuhn se gasesc celule caliciforme in exclusivitate.
Celulele caliciforme au putine granule de secretie, deoarece mucusul se secreta permanent si in cantitati mari, deci se stocheaza in granule de secretie doar putin mucus.
SubmucoasaPrezinta 2 particularitati:
1. vascularizatia este mai bogata decat in alte segmente ale tubului digestiv2. are mai multe adipocite.
MuscularaAre doua straturi de fibre musculare netede: circular inteern si longitudinal extern. Stratul longitudinal extern are o grosime neuniforma si formeaza 3 benzi (condensari de fibre
musculare netede) numite tenii. Intre tenii, stratul muscular longitudinal este subtire, uneori chiar discontinuu. Cele trei tenii sunt mezenterica, omentala si ...................
141

La intervale regulate, din tenii se desprind benzi care se orienteaza si patrund in stratul circular intern. Aceste benzi vor delimita arii musculare care se pot contracta independent de restul musculaturii. Alaturi de aceste arii, teniile determina si niste proeminente sacciforme spre exterior numite haustre. Intre doua haustre, mucoasa sufera si ea modificari, formand plicile semilunare.
Tunica externaPrezinta multe acumulari de tesut adipos care merg de-a lungul teniilor in special. Ele lipsesc
la nivelul cecului. Aceste acumulari formeaza apendicii epiploici.
ApendiceleAre patru tunici, cea externa fiind seroasa. Mucoasa este formata dintr-un epiteliu simplu cilindric in care apar celule absorbante, celule
M si mai putine celule caliciforme. Corionul prezinta glande Lieberkuhn si tesut limfoid. Glandele Lieberkuhn sunt mai putine si uneori mai scurte. De obicei sunt grupate, cu o lungime variabila si ocazional la baza lor pot sa apara celule Paneth si celule endocrine.
Tesutul limfoid este in special nodular , iar nodulii sunt dispusi ca o coroana si ei se extind si in submucoasa, fapt pentru care musculara mucoasei nu se vede. Tesutul limfoid se organizeaza astfel dupa nastere, in jurul varstei de 10 ani. La 40 de ani acest tesut limfoid se reduce si va fi inlocuit cu tesut conjunctiv care contine in special fibre de colagen.
Musculara prezinta doua straturi: circular intern si longitudinal extern care sunt continui si nu exista tenii. Musculara este subtire iar acesta este un element ce favorizeaza trecerea infectiei de la apendice la peritoneu (apendicita este insotita adesea de peritonita).
Rectul - structura histologicaEl are doua componente:
1. segmentul superior - rectul propriu-zis - 12 cm lungime2. segmentul inferior - canalul anal - 2-3 cm lungime
Rectul continua colonul sigmoid si se situeaza intre planul vertebrei S3 si diafragma pelvina. Ampula rectala reprezinta partea sa inferioara. Canalul anal continua rectul, de la diafragma pelvina pana la orificiul anal extern.
Rectul propriu-zisEste captusit de un epiteliu simplu cilindric. Are un corion cu multe glande Lieberkuhn, mai
lungi ca la nivelul colonului si cu multi noduli limfatici. Din aceasta cauza exista la suprafata interna a mucoasei niste proeminente determinate de nodulii limfoizi.
Musculara are doua straturi, stratul longitudinal redevenind continuu.
Canalul analAre trei componente:
1. zona columnara (hemoroidala), intre diafragma pelvina si valvele anale2. zona intermediara, intre valvele anale si linia anocutanata3. zona cutanata, intre linia anocutanata si orificiul anal
Linia anocutanata reprezinta tranzitia intre epiteliul simplu cilindric si epiteliul pluristratificat pavimentos keratinizat.
Zona columnaraLa acest nivel mucoasa formeaza 5-10 pliuri longitudinale care se numesc coloane anale
(columne rectale Morgagni). Aceste pliuri sunt mai evidente in copilarie, apoi se sterg. In partea inferioara ele se unesc prin intermediul unor pliuri transversale care se numesc valve anale. In
142

spatele valvelor (pliurilor) anale exista niste depresiuni, ca niste buzunare care se numesc sinusuri anale.
Mucoasa, in partea superioara a columnelor are un epiteliu simplu cilindric iar in partea inferioara a columnelor are un epiteliu pluristratificat pavimentos nekeratinizat. In sinusuri exista de asemenea epiteliu simplu cilindric. Tranzitia intre cele doua tipuri de epitelii se face dupa o linie dintata, linia pectinata, si se poate face brusc, de la epiteliu simplu cilindric la epiteliu pluristratificat pavimentos nekeratinizat sau lent de la epiteliu simplu cilindric la epiteliu pluristratificat cilindric - epiteliu pluristratificat cubic - epiteliu pluristratificat pavimentos nekeratinizat. In final, in ambele cazuri avem epiteliu pluristratificat pavimentos nekeratinizat.
Cand s-a facut trecerea la epiteliul pluristratificat pavimentos nekeratinizat, in corion nu mai exista glande Lieberkuhn. Musculara mucoasei dispare si ea insa inainte de tranzitia intre cele doua epitelii.
In sinusurile anale, la nivelul corionului exista glande - glandele anale care pot reprezenta frecvent sediul unor inflamatii. Ele se deschid in criptele anale. Ele sunt glande mucoase, simple tubulare ramificate iar in profunzime pot ajunge in submucoasa si chiar pana in musculara.
Zona intermediaraEste acoperita de epiteliu pluristratificat pavimentos. Are un corion care contine:
· fibre elastice· corpusculi Pacini· glande sebacee ce nu sunt asociate firelor de par.
Zona terminalaSe intinde de la linia anocutanata pana la nivelul sfincterului anal intern. Este acoperita de
epiteliu pluristratificat pavimentos moderat keratinizat. La nivelul dermului din aceasta regiune se gasesc:· glande sudoripare· foliculi pilosi· glande sebacee· vene care alcatuiesc plexul hemoroidal extern
Glandele sudoripare se grupeaza si formeaza glandele circumanale, situate in jurul orificiului anal. Acestea sunt glande apocrine care isi declanseaza activitatea la pubertate, secretia lor fiind controlata de hormonii sexuali.
Tunica musculara a canalului anal cuprinde si ea doua straturi musculare: circular intern si longitudinal extern. Stratul circular intern se ingroasa pe toata lungimea canalului anal si formeaza sfincterul anal intern.
Patura musculara longitudinala este subtire si se termina in regiunea cutanata intre sfincterul intern si cel extern.
Sfincterul anal extern este format din fibre musculare striate.
143
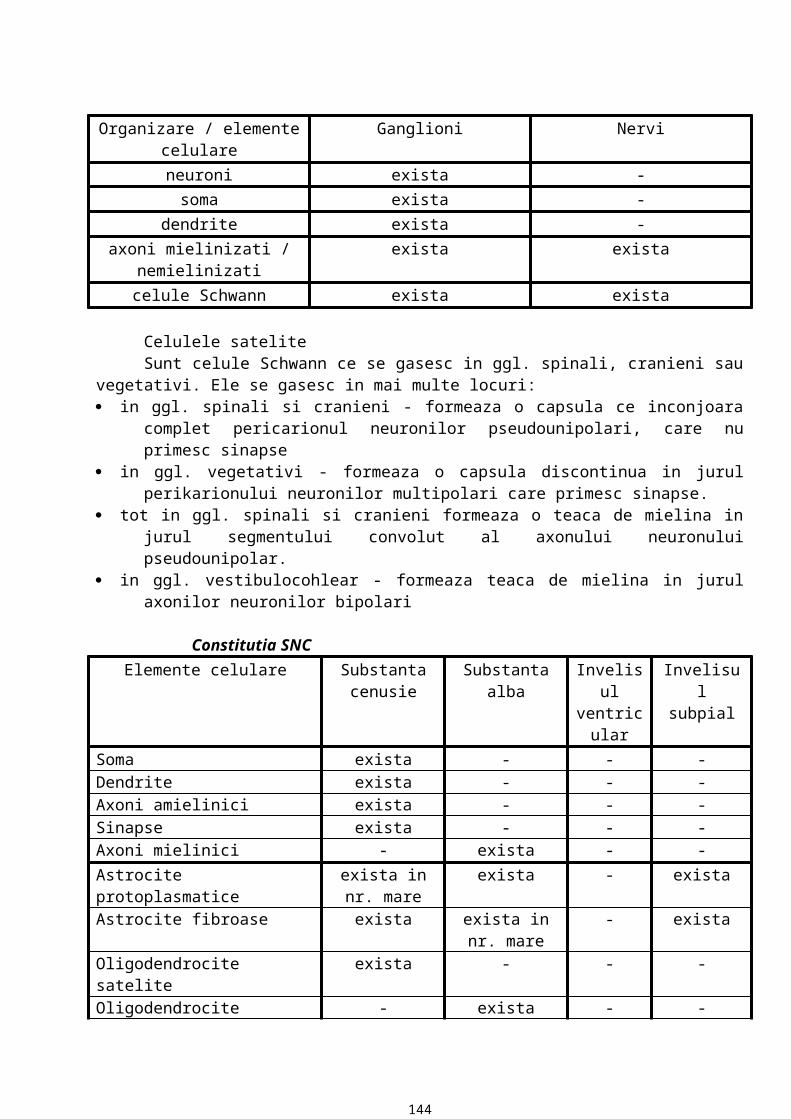
GLANDELE ANEXE
FICATUL
Este cea mai mare glanda din organism. Ea reprezinta a 50-a parte din greutatea organismului. Este un organ vital.
Ficatul este un organ parenchimatos care contine:· in majoritate exista celule de origine epiteliala - hepatocite care intra in structura lobulului
hepatic. Ele sunt dispuse sub forma de lame sau placi celulare perforate, ramificate si anastomozate. Ele vor realiza o retea tridimensionala, intre placi si intre celule existand capilarele sangvine si ductele biliare.
· tesut conjunctiv in proportie redusa.
Cu privire la structura parenchimului hepatic s-au emis mai multe ipoteze:· lobulul hepatic clasic· lobulul portal Sabourin· acinul hepatic Rappaport
144
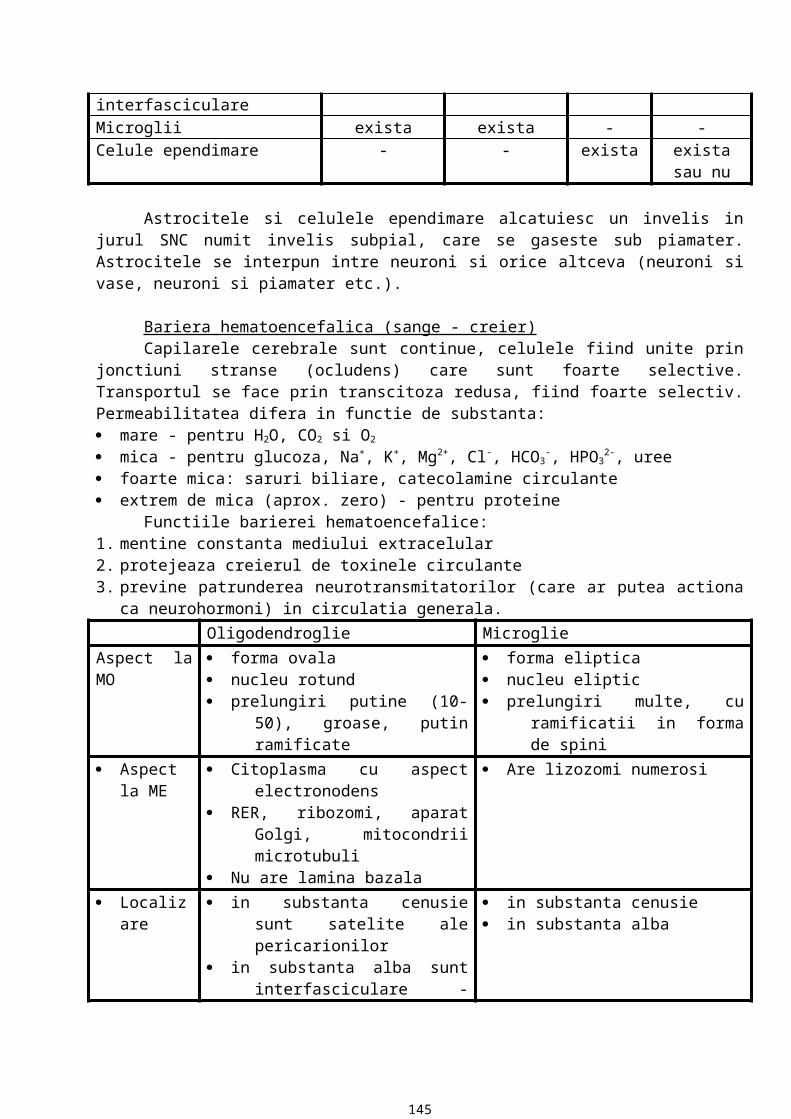
Lobulul hepatic clasicConform acestei teorii, lobulul hepatic clasic este unitatea morfo-functionala a ficatului.
Lobulul clasic reprezinta aria din parenchim a carui sange este drenat de aceeasi vena centrolobulara (venula hepatica terminala).
Lobulii sunt in numar de aproximativ 1 mil. Ei au forma poliedrica, fiind frecvent hexagonali. Lobulul are o inaltime de 2 mm si o latime de 0,7 mm. In alcatuirea lobulului intra placile de hepatocite cu dispozitie radiara si anastomozate. Intre placi exista capilarele sinusoide care sunt si ele anastomozate.
Pe o sectiune transversala, lobulul are forma de poligon, de regula cu 6 laturi. In colturile sale, la locul de vecinatate cu ceilalti lobuli exista un spatiu interlobular redus care se numeste spatiu portal Kiernan. In sectiunea transversala, placilor celulare le corespund cordoane celulare al caror aspect variaza in functie de varsta individului. La adult grosimea unui cordon este egala cu diametrul unui hepatocit cu exceptia locurilor de anastomoza.
Pe sectiune longitudinala fiecare cordon este gros de cel putin 2 randuri de celule, intre celule existand canaliculii biliari.
La periferie exista placa terminala care corespunde hepatocitelor de la periferia lobulului. Placa are grosimea unui hepatocit. La periferie, lobulul vine in raport direct cu tesutul conjunctiv din spatiile portale. Hepatocitele din placa terminal vor forma un perete aproape continuu , fiind strabatut doar de vasele sangvine si de ductele biliare.
Spatiile interlobulare, portale Kiernana contin 3 elemente care alcatuiesc triada portala:1. o vena interlobulara, ramura terminala a venei porte2. o artera interlobulara, ramura terminala a a. hepatice3. un canal biliar interlobular
Pe lang triada portala mai exista si 1 sau mai multe vase limfatice si fibre nervoase amielinice vegetative.
Lobulul portal SabourinLobulul portal repreazinta aria din parenchim a carei bila este tributara aceluiasi canalicul
biliar interlobular. El are forma de triunghi, avand in cetru spatiul portal si in varfuri trei vene centrolobulare de la trei lobuli clasici diferiti.
Acinul hepatic RappaportAcinul hepatic este aria de parenchim ce este tributara aceleiasi vascularizatii si aceluiasi
canalicul biliar. El are forma de romb, alcatuit din doua triunghiuri adiacente, unite prin marginile lor. Axul lung al rombului este dat de doua vene centrolobulare iar axul scurt este dat de 2 spatii portale. In el se descriu trei zone functionale cu dispozitie concentrica:1. in apropiera axului scurt2. spre a. hepatica3. spre vena centrolobulara
In functie de localizarea in cele trei zone, hepatocitele prezinta gradient metabolic, marime, activitatea enzimatica si continutul in organite depinzand si ele de localizarea celulei in acini.
In zona 1 se afla hepatocitele care primesc sangele cu cel mai mare procent de oxigen, fiind aproape de artera perilobulara. Aceste hepatocite au mai multe caracteristici:· sunt cele mai active metabolic· sunt primele care vor depune glicogen postprandial· sunt primele care se vor activa in hipoglicemie si vor produce glicogenoliza si gluconeogeneza· sunt primele care vor semnala aparitia semnelor de regenerare in caz de leziuni hepatice
145
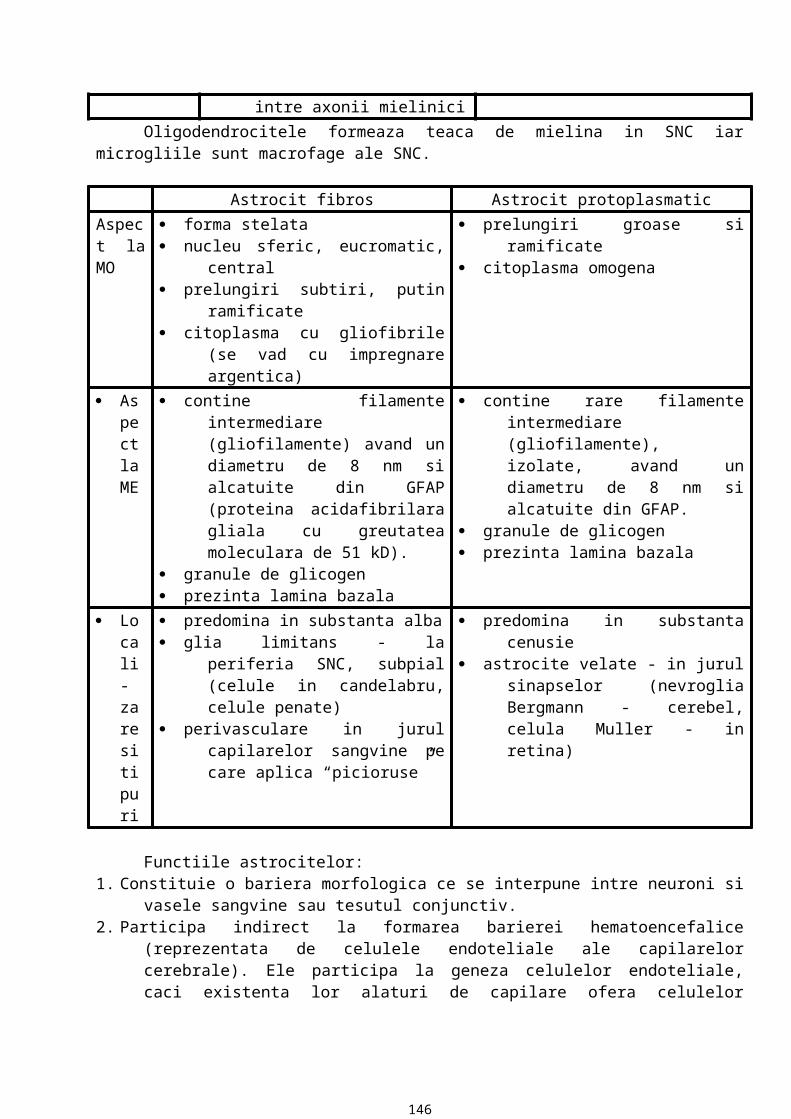
· sunt cele mai aproape de vasul sangvin si prin urmare sunt primele supuse actiunii toxinelor absorbite din tubul digestiv
· in caz de staza biliara sunt primele care sufera modificari.In zona 3 hepatocitele sunt supuse si ele actiunii toxinelor insa in cantitate mai mica. La fel
se exercita efectul si in caz de staza biliara. Aici ajunge sangele cu continutul cel mai mic de oxigen. De aceea in ischemie, primele care sufera sunt celulele din zona 3 (celulele din paropierea venei centrolobulare).
In zona 2 celulele au caracteristici intermediare celorlelte doua zone.De fapt, toate hepatocitele au d.p.d.v. metabolic acelasi potential insa caracteristicile lor
difera doar din cauza alimentarii cu sange cu concentratii diferite de oxigen.
Hepatocitele
Reprezinta 80% din totalitatea celulelor de la nivelul ficatului. Ele au forma poliedrica si diametru de 20-30 mm. Durata de viata este de aprox. 150 de zile.
Examinarea la MONucleulHepatocitele pot fi uni- sau binucleate, cele mai multe fiind uninucleate. Nucleul este mare,
rotund, situat central, eucromatic, avand 1-2 nucleoli. Frecvent (la 80% din hepatocitele uninucleate) se observa poliploidia, cea mai frecventa fiind diploidia insa. Prin urmare si volumul nucleului variaza.
25% din hepatocite sunt binucleate. Raportul intre celulele uninucleate si binucleate se modifica cu varsta (se mareste o data cu inaintarea in varsta) si in timpul proceselor de reparare dupa agresiuni.
La acest nivel se pot observa rare imagini de mitoza. Frecventa mitozelor creste in caz de leziuni sau consecutiv proceselor de reparare.
CitoplasmaEste eozinofila, fin granulara cu granule bazofile care alcatuiesc corpii Berg (ce se vad in
ME) si care sunt aglomerari de RER.Au un continut mare de incluziuni de glicogen si de lipide, prin urmare, in HE, au un aspect
vacuolar.Cu albastru de toluidina citoplasma se coloreaza albastru iar granulele de glicogen se
coloreaza metacromatic in grena.Lipidele se gasesc la periferie si se evidentiaza cu osmiu, cand apar negre.
Examinarea la MELa ME, hepatocitul apare ca o celula polarizata. Bogatia in organite celulare variaza in
functie de gradul de dezvoltare functionala, de activiate si, implicit, cu localizarea hepatocitului la nivelul lobulului.
Hepatocitul are trei tipuri de suprafete:· suprafata care vine in contact cu capilarul sinusoid, numita suprafata sinusoidala sau pol
vascular· suprafata care vine in raport cu alte hepatocite si delimiteaza pe o anumita distanta un spatiu
intercelular (canaliculul biliar intralobular), numita suprafata biliara sau pol biliar· suprafata care vine in raport cu celelalte hepatocite dar nu delimiteaza nici un spatiu intre ele,
celulele fiind unite prin jonctiuni. Aceasta suprafata se numeste suprafata intercelulara.
Reticulul endoplasmic rugos
146

Tinde sa se localizeze la polul vascular. El asigura procesele de sinteza a proteinelor de export hepatice. Hepatocitul sintetizeaza o mare varietate de proteine: majoritatea proteinelor din plasma: factori ai coagularii, lipoproteine, albumine, globuline.
Dupa sinteza proteinele trec prin aparatul Golgi iar apoi sunt exocitate direct, deoarece secretia este continua la nivelul hepatocitului. Prin urmare nu exista granule de secretie.
Reticulul endoplasmic netedEste dispersat in toata citoplasma. El contine seturi enzimatice necesare pentru procesele de
catabolism si pentru procesele de sinteza. REN participa in:· metabolismul glicogenului· sinteza colesterolului· procesele de detoxifiere si inactivare.
Procesele de detoxifiere si inactivare se bazeaza pe reactiile de oxidare si de conjugare. Aici se inactiveaza o mare parte din medicamente: anticonvulsive, h. steroizi, chimioterapice, antihistaminice. Atunci cand creste cantitatea de medicamente creste si activitatea enzimatica de la nivelul REN si prin urmare forma organitelor variaza in functie de necesitatile metabolice ale organismului. Aceste modificari sunt de aspect si sunt reversibile datorita interventiei lizozomilor.
REN participa la formarea bilei prin:· enzimele care realizeaza conjugarea acizilor biliari: acidul glicocolic si acidul taurocolic· enzimele care realizeaza conjugarea bilirubinei cu acidul glucuronic. cand capacitatea de
conjugare a bilirubinei in hepatocit este depasita, bilirubina neconjugata, insolubila se acumuleaza in sange si apare icterul.
MitocondriileSunt rotunde sau ovale, in numar de peste 1.000 mitocondrii / celula. Continutul in
mitocondrii difera, la periferia lobulului hepatocitele avand mai multe mitocondrii.Aparatul GolgiSe acumuleaza la nivelul polului biliar al hepatocitului. Acesta pare ca un organit multiplu.
Exista de fapt un singur aparat Golgi care se ramifica si da nastere la 50 de aparate Golgi in fiecare celula. El are mai multe roluri:· definitivarea sintezei si excretia proteinelor plasmatice· formarea lipoproteinelor· formarea lizozmilor din hepatocit
LizozomiiSunt localizati spre polul biliar. De aceea se mai numesc si corpi peribiliari. Ei au aspect
heteerogen, existand lizozomi primari, secundari si tertiari. Ei contin enzime prin care catabolizeaza diverse substante exogene si organite celulare perimate pana la molecule mici care vor fi utilizate ca substrat energetic sau pentru noi sinteze. Prin urmare lizozomii participa la mentinerea integritatii parenchimului hepatic. De asemenea ei mai stocheaza ioni, in cantitati mari sub forma de feroproteine.
PeroxizomiiPeroxizomii apar rar in hepatocit comparativ cu celelalte organite. Ei se gasesc cu precadere
la polul biliar, in apropierea REN si a incluziunilor de glicogen. Ei contin enzime oxidative air enzima marker a sa este catalaza. Sunt implicati in mai multe procese:· procesul de gluconeogeneza· metabolismul purinelor - metabolizeaza adenina si guanina la acid purinic· metabolismul alcoolului - oxideaza etanolul la acetaldehida
147

· in b-oxidarea acizilor grasi cu lant lung de atomi de carbon.
Incluziunile de glicogenSunt mai numeroase in hepatocitele de la periferia lobulului hepatic. Aspectul lor variaza in
functie de ritmul diurn si de starea de nutritie a individului: dupa o masa bogata in glucide numarul lor creste. Ele pot ajunge uneori si in zona centrala a lobulului hepatic. Au caracter tranzitoriu: apar dupa mese si dispar dupa un anumit timp.
Incluziunile lipidiceNumarul lor creste in trimestrul al II-lea de sarcina, in lactatie. In etilism cronic, hepatocitele
se incarca cu picaturi lipidice, mai ales cele din zona centrolobulara. Mai apoi se incarca toate hepatocitele.
Membrana celulara a hepatocituluiMembrana contine doua domenii specializate:
1. domeniu orientat spre polul vascular2. domeniu orientat spre polul biliar
Domeniul de la polul vascular reprezinta 70% din totalul suprafetei hepatocitului. Acest domeniu este in raport cu capilarul sangvin. Membrana de la acest nivel formeaza microvili care maresc suprafata de contact cu sangele. Membrana de la acest nivel este separata de peretele capilarului sinusoid prin spatiul perisinusoidal Disse in care se gasesc microvilii hepatocitelor si fibre de reticulina.
Domeniul de la polul biliar reprezinta aproximativ 15% din suprafata totala a membranei hepatocitului. Acest domeniu formeaza canaliculul biliar, care are diametrul de aprox. 1 mm. Membrana formeaza spre lumen microvili. Celulele formeaza jonctiuni stranse cu celulele invecinate, fapt pentru care produsii de secretie nu pot difuza in spatiul extrabiliar. La acest nivel exista o activitate enzimatica crescuta datorita glicoproteinelor de membrana. In citoplasma din apropierea polului biliar exista microfilamente de actina care circumscriu canalicului bilar si asigura prin contractie modificarea diametrului acestuia.
Celulele perisinusoidale sunt:· celula Ito· celula “pit” (cu gauri)
Ambele celule sunt asemanatoare cu celulele hematopoietice. Ele au acest rol (hematopoietic) in viata fetala si in caz de anemii cronice la adult.
Celulele Ito (lipocitele, celulele striate, celulele interstitiale)
Se intalnesc relativ rar (o celula Ito la 20-25 hepatocite). Au numeroase picaturi lipidice. Ele au raport strans cu fibrele conjunctive din spatiul Disse. De asemenea, o parte din ele inconjura capilarele sinusoide. Ele vin in contact cu hepatocitelel dar fara sa faca jonctiuni. Pot strabate cordoanele de hepatocite si ............ Ele au mai multe roluri:· secreta colagen II care se organizeaza in fibre de reticulina. La nivelul ficatului, daca au loc
colagenizari excesive la nivelul spatiului Disse este afectat raportul dintre capilarul sangvin si polul vascular al hepatocitului si apar tulburari de troficitate.
· secreta factorii de crestere ai hepatocitelor· depoziteaza lipide si vitamina A de natura exogena· in timpul vietii fetale intervin in hematopoieza.
In conditii patologice apar modificari ale fenotipului celulelor Ito: se transforma in fibroblaste, adipocite, celule stem hematopoietice.
148

Celulele “pit”
Se pot identifica doar la ME. Sunt mai rare. Ele sunt asemanatoare cu celulele endocrine: in citoplasma au numeroase granule electronodense cu diametrul de 300 nm. De aceea, initial s-a considerat ca au rol endocrin. Acum sunt considerate o varietate de limfocite circulante, fiind niste limfocite mari, cu granulatii si activitate de killer.
Tesutul conjunctiv
La un ficat normal, tesutul conjunctiv este in cantitate mica. El se gaseste in capsula organului si in hil de unde trimite prelungiri care insotesc vaselel si ductele biliare pana in spatiile interlobulare Kiernan.
Tesutul conjunctiv se continua in lobulul hepatic sub forma fibrelor de reticulina din spatiile Disse. Ele pot forma la nivelul lobulului hepatic stroma conjunctiva care asigura mentinerea deschisa a capilarelor sinusoide.
Ficatul are o mare putere de regenerare. Are loc atat hiperplazia (cresterea in numar) cat si hipertrofia (cresterea in volum) a celulelor restante. La sobolan, ficatul este regenerat in proportie de 75% in decurs de o luna.
Daca agresiunea la nivelul ficatului creste, creste si cantitatea de tesut conjunctiv si apare fibroza hepatica care debuteaza la nivelul spatiilor portale. Se vor forma in acest caz noi septuri conjunctive cu o orientare aleatorie fata de componentele parenchimului hepatic care vor impiedica raportul normal al hepatocitelor cu capilarele sinusoide si cu canaliculii biliari. De asemenea, aceste septuri conjunctive vor diminua hiperplazierea hepatocitelor care vor fi comprimate si se vor forma niste pseudonoduli. Cauze ale cresterii cantitatii de tesut conjunctiv pot fi tulburari ale nutritiei hepatocitului. Prin modificarea raportului normal intre hepatocit si capilarul sangvin se produce dezorganizarea parenchimului hepatic.
Circulatia prin ficat
Vascularizatia sangvinaLa nivelul ficatului ajunge aproximativ 25% din debitul cardiac de repaus.Ficatul beneficiaza de o dubla vascularizatie: nutritiva si functionala. Circulatia functionala
aduce 75% din sangele din ficat si provine din vena porta. Circulatia nutritiva contine 25% din volumul de sange din ficat si provine din artera hepatica.
Vena porta aduce:· substante absorbite din digestiv· toxine care trec de bariera digestiva· produsi de degradare ai hemoglobinei (de la splina)· produsi de secretie endocrina a pancreasului
Vascularizatia nutritiva se face prin ramurile a. hepatice proprii.Ultimele ramificatii ale v. porte sunt capilarele sinusoide care se gasesc intre hepatocite in
timp ce ultimele ramificatii ale arterei hepatice sunt capilarele a. hepatice ce se gasesc in spatiile portale. Ele se vor deschide in capilarele sinusoide, deci capilarele sinusoide vor contine sange venos amestecat cu sange arterial. Circulatia sangelui in capilarele sinusoide se face de la periferia lobulului spre centru si apoi ale se varsa in vena centrolobulara.
Capilarele sinusoidePrezinta un endoteliu discontinuu care are raport cu lamina bazala, ce contine fibre de
reticulina care formeaza o retea laxa in jurul endoteliului capilar. Peretii capilarului sinusoid nu
149

formeaza o adevarata bariera morfologica intre hepatocit si sange, existand si raporturi directe intre cele doua compartimente, fapt care face ca schimburile sa fie foarte usoare.
Capilarele sinusoide au un diametru mare, de 30-40 mm. Celulele capilarelor sinusoide sunt:· celulele endoteliale - sunt celule alungite, cu nucleu turtit, care proemina spre lumen. Celulele au
putine organite insa au fenestre care sunt grupate, neacoperite de diafragme, vezicule de pinocitoza, ceea ce denota o activitate intensa pentru enzimele de transport.
· celulele Kupffer - deriva din monocitele circulante si apartin astfel SFM (sistemul fagocitar mononuclear). Ele se pot insa divide. Sunt localizate la jonctiunea dintre doua capilare sinusoide si proemina spre lumen. Nu realizeaza jonctiuni cu celulele endoteliale. Au forma stelata, au nucleu eucromatic si prezinta mai multe organite decat celulele endoteliale. Au mai multe caracteristici:· Au organite implicate in sinteze (RER, mitocondrii) deoarece aceste celule sintetizeaza o
parte din proteinele de export ale ficatului· Contin si organite implicate in degradare: lizozomi, fagolizozomi si corpi reziduali.· Celulele Kupffer intervin si in imunitate prin capacitatea lor de a fagocita.· Ele fagociteaza pigmenti, eritrocite imbatranite sau denaturate care nu au fost distruse in
splina. Daca se face splenectomie, aceste celule devin foarte importante. · Au receptori pentru imunoglobuline si complement· Secreta citokine: IL-1
Vascularizatia limfaticaEste foarte bine dezvoltata extralobular. Vasele limfatice se gasesc:
· in capsula ficatului (foarte multe)· in septurile conjunctive· apar pana la periferia lobulului hepatic· nu se vad capilare limfatice la nivelul lobulului hepatic.
Originea limfei este in spatiile Disse. O parte din substantele de la acest nivel, care nu patrund in capilarele sinusoide trec in spatiul Mall (spatiul dintre hepatocitele din placa limitanta si tesutul conjunctiv portal) si de aici ajung in spatiul Kiernan unde vor forma capilarele limfatice.
Circulatia bileiLa acest proces participa hepatocitele si celulele Kupffer. Bila circula de la centrul lobulului
spre periferie. Canalele prin care circula bile pot fi:· intrahepatice:
· intralobulare· canaliculele biliare· canaliculele (pasajele) Hering
· extralobulare· canalele biliare perilobulare· canalele biliare interlobulare· canalele hepatice lobare (drept si stang)
· extrahepatice· canalul hepatic comun· canalul cistic· canalul coledoc (format din unirea canalului hepatic comun cu canalul cistic)Canaliculii biliariReprezinta componenta initiala a sistemului canalicular. Nu au pereti proprii ci sunt
delimitate de membranele a doua hepatocite invecinate. La MO apar ca niste mici cavitati. Folosind impregnarea argentica Gomori, se vede ca aceste canalicule alcatuiesc o retea.
Pasajele Hering
150
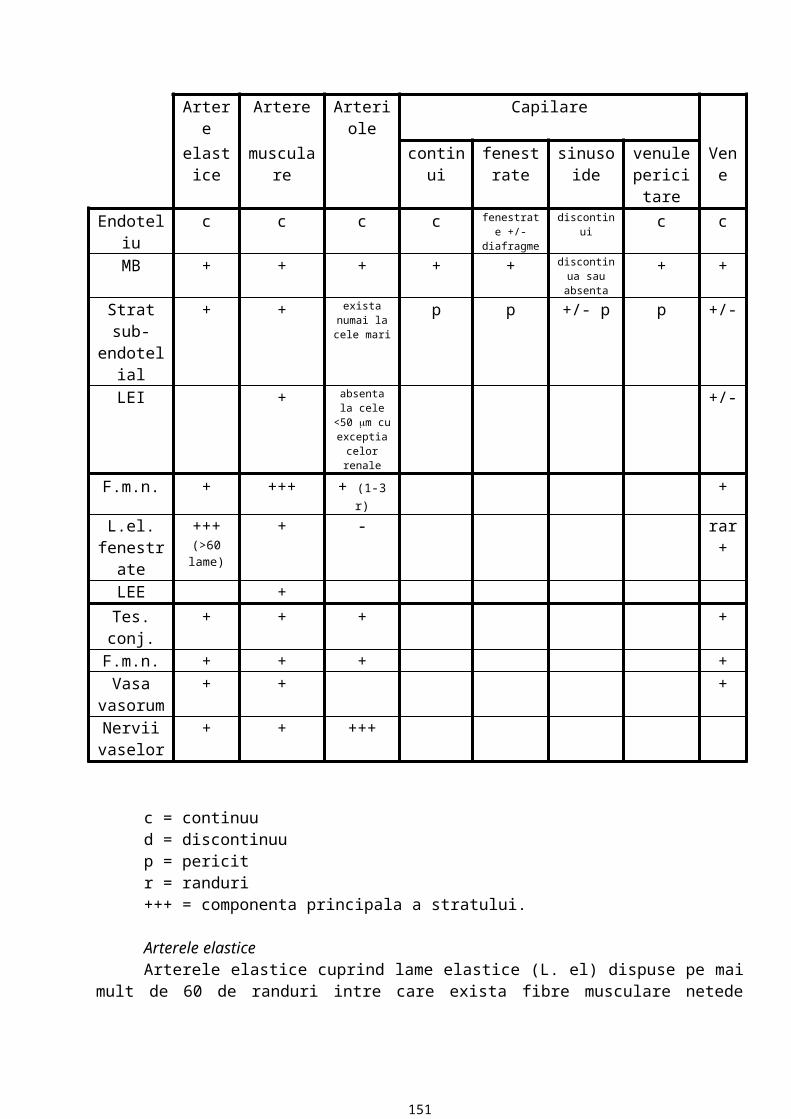
Fac jonctiunea intre canaliculii biliari si canalele extralobulare. Reprezinta portiunea de canalicul biliar aflata la nivelul placii limitante. Ele au perete propriu format dintr-un epiteliu simplu cubic inconjurat de fibre de reticulina.
Caile biliare extralobulareAu un perete format dintr-un epiteliu asezat pe o MB care este inconjurata de un strat de tesut
conjunctiv bogat in fibre elastice. Epiteliul poate fi simplu cubic sau simplu cilindric cu microvili.
Caile biliare extrahepaticePrezinta un perete format din mucoasa, musculara si adventice.Mucoasa contine un epiteliu simplu cilindric cu celule secretoare de mucus si un corion in
care se gasesc fibre elastice, infiltrat limfoid si glande tubuloacinoase de tip mucos.Canalul cistic prezinta in mod particular numeroase pliuri spiralate ale mucoasei care au un
ax muscular. Acestea se numesc valvele spiralate Heister. Canalul coledoc prezinta si el cateva particularitati:
· in epiteliu contine celule care secreta somatostatina. · musculara este initial discontinua. Musculara devine apoi continua si se ingroasa pentru a forma
sfincterele Boyden si Oddi. Musculara se ingroasa deci inainte de unirea cu canalul Wirsung si formeaza aici sfincterul Boyden. Musculara se ingroasa din nou la varsarea ampulei Vater in duoden si formeaza aici sfincterul Oddi.
APARATUL RESPIRATOR
Cavitatea nazala
Mucoasa este formata din epiteliu si corion. Mucoasa este foarte groasa. Epiteliul este de tip respirator (pseudostratificat cilindric ciliat). Majoritatea celulelor au cili,
exista si rare celule caliciforme. Caracteristica este prezenta din loc in loc a unor mici glande endoepiteliale.
Corionul este alcatuit din:· tesut conjunctiv lax· o retea vasculara complexa· glande
Reteaua vasculara pleaca de la ramurile arteriale din vecinatatea peretelui, atasate la periostul (pericondrul) cavitatii nazale. Ramurile arteriale dau diviziuni ce strabat corionul in intregime pana se impart in capilare fenestrate care vor asigura nutritia celulelor epiteliale. de aici sangele va fi colectat in venule postcapilare care se se aduna in lacunele venoase. Lacunele reprezinta vene modificate, cu diametrul mare si care prezinta din loc in loc ingrosari intimale. Aceste ingrosari contin celule musculare ce functioneaza ca sfinctere vasculare . Ele vor retine o cantitate mare de sange in mucoasa nazala. Spre exemplu, gripa sau alergie determina stocarea sangelui in cavitatea nazala. Din aceasta cauza se administreaza decongestionante care vor face sangele sa treaca mai departe, in ramurile venoase atasate de perioast (pericondru).
Printre ochiurile retelei vasculare exista glande compuse, ale caror ducte se deschid la suprafata epiteliului. Per ansamblu, mucoasa are:· celule caliciforme
151

· glande ce apartin corionului· glande endoteliale
Toate aceste formatiuni isi varsa produsii de secretie in lumenul cavitatii nazale si acesta va retine particulele din aer.
De asemenea, reteaua vasculara complexa determina incalzirea aerului. Tot aceasta retea complexa explica de ce in caz de epistaxis sangerarea este mare.
In cavitatea nazala exista o suprafata de aprox. 500 mm2 acoperita de mucoasa olfactiva, ce are rol senzorial. Componentele mucoasei senzoriale sunt:· epiteliu cu celule senzoriale (neuroni bipolari modificati)· celule de sustinere (cu cili)· din loc in loc exista celule cu microvili la polul apical· celule stem
Celulele senzorialeAu o dendrita ce la suprafata epiteliului este dilatata. Aceasta dilatatie se numeste bulb
olfactiv . De aici pleaca 3-4 cili imobili, modificati. Ultrastructura la ME a cililor este diferita de a unui cil obisnuit: in partea proximala este la fel ca un cil dar in partea distala prezinta chemoreceptori.
Axonul celulei senzoriale intra in alcatuirea nervuluui olfactiv. Acesti neuroni au o viata scazuta (aprox. 30 de zile). Ei sunt permanent reinnoiti. Ei reprezinta contactul cel mai direct al SNC cu mediul extern.
Laringele
Are o sstructura complexa. D.p.d.v. histologic prezinta:· tunica mucoasa· schelet cartilaginos· adventitie (tesut de invelis
laringele are mai multe roluri:· mecanismul fonatiei· impiedica trecerea aerului inspirat spre tubul digestiv· impiedica trecerea alimentelor ingerate spre caile respiratorii
MucoasaAre un contur neregulat si determina doua perechi de pliuri (falduri), numite pliuri
ventriculare. Intre ele determina spatiul numit reces ventricular. Aceste pliuri se mai numesc si corzi vocale false. Exista insa si corzi vocale adevarate.
In majoritate, mucoasa contine epiteliu de tip respirator. La nivelul corzilor vocale adevarate, epiteliul trece brusc in epiteliu pluristratificat pavimentos nekeratinizat. Acesta reprezinta o adaptare impotriva efectului abraziv al coloanei de aer.
In corzile vocale false, corionul este variabil ca grosime si are:· tesut conjunctiv lax· glande laringeale· foliculi limfoizi
Din loc in loc exista pachete de fibre elastice organizate sub forma “ligamentului ventricular”.
In corzile vocale adevarate, corionul este mai scazut ca grosime, se pastreaza insa asocierea de fibre elastice ce determina in acest caz ligemantul vocal. Acesta se insera pe cartilajul tiroid, in 1/3 mijlocie, merge spre cartilajele aritenoide iar spre trahee se prelungeste prin formatiunea numita corn elastic care se va prinde de cartilajul cricoid. Lateral de acest ligament este un m. striat (m.
152

tireoaritenoidian), portionea mediala a sa alcatuind m. vocal. Lig. vocal impreuna cu m. vocal participa in mecanismul fonatiei.
Scheletul cartilaginosExista doua tipuri de cartilaj:
· hialin (spre exterior):· cartilajul tiroid· cartilajul cricoid
· elastic (spre exterior)· epiglota· cartilajele aritenoidieneAdventitiaSe gaseste la periferia laringelui. Ea reprezinta o cale de acces pentru vase si terminatiuni
nervoase. De asemenea, ea este un instrument de legatura cu structurile din jur.
Traheea
Este un organ tubular, cu lungime de 16-20 cm, cu 16-20 de inele cartilaginoase incomplete alcatuite din cartilaj hialin. Prezzenta inelelor determina doua regiuni:
· regiunea anterioara - cartilaginoasa· regiunea posterioara - membranoasa (contine celule musculare)
Stratigrafie:· mucoasa (epiteliu si corion)· strat cartilaginos (anterior)· strat muscula (posterior)· tunica externa
Epiteliu este de tip respirator (pseudostratificat cilindric ciliat) cu o MB mai groasa. Corionul are grosimea inegala. El este denumit dupa unii tela submucosa. El contine:
· tesut conjunctiv lax· multe ramuri vasculare· glande traheale (seromucoase)
Tunica cartilaginoasa este formata din lame de cartilaj hialin asezate orizontal, solidarizate intre ele prin membrane fibroelastice (spatii prin care trec vase si nervi de la exterior la interior).
In regiunea posterioara, capetele sunt solidarizate prin fascicule musculare netede dispuse longitudinal spre adventice si circular spre lumen. Printre fasciculele musculare, la acest nivel exista glande traheale, ale caror ducte excretorii ajung la suprafata epiteliului
Adventitia este alcatuita din tesut conjunctiv.
Bronhiile primare
Au aceeasi structura histologica cu traheea.
Plamanii
Au trei elemente:· un invelis, de tip mezoteliu, pleura. Ea este alcatuita din doua foite, una parietala si alta
viscerala.· stroma, cu o retea de septuri conjunctive ce compartimenteaza organul in lobi, segmente si lobuli· parenchimul pulmonar care contine:
· caile aeriene intrapulmonare si· ramificatiile nervoase, vasculare si limfatice ce le insotesc.
153
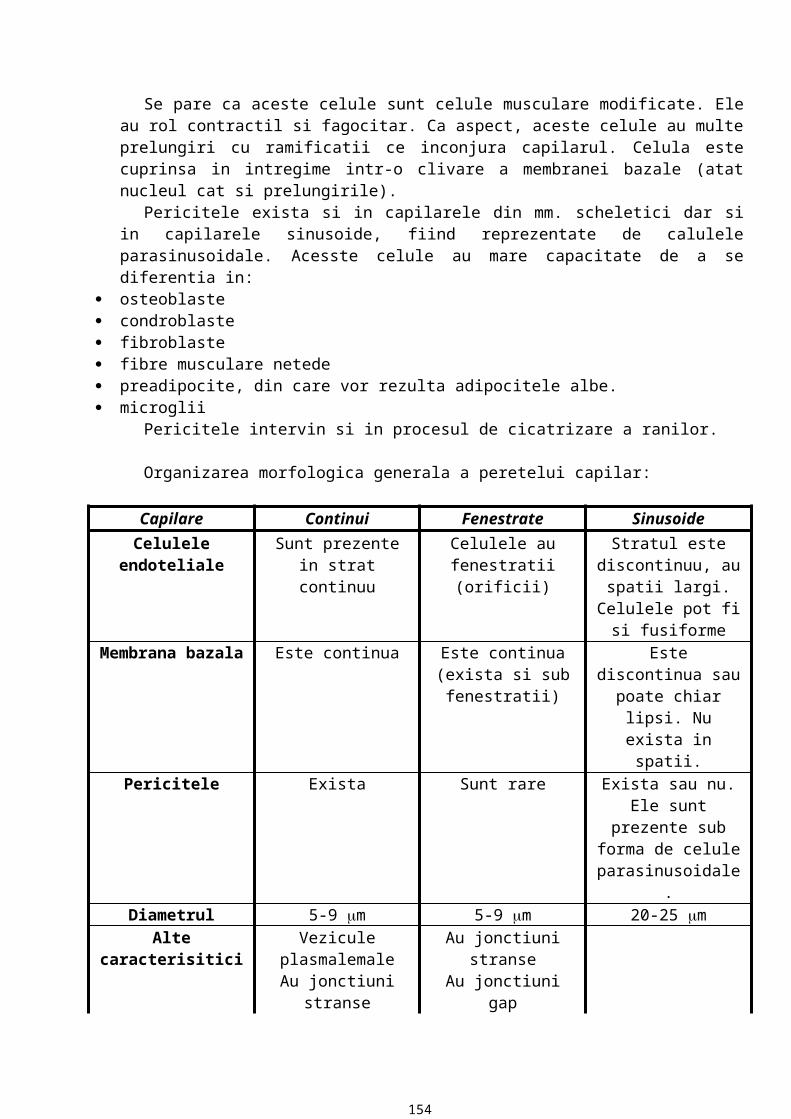
Caile aeriene intrapulmonareSunt acoperite de epiteliu pseudostratificat cilindric ciliat. Toate bronhiile intrapulmonare au
astfel de epiteliu. Falsa stratificare se datoreaza celulelor se vin in contact cu MB dar nu ajung pana la suprafata epiteliului. Celulele care sunt inalte cat toata grosimea epiteliului prezinta la polul apical cili cu dimensiunea de 7 mm si exista cca. 100-200 cili / celula. Ei se misca cu frecventa de 25 deplasari / secunda. Epiteliul mai contine:· Celule caliciforme, care sunt glande unicelulare mucoase, fara specializari la polul apical si care
prezinta multe granule de mucus in partea supranucleara a celulei.· Celule stem care asigura reinnoirea. Ele sunt mici, piramidale, se fixeaza la nivelul MB si cresc
treptat in volum· Celule endocrine (granulare) - sunt si ele asezate pe MB, intra in contact cu terminatiile
nervoase colinergice. La ME se observa multe organite si granule de secretie electronodense care se gasesc in regiunea bazala. D.p.d.v. histochimic exista doua tipuri de celule:· care sintetizeaza adrenalina sau noradrenalina· care sintetizeaza hh. peptidici
Produsul de secretie e eliberat la polul bazal iar prin capilare controleaza activitatea contractila a mm. netezi vasculari.
· Celule senzoriale (rare). Ele au la polul apical micrvili. La polul bazal au conexiiuni cu terminatiile nervoase. Caile aeriene intrapulmonare se divid dicotomic. Din traheerezulta 2 bronhii principale, care
determina doua bronhii secundare care determina doua bronhii tertiare etc.Structura histologica nu se modifica esential insa se modifica diametrul cailor aeriene
(dioametrul scaade). Elementele caracteristice (care deosebesc caile cu diametru mare fata de caile cu diametru mic) sunt elementele cartilaginoase:
· cartilajul hialin este prezent la caile cu diametru mare· treptat cartilajul hialin este inlocuit cu cartilaj elastic. Lamele cartilaginoase devin fine si acestea
se gasesc in bronhiile cu diametru mic. · Cand diametrul devine 0,5-2 mm structura cartilaginoasa nu mai exista iar caile devin bronhiole.
Bronhiile intrapulmonareTunica mucoasa este formata din epiteliu respirator asezat pe corion destul de putin inalt.
Corionul contine o retea de fibre elastice in care fasciculele sunt asezate longitudinal.Tunica musculara este formata din benzi (fascicule) de tesut muscular neted, spiralate, mai
aproape unele de altele in bronhiile intrapulmonare cu diametru mare iar pe masura ce diamtrul caii aeriene scade, fasciculele devin mai distantate.
Stratul cartilaginos este alcatuit din mai multe piese de cartilaj hialin cu contur foarte neregulat solidarizate prin benzi de tesut conjunctiv dens. Intre muschi si cartilaj exista glandele bronsice.
Adventitia ataseaza calea aeriana de calea arterei si venei pulmonare.
Bronhiola preterminalaBronhiola preterminala exista aproape de teritoriul de tranzit pentru aer unde acesta se
opreste. Structura histologica este diferita. Diametrul este de 0,5-2 mm. Epiteliul este simplu cilindric cu corpi neuroepiteliali si celule Clara. Dincolo de epiteliu exista o lama fina de tesut conjunctiv. Stratul muscular contine celule musculare spiralate. Prezinta si tunica de invelis (adventitie).
Mucoasa contine corpi neuroepiteliali. Acestia reprezinta asocieri de 10-80 celule strans atasate, cu proprietati argirofile. La ME se observa ca lapolul apical exista microvili. Au un singur nucleu, uneori indentat si multe organite. Aceste celule au su numeroase granule de secretie,
154
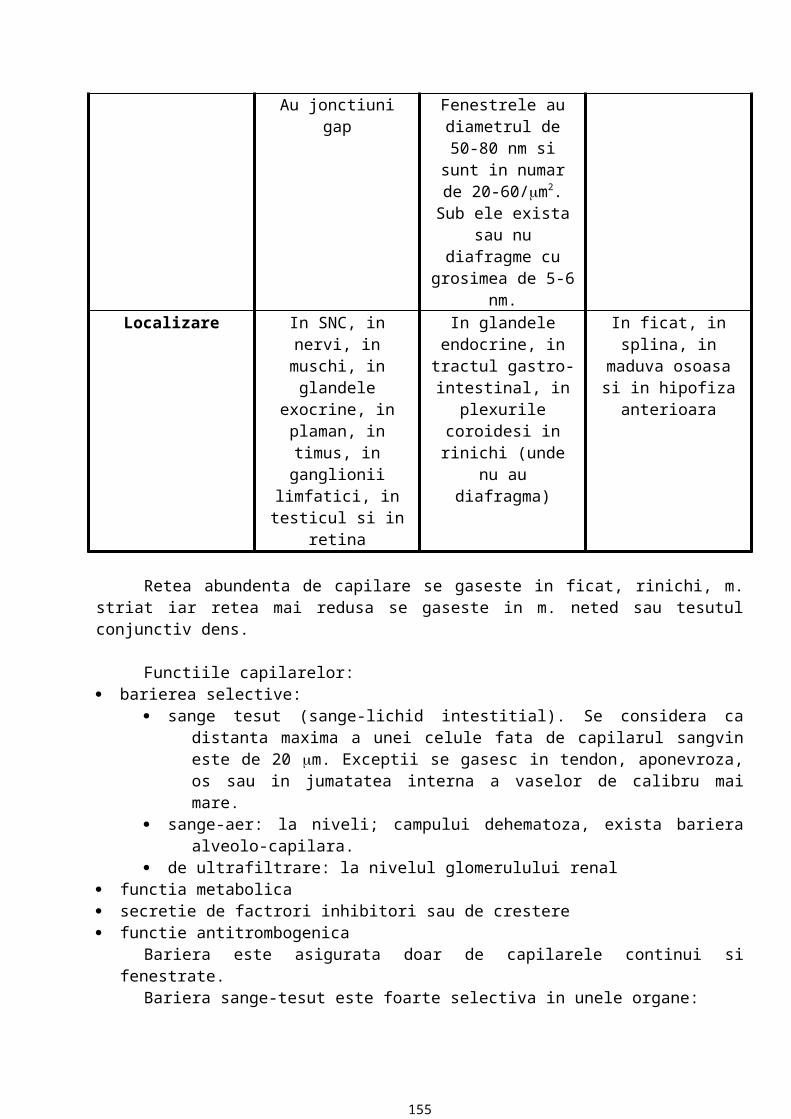
acumulate mai laes la polu bazal. Ele iau contact cu cate o terminatie nervoasa. Aceste celule sunt echivalentele celulelor endocrine din celelalte cai aeriene. Produsii de secretie pot fi de asemenea catecolamine sau hh. polipeptidici.
De regula corpi neuroepiteliali sunt inconjurati de celule Clara (asemanatoare cu celulele caliciforme). Aceste celule sunt bombate, fara specializari iar la ME se observa ca au un continut crescut de granule de secretie alcatuite din GAG si colesterol. Produsul de secretie eliberat de aceste celule are rol de protectie a suprafetei epiteliului.
Bronhiolele terminaleSunt asemanatoare cu bronhiolele preterminale dar au un diametru mai mic. Fiecare
bronhiola terminala determina 2-3 bronhiole respiratorii. Ele reprezinta spatii de schimb. La nivelul lor se deschid primele alveole.
Bronhiolele respiratoriiSunt acoperite de epiteliu simplu cubic. De asemenea ele mai contin o lama fina de tesut
conjunctiv si celule musculare netede dispuse circular, mai apropiate in portiunea initiala a bronhiolei respiratorii si apoi mai distantate in portiunea distala. La capatul bronhiolei respiratorii, din tunica musculara raman inele incomplete ce unesc orificiile de deschidere ale alveolelor.
Fiecare bronhiola respiratorie se imparte in doua ducte alveolare, care nu mai prezita pereti proprii. Aceste ducte iau nasterea datorita asezarii orificiilor de deschidere ale mai multor alveole pe aceeasi circumferinta.
Alveolele
Sunt spatii de schimb. In organism exista aprox. 150 mil. alveole. Fiecare este ca o sfera cu diametrul de 300 mm. Ele sunt separate unele de altele prin septuri alveolare, capilare si celule. Septurile prezinta din loc in loc discontinuitati numite perikohn cce au rolu de a egaliz presiunea intre alveolele invecinate.
Celulele ce alcatuiesc alveolele pulmonare sunt de doua feluri:· pneumocite de tip I · pneumocite de tip II
Pneumocitele de tip ISunt cele mai multe, asigura 90% din spatiul de schimb. Sunt celule foarte aplatizate (au doar
70 nm grosime) si au foarte putine organite. U o suprafata mare (4.000 mm2) si sunt atasate unele de altele prin jonctiuni stranse. Pe suprafata lor se observa din loc in loc microvezicule. Ele isi au originea din celulele alveolare de tip II.
Pneumocitele de tip IISunt mari, globulare, cu diametrul de 10-15 mm. Sunt atasate de pneumocitele de tip I si au
suprafata membranara libera spre alveole. La ME se observa ca au un singur nucleu rotund, central si contin aproape toate organitele: RER, ribozomi, mitocondrii dar nu contin lizozomi. Spre suprafata libera au doua caracteristici:· corpii multilamelari· corpii multiveziculari
Corpii multiveziculari se transforma in corpi multilamelari. Corpii au doua roluri:1. elimina produsul de secretie la suprafata epiteliului2. contin un factor antiatelectatic (atelectaia este procesul de alipire a peretilor alveolari. Acest
factor antiatelectatic este surfactantul care este un amestec de substante PC-like, lecitina etc. Acest amestec de substante se aseaza intr-o pelicula ca de film compusa din 3 straturi pe suprafata alveolara. Are grosimea de 30 nm. La suprafata totala pulmonara care este de 150 m2 corespund cateva mg de surfactant. Rolul sau este:3. modifica tensiunea superficiala la suprafata
155

4. ataseaza particulele de impuritati5. faciliteaa deplasarea macrofagelor alveolare pe suprafata alveoleiSub epiteliu exista o retea capilara cu foarte multe anastomoze, formata din capilare
nefenestrate. Printre ele exista fibre elastice ce se intind in inspir si protejeaza structurile alveolare de modificari bruste. Exista si o retea de fibre de colagen si celule:· celule conjunctive fixe - fibroblaste si fibrocite care produc fibrele elastice si de colagen· celule sangvine migrate: neutrofile, limfocite, mastocite.
Bariera alveolo-capilaraEste o membrana biologica cu urmatoarele elemente:
· portiunea aplatizata a celulelor alveolare de tip I· o membrana bazala alcatuita prin fuziunea MB a celulelor de tip I cu MB a celulelor endoteliale· portiunea aplatizata a celulelor endoteliale (de la nivelul capilarului)
Are grosimea de aproximativ 2,2 ± 3 mm. Ea este bariera aer-sange iar gazele respiratorii strabat aceasta structura prin difuziune pasiva: oxigenul trece din alveola spre capilar iar dioxidul de carbon trece din capilar spre alveola.
Macrofagele alveolareSunt celule cu diametru mare, au multe prelungiri (pseudopode) care le permit sa se
deplaseze pe suprafata alveolei si sa fagociteze diverse particule ajunse pana aici: carbune, praf, bacterii etc.
La ME se observa ca aceste celule au un singur nucleu foarte plicaturat, au o citoplasma bogata cu multe organite dintre care forte multi lizozomi si fagolizozomi. Acestia ajuta la descompunerea particulelor fagocitate.
Uneori aceste macrofage nu digera complet particulele fagocitate. Atunci maceste mecrofage vor evolua in doua feluri:migreaza in sens invers, ajungand in in caile aeriene cu diametru mare, aceasta petrecandu-se datorita secretiilor mucoase si cililor. Macrofagele ajung astfel in spatiul de unde vor fi eliminate prin tuse.
· migreaza strabatand peretele alveolar, la nivelul pneumocitelor de tip I, ajung in septul alveolar si apoi pe cale limfatica ajung in ganglionii limfatici unde aceste particule vor fi distruse in doua moduri:· macrofagele transfera particulele nedigerate macrofagelor fixe de aici care le vor distruge· macrofagele primesc in ganglion instructiunea de autodistrugere si apoi va fi fagocitat de
macrofagele fixe.Macrofagele alveoalre isi au originea in monocitele circulante iar in spatiul tisular se
modifica.
156

APARATUL GENITAL FEMININ
Aparatul genital feminin este alcatuit din:1. gonade - ovare2. cai genitale
3. trompa uterina4. uter5. vagin
6. glande anexe7. glanda mamara8. placenta
Trompa uterina
Este un organ pereche, cavitar, localizat in cavitatea peritoneala, dispus lateral de uter si medial de ovar. D.p.d.v. anatomic prezinta 4 rediuni:· zona infundibulara· zona ampulara· zona istmica· zona intramurala
Trompa uterina prezinta si 2 orificii prin care comunica:· cu cavitatea peritoneala - in zona infundibulara· cu cavitatea uterina - ostiumul tubar
D.p.d.v. histologic peretele are trei tunici:· mucoasa· musculara· seroasa
Mucoasa
157

Prezinta numeroase falduri sau pliuri longitudinale care umplu lumenul si ii dau un aspect labirintic. Aceste falduri se ramifica dar nu se anastomozeaza, astfel incat fenestrele dintre falduri comunica intre ele. Acesta este un element de diagnostic diferential intre trompa uterina si vezicula seminala unde faldurile se anastomozeaza si fenestrele nu mai comunica intre ele.
Faldurile sunt lungi si dese in zona infundibulara si se reduc ca numar in zona istmica. Ele dispar in zona intramurala unde intalnim o mucoasa aproape neteda. Aceste falduri au rol in deplasarea ovulului sau zigotului catre cavitatea uterina.
Mucoasa contine un epiteliu si un corion (lamina propria).Epiteliul contine trei tipuri de celule:
1. Celulele ciliate - cilii lor vin in contact cu lumenul. Aceste celule predomina in faza foliculara, cilii dezvoltandu-se sub actiunea astrogenilor. Cilii au rolul de a deplasa lichidul de la suprafata mucoasei, impreuna cu ovulul sau zigotul. Ei faciliteaza si deplasarea spermatozoizilor pana la zona ampulara.
2. Celulele secretorii - sunt mai scurte, au polul apical rotunjit. Ele nu vin in contact cu lumenul in faza foliculara cand numarul lor este si foarte redus. Ele cresc insa ca numar in faza luteinica si intra in contact cu lumenul deoarece celulele ciliate se reduc ca inaltime.
3. Celulele intercalate - vin in contact cu MB dar nu ajung niciodata in contact cu lumenul. Ele sunt precursori ai celulelor secretorii in care se vor transforma in timpul fazei luteinice.
Celulele secretorii produc un lichid care are rol in:· capacitatia spermatozoizilor· troficitatea oului sau ovulului· transportul ovulului sau al zigotului
Prin urmare, participa la transportul oului sau ovulului urmatoarele elemente:· cilii celulelor ciliate· faldurile mucoasei· musculara, prin miscarile sale peristaltice
Corionul este format din tesut conjunctiv lax bogat vascularizat dar fara glande. Vasele sunt mari, artere sau vene, care in timpul ovulatiei devin hiperemice, turgescente, conferind trompei uterine un grad mare de distensibilitate si o reducere a motilitatii ei care este necesara pentru a se putea aplica pe ovar pentru a capta ovulul.
Tunica muscularaEste alcatuita din muschi neted dispus in doua straturi: circular intern si longitudinal extern.Tunica musculara este mai groasa in zona intramurala si istmica si este mai subtire in zona
ampulara si infundibulara. Ea ajuta la deplasarea oului sau ovulului si a spermatozoizilor.
SeroasaEste alcatuita din spiteliu simplu pavimentos si tesut conjunctiv bogat vascularizat. De fapt
seroasa este alcatuita din mezoteliul peritoneal.
Rolurile trompei uterine· Trompa uterina preia ovocitul II in ovulatie. Acesta poate fi fertilizat in zona ampulara, caz in
care el isi va definitiva a doua diviziune meiotica si se va forma, prin unirea cu spermatozoidul, zigotul (oul).
· In trompa uterina incepe segmentatia oului timp de 4-5 zile· Dupa 4-5 zile, prin ostium, oul va fi impins in cavitatea uterina unde el va nida.· La nivelul trompei uterine se poate implanta oul segmentat. Aceasta se poate intampla din mai
multe cauze:· dupa anexite acute netratate, care s-au cronicizat si au dus la micsorarea sau chiar
astuparea ostiumului
158

· datorita malformatiilor trompui uterine (lumen prea ingust, foarte lungi, foarte inalte sau infantile)
Daca implantarea s-a facut in zona istmica sarcina nu se dezvolta mai mult de 2-3 saptamani dupa care trompa se rupe, apare hemoragia si este necesara interventia chirurgicala de urgenta.
Daca implantarea se face in zona ampulara sau infundibulara sarcina se poate dezvolta 2-3, maxim 4 luni dupa care se produce avort prin ruptura de trompa uterina sau evacuare in cavitatea peritoneala, in ambele cazuri existand hemoragie si necesitand interventia chirurgicala de urgenta.
Uterul
Este un organ cavitar, situat inapoia vezicii urinare si in fata rectului. El are forma de para cu varful in jos. D.p.d.v. anatomic i se descriu mai multe regiuni:· corpul uterin· istmul uterin· colul uterin
D.p.d.v histologic corpul si istmul uterin au structura asemanatoare iar colul uterin are structura diferita. La nivelul corpului se deschid cele doua ostiumuri tubare. Portiunea dintre cele doua ostiumuri se numeste fundul uterin si la acest nivel peretele este mai subtire. De aceea, in aceasta zona peretele se poate rupe foarte des in timpul chiuretajelor.
Corpul si istmul uterin
Au peretele format din trei tunici:1. mucoasa = endometru2. musculara = miometru3. seroasa = perimetru
EndometrulEste alcatuit dintr-un epiteliu si corion. Epiteliul se imparte in:
· epiteliu de acoperire - epiteliu simplu cilindric· epiteliu glandular - este o extensie a epiteliului de acoperire la nivelul glandelor uterine (cripte
uterine) care reprezinta invaginari ale epiteliului in corion. Aceste glande sunt simple, tubulare drepte, rareori fiind ramificate si daca sunt ramificate aceasta se intampla numai in portiunea profunda endometrului.
Corionul este format din tesut conjunctiv lax spinocelular bogat vascularizat si inervat.Portiunea bazala a endometrului reprezinta 1/4 din grosimea totala a endometrului (2 mm din
8-9 mm). Aceasta portiune alcatuieste endometrul bazal si nu este sensibila la actiunea hormonilor ovarieni, deci nu sufera modificari in timpul ciclului uterin. Endometrul bazal contine:· fundul glandelor uterine· corion dens citogen
Endometrul functional reprezinta 3/4 superioare din grosimea endometrului. El contine:· corpul si colul glandelor uterine· corion citogen mai putin dens
Endometrul functional este deosebit de sensibil la actiunea h. ovarieni si el sufera modificari importante de-a lungul unui ciclu ovarian.Aceasta portiune se va elimina in timpul menstruatiei. De asemenea, in faza secretorie si in timpul sarcinii, endometrul fonctional este subdivizat in 2 parti:· partea compacta (superficiala) - contine epiteliul de acoperire, colul glandelor uterine si
corionul aferent
159

· partea spongioasa - contine corpul glandelor uterine si corionul aferent
MiometrulEste format din fibre musculare netede dispuse in 3 straturi:
· stratul intern - fibre musculare netede dispuse circular si longitudinal· stratul extern - fibre musculare netede dispuse de asemenea circular si longitudinal· stratul mijlociu - fibre musculare netede oblic spiralate. In acest strat se gasesc arterele si venele
care alcatuiesc reteaua vasculara arcuata. Acestor vase le lipseste adventicea, deci tunica musculara a vasului se continua cu fibrele musculare netede ale miometrului. Aceasta particularitate are rol in oprirea foarte rapida a hemoragiei dupa nastere, dupa interventii chirurgicale etc.
PerimetrulEste format din mezoteliul peritoneal (epiteliu simplu pavimentos) si tesut conjunctiv dens.
Endometrul (continuare)Endometrul este tesutul cu cea mai mare sensibilitate la actiunea h. ovarieni. Au loc
modificari zilnice ale acestui tesut. Prin urmare acest tesut sufera transformari ciclice ce se repeta la fiecare 28 de zile (de ovbcei) si care alcatuiesc ciclul endometrial. Ciclul endometrial se suprapune peste ciclul ovarian.
Endometrul contine elemente epiteliale si elemente mezenchimale.
Elementele epitelialeSunt reprezentate de cele doua epitelii (de acoperire si glandular). Aceste epitelii vor fi
formate din mai multe tipuri de celule:· celule proliferative (celuel bazale). Aceste celule formeaza epiteliul de acoperire si epiteliul
glandular din endometrul functional in faza proliferativa. De asemenea, aceste celule vor forma glandele de la nivelul portiunii bazale a endometrului.
Celulele sunt inalte, cilindrice, cu citoplasma putina, usor bazofila sau amfofila. Raportul nucleo-citoplasmatic este >1. Nucleii sunt alungiti, tahicromatici, cu nucleoli mici. Acestea sunt celule care se divid intens. Din aceasta cauza apare pseudostratificarea spiteliului glandular si de acoperire.
· Celulele epiteliale ciliate - formeaza epiteliul de acoperire si epiteliul din zona colului uterin in faza proliferativa a ciclului endometrial. Ele descresc ca numar in faza secretorie. Falsa stratificare a epiteliului aproape dispare in faza secretorie. Aceste celule au citoplasma acidofila, mai abundenta si nucleul eucromatic, cu nucleoli evidenti.
· Celulele secretorii - sunt putine ca numar in faza proliferativa si mai numeroase in faza secretorie. Aceste celule nu se mai divid. Ele acumuleaza in citoplasma granule cu produsi de secretie care se pot localiza mai intai subnuclear (nucleul fiind situat apical), apoi acestea se impart (nucleul fiind situat central), apoi granulele se situeaza la polulapical (nucleul fiind situat bazal). Daca produsii de secretie sunt prezenti, avem de a face cu celula secretorie vacuolara. Dupa exocitoza produsilor de secretie, polul apical se rotunjeste si avem de a face cu celula secretorie nevacuolara. Celulele secretorii se gasesc predominant in epiteliul de acoperire in faza proliferativa si apoi se localizeaza predominant in corpul si colul glandelor uterine, din faza proliferativa tardiva pana la sfarsitul fazei secretorii.
Elementele de natura mezenchimala
Stroma conjunctiva
160

Este constituita din tesut conjunctiv lax spinocelular, in care predomina componenta celulara. Exista si putine fibre de reticulina si putina substanta fundamentala.
CeluleleCelulele se dispun in vartejuri in jurul glandelor si vaselor. Exista mai multe tipuri de celule:
· celula stromala reticulara de tip fibroblast. Aceasta celula este foarte sensibila la actiunea h. ovarieni:· In faza proliferativa, incipienta si mijlocie, celula este fusiforma, are dimensiuni mici,
citoplasma usor bazofila si nucleul tahicromatic.· In faza proliferativa tardiva celula creste in dimensiuni, apar prelungirile, se incarca cu
organite celulare si acest proces continua si in faza secretorie incipienta si mijlocie.· In faza secretorie tardiva are loc procesul de predecidualizare care consta in hipertroierea
celulelor care se vor incarca cu incluziuni de glicogen, citoplasma va deveni deci foarte acidofila iar nucleul va fi mare si eucromatic. In acest stadiu limitele intercelulare sunt foarte evidente. Aceste celule vor forma decidua materna.
· celula spinoasa (xantomatoasa) - are origine mezenchimala local si nu origine histiocitara. Ea apare in faza secretorie mijlocie si tardiva. Are o citoplasma cu multe vacuole lipidice si un nucleu mic, reniform, tahicromatic.
· celula granulomatoasa - nu provine din granulocitele sangvine ci are origine limfocitara, fiind asemanatoare cu limfocitele mari, citotoxice, cu granulatii in citoplasma. Apare in faza secretorie mijlocie si tardiva.
· limfocitele - sunt izolate si apar in faza secretoire mijlocie si tardiva.· plasmocite - secreta IgA. Si ele sunt izolate Daca sunt grupate semnifica existenta unei
endometrite.
Fibrele de reticulinaSunt un marker important pentru functionalitatea endometrului si pentru receptivitatea
hormonala. Fibrele de reticulina sunt produse de celula stromala reticulara de tip fibroblast. Numarul si localizarea lor sunt influentata de h. ovarieni:· Fibrele de reticulina sunt putine in faza proliferativa incipienta cand se dispun periglandular si
periarterial· Spre sfarsitul fazei proliferative ele vor creste ca numar si se vor dispune si in stroma
interglandulara si intervasculara.· In faza secretorie continua sa creasca numarul lor, devin mai groase.· In ziua 22 (faza secretorie mijlocie) se edematiaza si le dispare reactia argirofila. Prin urmare ele
nu vor mai putea fi evidentiate la MO insa nu dispar.· In ziua 23 reapare reactia argirofila iar fibrele se dispun sub forma de cuiburi in jurul celulelor
predeciduale. Din aceasta cauza limitele lor se vor obsrva atat de usor.
Substanta fundamentalaEste bogata in apa, proteine, glucide, lipide si mucopolizaharide cu greutate moleculara mare
ce apara in special in faza secretorie.
Vasele sangvineArteriolele spiralate provin din aa bazale ale endometrului. Arterele bazale ale endometrului
provin din aa. radiate si scurte de la limita endometru - miometru. Aa. radiate si scurte provin la randul lor din reteaua vasculara arcuata de la nivelul miometrului.
Aa. spiralate sunt foarte receptive la actiunea hormonilor ovarieni:· In faza proliferativa incipienta ele sunt drepte, au lumen ingust· In faza proliferativa tardiva ele devanseaza in crestere stroma si devin spiralate
161

· In faza secretorie pereteii se ingroasa foarte mult si vor prezenta contractii ritmice, sub influenta h. ovarieni cu frecventa de 60-70 / min.
· In faza secretorie tardiva, cand nu se mai secreta h. estrogeni sau progesteroni apar contractii lungi care vor duce la aparitia unor zone de ischemie. Api apare necroza si impastarea tesutului iar in final va avea loc eliminarea (descuamarea) endometrului functional impreuna cu o cantitate variabila de sange. Endometrul functional nu se elimina tot la un momant dat ci alterneaza zonele descuamate cu zonele de proliferare, motiv pentru care endometrul are un aspect foarte heterogen ce variaza de la o ora la alta.
Ciclul endometrial
Ciclul endometrial reprezinta ciclul menstrual, are o durata de 28 de zile de obicei si se suprapune perfect peste ciclul ovarian. El are mai multe faze:· faza menstruala (2-5 zile)· faza proliferativa (estrogenica) - corespunde fazei foliculinice din ciclul ovarian (din ziua 5 pana
in ziua 14)· faza secretorie (progesteronica) - corespunde fazei luteinice din ciclul ovarian (din ziua 14 pana
in ziua 28)Variabilitatea duratei ciclului este data de variabilitatea duratei fazei proliferative, durata
fazei secretorii fiind constanta.· faza proliferativa:
· endometrul se reface pe baza stratului bazal (de la 2 mm grosime acesta va ajunge la 8-9 mm)
· in epiteliul de acoperire predomina celulele ciliate si celulele proliferative· glandele uterine au lumen ingust, nu au produs de secretie in lumen, sunt rectilinii, avand
o crestere concomitent cu stroma. Predomina celulele proliferative iar in faza proliferativa tardiva apar si celulele secretorii. Epiteliul are un aspect de pseudostratificare deoarece in peretii glandelor uterine se evidentiaza numeroase mitoze.
· stroma este densa, bogat celulara, predominand celulele stromale reticulare de tip fibroblast. Si la acest nivel sunt prezente mitozele.
· arterele spiralate sunt drepte, au lumenul ingust si peretele subtire. Ele se spiralizeaza in faza proliferativa tardiva
· se evidentiaza numeroase fibre de reticulina· Faza secretorie
· glandele, in sectiune transversala au aspect spiralat sau stelat.· epiteliul de acoperire este format din celule ciliate si celule secretorii· glandele uterine devanseaza in crestere stroma, asupra lor se exercita o presiune si ele se
spiralizeaza. Glandele vor fi deci spiralate sau stelate. In epiteliul glandular predomina celulele secretorii. Epiteliul nu mai are aspect de pseudostratificare. Ele au lumen larg in care se evidentiaza produsul de secretie intens acidofil.
· in stroma nu mai exista mitoze si se gasesc celule stromale reticulare de tip fibroblast alaturi de celule xantomatoase, celule granulomatoase, limfocite si rare plasmocite.
· se evidentiaza fibrele de reticulina· arterele spiralate au peretele gros ce prezinta contractii ritmice cu frecventa de 6-0-70 /
min. La sfarsitul fazei secretorii apar sunturi arteriovenoase, adevarate lacuri de sange dispuse in corionul de sub epiteliul de acoperire. La sfarsitul fazei secretorii, cand nu se mai secreta estrogeni si progesteron apar contractii lungi ale aa. spiralate care vor duce la aparitia ischemieie, a necrozei, impastarii tesutului si in final a descuamarii sale.
162

Descuamarea este parcelara, se face prin eliminarea si a unei cantitati variabile de sange. Prin urmare apare sange in lumenul glandei uterine.
Colul uterin
Reprezinta partea inferioara a uterului, cu forma de butoias, la nivelul careia se insera vaginul. Prin urmare, colul uterin are doua portiuni:· supravaginala· infravaginala
Lumenul colului uterin se numeste canal cervical. El are doua orificii:· intern - asigura comunicarea cu istmul uterin· extern - esigura comunicarea cu vaginul
Pe o hemisectiune la nivelul colului uterin se observa ca central se afla un ax fibromuscular format din 75-85% tesut conjunctiv fibros si 15-25% muschi neted. Fibrele musculare netede sunt organizate in fascicule separate de tesut conjunctiv fibros. Colul uterin este mai dur decat corpul uterin. Sub influenta hormonilor ovarieni si placentari, axul conjunctiv muscular se ramoleste si colul se inmoaie si are aspect catifelat.
Colul uterin este impartit in doua parti:· endocolul (spre canalul cervical)· exocolul
EndocolulEpiteliul de acoperire este epiteliu simplu cilindric, bogat in celule mucoase. Acest epiteliu se
invagineaza in corion si da nastere glandelor cervicale care sunt tubulare ramificate. Ele au lumenul mai larg decat glandele uterine si prezinta adesea produs de secretie cu tenta acidofila in lumen.
Corionul este format din tesut conjunctiv lax bogat vascularizat. Uneori orificiile de varsare ale glandelor epiteliale se pot obstrua (in infectii). Atunci produsul de secretie se aduna in lumenul glandular, epiteliul glandular se turteste si se formeaze un ou (chist) al lui Naboth. Cand aceste chisturi sunt foarte multe ele proemina in canalul cervical si il pot chiar obstrua, caz in care este necesara interventia chirurgicala.
ExocolulEste acoperit de epiteliu pluristratificat pavimentos nekeratinizat. Prezinta un corion fara
glande dar care contine tesut conjunctiv dens bogat vascularizat cu vase de calibru mare (artere si vene) si filtrat limfocitar nodal sau difuz.
Secretia colului uterin este foarte importanta. Ea difera in functie de fazele ciclului ovarian. Aceasta secretie este mai apoasa in timpul ovulatieim facilitand patrunderea spermatozoizilor prin canalul cervical. In faza progesteronica secretia este vascoasa pentru a impiedica patrunderea germenilor. Secretia de la acest nivel poarta numele de glera. Glera participa si la formarea secretiei vaginale normale.
Jonctiunea epiteliu simplu cilindric - epiteliu pluristratificat pavimentos nekeratinizat este brosca. Aceasta jonctiune este frecvent sediul unor tumori, cancerul de col uterin fiind foarte frecvent dar nu se moare din aceasta cauza, indicele de mortalitate fiind de 8 : 100.000.
Vaginul
Este un conduct musculo-fibros cilindric care se insera la nivelul colului uterin unde determina aparitia a 4 funduri de sac.:· doua funduri de sac laterale· fundul de sac posterior - cel mai larg, locul de prelevare a secretiilor
163

· fundul de sac anterior - cel mai scurtPeretele vaginal are mai multe tunici:
· mucoasa· musculara· adventice
MucoasaNu are glande. Secretiile vaginale se formeaza insa din:
· glera de la nivelul endocolului· transudatia plasmatica ce are loc la nivelul vaselor mari ale corionului din mucoasa vaginala.
Secretia vaginala are un pH acid. Daca pH-ul ar fi neutru sau chiar bazic mucoasa ar fi foarte expusa la infectii. PH-ul este acid deoarece celulele mucoase sintetizeaza glicogen. Celulele superficiale, incarcate cu glicogen, se descuameaza iar glicogenul se raspandeste in lumen. El va fi metabolizat la acid lactic de bacilul Dodderlain care constituie flora bacteriana saprofita a vaginului. Astfel pH-ul mucoasei vaginale este acid.
Epiteliul este pluristratificat pavimentos nekeratinizat in care se gasesc zone de precornificare (prekeratinizare) deoarece in celulele superficiale epiteliale exista keratohialina. Epiteliul vaginal are o grosime variabila, intre 150-200 mm si e format din urmatoarele straturi celulare:· stratul bazal, la nivelul MB - un singur rand de celule· stratul parabazal - 3-4 straturi de celule· stratul intermediar· stratul superficial
Celulele stratului superficial se pot descuama si prin urmare ele se pot preleva in secretia vaginala. Din secretie se va face un frotiu care se va colora prin tehnica Babes-Papa-Nicolau care foloseste mai multi coloranti:· pentru nucleu se foloseste hemalaunul· pentru citoplasma se foloseste fuxina acida si verdoglobina???????
Celulele mucoase:· Celulele bazale sunt celule cubice sau usor rotunjite, cu diametrul de 15 mm, cu un nucleu de 10
mm diametru, eucromatic si cu citoplasma bazofila.· Celulele parabazale au diametrul de 20 mm, cu un nucleu de 8-9 mm diametru, citoplasma
bazofila.· Celulele intermediare sunt de doua feluri:
· celule adevarate - au aspect de nacela (naviculare). Ele apar doar pe frotiurile de sarcina. Astfel se poate pune diagnosticul de sarcina inainte sa se pozitive testele hormonale.
· celule mari - au diametrul de 25 mm iar nucleul are un diametru de 5-6 mm. Celula sufera fenomene de prepicnoza si apoi picnoza prin cariorecsis (fragmentarea nucleului) si carioliza (liza nucleului). Aceste celule au citoplasma bazofila sau amfofila deoarece incepe secretia de glicogen care se dispune perinuclear
· Celulele superficiale - prezinta acumulari lipidice perinuclear.Sub influenta estrogenilor celulele epiteliale se matureaza rapid si vor fi multe celule
acidofile cu nuclei picnotici. In faza estrogenica deci:· indicele acidofil este de 70%· indicele picnotic este de 90%
Progesteronul faciliteaza descuamarea celulelor sin stratul intermediar si parabazal. Prin urmare el impiedica maturarea. In faza progesteronica deci:· indicele acidofil este de 5-10%· indicele picnotic este de 5%
Prin stabilirea celor 2 indici este cel mai usor de stabilit profilul hormonal al persoanei.
164

In faza luteinica, in epiteliul vaginal exista si limfocite. Acestea nu trebuie interpretate ca o stare de infectie.
Glanda mamara
Se gaseste in partea anterioara a toracelui, prepectoral, in hipodermul regiunii.Fiecare din cele doua glande are in structura sa 15-20 lobi glandulari, fiecare lob fiind o
glanda separata, cu canal de extcretie propriu (canal galactofor) care se deschide la nivelul mamelonului prin porul galactofor.
Lobul glandular este format din lobuli glandulari delimitati intre ei de tesut conjunctiv fibros si adipos. In lobuli este stroma, formata din tesut conjunctiv lax, si parenchimul. In tesutul conjunctiv lax stromal exista limfocite si plasmocite care cresc ca numar in faza progesteronica si in timpul sarcinii. Se secreta si IgA care se transmite prin laptele matern la nou-nascut si se realizeaza astfel imunizarea pasiva.
In stroma, premenstrual, apare un edem stromal si se produce mastrodinia premenstruala.
Glanda mamara in repausParenchimul are canalele de secretie intralobulare delimitate de epiteliu simplu cubic si
insule de celule secretorii in repaus care pot fi izolate sau atasate la un canal secretor intralobular. Canlul intralobular se varsa intr-un canal extralobular (canal galactofor) care sunt delimitate de un epiteliu bistratificat cubic sau cilindric. Inainte de varsare, canalul galactofor se dilata si formeaza un sinus galactofor care este delimitat de derm pe de o parte si de epiteliul bistratificat al canalului galactofor pe de alta parte.
Glanda mamara in lactatieCelulele secretorii aflate in repaus devin active, inmuguresc si vor da nastere acinilor
glandulari care au forme si marimi diferite. Acinii vor fi delimitati de epiteliu simplu cubic. In lumenul acinilor exista laptele, produsul de secretie care este acidofil. Stroma se reduce, in rest, celelalte structuri aau aceeasi alcatuire ca in repaus.
Secretia este apocrina.
165

APARATUL GENITAL MASCULIN
Este format din:· gonada (glanda genitala) = testiculul care are secretie mixta:
· exocrina - spermatozoizii· endocrina - h. testosteron
· caile genitale - cu rol de depozitare si transport al spermatozoizilor· glandele genitale accesorii:
· doua vezicule seminale· prostata· doua glande bulbouretrale
· organul copulator
Testiculul
Se dezvolta in cavitatea abdominala si apoi coboara in bursele scrotale, acest lucru fiind esential pentru generarea spermatozoizilor (este necesara o temperatura cu 2-3ºC mai mica decat tenperatura corpului). Cand acestea nu coboara, spermatozoizii nu sunt generati si apare sterilitatea (la om). Spre exemplu, la om testiculele coboara doar in perioada imperecherii.
Ele ou forma ovoidala, diametrul longitudinal de 5 cm, latimea de 2,5 cm iar lungimea axului antero-posterior este de 3 cm. Greutatea medie este de 14 g dar exista variatii, in functie de talia individului etc.
Cei doi poli, fata anterioara si fetele laterale sun acoperite de tunica vaginala (foita viscerala a peritoneului). Deci tunica vaginala este alcatuita dintr-un mezoteliu si o membrana bazala. Foita viscerala se muleaza si pe capul epididimului.
Dupa tunica vaginala urmeaza albugineea care este o capsula fibroasa, inextensibila, formata din tesut conjunctiv, fibre de colagen si elastina cu dispozitie lamelara si fibre musculare netede rare. Posterior, tunica albuginee formeaza o ingrosare numita mediastinum testis (corpul Highmore) din care pornesc septuri conjunctive care devin incomplete in partea opusa mediastinului si care delimiteaza niste camarute, lobulii testiculari, ca niste trunchiuri de piramida cu baza mare spre tunica albuginee.
Fiecare testicul are aprox. 250 lobuli. In fiecare lobul exista tubii seminiferi (1-4 / lobul). Ei au traseu foarte sinuos si intre ei exista tesut interstitial care reprezinta prox. 35% din volumul testicular. Tesutul interstitial este format dintr-o retea de tesut conjunctiv lax cu putine fibre de colagen si reticulina, fibroblasti si rare macrofage si mastocite. Prin acest tesut patrund vase de sange, capilare fenestrate cu diafragme. Mai exista vase limfatice, nervi si grupuri de celule endocrine (celulele Leydig) care secreta testosteron.
Sub albuginee se vede tunica vasculosa care contine vase de sange cu calibru mult mai mare. Aceste vase trec de la un lobul la altul prin zona unde septurile conjunctive sunt incomplete.
Celulele LeydigAu forma poligonala, diametrul de 15-20 mm, sunt grupate si sunt inconjurate de o bogata
retea de capilare sangvine. Au nucelul mare, eucromatic, in general excentric. Rareori pot fi
166

binucleate. Citoplasma este acidofila si poate avea aspect spongios, vacuolar, datorita picaturilor lipidice din interior.
La ME se observa ca aceste celule au caracteristici comune cu cele ale celulelor secretoare de hormoni steroizi: REN abundent, aparat Golgi bine dezvoltat, mitocondrii cu criste tubulo-veziculare, picaturi lipidice. La om se intalnisc si niste formatiuni cristaloide de natura proteica, numite cristale Reinke care cresc o data cu inaintarea in varsta. Exista si incluziuni de lipofuscina.
Secretia celulelor Leydig este stimulata de:· LH care se mai numeste si interstitial cell stimulating hormone - ICSH.· prolactina
Celulele Leydug uncep sa se diferentieze in viata fetala si incep sa secrete testosteron, cu rol in dezvoltarea aparatului genital masculin. Din L4 incep sa involueze, pana in L8 iar la nastere nu mai exista (nu se pot diferentia de fibroblaste). La pubertate se diferentiaza din nou si vor ramane toata viata. Aici se elibereaza 95% din testosteron iar 5% se elibereaza la nivelul corticosuprarenalei. In fiecare i se secreta 7 mg de testosteron din care:· O parte trece in circulatie si devine testosteron circulant. El ajuta la:
· diferentierea aparatului genital masculin· diferentierea si dezvoltarea SNC· imprimarea caracterele sexuale secundare· intervine in procesele anabolice· determina comportamentul de tip masculin
· Alta parte intra in epiteliul tubilor seminiferi si ajuta la diferentierea si proliferarea celulelor liniei seminale. Pentru aceasta este necesar ca testosteronul sa fie foarte concentrat in epiteliul seminifer ( de 200 de ori mai concentrat ca in sange.). Concentrarea testosteronului se face cu ajutorul celulelor de sustinere Sertoli prin proteina ABP (androgen binding protein).
Tubul seminifer contortFiecare tub are o lungime de 40-60 cm si un diametru de 200-300 mm. In fiecare testicul
exista 4.000 - 6.000 de tubi seminiferi. Prin urmare, lungime totala a tubilor seminiferi este de 250 m in fiecare testicul. Ambele capete ale tubului sunt orientate spre mediastinum testis. Unul din capete este orb (in deget de manusa) iar celalalt capat se continua cu o portiune scurta, tubii drepti.
Organizare generala (dinspre periferie spre lumen):· Tunica de tesut conjunctiv format din fibroblaste si celule mioide (cu capacitate de contractie)
dar la om predomina fibroblastele. Toate celulele sunt dispuse pe 2-3 randuri.· Membrana bazala bine definita. Exista cazuri cand aceasta MB se ingroasa, cazuri in care apare
infertilitatea.· Epiteliul seminifer, germinativ, care contine mai multe randuri de celule (4-8). La nivelul
lumenului se observa spermatozoizii care au flagel. Celulele epiteliului seminifer sunt:· celule germinative (proliferative) - apartin liniei spermatogenetice (spermatogonii A /
spermatogonii B ® spermatocite I ® spermatocite II ® spermatide ® spermatozoizi imaturi.
· celule de sustinere (neproliferative) - celulele Sertoli, care nu apartin liniei seminale.O spermatogonie produce in final aprox. 16 spermatozoizi. In testicul, in 74 de zile, iau
nastere spermatozoizii imaturi. Ei nu au capacitate fecundanta si nici motilitate proprie. Mai urmeaza inca 10 - 12 zile de maturare la nivelul cozii epididimului.
Spermatogoniile sunt de doua feluri: A si B. Ele sunt situate la baza epiteliului germinativ, deasupra MB. Celulele germinative se gasesc intre faldurile de sustinere ale celulelor Sertoli care sunt ca niste buzunare. Spermatogoniile A alcatuiesc compartimentul susa pentru celulele
167

germinative si nu sunt diferentiate. Spermatogoniile A au nucleul ovalar si, in functie de cromatina, se impart in:· spermatogonii A dark, care au cromatina fin granulara dar mai condensata la periferie. Au si doi
nucleoli atasati la membrana nucleara. · spermatogonii A pale, care au cromatina tot fin granulara dar nucleul este mai deschis la culoare
decat in cazul spermatogoniilor A dark.Spermatogoniile A, prin diviziune, dau nastere spermatogoniilor A dark sau A pale care prin
diviziune, mai departe (4-8 diviziuni), dau nastere spermatogoniilor B. Spermatogoniile B sunt celule diferentiate si prin diviziune dau nastere spermatocitelor I. Ele au nucleul rotund, cromatina este grunjoasa, mai condensata, mai ales in celulele de la periferia lobulului. Nucleolul este central.
Spermatogoniile B cau initial 12 mm, apoi ele cresc in dimensiuni si prin diviziune mitotica dau nastere spermatocitelor I. Acestea sunt cele mai mari celule din seria seminala, avand diametrul de 16 mm. Ele au un set diploid de cromozomi (46 cromozomi bicromatidici).
Spermatocitele intra apoi in prima diviziune meiotica. Profaza dureaza 22 de zile si deci ea se intalneste frecvent in preparatele histologice. Dintr-un spermatocit I rezulta 2 spermatocite II, fiecare cu cate 23 de cromozomi bicromatidici. Dupa o perioada scurta ( de cateva ore) spermatocitele II intra in a doua diviziune meiotica in care nu se mai dubleaza cantitatea de ADN. Astfel, spermatocitele II se intalnesc rar pentru ca ele traiesc putin. Ele au diametrul de 9 mm si in final, din cele doua spermatocite II rezulta 4 spermatide care au cate 23 de cromozomi monocromatidici. Spermatidele vor da nastere apoi la 4 spermatozoizi fara a se mai divide.
SpermatogenezaSpermatogeneza cuprinde trei etape:
1. spermatocitogeneza (formarea spermatocitelor)2. spermiogeneza (formarea spermatozoizilor)3. spermiere (eliberarea spermatozoizilor in lumenul tubului seminifer)
In timpul etapelor spermatogenezei exista o cariokineza completa (de la o etapa la alta, nucleii celulelor se separa complet) dar exista o citokineza incompleta (intre celulele de un fel exista punti de citoplasma). Aceste punti au rolul de a sincroniza diferentierea celulelor care are loc in valuri de aceeasi generatie. La om diferentierea in valuri se face pe generatii diferite in acelasi tub seminifer. O spermatogonie A intra in procesul de diferentiere si diviziune la un moment dat. Toate celulele ce rezulta din aceasta spermatogonie se vor divide toate in acelasi moment datorita prezentei puntilor citoplasmatice. Insa nu toate spermatogoniile A intra in acelasi moment in diviziune. Vor exista prin urmare valuri de celule aflate in diferite etape de diferentiere in acelasi tub seminifer.
spermiogeneza reprezinta transformarea spermatidelor in spermatozoizi. Initial, spermatidele sunt rotunde, cu diametrul de 6 mm,fiind cele mai mici celule din linia seminala. Modificarile care urmeaza au loc concomitent dar, din motive didactice, le impartim in etape:1. Formarea acrozomului
Aparatul Golgi este bine dezvoltat. El da nastere la macrovezicule care vor fuziona si vor da nastere unei vezicule mai mari care se ataseaza la suprafata dinspre aparatul Golgi a nucleului. Aceasta vezicula se numeste vezicula acrozomala. In continuare, aparatul Golgi sintetizeaza macrovezicule care se ataseaza la vezicula acrozomala. Ea va creste si va incepe sa se muleze pe suprafata nucleuluui. In final va acoperi 2/3 din supraafata nucleului si se va numi acrozom. Acrozomul contine enzime hidrolitice care au rolu de a digera zona pellucida si de a patrunde in nucleul ovulului spre a fuziona cu materialul genetic feminin.
2. Formarea flageluluiInitial centriolii migreaza in partea opusa aparatului Golgi. Centriolii vor sintetiza
microtubuli care se vor asambla si vor forma in final flagelul.3. Modificarea nucleului
168

Initial nucleul este central si rotund. Apoi el va deveni excentric, de forma discoidala (turtit antero-posterior) iar cromatina se va condensa foarte mult. Din nucleul initial, eucromatic, se obtine un nucleu tahicromatic, care contine numai heterocromatina si, uneori, niste vacuole interne.
4. Mitocondriile migreaza si se dispun in portiunea initiala a flagelului. Ele se dispun intr-un singur rand, cap la cap, helicoidal.
5. Dupa aceea, excesul de citoplasma este indepartat, spermatozoizii formati fiind eliberati in lumen iar din excesul de citoplasma formandu-se corpii reziduali Reynolds care vor fi fagocitati de celulele Sertoli.
La inceput, spermatozoidul este imatur. El are 65-70 mm lungime. El are urmatoarea structura:· cap - contine:
· nucleul foarte condensat, avand forma in functie de specie· acrozomul· membrana plasmalemala
· gatul - contine 1 centriol· flagelul - are trei parti:
· piesa mijlocie - are aceeasi lungime ca si capul (aprox 5 mm)· piesa rpincipala - are aprox. 50 mm, fiind cea mai mare· piesa terminala - are aprox. 5 mm.
· Pe toata lungimea flagelului exista axonema alcatuita din 9 perechi de microtubuli periferice si o pereche centrala. La partea terminala, peste axonema vine membrana plasmatica. Peste piesa principala vine un invelis fibros format din mai multe coloane (ca niste coaste). La piesa mijlocie, se gaseste mai intai axonema, apoi invelisul fibros, teaca mitocondriala si apoi plasmalema.In sindromul Kartagena?? apar defecte la nivelul microtubulilor. Apar spermatozoizi imobili
si apare sterilitatea. Dar microtubulii intra si in structura cililor, deci persoana va avea si afectiuni respiratorii.
Celulele de sustinere SertoliSunt celule inalte, asezate pe un strat continuu pe MB iar polul apical ajunge pana in lumen.
Faldurile laterale patrund intre celulele liniei germinative. Prin urmare este greu de urmarit conturul unei celule Sertoli.
Intre doua celule Sertoli vecine se stabilesc jonctiuni nepermeabile care vor determina doua compartimente in care se gasesc celule germinative:
· compartimentul bazal (in care exista doar spermatogonii)· compartimentul adluminal - spre lumen, in care se afla restul celulelor liniei seminale.
Rolul celulelor Sertoli este de a constitui bariera sange-testicul. Aceasta bariera este necesara pentru a proteja celulele haploide de sistemul imun intrucat aceste celule prezinta pe suprafata alte antigene decat cele proprii organismului respectiv si ar putea fi distruse de componentele sistemului imun.
Aspectul celulelor la MO:· nucleul este in treimea bazala a celulei, are forma ovalara, triunghiulara sau neregulata si este
eucromatic, cu nucleol evident.· citoplasma este abundenta, acidofila, cateodata usor vacuolara.
Aspectul la ME:· in jurul nucleolului se vad doua mase de heterocromatina.· citoplasma contine REN si RER bine dezvoltat, mitocondrii numeroase, alungite. Exista
incluziuni lipidice, multi lizozomi, pigment de imbatranire - de lipofuscina. Mai exista niste
169

formatiunicristaloide, corpii Charcot-Bottcher. La polul apical, citoscheletul este bine dezvoltat si se vad multe mmicrofilamente de actina si miozina.
La polul apical, exista jonctiuni intre celulele Sertoli si spermatide care tin strans legate spermatidele de celulele Sertoli pana cand se diferentiaza in spermatozoizi imaturi. Cand diferentierea s-a terminat, jonctiunea se desface si, prin miscarile polului apical (format din microfilamente de actina si de miozina) spermatozoizii sunt eliminati in lumen. Dar corpii Reynolds (excesul de citoplasma al spermatidei ) raman legati de celula Sertoli si vor fi fagocitati.
Rolu celulelor Sertoli:1. Au rol de suport si nutritie pentru celulele germinative. Celula Sertoli mai este numita si
“nursing cell” deoarece prin intermediul ei ajung substantele nutritive din sange la celulele liniei seminale.
2. Secreta fluidul din tubii seminiferi, necesar pentru mobilitatea si nutritia spermatozoizilor.3. Secreta ABP care are rol in concentrarea testosteronului in epiteliul seminifer. Acesta secretie
este influentata de FSH.4. Fagociteaza corpii reziduali si celulele germinative moarte.5. Asigura eliberarea spermatozoizilor in lumen. 6. Secreta mici cantitati de hormoni estrogeni.
Celulele Sertoli nu se divid in perioada reproductiva. Ele confera rezistenta la infectii, iradiere etc.
Exista si tubi seminiferi aspermatogeni, mai numerosi o data cu inaintarea in varsta. Ei predomina la batrani. Acesti tubi sunt alcatuiti numai din celule Sertoli. Celulele Sertoli si in partea distala a tubului seminifer unde acesta se ingusteaza si apoi se continua cu tubii drepti.
Caile genitale
Caile genitale sunt:· tubii seminiferi· tubii drepti· rete testis· tubii eferenti· canalul epididimal· canalul deferent care se uneste cu:· canalul ejaculator si impreuna se varsa in:· uretra prostatica· uretra membranoasa· uretra peniana
Tubii drepti contin un epiteliu simplu cubic asezat pe o MB si tesut conjunctiv dens. Ei au un traiect scurt.
Rete testis se gaseste in mediastinum testis. Este o retea de tubi ce [rezinta epiteliu simplu cu celule turtite, cubice sau columnare (cu umai multi microvili si un cil). Rolul cililor si microvililor este de a impingefluidul testicular mai departe, in ductii eferenti.
Tubii eferenti sunt 10-20 de tubi care pornesc din partea superioara a mediastinumului. Initial au un traict rectiliniu, apoi vor avea un traiect foarte sinuos, formand capul epididimului. Ei au un epiteliu simplu cu celule cubice si columnare care au mai multi cili. Sub MB a epiteliului exista un strat de fibre musculare netede. Tubii eferenti au doua roluri:· asigura mobilitatea fluidului, impingandu-l spre epididim· absoarbe fluidul seminal (in mai mica masura)
Canalul epididimal se formeaza prin unirea tubilor eferenti intr-unul singur. El are traiect foarte intortocheat. El formeaza corpul si coada epididimului. Intins, ar avea o lungime de 5 m. Diametrul tubului creste spre coada epididimului: initial are un diametru de 150 mm iar in final are
170

diametrul de 400 mm. Este captusit de un epiteliu pseudostratificat care contine celule bazale mici (celule de inlocuire) si celule principale care sunt inalte in partea initiala (80 mm) iar in partea finala sunt mai scunde (40 mm). La polul apical ei prezinta stereocili. Canalul epididimal are mai multe roluri:· asigura absorbtia fluidului testicular (absorbindu-se aici 90% din fluidul testicular)· secreta glicoproteine cu rol nutritiv pentru spermatozoizi.
In partea initiala a canalului exista 1-3 randuri de fibre musculare netede care au contractii ritmice. In partea finala a canalului contractia este redusa. Aici se depoziteaza spermatozoizii aprox. 12 zile si se matureaza pana devin fertili.
Canalul (ductul) deferent este continuarea epididimului si el intra in alcatuirea cordonului spermatic. El ajunge in veciantatea prostatei si in partea terminala are o dilatatie numita ampula ductului deferent.
171