Școala doctoralĂ interdisciplinarĂ facultatea de ......4.2.3.1 valori medii fenotipice și...
TRANSCRIPT

ȘCOALA DOCTORALĂ INTERDISCIPLINARĂ
Facultatea de Silvicultură și Exploatări Forestiere
Ing. Ecaterina – Nicoleta CHESNOIU căs. APOSTOL
Discriminarea taxonomică prin descriptori ai frunzelor şi analiza
fenologiei de primăvară la stejarul pedunculat (Quercus robur L.) şi
stejarul brumăriu (Quercus pedunculiflora K. Koch)
Taxonomic discrimination through leaf descriptors and analysis of
spring phenology in pedunculate oak (Quercus robur L.) and grayish
oak (Quercus pedunculiflora K. Koch)
REZUMAT / ABSTRACT
Conducător ştiinţific
Prof. dr. ing. Neculae ȘOFLETEA
BRAȘOV, 2017

i
Investeşte în oameni! FONDUL SOCIAL EUROPEAN Programul Operaţional Sectorial Dezvoltarea Resurselor Umane 2007 – 2013 Axa prioritară 1 „Educaţie şi formare profesională în sprijinul creşterii economice şi dezvoltării societăţii bazate pe cunoaştere” Domeniul major de intervenţie 1.5. „Programe doctorale şi post-doctorale în sprijinul cercetării” Titlul proiectului: Burse doctorale si postdoctorale pentru cercetare de excelenta Numărul de identificare al contractului: POSDRU/159/1.5/S/134378 Beneficiar: Universitatea Transilvania din Braşov
D-lui (D-nei) ..............................................................................................................
COMPONENȚA
Comisiei de doctorat
Numită prin ordinul Rectorului Universității Transilvania din Braşov Nr. 8955 din 6.11.2017
PREŞEDINTE: Prof.dr.ing. Ovidiu IONESCU
Prodecan Facultatea de Silvicultură și Exploatări Forestiere
Universitatea Transilvania din Brașov
CONDUCĂTOR ŞTIINŢIFIC: Prof.dr.ing. Neculae ȘOFLETEA
Universitatea Transilvania din Brașov
REFERENȚI: Prof.dr.ing. Gheorghe Florian BORLEA
Universitatea de Științe Agricole și Medicină Veterinară a
Banatului din Timișoara
Conf.dr.ing. Florin CLINOVSCHI
Universitatea “Ștefan cel Mare” din Suceava
Prof.dr.ing. Alexandru Lucian CURTU
Universitatea Transilvania din Brașov
Data, ora şi locul susținerii publice a tezei de doctorat: 18.12.2017, ora 11.00, sala SI2, Facultatea de
Silvicultură și Exploatări Forestiere
Eventualele aprecieri sau observații asupra conținutului lucrării vor fi transmise electronic, în timp util,
pe adresa [email protected].
Totodată, vă invităm să luați parte la şedința publică de susținere a tezei de doctorat.
Vă mulțumim.

ii
CUPRINS
Pg. Pg.
teza rezumat
1. Mulțumiri........................................................................................................................................... 2 1
2. Introducere………………………………………………………………………………………… 3 2
1. Stadiul actual al cunoștințelor privind taxonomia, corologia, variabiliatea fenologică şi
fenotipică ale stejarului pedunculat (Q. robur L.) şi stejarului brumăriu (Quercus
pedunculiflora K. Koch).................................................................................................................... 4 3
1.1 Arealul și ecologia stejarului pedunculat și stejarului brumăriu............................................... 4 3
1.2 Poziția taxonomică a stejarului pedunculat și stejarului brumăriu reflectată în literatura de
specialitate....................................................................................................................................... 6 3
1.3 Variabilitatea morfologică şi ecotipică a stejarului pedunculat (Q. robur L.).......................... 7 4
1.3.1 Descriptori fenotipici şi variabilitatea intraspecifică a stejarului pedunculat.................. 7 4
1.3.2 Rezultate ale studiilor privind variabilitatea morfologică şi ecotipică a stejarului
pedunculat (Q. robur L.)........................................................................................................... 9 4
1.4 Variabilitatea morfologică şi ecotipică a stejarului brumăriu (Quercus pedunculiflora K.
Koch)............................................................................................................................................... 10 5
1.4.1 Descriptori fenotipici şi variabilitatea intraspecifică a stejarului brumăriu..................... 10 5
1.4.2 Rezultate ale studiilor privind variabilitatea morfologică şi ecotipică a stejarului
brumăriu (Quercus pedunculiflora K. Koch)............................................................................ 13 5
1.5 Fenologia arborilor.................................................................................................................... 14 5
1.5.1 Importanța studierii fenologiei arborilor.......................................................................... 14 5
1.5.2 Rezultate obținute în România și pe plan internațional.................................................... 17 6
2. Scop şi obiective............................................................................................................................. 20 7
2.1 Scopul cercetărilor..................................................................................................................... 20 7
2.2 Obiectivele cercetărilor............................................................................................................. 21 7
3. Materialul şi metoda de cercetare................................................................................................ 22 8
3.1 Materialul de cercetare.............................................................................................................. 22 8
3.2 Metoda de cercetare……………………………………………………………………...…… 28 9
3.2.1 Analiza fenotipică............................................................................................................. 28 9
3.2.2 Analiza fenologiei............................................................................................................ 40 15
3.2.3 Fazele formării inelului anual........................................................................................... 43 17
4. Rezultate şi discuţii........................................................................................................................ 45 18
4.1 Variabilitatea taxonomică intraspecifică – diferențieri de varietăți și forme după caractere
ale frunzelor..................................................................................................................................... 45 18
4.1.1 Unități taxonomice infraspecifice ale stejarului pedunculat-analiză intra- și
interpopulațională.................................................................................................................... 45 18
4.1.2 Unități taxonomice infraspecifice ale stejarului brumăriu-analiză intra și
interpopulațională.................................................................................................................... 51 20
4.1.3 Analiza comparativă a structurii taxonomice infraspecifice a stejarului pedunculat și
stejarului brumăriu................................................................................................................... 57 22
4.1.4 Structura taxonomică infraspecifică pentru speciile coabitante stejarului brumăriu și
stejarului pedunculat în pădurea de amestec Grivița............................................................... 61 24
4.2 Variabilitatea fenotipică pe baza descriptorilor morfologici ai frunzelor................................. 63 26
4.2.1 Variabilitatea intrapopulațională și interpopulațională a descriptorilor morfologici ai
frunzelor stejarului pedunculat................................................................................................... 63 26
4.2.2 Variabilitatea intrapopulațională și interpopulațională a descriptorilor morfologici ai
frunzelor stejarului brumăriu..................................................................................................... 69 27
4.2.3 Evaluări comparative stejar pedunculat-stejar brumăriu................................................... 74 28
4.2.3.1 Valori medii fenotipice și analiza varianței (ANOVA) a descriptorilor morfologici
ai frunzelor.............................................................................................................................. 75 28
4.2.3.2 Analiza componentelor principale (PCA).................................................................. 83 30
4.2.3.3 Analiza discriminantă................................................................................................. 87 32
4.2.3.4 Analiza cluster............................................................................................................ 91 34

iii
4.2.3.5 Sinteza evaluării morfologice comparative stejar pedunculat - stejar brumăriu
după descriptorii frunzelor..................................................................................................... 95 35
4.3 Analiza fenologiei..................................................................................................................... 98 37
4.3.1 Analiza fenologiei de primăvară privind înfrunzirea și înflorirea..................................... 98 37
4.3.1.1 Analiza fenologiei de primăvară privind înfrunzirea în pădurea de amestec Grivița. 98 37
4.3.1.2 Analiza fenologiei de primăvară privind înflorirea în pădurea de amestec Grivița... 112 42
4.3.1.3 Analiza fenologiei de primăvară privind înfrunzirea în plantajele Mândrești -
Tecuci și Cobadin - Constanța................................................................................................ 116 45
4.3.1.4 Analiza fenologiei de primăvară privind înflorirea în plantajele Mândrești - Tecuci
și Cobadin - Constanța............................................................................................................ 119 46
4.3.2 Analiza comparativă privind fenologia de primăvară în pădurea de amestec Grivița și
în plantajele Mândrești - Tecuci și Cobadin – Constanța........................................................... 121 47
4.3.3 Interpretarea corelativă a datelor din cei 4 ani de observații fenologice în pădurea de
amestec Grivița........................................................................................................................... 122 48
4.3.4 Interpretarea corelativă a datelor din anii de observații fenologice în plantajele Cobadin
și Mândrești................................................................................................................................. 127 49
4.3.5 Interpretarea datelor fenologice în raport cu datele meteorologice................................... 128 50
4.3.6 Analiza xilologică a speciilor de cvercinee din pădurea de amestec Grivița..................... 132 51
4.3.6.1 Activitatea cambială și formarea xilemului în anul 2016........................................... 132 51
4.3.6.2 Evaluări comparative între fenologia înfrunzirii, senescența și fenologia cambială.. 134 52
5. Concluzii. Contribuții originale. Diseminarea rezultatelor. Direcții viitoare de cercetare… 136 54
5.1. Concluzii generale.................................................................................................................... 136 54
5.1.1 Concluzii privind discriminarea taxonomică prin descriptori ai frunzelor la stejarul
pedunculat şi stejarul brumăriu .................................................................................................. 136 54
5.1.2 Concluzii privind analiza fenologiei de primăvară la stejarul pedunculat şi stejarul
brumăriu ..................................................................................................................................... 137 55
5.2 Contribuții originale.................................................................................................................. 140 57
5.3 Diseminarea rezultatelor............................................................................................................ 142 58
5.4 Direcții viitoare de cercetare..................................................................................................... 144 59
Bibliografie......................................................................................................................................... 145 60
Scurt rezumat………………………………………………………………………………………. 181 66
Curriculum vitae................................................................................................................................. 182 67

iv
CONTENTS Pg. Pg.
thesis abstract
Acknowledgements………………………………………………………………………………… 2 1
Introduction …………………………………................................................................................... 3 2
1. Current state of knowledge regarding the taxonomy, chorology, phenological and
phenotypic variability of pedunculate oak (Q. robur L.) and grayish oak (Quercus
pedunculiflora K. Koch)............................................................................................................... 4 3
1.1 Area and ecology of pedunculate oak and grayish oak............................................................. 4 3
1.2 Taxonomic position of pedunculate oak and grayish oak reflected in the literature................. 6 3
1.3 Morphological and ecotypical variability of pedunculate oak (Q. robur L.)............................ 7 4
1.3.1 Phenotypic descriptors and infraspecific variability of pedunculate oak......................... 7 4
1.3.2 Results of studies on the morphological and ecotypic variability of pedunculate oak
(Q. robur L.).............................................................................................................................. 9 4
1.4 Morphological and ecotypical variability of grayish oak (Quercus pedunculiflora K.
Koch)............................................................................................................................................... 10 5
1.4.1 Phenotypic descriptors and infraspecific variability of the grayish oak........................... 10 5
1.4.2 Results of studies on the morphological and ecotypic variability of grayish oak
(Quercus pedunculiflora K. Koch)…………………………………………………………… 13 5
1.5 Phenology of trees..................................................................................................................... 14 5
1.5.1 The importance of studying tree phenology..................................................................... 14 5
1.5.2 Results achieved in Romania and internationally............................................................. 17 6
2. Aim and objectives......................................................................................................................... 20 7
2.1 Research aim............................................................................................................................. 20 7
2.1 Research objectives................................................................................................................... 21 7
3. Research material and method..................................................................................................... 22 8
3.1 Research material...................................................................................................................... 22 8
3.2 Research method …………….....................…………………………………………………. 28 9
3.2.1 Phenotypic analysis.......................................................................................................... 28 9
3.2.2 Analysis of phenology...................................................................................................... 40 15
3.2.3 Phases of annual ring formation....................................................................................... 43 17
4. Results and discussions.................................................................................................................. 45 18
4.1 Intraspecific taxonomical variability– differentiations varieties and forms, depending on the
character of the leaves..................................................................................................................... 45 18
4.1.1 Intraspcific taxonomic units of common oak-intra and inter population analysis.......... 45 18
4.1.2 Intraspcific taxonomic units of grayish oak-intra and inter population analysis............ 51 20
4.1.3 Comparative analysis of intraspecific taxonomic structure of pedunculate oak and
grayish oak .............................................................................................................................. 57 22
4.1.4 Intraspecific taxonomical structure to species cohabiting with pedunculate oak and
grayish oak in the mixed forest Grivita.................................................................................... 61 24
4.2 Phenotypic variability based on morphological descriptors of leaves.. 63 26
4.2.1 Intrapopulational și interpopulațtonal variability of morphological descriptors of leaves
for pedunculate oak..................................................................................................................... 63 26
4.2.2 Intrapopulational și interpopulațtonal variability of morphological descriptors of
leaves for grayish oak.................................................................................................................. 69 27
4.2.3 Comparative evaluations pedunculate oak-grayish oak..................................................... 74 28
4.2.3.1 Phenotypic mean values and variance analysis (ANOVA) of leaf morphological
descriptors............................................................................................................................... 75 28
4.2.3.2 Principal Component Analysis (PCA)........................................................................ 83 30
4.2.3.3 Discriminant analysis.................................................................................................. 87 32
4.2.3.4 Cluster analysis........................................................................................................... 91 34
4.2.3.5 Synthesis of the comparative morphological evaluation of pedunculating oak -
ghrayish oak after leaf descriptors.......................................................................................... 95 35
4.3 Analysis of phenology………………………………………………………………………... 98 37

v
4.3.1 Analysis of spring phenology regarding bud burst and and flowering.............................. 98 37
4.3.1.1 Analysis of spring phenology regarding bud burst in the mixed forest of Grivita..... 98 37
4.3.1.2 Analysis of spring phenology regarding flowering in the mixed forest of Grivita..... 112 42
4.3.1.3 Analysis of spring phenology regarding bud burst in Mândrești - Tecuci and
Cobadin – Constanța seed orchards........................................................................................ 116 45
4.3.1.4 Analysis of spring phenology regarding flowering in Mândrești - Tecuci and
Cobadin – Constanța seed orchards........................................................................................ 119 46
4.3.2 Comparative analysis regarding spring phenology in the mix forest of Grivița and in
Mândrești - Tecuci and Cobadin – Constanța seed orchards...................................................... 121 47
4.3.3 Correlative interpretation of data from the 4 years of phenological observations in the
mixed forest of Grivita................................................................................................................ 122 48
4.3.4 Correlative interpretation of data from years of phenological observations in the
Cobadin and Mândreşti seed orchards........................................................................................ 127 49
4.3.5 Interpretation of phenological data in relation to meteorological data.............................. 128 50
4.3.6 Xylological analysis of the oak species in the mixed forest Grivița.................................. 132 51
4.3.6.1 Cambial activity and xilem fromation in 2016........................................................... 132 51
4.3.6.2 Comparative evaluations on the bud burst phenology, senescence and cambial
phenology................................................................................................................................ 134 52
5. Conclusions. Original contributions. Dissemination of results. Future research directions.. 136 54
5.1 General conclusions………………………………………………………………………… 136 54
5.1.1 Conclusions on taxonomic discrimination through leaf descriptors of pedunculate oak
and grayish oak........................................................................................................................... 136 54
5.1.2 Conclusions on the analysis of spring phenology in pedunculate oak and grayish oak.... 137 55
5.2 Original contributions................................................................................................................ 140 57
5.3 Dissemination of results............................................................................................................ 142 58
5.4 Future research directions.......................................................................................................... 144 59
Bibliography ……………………………………………………………………………………….. 145 60
Short summary………….………………………………………………………………………….. 181 66
Curriculum vitae................................................................................................................................. 183 68

1
Mulțumiri
Această lucrare personală nu ar fi fost posibilă fără sprijinul necontenit al conducătorului științific,
Prof.dr.ing. Neculae ȘOFLETEA, căruia îi adresez întreaga mea recunoștință și sincere mulțumiri pentru
efortul depus, generozitatea, susținerea și răbdarea de care a dat dovadă în îndrumarea competentă și
permanentă pe întreg parcursul elaborării și realizării acestei teze de doctorat, precum și pentru formarea mea
profesională în domeniile dendrologiei și geneticii forestiere.
Prețioase mulțumiri aduc domnului Prof.dr.ing. Lucian CURTU, decanul Facultății de Silvicultură și
Exploatări Forestiere din Brașov, pentru buna colaborare, amabilitatea, sfaturile și recomandările acordate cu
ocazia susținerii rapoartelor științifice, precum şi pentru disponibilitatea domniei sale de a face parte din
comisiile de îndrumare și de analiză a tezei de doctorat.
Îmi exprim gratitudinea față de Prof.dr.ing. Victor PĂCURAR și Conf.dr.ing. Adrian INDREICA,
pentru recomandările exigente și prețioase oferite cu ocazia susținerii referatelor științifice, și pentru
amabilitatea de a face parte din comisia de îndrumare a tezei.
Mă simt onorată și mulțumesc pe această cale domnilor, Prof.dr.ing. Ovidiu IONESCU, Prof.dr.ing.
Gheorghe Florian BORLEA și Conf.dr.ing. Florin CLINOVSCHI, pentru amabilitatea de a accepta să fie
membri în comisia de analiză a tezei de doctorat, în vederea susținerii publice.
Aduc mulțumiri Regiei Naționale a Pădurilor – RNP Romsilva, Direcției Silvice Galați, în special
domnilor, ing. Theodor HOTNOGU, ing. Dragoș BUTUNOI – șef Ocol Silvic Grivița, ing. Radu BUȘCĂ -
șef Ocol Silvic Tecuci, precum și regretatului silvicultor, alături de care am pus bazele acestei cercetări, ing.
Constantin LOGHIN , fost șef al Ocolului Silvic Grivița.
Alese mulţumiri aduc conducerii Institutului Naţional de Cercetare - Dezvoltare în Silvicultură
„Marin Drăcea“, în special domnului Director Științific, CSI dr. ing. Ovidiu BADEA, pentru formarea mea ca
cercetător, încrederea și suportul tehnico-științific permanent, fără de care aceste cercetări nu ar fi fost
posibile, precum și pentru sugestiile valoroase asupra lucrării.
Sincere mulțumiri adresez domnului CSI dr.ing. Flaviu POPESCU, pentru suportul continuu acordat
în cadrul echipei de cercetare, dar și pentru recomandările și sfaturile prețioase.
Îmi exprim aprecierea sinceră și le mulțumesc pentru ajutorul tehnic și încurajările permanente,
colegelor mele, Cristiana DINU și Anca SEMENIUC. Totodată, doresc să le mulțumesc pentru suport,
colegilor mei, Robert IVAN, Marin TUDOROIU, Lucia IONIȚĂ, Diana PITAR, Adrian LORENȚ, Cristian
SIDOR, Andraș TOTPHAL, Alin TOADER.
Va rămâne mereu în amintirea mea și îi sunt profund recunoscătoare pentru tot, domnului
CS I dr. ing. Gheorghe PÂRNUȚĂ, pentru încrederea și dăruința cu care mi-a deschis acest drum al cercetării
și pentru formarea mea ca genetician.
Recunoștință incomensurabilă adresez familiei mele, părinţilor mei și ai soțului meu, surorii și
cumnatului meu, nașilor noștri și celor trei nepoței zglobii, Daria, Maria și Albert, însă, în mod special, îi
mulțumesc soțului meu Bogdan, pentru iubirea profundă pe care mi-o împărtășește, pentru susținerea morală,
răbdarea și înțelegerea manifestată cu generozitate pe întreaga perioadă de desfășurare a tezei de doctorat, și
nu numai.
În memoria bunicului meu, Ion NISTOR, silvicultor de pe Valea Mamului...!

2
INTRODUCERE
În condiţiile actuale privind schimbările globale de mediu, asigurarea adaptabilităţii şi
plasticităţii speciilor forestiere, prin menţinerea unui nivel corespunzător de variabilitate genetică
(Enescu et al. 1997, Ledig 1993), trebuie să constituie o prioritate.
Odată cu aceste schimbări climatice, specii de stejar, în special Quercus robur L. (stejarul
pedunculat) şi Quercus pedunculiflora K. Koch (stejarul brumăriu) fructifică din ce în ce mai rar, cu
o periodicitate chiar de până la 10 ani (Curtu et al. 2015), ceea ce influențează în mod negativ
regenerarea naturală, fiind posibil să asistăm la un regres biologic al speciilor, discutându-se chiar
de o recesiune ontogenetică a acestora (Bândiu et al. 1995). În mod imperios, se impune să se
reflecte asupra variabilităţii cvercineelor în scopul conservării genofondului acestora, a „fondurilor
de gene valoroase caracterizate prin diversitate genetică accentuată, capacitate deosebită de
bioacumulare, calitate excepţională a lemnului şi însuşiri adaptive remarcabile” (Şofletea 2005 citat
de Apostol et al. 2017).
În acest context, cunoașterea fenologiei speciilor de cvercinee și a variabilității fenotipice, în
scopul identificării descriptorilor de diferențiere dintre aceștia, precum și a elucidării, sau nu, a
posibilității interfecundării potențiale dintre acestea, constituie elemente fundamentale în vederea
asigurării unui management corespunzător al ecosistemelor forestiere. De asemenea, formularea de
recomandări privind transferul şi utilizarea în cultură a materialelor forestiere de reproducere pentru
speciile avute în vedere, va contribui la o mai bună utilizare a materialului forestier de reproducere,
la îmbunătăţirea monitorizării. Totodată, vor putea fi evaluate şi prognozate variațiile fenologice în
contextul actual al schimbărilor climatice.
Având în vedere că în România cvercineele sunt specii de mare valoare tehnico-economică,
ecologică, socială și cu rol sanogen important (Beldeanu 2008), prin produsele lemnoase şi
nelemnoase, prin creşterea stabilităţii, calităţii, productivităţii şi rezistenţei la adversităţi a
ecosistemelor forestiere de cvercinee se va asigura ridicarea valorii acestora, menţinerea speciei în
arealul natural de răspândire, creşterea adaptabilităţii şi plasticităţii lor prin asigurarea unei
dezvoltări durabile a pădurii.
Conservarea in situ și ex situ, precum şi monitorizarea celor mai valoroase resurse genetice
forestiere vor avea efecte directe și pozitive asupra productivităţii pădurilor și conservării
biodiversității (Lefort et al. 1998) în contextul bioeconomic actual.

3
1. STADIUL ACTUAL AL CUNOȘTINȚELOR PRIVIND TAXONOMIA,
COROLOGIA, VARIABILITATEA FENOTIPICĂ ŞI FENOLOGICĂ ALE
STEJARULUI PEDUNCULAT (Q. ROBUR L.) ŞI STEJARULUI BRUMĂRIU
(QUERCUS PEDUNCULIFLORA K. KOCH)
1.1 Arealul și ecologia stejarului pedunculat și stejarului brumăriu
Specie cu areal european, stejarul pedunculat (sin. stejar, tufan) se întâlnește în bazinul
Mării Mediterane, se întinde până în Asia Mică și Marea Caspică, urcă spre nord, până în Norvegia
(63°), de unde îşi continuă arealul spre răsărit, de-a lungul paralelei de 60°, până în Munţii Ural şi
coboară înspre sud, pe un traseu relativ ondulat, până la Marea Azov. De aici, arealul său se
derulează spre răsărit, până la Marea Caspică (Șofletea & Curtu 2007).
1.2 Poziția taxonomică a stejarului pedunculat și stejarului brumăriu reflectată în
literatura de specialitate
Genul Quercus aparține familiei Fagaceae și cuprinde în jur de 350 - 500 de specii cu un
ridicat potențial tehnico - economic și ecologic (Kubitzki 1993, Viscosi et al. 2009a). La noi în ţară,
în funcţie de diferite încadrări taxonomice, este reprezentat de 5-9 specii (Georgescu & Morariu
1948, Stănescu et al. 1997, Şofletea & Curtu 2007). Dintre stejarii albi europeni, la noi în țară se
întâlnesc Quercus robur L., Q. petraea (Matt.) Liebl., Q. pubescens Willd., Q. frainetto Ten., Q.
cerris L., Q. faginea Lam., Q. macranthera Fisch. & C.A.Mey, Q. pyrenaica Willd., Q. canariensis
Willd., Q. alnifolia Poech, Q. infectoria Oliv., Q. coccifera L., precum și Q. virgiliana Ten. și Q.
pedunculiflora K. Koch. Quercus robur L. și Quercus pedunculiflora K. Koch sunt specii ce aparțin
subgenului Lepidobalanus (Endl.) Oerst, secția Robur Schb., seria Pedunculatae Schwz și prezintă
caractere morfologice asemănătoare.
Q. pedunculiflora, cea mai valoroasă specie forestieră din silvostepa ţării noastre (Enescu
1993, Apostol et al. 2015, Apostol et al. 2017) este evidenţiat, din punct de vedere taxonomic, ca
specie de sine-stătătoare în diverse lucrări din România, Monografia stejarilor din România
(Georgescu & Morariu 1948), Consideraţiuni sistematice asupra speciei Quercus pedunculiflora K.
Koch în România (Georgescu & Cretzoiu 1941), Flora Ramâniei. Determinator ilustrat al
plantelor vasculare (Beldie 1977), Flora ilustrată a României (Ciocârlan 2000), Habitatele din
România (Doniţă et al. 2005), Fitocenozele din România - Sintaxonomie, Structură şi Evoluţie
(Sanda et al. 2008), Plante vasculare din România (Sârbu et al. 2013), precum și din alte țări ca
Serbia (Vuckovic 1984), Grecia (Athanasiadis et al. 2000) şi chiar în trecut în Flora Europaea
(Schwarz, O. în Tutin et al. 1964). Considerat uneori ca o unitate infraspecifică a lui Quercus robur

4
(Bordács et al. 2002), acesta apare nominalizat ca Q. robur ssp. pedunculiflora (Davis 1965-1988,
Govaerts & Frodin 1998, Ozturk et al. 2004, Broshtilov 2006, Mehrnia et al. 2013).
1.3 Variabilitatea morfologică şi ecotipică a stejarului pedunculat (Q. robur L.)
1.3.1 Descriptori fenotipici şi variabilitatea intraspecifică a stejarului pedunculat
Principalele caractere morfologice de identificare a stejarului pedunculat sunt cele ale
frunzei, ale lujerilor, ale pețiolului şi ale ghindei. Cele mai importante unităţi infraspecifice ale
stejarului pedunculat, specie cu variabilitate intraspecifică mare (Masarovičová 1991) sunt descrise
în Monografia stejarilor din România (Georgescu & Morariu 1948), Flora R.P.R. vol. I (Săvulescu
et al. 1952 ) și în Dendrologie (Şofletea & Curtu 2002), și prezentate detaliat în cadrul capitolului 3
(Materialul şi metoda de cercetare).
Stejarul din ţara noastră, care aparţine unei rase de tranziţie, de la climatul moderat sudic şi vest-
european, la cel aspru din nord-estul arealului speciei, include o serie de populaţii cu caracteristici
de adaptabilitate ecotipică, cum sunt: “Climatipul submezoterm, Climatipul de silvostepă,
Edafotipul de luncă, Edafotipul de terasă, Edafotipul de gleiosoluri, Edafotipul de soluri relativ
alcaline, sărăturoase, Edafotipul de soluri cu drenaj activ” (Şofletea & Curtu 2002).
1.3.2 Rezultate ale studiilor privind variabilitatea morfologică şi ecotipică a stejarului
pedunculat (Q. robur L.)
În Bulgaria, Broshtilov (2006) a studiat variabilitatea formei frunzelor la stejar, caz în care
nu s-au identificat diferenţe cantitative şi morfologice semnificative între Q. robur subsp. longipes
Stev. şi Q. robur. subsp. pedunculiflora K. Koch. Într-un arboret natural, Bacilieri et al. au studiat,
în anul 1994, diferenţele morfologice şi fenologice dintre stejar şi gorun, concluzionându-se faptul
că, analizele morfologice ar trebui suplimentate cu cele moleculare în vederea discriminării
speciilor. Kremer et al. (2002) au evaluat diferenţele morfologice între gorun şi stejar în nouă
arborete mixte de cvercinee din Europa de vest. În urma analizelor făcute, cele nouă arborete mixte
s-au separat în două populaţii corespunzătoare celor două specii de cvercinee. Într-un studiu
morfologic comparativ privind variabilitatea descriptorilor frunzelor la stejarul pedunculat, gorun şi
stejarul pufos din partea de nord-est a Franţei, Depouez şi Badeau (1991) au ajuns la concluzia că
pilozitatea şi prezenţa nervurilor intercalare permit diferenţierea acestor cvercinee, mai ales a
stejarului pedunculat de celelalte două specii. De asemenea, autorii amintiţi au precizat că lungimea
peţiolului şi numărul de nervuri intercalare prezintă o rată de succes de 99% pentru diferenţierea
stejarului pedunculat de gorun. Kleinschmit (1993) a studiat variaţia intraspecifică privind creşterea
şi caracterele adaptative la cvercinee, concluzionând faptul că speciile de stejari europeni
hibridează.

5
1.4 Variabilitatea morfologică şi ecotipică a stejarului brumăriu (Quercus
pedunculiflora K. Koch)
1.4.1 Descriptori fenotipici şi variabilitatea intraspecifică a stejarului brumăriu
Georgescu şi Cretzoiu (1941) au realizat o analiză de detaliu asupra variabilităţii fenotipice a
stejarului brumăriu, prezentând caracterele sale de recunoaştere, formele şi varietăţile pe care acesta
le prezintă în arboretele din ţara noastră, fiind identificate: var. atrichoclados (Borb. et Born.)
Schwz., f. typica Schwz., cu subformele: subf. rotundiloba Georg. et Cretz; subf. obtusiloba
Schwz.; subf. goniolobula (Borb.) Schwz.; f. Borzae Georg. et Cretz., cu subformele: subf.
stellatoides Georg. et Cretz.; subf. acutiloba Georg. et Cretz.; subf. simplex Georg. et Cretz.; subf.
frainettoides Georg. et Cretz.; f. maxima Georg. et Cretz. și var. virescens K.Koch, diferenţiindu-se
formele: f. normalis Georg. et Cretz., f. lobulosa Georg. et Cretz.
1.4.2 Rezultate ale studiilor privind variabilitatea morfologică şi ecotipică a stejarului
brumăriu ( Quercus pedunculiflora K. Koch)
Deşi Quercus pedunculiflora este evidenţiat ca specie de sine stătătoare, există şi părerea că
acesta ar reprezenta o unitate infraspecifică a stejarului pedunculat. Astfel, Gömöry et al. (2001)
aduce în discuţie aprecierile făcute de Kleinschmit et al. (1995) şi respectiv de Steinhoff (1997)
potrivit cărora Quercus pedunculiflora ar trebui sa fie considerat ecotip sau subspecie a lui Quercus
robur. În Monografia stejarilor din România (Georgescu & Morariu 1948) nu se evidenţiază
existenţa hibridului natural dintre cele două specii. Broshtilov (2007) precizează că în Bulgaria nu a
identificat diferenţe morfologice semnificative între Q. robur ssp. longipes şi Q. robur ssp.
pedunculiflora, infirmând astfel datele comunicate în acest sens de Cherniavski (1957 – citat de
Broshtilov 2007). De asemenea, tipul de variaţie a principalilor descriptori ai frunzelor de Q. robur
a fost similar cu cel comunicat de Franjic (1996) pentru populaţiile analizate în Croaţia. Carella
(2013) susține că a identificat pentru prima dată stejarul brumăriu în Italia, determinarea speciei
bazându-se în principal pe caracteristici morfologice ale frunzelor.
1.5 Fenologia arborilor
1.5.1 Importanța studierii fenologiei arborilor
La nivelul arborilor, procesele fenologice, cum ar fi înmugurirea, înfrunzirea, înflorirea,
fructificația, coacerea, senescența și căderea frunzelor sunt controlate atât genetic, cât și de factorii
climatici (Crabbe et al. 2016).
Scenariile actuale arată că, în contextul încălzirii globale, cu temperaturi în creștere și
precipitații în scădere (Dubrovský et al. 2015, IPCC 2013, Simota et al. 2014), speciile forestiere ar
putea migra spre nord sau la o altitudine mai mare, ar putea avea un sezon de creștere mai lung și

6
marcat de prezența fenomenelor climatice extreme, cum ar fi înghețurile târzii și seceta prelungită,
ceea ce ar produce perturbări privind fenologia înfrunzirii și reproducerii, în principal prin creșterea
perioadei fructificațiilor și scăderea intensității acesteia (Borlea 1998, Lechowicz 1995, Norby et al.
2003, Stănescu et al. 1997, Martinik et al. 2014). Se poate astfel afirma că, înțelegerea mecanicistă
a fenologiei arborilor este esențială pentru prognozarea schimbărilor fenofazelor, a reglării ciclului
de carbon și apă dintre sol, vegetație și atmosferă (Menzel & Sparks 2006), precum și pentru
evaluarea impactului bolilor și dăunătorilor asupra arborilor într-un mediu în continuă schimbare.
1.5.2 Rezultate obținute în România și pe plan internațional
Primele observații asupra speciilor de arbori în țara noastră au fost cele făcute în sezonul de
vegetaţie din anul 1938 de către dr. Rădulescu la speciile de cvercinee din Bucureşti. Ca o
continuare a observaţiilor începute în anul 1946, Institutul de Cercetări Silvice a continuat
observaţiile fenologice şi în anul 1949 prin, ing. Bălănică T. şi ing. Tomescu A. S-a observat că la
stejarul pedunculat, în regiunea de câmpie şi de deal din Muntenia începutul înfrunzirii a avut loc în
25 aprilie, iar în Moldova către 3 mai. Începutul înfrunzirii s-a produs după 6-7 zile de la data când
s-au înregistrat, în regiunea respectivă, temperaturi minime cu valori sub 0 grade. Tomescu et al.
(1964) au studiat în anul 1961 influenţa mediului extern şi a celui intern asupra producerii
fenofazelor. Astfel, în funcţie de longitudine, stejarul pedunculat are nevoie de o perioadă de 2,7
zile de la un grad de longitudine la altul. În raport cu zona de vegetaţie, Ştefăneşti şi Bărăgan,
diferenţa la înmugurire a fost de 18 zile pentru stejarul pedunculat. În staţiunea Ştefăneşti stejarul
brumăriu a început să înfrunzească pe 17 aprilie.
În ICP Forests s-au realizat observații fenologice asupra principalelor specii forestiere din
țara noastră, inclusiv asupra cvercineelor, cu scopul determinării poluării aerului, evaluării
schimbărilor climatice și al corelației fenofazelor cu creșterea arborilor, starea de sănătate, litiera
(Badea 1998). În sezonul de vegetaţie 2004, Teodosiu et al. au făcut observaţii fenologice asupra
stejarului pedunculat în staţiunile Simeria şi Ştefăneşti, unde s-au observat diferenţe semnificative
la perioada de colorare a frunzelor - 16 septembrie, respectiv 21 octombrie şi la căderea frunzelor -
23 octombrie, respectiv 5 noiembrie. Crăciunesc (2013) a studiat hibridarea naturală între speciile
de cvercinee existente în fitocenoza Bejan-Deva, recunoscută ca centru de hibridare a speciilor
Quercus robur L. (stejar pedunculat), Q. petraea (Matt.) Liebl. (gorun), Q. frainetto Ten. (gârniţă)
şi Q. pubescens Willd. (stejar pufos) concluzionînd că nu există bariere prezigotice pentru
împiedicarea fenomenului de hibridare naturală și, ca atare, există premisele necesare realizării
acestui fenomen. Bacilieri et al. (1995) au studiat și diferenţele fenologice dintre stejar şi gorun. Ca
un rezultat, se poate spune că în acelaşi an nu se realizează toate încrucişările între indivizi.

7
2. SCOP ŞI OBIECTIVE
2.1 Scopul cercetărilor
Chiar dacă Quercus robur L. și Quercus pedunculiflora K. Koch prezintă o adaptabilitate
diferită, primul fiind mezofit – mezohigrofit și mezoterm (Becker & Levy 1990), chiar eurifit, iar al
doilea termofil și xerofit – mezoxerofit (Beldie 1977, Curtu et al. 2011, Enescu 1993, Tomescu
2000, Sârbu et al. 2013, Apostol et al. 2015), acolo unde se întâlnesc, în special în zona de contact
cu silvostepa externă și în pădurile de luncă cu Populus spp., Salix spp., Alnus glutinosa, din punct
de vedere morfologic, cei doi taxoni se diferențiază foarte greu, sau deloc. În vederea clarificării
statutului taxonomic al celor două specii și de evaluare a diversităţii genetice intra- şi
interpopulaţionale, în ultimul deceniu au fost desfășurate tot mai multe studii de genetică
populațională, însă rezultatele acestora au reliefat un nivel foarte scăzut de diferențiere între cei doi
taxoni, indicele de diferențiere genetică fiind foarte mic: FST = 0.020 (Curtu et al. 2011), FST=0,039
(Crăciunesc et al. 2015). Totodată, lipsa unei bariere reproductive între cei doi taxoni (Steinhoff
1997, Chesnoiu et al. 2009) face posibilă interfecundarea acestora, a apariției hibrizilor
interspecifici, ceea ce implică o identificare și mai greoaie a celor două specii, acolo unde acestea se
întrepătrund.
În acest context, cercetările efectuate cu ocazia elaborării tezei de doctorat au avut ca scop
evaluarea fenotipică la nivelul descriptorilor morfologici foliari ai populaţiilor autohtone de stejar
pedunculat şi stejar brumăriu, cartate ca resurse genetice forestiere (RGF), materiale de bază (MB)
sau surse identificate (SI), în vederea clarificării existenţei unor diferenţe morfologice decelabile
între cei doi taxoni și a stabilirii statutului taxonomic al celor două entităţi, Q. robur L. şi Q.
pedunculiflora K. Koch, considerate uneori specii, alteori unităţi infraspecifice. De asemenea, s-a
urmărit evaluarea nivelului de variabilitate a populaţiilor luate în studiu, atât la nivel interspecific,
cât și, în premieră, la nivelul unităților infraspecifice. Totodată, evaluări fenologice, în special a
celei de primăvară, s-au realizat atât în plantaje pure de stejar pedunculat, de hibridare interspecifică
stejar pedunculat – stejar brumăriu, cât și într-o pădure de amestec, în compoziția căreia se regăsesc
stejarul pedunculat şi stejarul brumăriu, pentru a identifica decalajele sau suprapunerea fenofazelor,
caz în care se poate discuta de problema interfertilităţii potenţiale.
2.2 Obiectivele cercetărilor
În raport cu scopul cercetărilor, obiectivele urmărite au fost:
➢ Evaluarea variabilităţii taxonomice intraspecifice, a nivelului de variabilitate fenotipică (intra-
şi interpopulaţională) la nivelul descriptorilor morfologici foliari.
➢ Identificarea și validarea unor caractere de diferențiere între cei doi taxoni, utilizând analize
statistice multivariate.
8
➢ Analiza fenologiei de primăvară în scopul identificării derulării fenofazelor antezei, pornind
de la considerente că suprapunerea fenofazelor înfloririi ar putea face posibil fluxul de gene
între cei doi taxoni, ceea ce ar presupune posibilitatea interfertilităţii potenţiale, iar decalajul
fenofazelor ar putea sugera individualitatea celor doi taxoni ca specii.
➢ Evaluarea fazelor formării inelului anual și coroborarea cu categoriile de arbori precoci,
intermediari și tardivi.
➢ Identificarea de populații valoroase, după criteriul diversității fenotipice, inclusiv pentru
formularea de recomandări privind strategia de conservare a genofondurilor de stejar
pedunculat și stejar brumăriu din România.
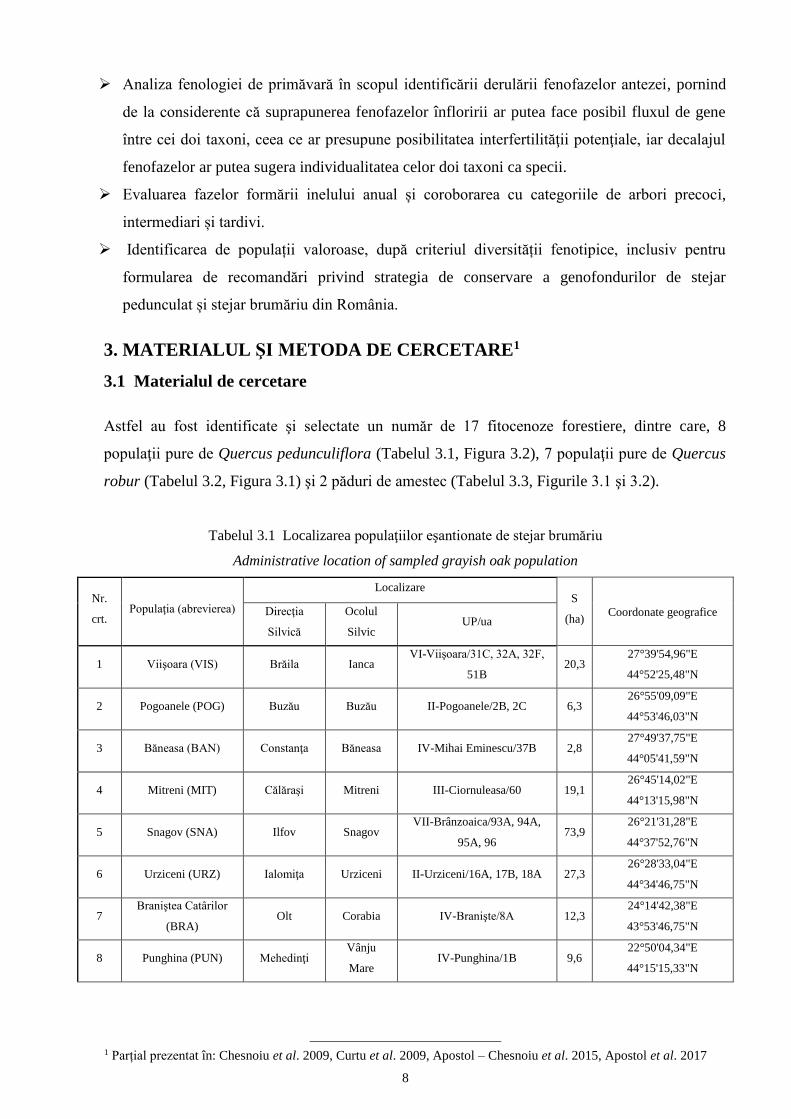
3. MATERIALUL ŞI METODA DE CERCETARE1
3.1 Materialul de cercetare
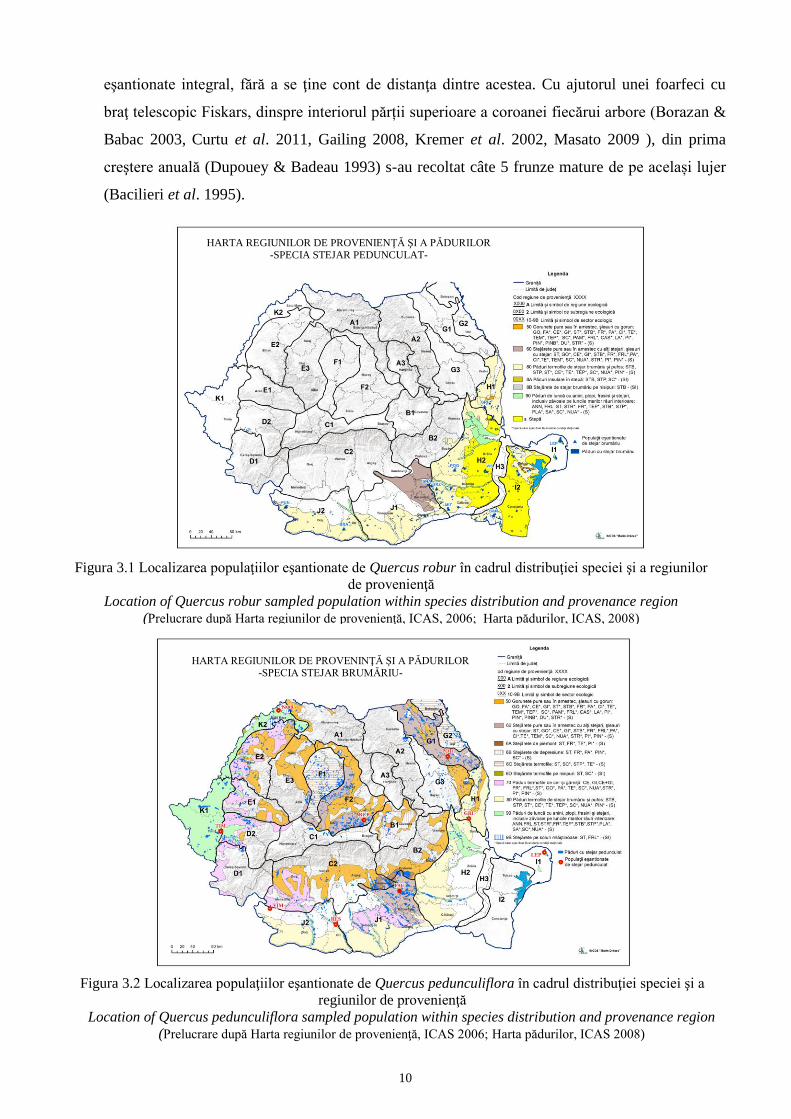
Astfel au fost identificate şi selectate un număr de 17 fitocenoze forestiere, dintre care, 8
populaţii pure de Quercus pedunculiflora (Tabelul 3.1, Figura 3.2), 7 populaţii pure de Quercus
robur (Tabelul 3.2, Figura 3.1) şi 2 păduri de amestec (Tabelul 3.3, Figurile 3.1 şi 3.2).
Tabelul 3.1 Localizarea populaţiilor eşantionate de stejar brumăriu
Administrative location of sampled grayish oak population
Nr.
crt. Populaţia (abrevierea)
Localizare S
(ha) Coordonate geografice Direcţia
Silvică
Ocolul
Silvic UP/ua
1 Viişoara (VIS) Brăila Ianca VI-Viişoara/31C, 32A, 32F,
51B 20,3
27°39'54,96"E
44°52'25,48"N
2 Pogoanele (POG) Buzău Buzău II-Pogoanele/2B, 2C 6,3 26°55'09,09"E
44°53'46,03"N
3 Băneasa (BAN) Constanţa Băneasa IV-Mihai Eminescu/37B 2,8 27°49'37,75"E
44°05'41,59"N
4 Mitreni (MIT) Călăraşi Mitreni III-Ciornuleasa/60 19,1 26°45'14,02"E
44°13'15,98"N
5 Snagov (SNA) Ilfov Snagov VII-Brânzoaica/93A, 94A,
95A, 96 73,9
26°21'31,28"E
44°37'52,76"N
6 Urziceni (URZ) Ialomiţa Urziceni II-Urziceni/16A, 17B, 18A 27,3 26°28'33,04"E
44°34'46,75"N
7 Braniştea Catârilor
(BRA) Olt Corabia IV-Branişte/8A 12,3
24°14'42,38"E
43°53'46,75"N
8 Punghina (PUN) Mehedinţi Vânju
Mare IV-Punghina/1B 9,6
22°50'04,34"E
44°15'15,33"N
1 Parțial prezentat în: Chesnoiu et al. 2009, Curtu et al. 2009, Apostol – Chesnoiu et al. 2015, Apostol et al. 2017
9
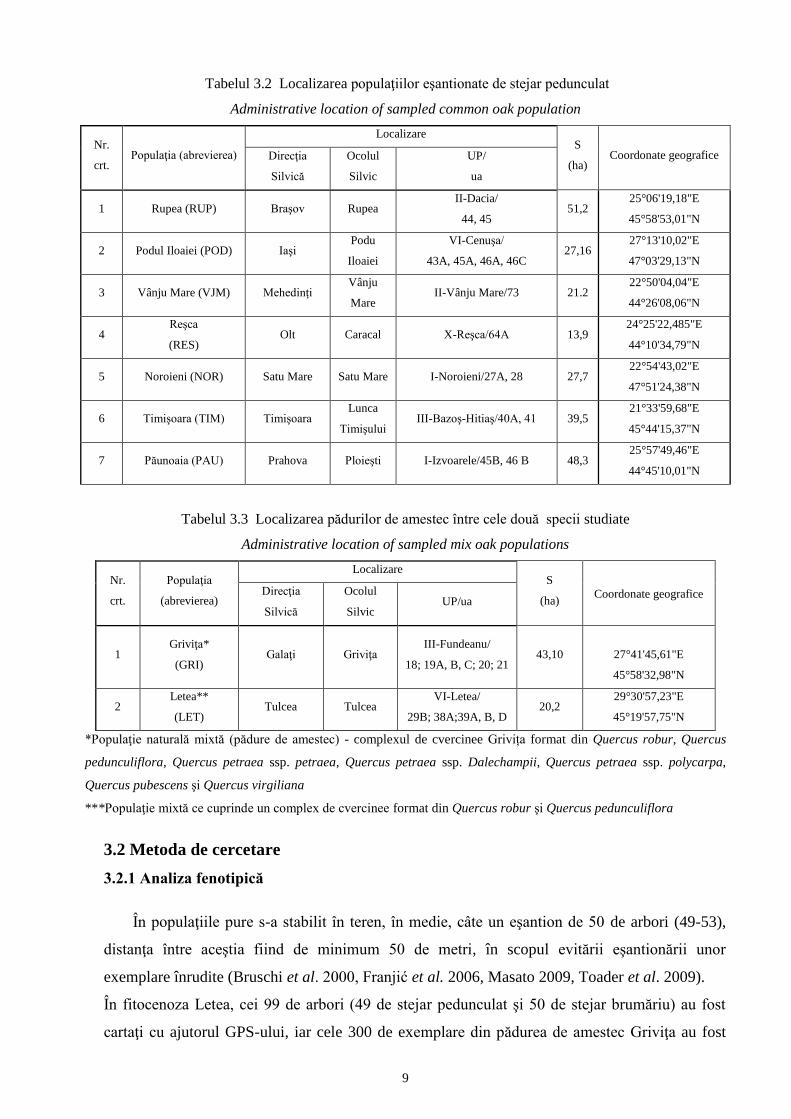
Tabelul 3.2 Localizarea populaţiilor eşantionate de stejar pedunculat
Administrative location of sampled common oak population
Nr.
crt. Populaţia (abrevierea)
Localizare S
(ha) Coordonate geografice Direcţia
Silvică
Ocolul
Silvic
UP/
ua
1 Rupea (RUP) Braşov Rupea II-Dacia/
44, 45 51,2
25°06'19,18"E
45°58'53,01"N
2 Podul Iloaiei (POD) Iaşi Podu
Iloaiei
VI-Cenuşa/
43A, 45A, 46A, 46C 27,16
27°13'10,02"E
47°03'29,13"N
3 Vânju Mare (VJM) Mehedinţi Vânju
Mare II-Vânju Mare/73 21.2
22°50'04,04"E
44°26'08,06"N
4 Reşca
(RES) Olt Caracal X-Reşca/64A 13,9
24°25'22,485"E
44°10'34,79"N
5 Noroieni (NOR) Satu Mare Satu Mare I-Noroieni/27A, 28 27,7 22°54'43,02"E
47°51'24,38"N
6 Timişoara (TIM) Timişoara Lunca
Timişului III-Bazoş-Hitiaş/40A, 41 39,5
21°33'59,68"E
45°44'15,37"N
7 Păunoaia (PAU) Prahova Ploieşti I-Izvoarele/45B, 46 B 48,3 25°57'49,46"E
44°45'10,01"N
Tabelul 3.3 Localizarea pădurilor de amestec între cele două specii studiate
Administrative location of sampled mix oak populations
Nr.
crt.
Populaţia
(abrevierea)
Localizare S
(ha) Coordonate geografice Direcţia
Silvică
Ocolul
Silvic UP/ua
1 Griviţa*
(GRI) Galaţi Griviţa
III-Fundeanu/
18; 19A, B, C; 20; 21 43,10
27°41'45,61"E
45°58'32,98"N
2 Letea**
(LET) Tulcea Tulcea
VI-Letea/
29B; 38A;39A, B, D 20,2
29°30'57,23"E
45°19'57,75"N
*Populaţie naturală mixtă (pădure de amestec) - complexul de cvercinee Grivița format din Quercus robur, Quercus
pedunculiflora, Quercus petraea ssp. petraea, Quercus petraea ssp. Dalechampii, Quercus petraea ssp. polycarpa,
Quercus pubescens şi Quercus virgiliana
***Populaţie mixtă ce cuprinde un complex de cvercinee format din Quercus robur şi Quercus pedunculiflora
3.2 Metoda de cercetare
3.2.1 Analiza fenotipică
În populaţiile pure s-a stabilit în teren, în medie, câte un eşantion de 50 de arbori (49-53),
distanţa între aceştia fiind de minimum 50 de metri, în scopul evitării eşantionării unor
exemplare înrudite (Bruschi et al. 2000, Franjić et al. 2006, Masato 2009, Toader et al. 2009).
În fitocenoza Letea, cei 99 de arbori (49 de stejar pedunculat şi 50 de stejar brumăriu) au fost
cartaţi cu ajutorul GPS-ului, iar cele 300 de exemplare din pădurea de amestec Griviţa au fost
10
eşantionate integral, fără a se ţine cont de distanţa dintre acestea. Cu ajutorul unei foarfeci cu
braţ telescopic Fiskars, dinspre interiorul părții superioare a coroanei fiecărui arbore (Borazan &
Babac 2003, Curtu et al. 2011, Gailing 2008, Kremer et al. 2002, Masato 2009 ), din prima
creştere anuală (Dupouey & Badeau 1993) s-au recoltat câte 5 frunze mature de pe același lujer
(Bacilieri et al. 1995).
Figura 3.1 Localizarea populaţiilor eşantionate de Quercus robur în cadrul distribuţiei speciei şi a regiunilor
de provenienţă
Location of Quercus robur sampled population within species distribution and provenance region
(Prelucrare după Harta regiunilor de provenienţă, ICAS, 2006; Harta pădurilor, ICAS, 2008)
Figura 3.2 Localizarea populaţiilor eşantionate de Quercus pedunculiflora în cadrul distribuţiei speciei şi a
regiunilor de provenienţă
Location of Quercus pedunculiflora sampled population within species distribution and provenance region
(Prelucrare după Harta regiunilor de provenienţă, ICAS 2006; Harta pădurilor, ICAS 2008)
HARTA REGIUNILOR DE PROVENINŢĂ ŞI A PĂDURILOR -SPECIA STEJAR BRUMĂRIU-
HARTA REGIUNILOR DE PROVENIENŢĂ ŞI A PĂDURILOR -SPECIA STEJAR PEDUNCULAT-

11
Cunoscându-se că tipul de părozitate poate fi un indiciu excelent privind hibridizarea şi
introgresia, în funcție de tipul de părozitate (Figura 3.3), încadrarea taxonilor s-a făcut astfel:
stejarul pedunculat este în general glabru, dar prezintă şi părozitate în cadrul var. puberula – peri
simpli; stejarul brumăriu prezintă peri fasciculaţi, în cazul var. virescens doar la inserţia
nervurilor. Stejarul pufos şi italian prezintă puţini peri simpli uniseraţi, peri fasciculaţi şi
fasciculat-bifurcaţi şi un grad ridicat de părozitate, iar gorunul peri stelaţi sau stelat-bifurcaţi
(Fortini et al. 2015).
Figura 3.3 Tipuri de părozitate identificate la taxonii analizați
Trichomes types identified at the analyzed taxa (Foto: Apostol E)
Pentru realizarea analizei variabilităţii taxonomice intraspecifice la nivelul frunzelor, atât
a celor doi taxoni, cât şi a celorlalţi taxoni identificaţi în populaţia mixtă Griviţa, şi anume Q.
petraea ssp. petraea, Q. petraea ssp. dalechampii, Q. petraea ssp. polycarpa, Q. pubescens şi Q.
virgiliana, s-au utilizat unităţile morfologice intraspecifice după frunze ( Tabelele 3.4, 3.5) descrise
în literatura de specialitate (Beldie & Cretzoiu 1941; Georgescu & Cretzoiu 1941; Georgescu &
Morariu 1948; Beldie 1952, 1977; Tătăranu et al. 1960; Stănescu et al. 1997; Şofletea & Curtu
2007). Astfel de unităţi infraspecifice sunt menționate şi în alte studii, sau în literatura de circulaţie
internaţională (Broshtilov 2006).
12
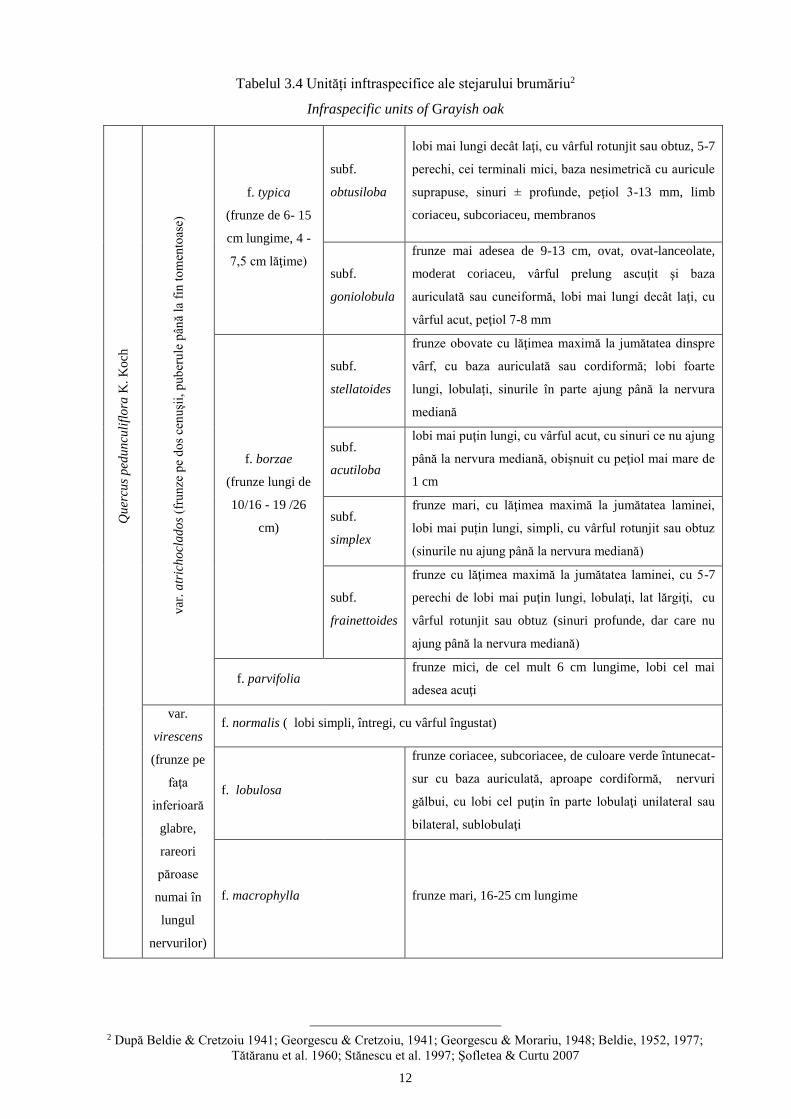
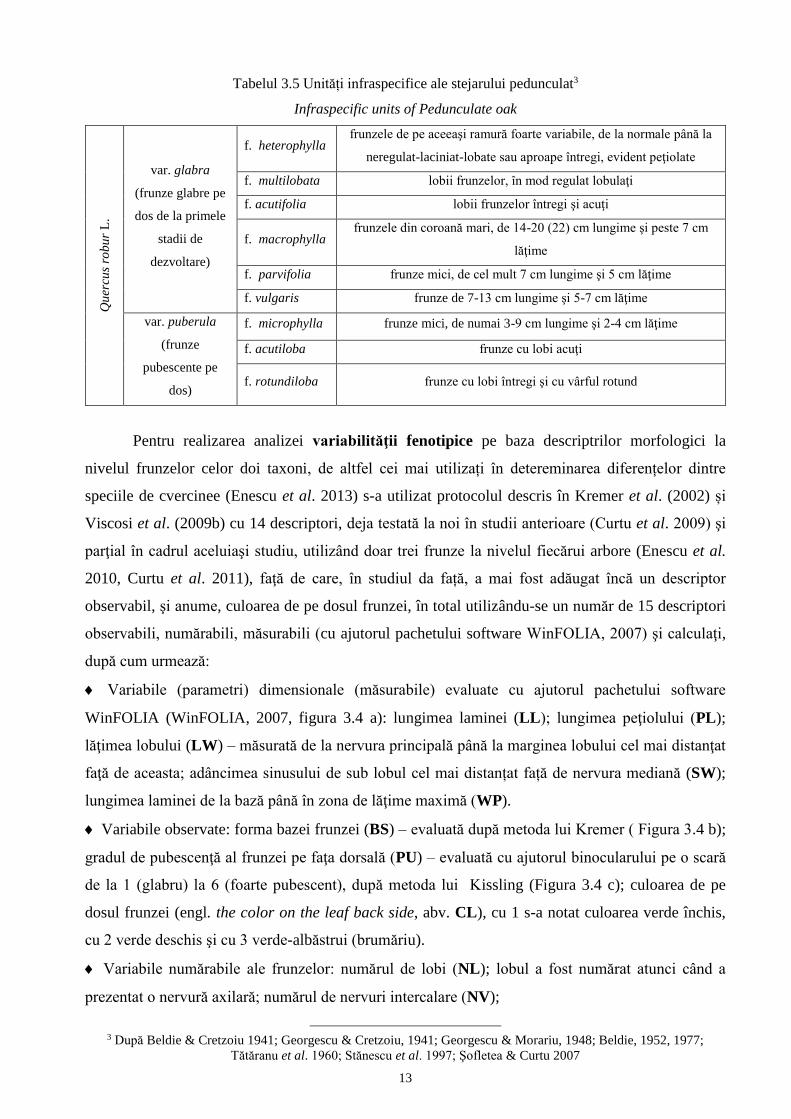
Tabelul 3.4 Unități inftraspecifice ale stejarului brumăriu2
Infraspecific units of Grayish oak Q
uer
cus
ped
un
culi
flo
ra K
. K
och
var
. a
tric
ho
clad
os
(fru
nze
pe
do
s ce
nu
şii,
pub
eru
le p
ână
la f
in t
om
ento
ase)
f. typica
(frunze de 6- 15
cm lungime, 4 -
7,5 cm lăţime)
subf.
obtusiloba
lobi mai lungi decât laţi, cu vârful rotunjit sau obtuz, 5-7
perechi, cei terminali mici, baza nesimetrică cu auricule
suprapuse, sinuri ± profunde, peţiol 3-13 mm, limb
coriaceu, subcoriaceu, membranos
subf.
goniolobula
frunze mai adesea de 9-13 cm, ovat, ovat-lanceolate,
moderat coriaceu, vârful prelung ascuţit şi baza
auriculată sau cuneiformă, lobi mai lungi decât laţi, cu
vârful acut, peţiol 7-8 mm
f. borzae
(frunze lungi de
10/16 - 19 /26
cm)
subf.
stellatoides
frunze obovate cu lăţimea maximă la jumătatea dinspre
vârf, cu baza auriculată sau cordiformă; lobi foarte
lungi, lobulaţi, sinurile în parte ajung până la nervura
mediană
subf.
acutiloba
lobi mai puţin lungi, cu vârful acut, cu sinuri ce nu ajung
până la nervura mediană, obişnuit cu peţiol mai mare de
1 cm
subf.
simplex
frunze mari, cu lăţimea maximă la jumătatea laminei,
lobi mai puțin lungi, simpli, cu vârful rotunjit sau obtuz
(sinurile nu ajung până la nervura mediană)
subf.
frainettoides
frunze cu lăţimea maximă la jumătatea laminei, cu 5-7
perechi de lobi mai puţin lungi, lobulaţi, lat lărgiţi, cu
vârful rotunjit sau obtuz (sinuri profunde, dar care nu
ajung până la nervura mediană)
f. parvifolia
frunze mici, de cel mult 6 cm lungime, lobi cel mai
adesea acuţi
var.
virescens
(frunze pe
faţa
inferioară
glabre,
rareori
păroase
numai în
lungul
nervurilor)
f. normalis ( lobi simpli, întregi, cu vârful îngustat)
f. lobulosa
frunze coriacee, subcoriacee, de culoare verde întunecat-
sur cu baza auriculată, aproape cordiformă, nervuri
gălbui, cu lobi cel puţin în parte lobulaţi unilateral sau
bilateral, sublobulaţi
f. macrophylla frunze mari, 16-25 cm lungime
2 După Beldie & Cretzoiu 1941; Georgescu & Cretzoiu, 1941; Georgescu & Morariu, 1948; Beldie, 1952, 1977;
Tătăranu et al. 1960; Stănescu et al. 1997; Şofletea & Curtu 2007
13
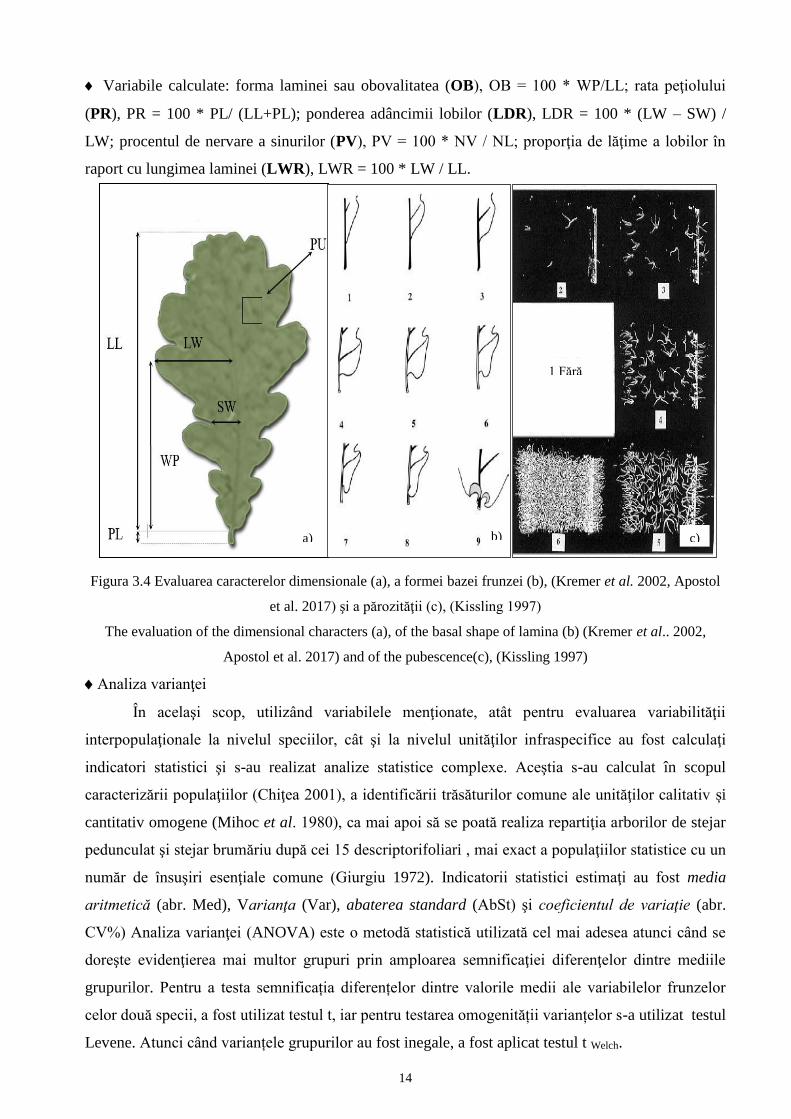
Tabelul 3.5 Unități infraspecifice ale stejarului pedunculat3
Infraspecific units of Pedunculate oak Q
uer
cus
rob
ur
L.
var. glabra
(frunze glabre pe
dos de la primele
stadii de
dezvoltare)
f. heterophylla frunzele de pe aceeaşi ramură foarte variabile, de la normale până la
neregulat-laciniat-lobate sau aproape întregi, evident peţiolate
f. multilobata lobii frunzelor, în mod regulat lobulaţi
f. acutifolia lobii frunzelor întregi şi acuţi
f. macrophylla frunzele din coroană mari, de 14-20 (22) cm lungime şi peste 7 cm
lăţime
f. parvifolia frunze mici, de cel mult 7 cm lungime şi 5 cm lăţime
f. vulgaris frunze de 7-13 cm lungime şi 5-7 cm lăţime
var. puberula
(frunze
pubescente pe
dos)
f. microphylla frunze mici, de numai 3-9 cm lungime şi 2-4 cm lăţime
f. acutiloba frunze cu lobi acuţi
f. rotundiloba frunze cu lobi întregi şi cu vârful rotund
Pentru realizarea analizei variabilităţii fenotipice pe baza descriptrilor morfologici la
nivelul frunzelor celor doi taxoni, de altfel cei mai utilizați în detereminarea diferențelor dintre
speciile de cvercinee (Enescu et al. 2013) s-a utilizat protocolul descris în Kremer et al. (2002) și
Viscosi et al. (2009b) cu 14 descriptori, deja testată la noi în studii anterioare (Curtu et al. 2009) şi
parţial în cadrul aceluiaşi studiu, utilizând doar trei frunze la nivelul fiecărui arbore (Enescu et al.
2010, Curtu et al. 2011), faţă de care, în studiul da față, a mai fost adăugat încă un descriptor
observabil, şi anume, culoarea de pe dosul frunzei, în total utilizându-se un număr de 15 descriptori
observabili, numărabili, măsurabili (cu ajutorul pachetului software WinFOLIA, 2007) şi calculaţi,
după cum urmează:
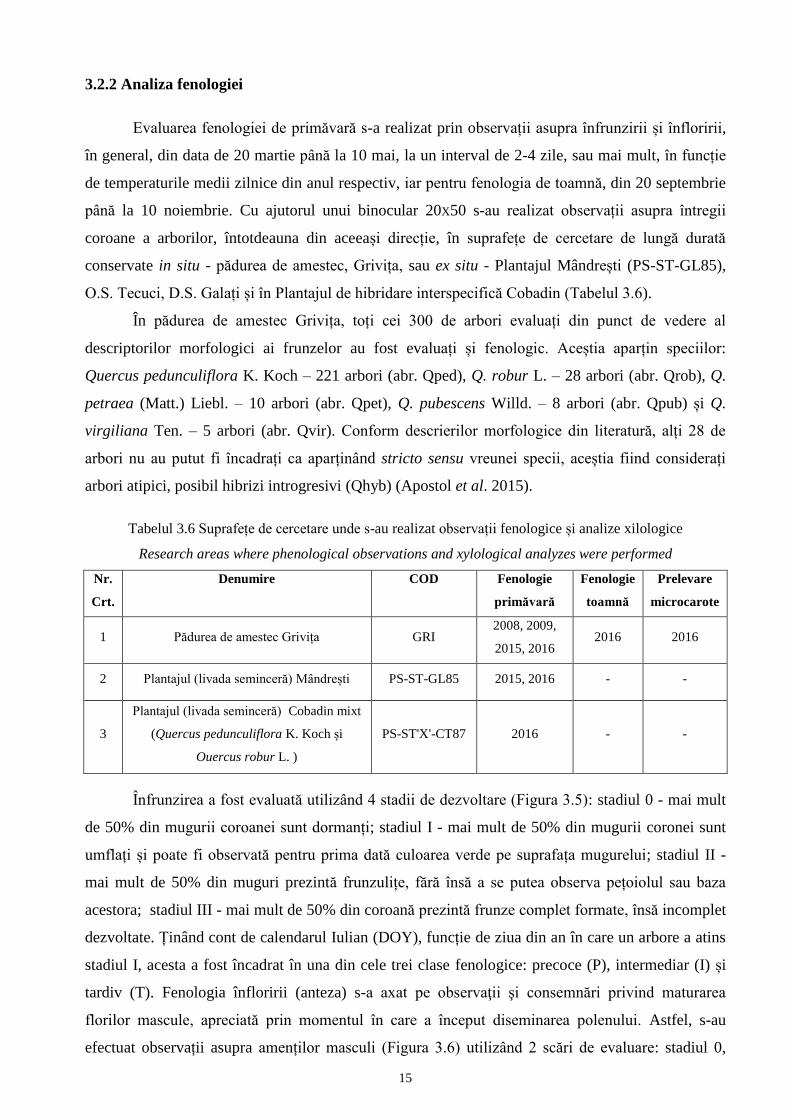
Variabile (parametri) dimensionale (măsurabile) evaluate cu ajutorul pachetului software
WinFOLIA (WinFOLIA, 2007, figura 3.4 a): lungimea laminei (LL); lungimea peţiolului (PL);
lăţimea lobului (LW) – măsurată de la nervura principală până la marginea lobului cel mai distanţat
faţă de aceasta; adâncimea sinusului de sub lobul cel mai distanțat față de nervura mediană (SW);
lungimea laminei de la bază până în zona de lăţime maximă (WP).
Variabile observate: forma bazei frunzei (BS) – evaluată după metoda lui Kremer ( Figura 3.4 b);
gradul de pubescenţă al frunzei pe faţa dorsală (PU) – evaluată cu ajutorul binocularului pe o scară
de la 1 (glabru) la 6 (foarte pubescent), după metoda lui Kissling (Figura 3.4 c); culoarea de pe
dosul frunzei (engl. the color on the leaf back side, abv. CL), cu 1 s-a notat culoarea verde închis,
cu 2 verde deschis şi cu 3 verde-albăstrui (brumăriu).
Variabile numărabile ale frunzelor: numărul de lobi (NL); lobul a fost numărat atunci când a
prezentat o nervură axilară; numărul de nervuri intercalare (NV);
3 După Beldie & Cretzoiu 1941; Georgescu & Cretzoiu, 1941; Georgescu & Morariu, 1948; Beldie, 1952, 1977;
Tătăranu et al. 1960; Stănescu et al. 1997; Şofletea & Curtu 2007
14
Variabile calculate: forma laminei sau obovalitatea (OB), OB = 100 * WP/LL; rata peţiolului
(PR), PR = 100 * PL/ (LL+PL); ponderea adâncimii lobilor (LDR), LDR = 100 * (LW – SW) /
LW; procentul de nervare a sinurilor (PV), PV = 100 * NV / NL; proporţia de lăţime a lobilor în
raport cu lungimea laminei (LWR), LWR = 100 * LW / LL.
Figura 3.4 Evaluarea caracterelor dimensionale (a), a formei bazei frunzei (b), (Kremer et al. 2002, Apostol
et al. 2017) şi a părozităţii (c), (Kissling 1997)
The evaluation of the dimensional characters (a), of the basal shape of lamina (b) (Kremer et al.. 2002,
Apostol et al. 2017) and of the pubescence(c), (Kissling 1997)
Analiza varianţei
În acelaşi scop, utilizând variabilele menţionate, atât pentru evaluarea variabilităţii
interpopulaţionale la nivelul speciilor, cât şi la nivelul unităţilor infraspecifice au fost calculaţi
indicatori statistici şi s-au realizat analize statistice complexe. Aceştia s-au calculat în scopul
caracterizării populaţiilor (Chiţea 2001), a identificării trăsăturilor comune ale unităţilor calitativ și
cantitativ omogene (Mihoc et al. 1980), ca mai apoi să se poată realiza repartiţia arborilor de stejar
pedunculat şi stejar brumăriu după cei 15 descriptorifoliari , mai exact a populaţiilor statistice cu un
număr de însuşiri esenţiale comune (Giurgiu 1972). Indicatorii statistici estimaţi au fost media
aritmetică (abr. Med), Varianţa (Var), abaterea standard (AbSt) şi coeficientul de variaţie (abr.
CV%) Analiza varianţei (ANOVA) este o metodă statistică utilizată cel mai adesea atunci când se
doreşte evidenţierea mai multor grupuri prin amploarea semnificaţiei diferenţelor dintre mediile
grupurilor. Pentru a testa semnificația diferențelor dintre valorile medii ale variabilelor frunzelor
celor două specii, a fost utilizat testul t, iar pentru testarea omogenității varianțelor s-a utilizat testul
Levene. Atunci când varianțele grupurilor au fost inegale, a fost aplicat testul t Welch.
1 Fără
părozitate
a) b) c)
15
3.2.2 Analiza fenologiei
Evaluarea fenologiei de primăvară s-a realizat prin observații asupra înfrunzirii și înfloririi,
în general, din data de 20 martie până la 10 mai, la un interval de 2-4 zile, sau mai mult, în funcție
de temperaturile medii zilnice din anul respectiv, iar pentru fenologia de toamnă, din 20 septembrie
până la 10 noiembrie. Cu ajutorul unui binocular 20X50 s-au realizat observații asupra întregii
coroane a arborilor, întotdeauna din aceeași direcție, în suprafețe de cercetare de lungă durată
conservate in situ - pădurea de amestec, Grivița, sau ex situ - Plantajul Mândrești (PS-ST-GL85),
O.S. Tecuci, D.S. Galați și în Plantajul de hibridare interspecifică Cobadin (Tabelul 3.6).
În pădurea de amestec Grivița, toți cei 300 de arbori evaluați din punct de vedere al
descriptorilor morfologici ai frunzelor au fost evaluați și fenologic. Aceștia aparțin speciilor:
Quercus pedunculiflora K. Koch – 221 arbori (abr. Qped), Q. robur L. – 28 arbori (abr. Qrob), Q.
petraea (Matt.) Liebl. – 10 arbori (abr. Qpet), Q. pubescens Willd. – 8 arbori (abr. Qpub) și Q.
virgiliana Ten. – 5 arbori (abr. Qvir). Conform descrierilor morfologice din literatură, alți 28 de
arbori nu au putut fi încadrați ca aparținând stricto sensu vreunei specii, aceștia fiind considerați
arbori atipici, posibil hibrizi introgresivi (Qhyb) (Apostol et al. 2015).
Tabelul 3.6 Suprafețe de cercetare unde s-au realizat observații fenologice și analize xilologice
Research areas where phenological observations and xylological analyzes were performed
Nr.
Crt.
Denumire COD Fenologie
primăvară
Fenologie
toamnă
Prelevare
microcarote
1 Pădurea de amestec Grivița GRI 2008, 2009,
2015, 2016 2016 2016
2 Plantajul (livada seminceră) Mândrești PS-ST-GL85 2015, 2016 - -
3
Plantajul (livada seminceră) Cobadin mixt
(Quercus pedunculiflora K. Koch și
Ouercus robur L. )
PS-ST'X'-CT87 2016 - -
Înfrunzirea a fost evaluată utilizând 4 stadii de dezvoltare (Figura 3.5): stadiul 0 - mai mult
de 50% din mugurii coroanei sunt dormanți; stadiul I - mai mult de 50% din mugurii coronei sunt
umflați și poate fi observată pentru prima dată culoarea verde pe suprafața mugurelui; stadiul II -
mai mult de 50% din muguri prezintă frunzulițe, fără însă a se putea observa pețoiolul sau baza
acestora; stadiul III - mai mult de 50% din coroană prezintă frunze complet formate, însă incomplet
dezvoltate. Ținând cont de calendarul Iulian (DOY), funcție de ziua din an în care un arbore a atins
stadiul I, acesta a fost încadrat în una din cele trei clase fenologice: precoce (P), intermediar (I) și
tardiv (T). Fenologia înfloririi (anteza) s-a axat pe observaţii şi consemnări privind maturarea
florilor mascule, apreciată prin momentul în care a început diseminarea polenului. Astfel, s-au
efectuat observații asupra amenților masculi (Figura 3.6) utilizând 2 scări de evaluare: stadiul 0,

16
când peste 50% din amenți sunt de culoare verde-gălbui, alungiți, însă nu eliberează polen și stadiul
I, când peste 50% din amenți sunt complet alungiți, complet pendenți, galben-maronii și eliberează
polen.
Pentru fenologia de toamnă s-au utilizat 3 stadii: stadiul I, când peste 50% din frunzele
coroanei arborelui sunt verde-închis, însă prezintă primele pete de culoare galbenă; stadiul II, când
peste 50% din frunzele coroanei arborelui sunt verde-deschis și prezintă numeroase pete de culoare
galbenă; stadiul III, când peste 50% din frunzele coroanei arborelui sunt galben-maronii.
Figura 3.5 Stadiile de dezvoltare privind înfrunzirea cvercineelor, cu cele 4 scări de evaluare
The main stages of bud flushing for oak and the corresponding 4 grades- scoring scale (Foto:Apostol E.)
Foto: Apostol E.
Figura 3.6 Stadiile de înflorire ale florilor mascule, cu cele 2 scări de evaluare
Two main stages of male flowering for oak and the corresponding 2 grades - scoring scale
Foto: Apostol E.
Figura 3.7 Stadiile de colorare a frunzelor, cu 3 scări de evaluare
Three main stages of senescence for oak and the corresponding 3 grades- scoring scale
Stadiul 0 Stadiul I Stadiul II Stadiul III

17
3.2.3 Fazele formării inelului anual
Analiza fazelor de formare a inelului anual s-a realizat prin metoda xilologică de analiză a
microcarotelor de creștere (Popa & Semeniuc 2009), prin această metodă putându-se obține
informații cu privire la perioada formării inelului anual, precum și la numărul de celule aflate în
diferite faze de dezvoltare (Antonova & Stasova 1997, Rossi et al. 2006, Popa & Semeniuc 2009).
Cu ajutorul unui burghiu de dimensiuni mici (Trephor) (Rossiet et al. 2006), pe parcursul sezonului
de vegetație al anului 2016, și la un interval de 2 săptămâni, s-au prelevat microcarote (1-2 mm
grosime și 1-2 cm lungime) (Popa & Semeniuc 2009) din 16 arbori (Q. pedunculiflora, arborele -
18; Q. pubescens, arborele - 20; Q. pedunculiflora , arborele – 21; Q. robur, arborele - 22; Q.
petraea, arborele - 129; Q. pedunculiflora, arborele – 164; Q. robur, arborele – 166; Q.
pedunculiflora, arborele - 167; exemplar atipic, arborele - 168; exemplar atipic, arborele - 179;
exemplar atipic, arborele - 180; exemplar atipic, arborele - 182, Q. pedunculiflora, arborele – 293;
Q. pedunculiflora, arborele – 294; Q. robur, arborele - 295; Q. pedunculiflora, arborele - 296). La
alegerea arborilor s-a ținut cont ca fiecare taxon să fie reprezentat de arbori din toate categoriile
fenologice (aferente anilor de observații) și să respecte proporția de reprezentare în fitocenoză.
Probele prelevate s-au introdus în microtuburi de plastic umplute cu amestec de alcool și apă în
proporție de 1:1, în scopul evitării deshidratării acestora (Popa & Semeniuc 2009. Microcarotele au
fost păstrate în biocasete speciale și introduse în procesator de țesuturi, având rol în eliminarea apei
din celule şi de fixare a parafinei în elementele anatomice ale lemnului și imediat scufundate în
RotiClear și parafină (Rossi et al. 2006). Următoarea etapă a constat în realizarea microsecţiunilor
cu ajutorul microtomului rotativ semiautomat (Microm HM335). În procedura de colorare s-au
folosit suporții de lame microscopice, recipienți de sticlă și substanțe chimice. S-au identificat patru
faze de formare a inelului anual: activitatea cambială, dezvoltarea celulelor, lignificare și maturare
celule (Rossi et al. 2006, Popa & Semeniuc 2009).

18
4. REZULTATE ŞI DISCUŢII
4.1 Variabilitatea taxonomică intraspecifică – diferențieri de varietăți și forme după
caractere ale frunzelor
Evaluarea variabilităţii intrapopulaţionale pentru 8 populaţii de stejar pedunculat (Rupea,
Podul Iloaiei, Vânju Mare, Reşca, Noroieni, Timişoara, Păunoaia, Letea robur) şi 9 populaţii de
stejar brumăriu (Viişoara, Pogoanele, Băneasa, Mitreni, Snagov, Urziceni, Braniştea Catârilor,
Punghina, Letea pedunculiflora) s-a realizat prin identificarea varietăţilor, formelor și subformelor
frunzelor conform unităţilor morfologice infraspecifice descrise în literatura de specialitate şi
sintetizate în metoda de lucru, capitolul 3.2.
4.1.1 Unități taxonomice infraspecifice ale stejarului pedunculat - analiză intra- și
interpopulațională
În vederea încadrării în unităţi infraspecifice a arborilor de stejar pedunculat, au fost
analizate 2030 de frunze, câte 5 frunze din fiecare din cei 406 arbori. Dintre cele nouă unităţi
infraspecifice corespunzătoare morfologiei frunzei (var. glabra cu f. heterophylla, f. multilobata, f.
acutifolia, f. macrophylla, f. parvifolia, f. vulgaris şi var. puberula cu f. acutiloba, f. microphylla,
f. rotundiloba), în nicio populaţie nu au fost identificate frunze care să aparțină var. glabra, f.
parvifolia, toate celelalte varietăți fiind prezente (Figura 4.1). Din cei 406 arbori, la 19 dintre aceştia
nu toate cele 5 frunze au prezentat caractere specifice unei singure unităţi, aşa încât au fost
consideraţi arbori cu caractere intermediare (INT) şi au fost alocaţi unei unităţii infraspecifice sau
alteia prin asimilare cu taxonul față de care au prezentat cea mai mare similitudine a descriptorilor
de bază utilizaţi în determinarea taxonilor: Rupea - 7 arbori, dintre care 1 exemplar f. acutifolia, 2
exemplare f. macrophylla şi 4 exemplare f. vulgaris; Podul Iloaiei - 3 arbori asimilaţi f. vulgaris;
Vânju Mare - 4 arbori f. vulgaris; Reşca - 2 arbori, din care 1 exemplar f. macrophylla şi unul f.
vulgaris; Noroieni - 1 arbore asimilat f. acutifolia; Letea robur 2 arbori asimilaţi f. multilobata;
Timişoara şi Păunoaia nu au prezentat arbori intermediari. În cei 406 arbori din populaţiile de stejar
pedunculat, forma acutifolia a fost cel mai bine reprezentată, fiind identificată la 103 exemplare
(25,4% din totalul arborilor analizați). Formele multilobata şi vulgaris au fost, de asemenea, bine
reprezentate, fiind identificate, câte 92 (22,7%) şi respectiv 90 de exemplare (22,2%) pentru
fiecare unitate infraspecifică.

19
Figura 4.1 Varietăţile şi formele identificate în populaţiile pure la Quercus robur L. (Foto: Apostol E.)
Varieties and shapes identified in Quercus robur L pure populations
Tabelul 4.1 Distribuția arborilor analizați pe varietăţi identificate în populaţiile de stejar pedunculat
Tree distribution based on varieties identified for pedunculate oak
Quercus robur L.
Populaţia
var
.
gla
bra
var
.
pu
ber
ula
To
tal
arb
ori
p
op
ula
ţii
f. h
eter
op
hyl
la
f. m
ult
ilo
ba
ta
f. a
cuti
foli
a
f. m
acr
op
hyl
la
f. v
ulg
ari
s
f. a
cuti
loba
f. m
icro
ph
ylla
f. r
otu
nd
ilob
a
RUP 2 10 9 4 16 0 0 9 50
POD 8 13 9 2 4 0 0 14 50
VJM 6 12 15 0 14 0 0 6 53
RES 0 12 17 8 7 0 0 7 51
NOR 3 20 8 0 8 3 2 7 51
TIM 5 9 24 2 8 2 0 0 50
LER 6 13 7 0 12 3 0 8 49
PAU 5 3 14 9 21 0 0 0 52
Total arbori pe unităţi 35 92 103 25 90 8 2 51 406
În pădurea de amestec Grivița, studiat în cadrul UP III Fundeanu, din totalul celor 300 de
arbori eşantionaţi, 28 de arbori (9,3% din totalul arborilor analizați, inclusiv speciile coabitante)
sunt Quercus robur (stejar pedunculat). Dintre cele nouă tipuri de unităţi infraspecifice
caracteristice stejarului pedunculat, arborii au fost asimilaţi la şapte dintre acestea (Tabelul 4.2). F.
parvifolia nu a fost identificată nici în populaţiile pure de stejar pedunculat.

20
Tabelul 4.2 Distribuția arborilor de stejar pedunculat analizați pe varietăţi, în pădurea de amestec Grivița
Tree distribution based on varieties identified in Griviţa mix forest
Quercus robur L. Total
ST
Po
pu
laţi
a
Gri
viţ
a var. glabra var. puberula
f.
heterophylla
f.
multilobata
f.
acutifolia
f.
macrophylla
f.
vulgaris
f.
acutiloba
f.
rotundiloba
Nr. Arb. 6 5 6 1 5 2 3 28
4.1.2 Unități taxonomice infraspecifice ale stejarului brumăriu – analiză intra și
interpopulațională
În vederea încadrării în unităţi infraspecifice, în fitocenozele pure de stejar brumăriu au fost
analizate 2030 de frunze, câte 5 frunze din fiecare cei 406 arbori eşantionaţi în populaţiile
Punghina, Băneasa, Braniştea Catârilor, Mitreni, Pogoanele, Snagov, Urziceni şi Viişoara şi 250 de
frunze din cei 50 de arbori în fitocenoza mixtă Letea pedunculiflora, în total 2280 de frunze. Dintre
unitățile infraspecifice corespunzătoare, în populațiile cercetate au fost identificate: varietatea
atrichoclados cu forma typica- subformele obtusiloba şi goniolobula, forma maxima – subformele
stellatoides, acutiloba, simplex şi frainettoides (Figura 4.2). De menționat este faptul că, față de
descrierea din literatura de specialitate, forma maxima, subforma frainettoides prezintă cel mai
adesea frunze obovate, și nu eliptice (Figura 4.2), care pot avea dimensiuni mai mici de 10 cm, ca în
cazul celor din populația Băneasa – Constanța, o explicație putând fi, pentru cel din urmă caz,
condițiile staționale deficitare în precipitații din sezonul de vegetație. La forma typica, subforma
obtusiloba au fost identificate și frunze care prezintă doar 4 perechi de lobi, adesea lobulați inferior.
Dintre cei 456 de arbori supuşi analizelor, 45 dintre aceştia (circa 10%) nu au toate cele 5
frunze cu caractere specifice unei singure unităţi, aşa încât au fost consideraţi arbori cu caractere
intermediare (INT) şi au fost alocaţi unei unităţi infraspecifice sau alteia, funcţie de caracteristicile
frunzelor majoritare. În populaţiile Punghina, Snagov şi în populaţia Letea pedunculiflora nu au fost
identificaţi arbori intermediari; populaţia Băneasa are cel mai mare număr de arbori intermediari –
11 (2,4%) dintre care, 7 arbori au fost asimilaţi subformei obtusiloba şi 4 subformei frainettoides;
populaţia Braniştea Catârilor prezintă 8 arbori intermediari (1,7%), 4 asimilaţi subformei
obtusiloba, şi câte doi asimilaţi subformelor stellatiodes şi simplex; din cei 9 arbori (2%)
intermediai ai populaţiei Mitreni, 5 au fost asimilaţi subformei obtusiloba şi câte 2 subformelor
simplex şi frainettoides; cu un arbore mai puţin decât în populaţia Băneasa, populaţia Pogoanele are
10 arbori intermediari (2,2%), dintre care 4 au frunze corespunzătoare subformei stellatoides, 3
subformei obtusiloba, 2 subformei frainettoides şi 1 subformei simplex; cel mai mic număr de
arbori intermediari a fost identificat în populaţia Urziceni - 1 arbore (0,2%) aparținând f. maxima,
21
subf. simplex; populaţia Viişoara prezintă 4 arbori asimilaţi subf. stellatoides şi 2 asimilaţi subf.
frainettoies.
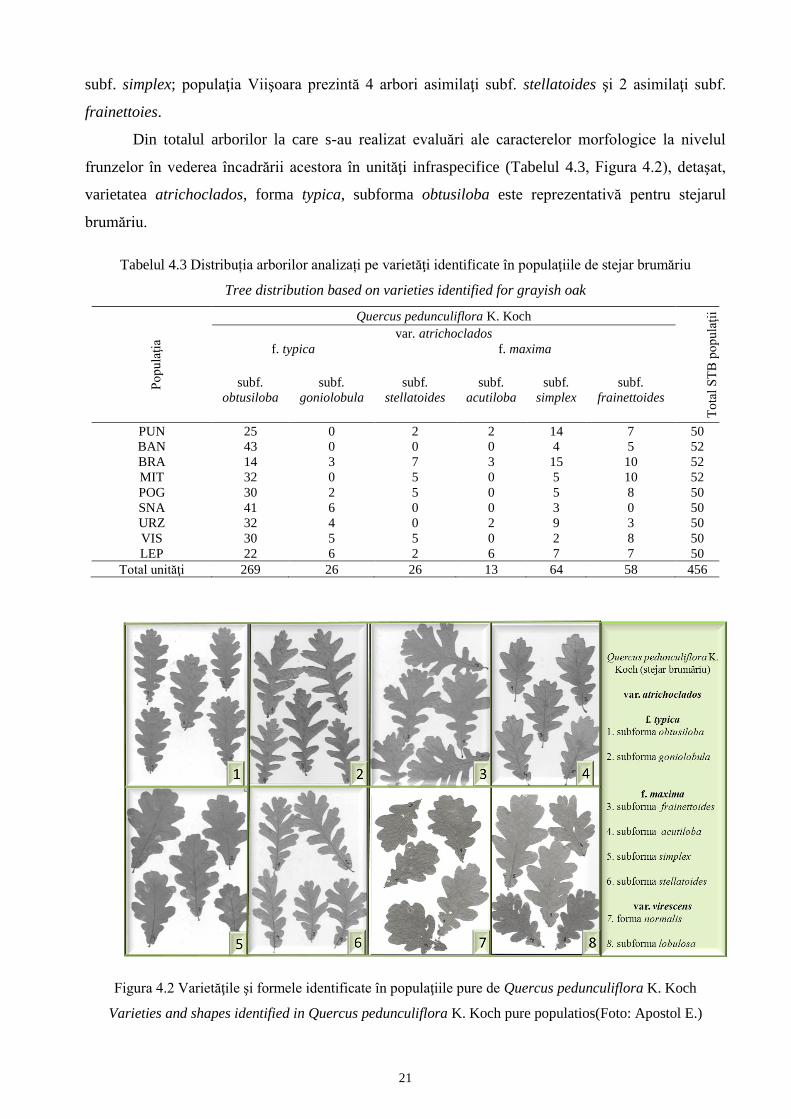
Din totalul arborilor la care s-au realizat evaluări ale caracterelor morfologice la nivelul
frunzelor în vederea încadrării acestora în unităţi infraspecifice (Tabelul 4.3, Figura 4.2), detaşat,
varietatea atrichoclados, forma typica, subforma obtusiloba este reprezentativă pentru stejarul
brumăriu.
Tabelul 4.3 Distribuția arborilor analizați pe varietăţi identificate în populaţiile de stejar brumăriu
Tree distribution based on varieties identified for grayish oak
Po
pu
laţi
a
Quercus pedunculiflora K. Koch
To
tal
ST
B p
op
ula
ţii
var. atrichoclados
f. typica f. maxima
subf.
obtusiloba
subf.
goniolobula
subf.
stellatoides
subf.
acutiloba
subf.
simplex
subf.
frainettoides
PUN 25 0 2 2 14 7 50
BAN 43 0 0 0 4 5 52
BRA 14 3 7 3 15 10 52
MIT 32 0 5 0 5 10 52
POG 30 2 5 0 5 8 50
SNA 41 6 0 0 3 0 50
URZ 32 4 0 2 9 3 50
VIS 30 5 5 0 2 8 50
LEP 22 6 2 6 7 7 50
Total unităţi 269 26 26 13 64 58 456
Figura 4.2 Varietăţile şi formele identificate în populaţiile pure de Quercus pedunculiflora K. Koch
Varieties and shapes identified in Quercus pedunculiflora K. Koch pure populatios(Foto: Apostol E.)

22
În populaţia mixtă Griviţa, cu o variabilitate intrapopulaţională foarte ridicată, cei 221 de
stejari brumării prezintă aproape toate unităţile infraspecifice caracteristice acestuia. Spre
deosebire de celelalte populaţii, în aceasta este prezentă var. virescens, cu f. normalis (Tabelul
4.4). Această varietate a fost identificată şi în pădurile cu stejar brumăriu de pe raza Ocolului
Silvic Huşi (Dumitriu - Tătăranu 1954).
Tabelul 4.4 Distribuția arborilor analizați pe varietăţi de frunze identificate în pădurea de amestec Griviţa
Tree distribution based on leaf varieties identified in Griviţa mix forest
Quercus pedunculiflora K. Koch Total STB
var. atrichoclados var. virescens
f. typica f. maxima f. normalis
subf.
obtusiloba
subf.
goniolobula
subf.
stellatoides
subf.
acutiloba
subf.
simplex
subf.
frainettoides
88 12 6 13 70 27 5 221
4.1.3 Analiza comparativă a structurii taxonomice intraspecifice a stejarului pedunculat și
stejarului brumăriu
Făcând o analiză la nivelul ponderilor unităţilor taxonomice infraspecifice la stejarul
pedunculat, în toate populaţiile studiate, se poate constata că, variabilitatea cea mai ridicată este
întâlnită în populaţiile Noroieni şi Griviţa, aici fiind prezente 7 din cele 9 unităţi infraspecifice
(Figura 4.3 a). Dacă var. glabra, f. parvifolia lipseşte în ambele populaţii, iar în populaţia Griviţa
mai lipseşte var. puberula f. microphylla, în populaţia Noroieni cea de-a doua unitate infraspecifică
absentă este f. macrophylla, și mai mult, este singura populaţie în care a fost întâlnită var. puberula
f. microphylla.
Populaţiile Rupea, Podul Iloaiei, Populaţia Letea robur şi Timişoara au aceași pondere a
unităţilor infraspecifice identificate (66%). Populaţiile cu cele mai puţine tipuri de unităţi
infraspecifice sunt Vânju Mare, Reşca şi Păunoaia. În cea din urmă populaţie nu au fost identificate
exemplare asimilabile var. puberula. Varietatea glabra cu f. multilobata, f. acutifolia şi f. vulgaris a
fost prezentă în toate populaţiile luate în studiu, fie ele pure sau de amestec.
În cadrul populaţiilor de stejar brumăriu (Figura 4.3 b), ponderea tipurilor de unităţi
infraspecifice prezente, raportate la numărul total de unităţi descrise din literatura de specialitate, a
fost cuprinsă între 33,33% şi 66,66%. Cele mai multe tipuri de unităţi infraspecifice se găsesc în
populaţiile Braniştea Catârilor, Letea şi Griviţa, iar cele mai puţine în Băneasa şi Snagov. Varietatea
atrichoclados subformele obtusiloba şi simplex au fost prezente în toate populaţiile. Varietatea
virescens, forma normalis a fost identificată doar în pădurea de amestec Griviţa. Dacă ne raportăm
doar la varietatea atrichoclados, cel mai des întâlnită, populaţiile Braniştea Catârilor şi Letea
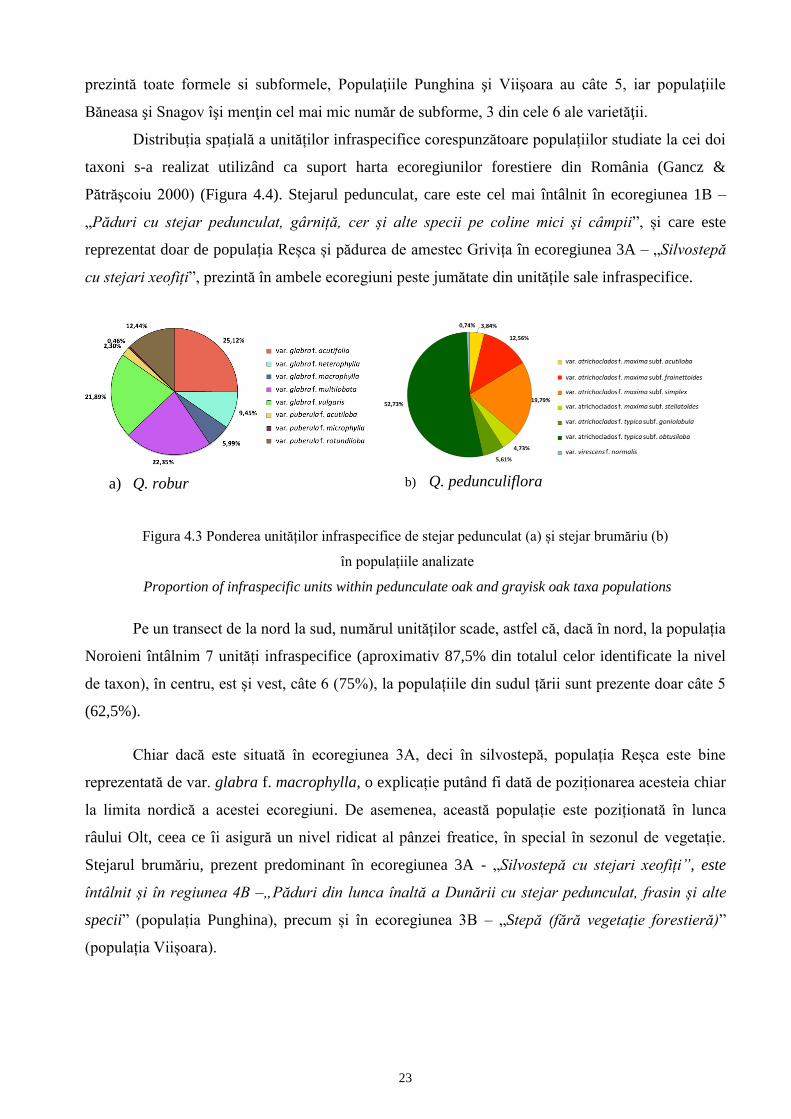
23
prezintă toate formele si subformele, Populaţiile Punghina şi Viişoara au câte 5, iar populaţiile
Băneasa şi Snagov îşi menţin cel mai mic număr de subforme, 3 din cele 6 ale varietăţii.
Distribuția spațială a unităților infraspecifice corespunzătoare populațiilor studiate la cei doi
taxoni s-a realizat utilizând ca suport harta ecoregiunilor forestiere din România (Gancz &
Pătrăşcoiu 2000) (Figura 4.4). Stejarul pedunculat, care este cel mai întâlnit în ecoregiunea 1B –
„Păduri cu stejar pedunculat, gârniță, cer și alte specii pe coline mici și câmpii”, și care este
reprezentat doar de populația Reșca și pădurea de amestec Grivița în ecoregiunea 3A – „Silvostepă
cu stejari xeofiți”, prezintă în ambele ecoregiuni peste jumătate din unitățile sale infraspecifice.
a) Q. robur
b) Q. pedunculiflora
Figura 4.3 Ponderea unităților infraspecifice de stejar pedunculat (a) și stejar brumăriu (b)
în populațiile analizate
Proportion of infraspecific units within pedunculate oak and grayisk oak taxa populations
Pe un transect de la nord la sud, numărul unităților scade, astfel că, dacă în nord, la populația
Noroieni întâlnim 7 unități infraspecifice (aproximativ 87,5% din totalul celor identificate la nivel
de taxon), în centru, est și vest, câte 6 (75%), la populațiile din sudul țării sunt prezente doar câte 5
(62,5%).
Chiar dacă este situată în ecoregiunea 3A, deci în silvostepă, populația Reșca este bine
reprezentată de var. glabra f. macrophylla, o explicație putând fi dată de poziționarea acesteia chiar
la limita nordică a acestei ecoregiuni. De asemenea, această populație este poziționată în lunca
râului Olt, ceea ce îi asigură un nivel ridicat al pânzei freatice, în special în sezonul de vegetație.
Stejarul brumăriu, prezent predominant în ecoregiunea 3A - „Silvostepă cu stejari xeofiți”, este
întâlnit și în regiunea 4B –„Păduri din lunca înaltă a Dunării cu stejar pedunculat, frasin și alte
specii” (populația Punghina), precum și în ecoregiunea 3B – „Stepă (fără vegetație forestieră)”
(populația Viișoara).

24
4.1.4 Structura taxonomică intraspecifică pentru speciile coabitante stejarului brumăriu și
stejarului pedunculat în pădurea de amestec Grivița
Pe lângă stejar brumăriu şi stejar pedunculat, în suprafaţa de cercetare intensivă au mai fost
identificate 51 de exemplare aparținând altor specii de cvercinee (Tabelele 4.6 și 47), ceea ce
reprezintă 17% din totalul numărului de arbori eșantionați. Aceste specii sunt: gorun, gorun
balcanic, gorun transilvănean, stejar pufos, stejar italian, precum şi o serie de 28 de arbori (9,3%)
care, din punct de vedere morfologic, nu corespund sensu stricto niciunei specii. Acestea din urmă
au fost denumite exemplare atipice şi au fost notate cu H (posibil hibrizi introgresivi). Având în
vedere faptul că analizele fenologice s-au realizat tocmai în vederea elucidării posibilității
interfecundării potențiale dintre specii de cvercinee, chiar dacă aceste specii nu constituie o
prioritate a obiectivului fenotipic, acestea au fost analizate, fiind astfel posibil ca anumite
caracteristici particulare, în special ale frunzelor, să poată fi explicate ca urmare a hibridării dintre
acestea.
Figura 4.4 Distribuția spațială a unităților infraspecifice de stejar pedunculat și stejar brumăriu în România
(Harta Ecoregiunilor Forestiere din România - Gancz & Pătrăşcoiu 2000)
Q. robur and Q. pedunculiflora infraspecific units spatial distribution across Romania (Romania Forest
Ecoregion map - Gancz & Pătrăşcoiu 2000)

25
Tabelul 4.5 Distribuția arborilor analizați pe varietăţi identificate la gorun, gorun balcanic şi gorun
transilvănean în pădurea de amestec Griviţa
Tree distribution based on varieties of sessile oak, balcan oak and transilvanian oak identified in Griviţa
mixed forest
Quercus petraea ssp. petraea Q. petraea ssp. dalechampii Q. petraea ssp.
polycarpa
Total
GO
var. typica
f. platyhilla f. longifolia f. lancifolia f. pinnatifida
subf. normalis subf. angustifolia
3 2 1 2 2 10
În pădurea de amestec Griviţa au fost identificate şi analizate un număr de 10 exemplare de
gorun (Tabelul 4.5). În cadrul aceleiaşi păduri au fost identificate biogrupe cu stejar pufos şi stejar
italian în totalitate fiind reprezentate cele două specii de 8 şi respectiv 5 exemplare (Tabelul 4.6).
Tabelul 4.6 Distribuția arborilor analizați pe varietăţi identificate la stejar pufos şi stejar italian în pădurea de
amestec Griviţa
Tree distribution based on varieties of pubescent oak and italian oak identified in Griviţa mix forest
Quercus pubescens Willd. Total
STP Quercus virgiliana Ten.
Total
STV
var.
lanuginosa var. undulata
var.
glomerata
var. Tenorei var. ambigua
f. pinnatifida f. dissecata f. sublobata
f. typica f. pungens
1 4 3 8 4 1 5
Un procent de aproximativ 9% din totalul numărului de cvercinee studiate, funcţie de
caracteristicile morfologice, nu au putut fi încadrate într-o specie sau alta, ele fiind considerate
exemplare atipice (Tabelul 4.7). Asemenea exemplare intermediare au fost identificate şi într-un
studiu din Franţa (Bacilieri et al. 1995), procentul acestora fiind de 3% dintr-un total de 426 de
arbori inventariaţi, sau din România (Curtu et al. 2007), unde procentul exemplarelor intermediare a
fost de 6%. De obicei, aceşti arbori intermediari se întâlesc, ca şi în cazul de faţă, la speciile
simpatrice.
În fitocenoza Grivița, Q. pedunculiflora K. Koch, Q. pubescens Willd și Q. virgiliana Ten,
reprezintă împreună 86% din totalul exemplarelor luate în studiu în această pădure de amestec.
Stejarul brumăriu nu prezintă infrataxoni cu frunze mici (f. parvifolia) sau mari (f. macrophylla),
însă prezintă arbori var. virescens f. normalis, toate aceste caracteristici morfologice fiind direct
influențate de condițiile climatice specifice silvostepei externe (Apostol - Chesnoiu et al. 2015).

26
Tabelul 4.7 Distribuția arborilor analizați pe varietăţi de frunze identificate la stejarii atipici morfologic în
pădurea de amestec Griviţa
Tree distribution based on leaf varieties ambigous oak leaf varieties identified in Griviţa mix forest
Exemplare atipice - posibil hibrizi introgresivi (H) Total
H
var. atrichoclados var. virescens
f. typica
f. maxima
f.
normalis
subf.
obtusiloba
subf.
goniolobula
subf.
acutiloba
subf.
simplex
subf.
frainettoides
subf.
lobulosa
13 1 2 5 2 1 4 28
4.2 Variabilitatea fenotipică pe baza descriptorilor morfologici ai frunzelor
Pentru evidențierea variabilității intrapopulaționale și interpopulaționale a descriptorilor
morfologici ai frunzelor stejarului pedunculat și stejarului brumăriu, s-a procedat la determinarea
principalilor indicatori statistici.
4.2.1 Variabilitatea intrapopulațională și interpopulațională a descriptorilor morfologici ai
frunzelor stejarului pedunculat
Variabile observabile şi numărabile
Pentru stejarul pedunculat, din punct de vedere al gradului de pubescenţă (descriptorul PU),
valoarea medie maximă întâlnită a fost 1,6, corespunzătoare primei trepte a scării de evaluare a
gradului de pubescenţă, ceea ce semnifică o incidență foarte redusă a frunzelor cu nivel redus de
pubescență. Din punct de vedere al culorii frunzelor, la stejarul pedunculat și cel brumăriu aceasta
variază de la verde-închis la verde-albăstrui, cea din urmă fiind specifică stejarului brumăriu. În
general, la nivelul infrataxonilor, forma bazei limbului este auriculată (BS cu valori medii la nivelul
infrataxonilor între 7,9 şi 8,9). În ceea ce priveşte variabilele numărabile pe unităţi infraspecifice, în
medie, numărul perechilor de lobi a fost de 4-5 pentru fiecare frunză. La nivelul speciei, numărul
mediu al nervurilor intercalare (NV) este de 4,4 (între 3,3 şi 5,6).
Variabile măsurabile
La nivelul unităţilor infraspecifice ale stejarului pedunculat, dimensiunile cele mai mari ale
frunzelor se regăsesc în populaţiile Păunoaia, Podul Iloaiei şi Reşca şi sunt cele corespunzătoare
varietăţii glabra, formei macrophylla, cu valori medii între 14,7- 15,1 cm). În literatură,
dimensiunile frunzei pentru f. macrophylla sunt descrise ca fiind peste 14 cm şi ajungând chiar până
la 20 (22) cm. La nivelul unităţilor infraspecifice nicio frunză de stejar pedunculat nu a depăşit
lungimea de 17 cm, iar valorile maxime pentru lungimea laminei s-au înregistrat tot în cele trei
populaţii care s-au evidenţiat prin valorile medii cele mai mari. Cele mai mici dimenesiuni s-au
înregistrat în populaţia Noroieni şi sunt cele corespunzătoare formei microphylla (7 cm), de altfel
populaţia Noroieni este singura dintre cele analizate în cadrul căreia a fost identificată această

27
formă. De aceea, ar fi utilă o cercetare ecofiziologică prin care să se clarifice dacă această formă
regăsită în populația Noroieni poate fi o adaptare la condiții ecologice locale particulare.
Variabile calculate
În ceea ce priveşte forma laminei sau obovalitatea OB%, acest descriptor are în general valori
peste 55, ceea ce denotă un caracter al frunzelor de stejar pedunculat predominant de obovalitate.
Pentru var. glabra f. heterophylla valoarea OB% este între 52 şi 61, astfel ca frunzele arborilor
încadraţi la această unitate infraspecifică sunt de la eliptice la ovate şi chiar obovate; f. multilobata
şi f. acutifolia au valori ale descriptorului peste 57%, ceea ce presupune că frunzele sunt obovate; f.
vulgaris şi macrophylla au OB% cu valori peste 54-55, deci frunze eliptice, ovate şi obovate.
4.2.2 Variabilitatea intrapopulațională și interpopulațională a descriptorilor
morfologici ai frunzelor stejarului brumăriu
Variabile observabile şi numărabile
Din punct de vedere al gradului de pubescenţă (descriptorul PU), pentru stejarul brumăriu
valorile au variat între 4 şi 6. Printre populaţiile cu gradul de pubescenţă cel mai ridicat (PU=6) se
numără: Braniştea Catârilor, pentru var. atrichoclados, f. typica, subf. gniolobula, Punghina, pentru
var. atrichoclados, f. maxima subf. acutiloba şi Urziceni - f. maxima, subf. acutiloba. Analizând
acest descriptor pentru ansamblul populaţiilor de stejar brumăriu, cu includerea tuturor unităților
infraspecifice identificate, PU variază între 4,2 (Letea) şi 5 (Punghina). Cea mai mare amplitudine
de variaţie din punct de vedere al părozităţii de pe spatele frunzei se înregistrează în populaţia Letea
(CV% = 22,7), iar cea mai mica în populaţia Punghina, aici frunzele prezentând un grad ridicat de
pubescenţă.
În ceea ce priveşte evaluarea culorii de pe dosul frunzelor (descriptorul CL) aceasta variază
de la verde-deschis la brumăriu, valoarea cea mai ridicată (treapta a 3-a pe scara de evaluare) o
întâlnim la subformele obtusiloba (populaţia Punghina) și stellatoides (populaţia Letea
pedunculiflora). De remarcat faptul că, per ansamblul unităților infraspecifice existente, cele mai
brumării sunt cele din populaţia Punghina (CL=2,9), unde, ca şi în populaţia Viişoara, nu au fost
identificate frunze de culoare verde. Acest aspect se corelează și cu gradul ridicat de părozitate.
Valorile medii ale parametrului BS (forma bazei limbului) la nivelul infrataxonilor sugerează o
formă a acesteia de la cordat auriculată la slab, sau chiar puternic auriculată (BS=6,8-8). La nivel
interpopulațional, forma bazei limbului este cordat-auriculată (BS cu valori medii la nivel
populațional între 7,3 şi 8,3), formă ce se regăseşte în descrierea morfologică a bazei frunzei în
literatura de specialitate (Dumitriu-Tătăranu 1960, Şofletea şi Curtu 2007).

28
Variabile măsurabile
Stejarul brumăriu a prezentat dimensiuni medii mai mari ale lungimii laminei faţă de stejarul
pedunculat (Georgescu & Cretzoiu 1941). Realizând o analiză a valorilor descriptorului lungimea
laminei se poate spune că exemplarele populaţiilor eşantionate aparţinând var. atrichoclados, f.
typica, subf. obtusiloba prezintă valori medii ale LL între 9,5-12,5 cm, subf. gonilobula între 9,5 şi
14 cm. La var. atrichoclados, f. maxima, subf. stellatoides LL prezintă dimensiuni între 10,5-13,5
cm, subf. acutiloba 9,5-14 cm, subf. simplex 11-15.5 cm şi subf. frainettoides 10-14 cm.
Variabile calculate
Descriptorul forma laminei sau obovalitatea OB% a avut în general valori între 52% şi 66%,
ceea ce confirmă formele descrise în literatura, şi anume că frunzele sunt eliptice, obovate, chiar lat
obovate, deci cu lăţimea maximă la mijlocul frunzei sau mai sus (Beldie, 1952; Dumitriu -Tătăranu,
1960; Şofletea & Curtu, 2007). Pentru var. glabra f. heterophylla valoarea OB% este între 52% şi
61%, astfel că frunzele arborilor încadraţi la această unitate infraspecifică sunt de la eliptice la ovate
şi chiar obovate; f. multilobata şi f. acutifolia au valori ale descriptorului peste 57%, ceea ce
presupune că frunzele sunt obovate; f. vulgaris şi macrophylla au OB% cu valori peste 54-55, deci
frunze eliptice, ovate şi obovate.
4.2.3 Evaluări comparative stejar pedunculat - stejar brumăriu prin analize statistice ale
descriptorilor morfologici ai frunzelor
În scopul identificării descriptorilor morfologici foliari care ar putea contribui la
diferențierea celor doi taxoni luați în studiu, Q. robur și Q. pedunculiflora, s-au utilizat o serie de
calcule și analize statistice precum: valori medii, minime și maxime, ANOVA, Analiza
componentelor principale (PCA), Analiza discriminantă (DA), Analiza Cluster. Mai apoi, pe baza
rezultatelor obținute, s-a realizat o sinteză a acestora în scopul evidențierii deosebirilor dintre cei
doi taxoni.
4.2.3.1 Valori medii fenotipice și analiza varianței (ANOVA) a descriptorilor morfologici ai
frunzelor
Analiza simplă a varianţei s-a aplicat cu scopul de a valida variabile ce pot contribui la
exprimarea unor diferenţe semnificative (p<0,05) între stejarul pedunculat şi stejarul brumăriu. În
studii anterioare din ţara noastră s-a consemnat faptul că diferenţele morfologice remarcabile la
nivelul descriptorilor frunzei între cei doi taxoni se întâlnesc în pădurile din sudul ţării însă, pe un
transect de la sud la nord-est aceste diferenţe se atenuează (Dumitriu - Tătăranu 1954). Atât la
stejarul pedunculat, cât și la stejarul brumăriu, s-au obținut diferenţe semnificative (p<0,05) între
populaţii la nivelul tuturor celor 15 variabile ale descriptorilor. În studiul lui Bacilieri et al. (1995)
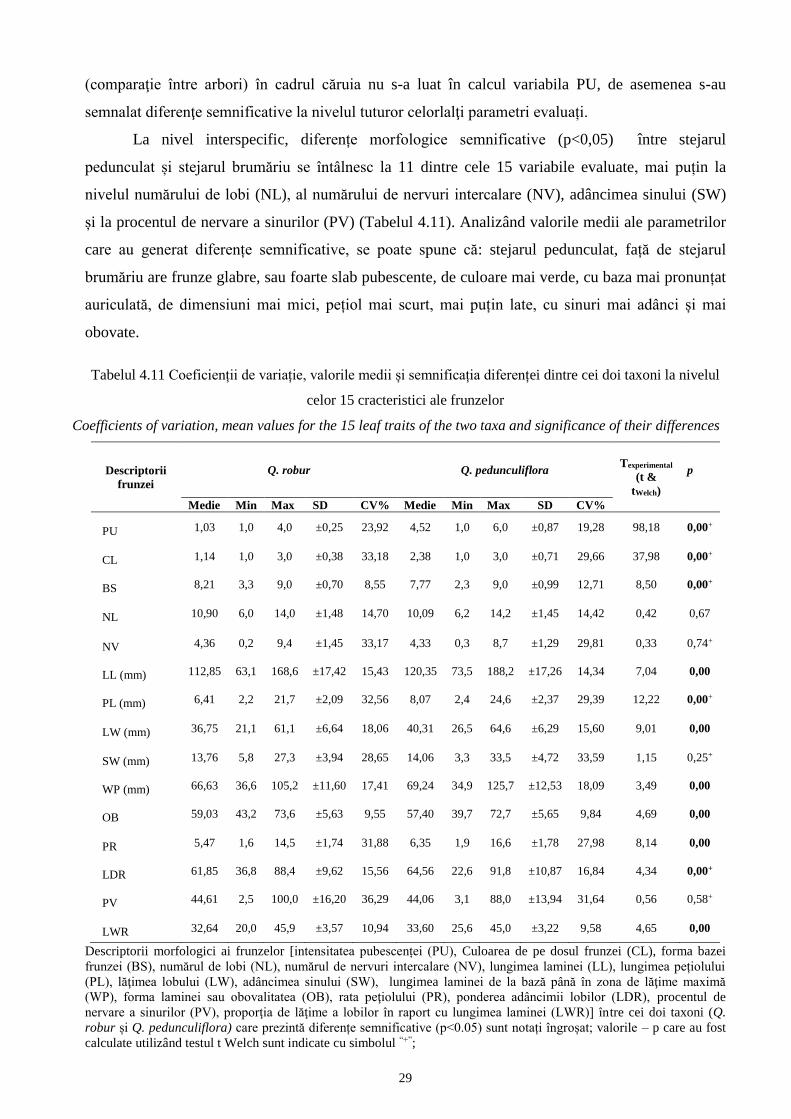
29
(comparaţie între arbori) în cadrul căruia nu s-a luat în calcul variabila PU, de asemenea s-au
semnalat diferenţe semnificative la nivelul tuturor celorlalţi parametri evaluați.
La nivel interspecific, diferențe morfologice semnificative (p<0,05) între stejarul
pedunculat și stejarul brumăriu se întâlnesc la 11 dintre cele 15 variabile evaluate, mai puțin la
nivelul numărului de lobi (NL), al numărului de nervuri intercalare (NV), adâncimea sinului (SW)
și la procentul de nervare a sinurilor (PV) (Tabelul 4.11). Analizând valorile medii ale parametrilor
care au generat diferențe semnificative, se poate spune că: stejarul pedunculat, față de stejarul
brumăriu are frunze glabre, sau foarte slab pubescente, de culoare mai verde, cu baza mai pronunțat
auriculată, de dimensiuni mai mici, pețiol mai scurt, mai puțin late, cu sinuri mai adânci și mai
obovate.
Tabelul 4.11 Coeficienții de variație, valorile medii și semnificația diferenței dintre cei doi taxoni la nivelul
celor 15 cracteristici ale frunzelor
Coefficients of variation, mean values for the 15 leaf traits of the two taxa and significance of their differences
Descriptorii
frunzei
Q. robur Q. pedunculiflora
Texperimental
(t &
tWelch)
p
Medie Min Max SD CV% Medie Min Max SD CV%
PU 1,03 1,0 4,0 ±0,25 23,92 4,52 1,0 6,0 ±0,87 19,28 98,18 0,00+
CL 1,14 1,0 3,0 ±0,38 33,18 2,38 1,0 3,0 ±0,71 29,66 37,98 0,00+
BS 8,21 3,3 9,0 ±0,70 8,55 7,77 2,3 9,0 ±0,99 12,71 8,50 0,00+
NL 10,90 6,0 14,0 ±1,48 14,70 10,09 6,2 14,2 ±1,45 14,42 0,42 0,67
NV 4,36 0,2 9,4 ±1,45 33,17 4,33 0,3 8,7 ±1,29 29,81 0,33 0,74+
LL (mm) 112,85 63,1 168,6 ±17,42 15,43 120,35 73,5 188,2 ±17,26 14,34 7,04 0,00
PL (mm) 6,41 2,2 21,7 ±2,09 32,56 8,07 2,4 24,6 ±2,37 29,39 12,22 0,00+
LW (mm) 36,75 21,1 61,1 ±6,64 18,06 40,31 26,5 64,6 ±6,29 15,60 9,01 0,00
SW (mm) 13,76 5,8 27,3 ±3,94 28,65 14,06 3,3 33,5 ±4,72 33,59 1,15 0,25+
WP (mm) 66,63 36,6 105,2 ±11,60 17,41 69,24 34,9 125,7 ±12,53 18,09 3,49 0,00
OB 59,03 43,2 73,6 ±5,63 9,55 57,40 39,7 72,7 ±5,65 9,84 4,69 0,00
PR 5,47 1,6 14,5 ±1,74 31,88 6,35 1,9 16,6 ±1,78 27,98 8,14 0,00
LDR 61,85 36,8 88,4 ±9,62 15,56 64,56 22,6 91,8 ±10,87 16,84 4,34 0,00+
PV 44,61 2,5 100,0 ±16,20 36,29 44,06 3,1 88,0 ±13,94 31,64 0,56 0,58+
LWR 32,64 20,0 45,9 ±3,57 10,94 33,60 25,6 45,0 ±3,22 9,58 4,65 0,00
Descriptorii morfologici ai frunzelor [intensitatea pubescenței (PU), Culoarea de pe dosul frunzei (CL), forma bazei
frunzei (BS), numărul de lobi (NL), numărul de nervuri intercalare (NV), lungimea laminei (LL), lungimea pețiolului
(PL), lăţimea lobului (LW), adâncimea sinului (SW), lungimea laminei de la bază până în zona de lăţime maximă
(WP), forma laminei sau obovalitatea (OB), rata peţiolului (PR), ponderea adâncimii lobilor (LDR), procentul de
nervare a sinurilor (PV), proporţia de lăţime a lobilor în raport cu lungimea laminei (LWR)] între cei doi taxoni (Q.
robur și Q. pedunculiflora) care prezintă diferențe semnificative (p<0.05) sunt notați îngroșat; valorile – p care au fost
calculate utilizând testul t Welch sunt indicate cu simbolul “+”;

30
Evaluări comparative cu ajutorul variaţiei cumulate a descriptorilor (VCD) - analiza
variabilităţii intra- şi interpopulaţionale şi interspecifice
Luând în considerare descriptorii generaţi de funcţia discriminantă pentru cei doi taxoni, se
observă că, cea mai mare variabilitate la stejarul pedunculat, la nivelul acestor descriptori, este în
populaţia Podul Iloaiei, iar cea mai mică în populaţia Vânju Mare.
La stejarul brumăriu cea mai mare variaţie cumulată a descriptorilor s-a înregistrat în
pădurea de amestec Griviţa, urmată de populațiile Băneasa, Urziceni şi Letea, care prezintă valori
apropiate. Punghina este populaţia cu cea mai mică variaţie cumulată. Culoarea este cea care
variază cel mai mult la stejarul brumăriu, urmată de rata pețiolului, pubescență, forma bazei și în
final de proporţia de lăţime a lobilor în raport cu lungimea laminei.
Analiza corelaţiei
În ceea ce privește corelațiile dintre variabilele măsurate, observate și numărate la stejarul
pedunculat, se poate remarca o corelație foarte strânsă, directă și foarte puternic semnificativă între
LL și LW (r=0,79), LL și WP (r=0,83) și o corelație strânsă, directă și foarte puternic semnificativă
între LW și WP (r=0,51). Aceste corelații confirmă faptul că frunzele stejarului pedunculat, fie că
sunt mici sau mari, acestea respectă proporțiile dimensionale care le conferă forma acestora, în
general obovată. Corelația strânsă dintre lățimea lobului și lungimea laminei de la bază până în zona
de lățime maximă întărește afirmația conform căreia forma frunzelor de stejar pedunculat este în
general constantă. La stejarul brumăriu se poate remarca aceeași corelație foarte strânsă, directă și
foarte puternic semnificativă între LL și LW (r=0,79), LL și WP (r=0,84) și o corelație strânsă,
directă și foarte puternic semnificativă între LW și WP (r=0,60).
4.2.3.2 Analiza componentelor principale (PCA)
În analiza componentelor principale s-au folosit toate seturile de date corespunzătoare
speciilor evaluate, inclusiv cele din pădurea de amestec Grivița, constituindu-se astfel o populaţie
sintetică alcătuită din 1162 de arbori din speciile: Quercus robur - 434 arbori (37,35%), Q. petraea
- 10 arbori (0,86%), Q. pubescens – 8 arbori (0,69%), Q. virgiliana – 5 arbori (0,43%), Q.
pedunculiflora – 677 arbori (58,26%%), precum și 28 de exemplare atipice din pădurea de amestec
Grivița (H) (2,41%). În cadrul populaţiei sintetice au fost introduse valorile medii ale tuturor celor
15 descriptori pentru toate populațiile și a rezultat un grafic PCA (un singur nor de puncte) pentru
toate speciile. Prin evidențierea în culori distincte ale arborilor, funcție de specia la care aceștia au
fost încadrați în urma evaluărilor morfologice, se observă o zonare incipientă (Figura 4.5) de-a
lungul Factorului 2 (18,09) (Figura 4.6). După aplicarea analizei discriminante, în urma căreia s-au
evidenţiat variabilele care pot alcătui funcţia discriminantă dintre cei doi taxoni, s-a repetat analiza
componentelor principale utilizând doar aceste variabile (PU, CL, BS, PR şi LWR), de această dată
formarea celor doi nori de puncte corespunzători celor două specii fiind foarte evidentă (Figura 4.7).
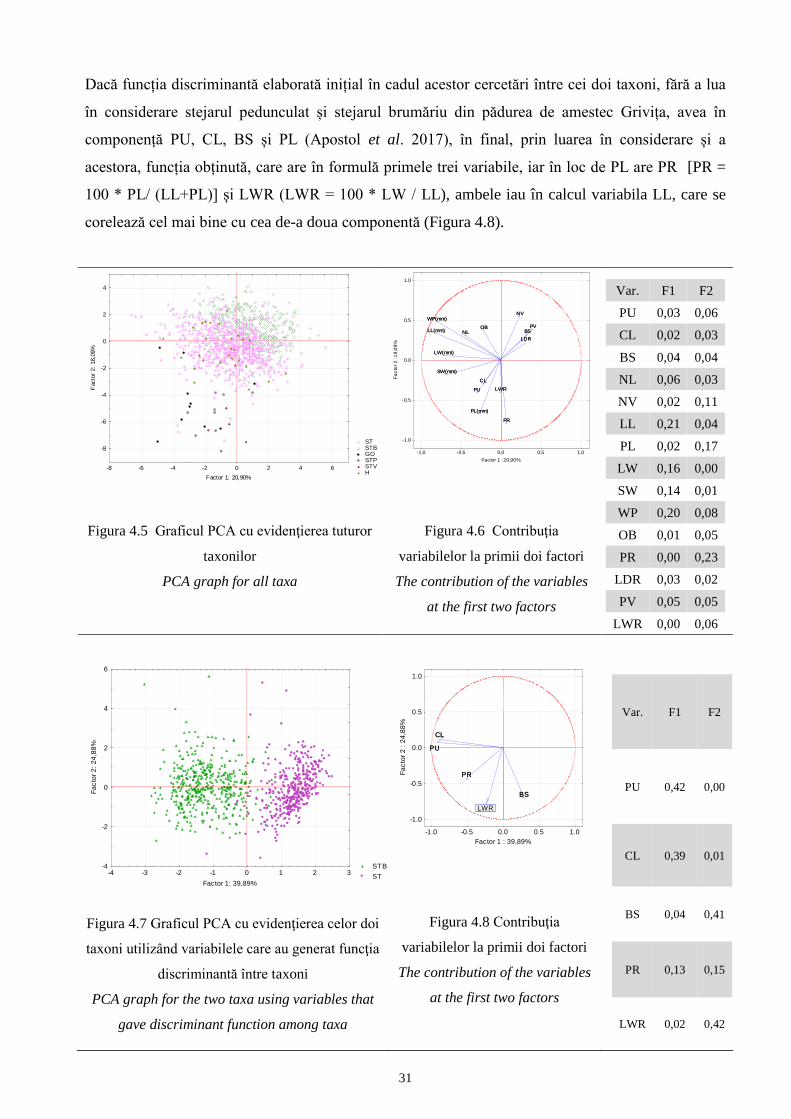
31
Dacă funcția discriminantă elaborată inițial în cadul acestor cercetări între cei doi taxoni, fără a lua
în considerare stejarul pedunculat și stejarul brumăriu din pădurea de amestec Grivița, avea în
componență PU, CL, BS și PL (Apostol et al. 2017), în final, prin luarea în considerare și a
acestora, funcția obținută, care are în formulă primele trei variabile, iar în loc de PL are PR [PR =
100 * PL/ (LL+PL)] și LWR (LWR = 100 * LW / LL), ambele iau în calcul variabila LL, care se
corelează cel mai bine cu cea de-a doua componentă (Figura 4.8).
ST STB GO STP STV H
-8 -6 -4 -2 0 2 4 6
Factor 1: 20,90%
-8
-6
-4
-2
0
2
4
Facto
r 2:
18,0
9%
PU
CL
BSNL
NV
LL(mm)
PL(mm)
LW(mm)
SW(mm)
WP(mm)
OB
PR
LDR
PV
LWR
-1.0 -0.5 0.0 0.5 1.0
Factor 1 : 20,90%
-1.0
-0.5
0.0
0.5
1.0
Fa
cto
r 2
: 1
8,0
9%
PU
CL
BSNL
NV
LL(mm)
PL(mm)
LW(mm)
SW(mm)
WP(mm)
OB
PR
LDR
PV
LWR
Var. F1 F2
PU 0,03 0,06
CL 0,02 0,03
BS 0,04 0,04
NL 0,06 0,03
NV 0,02 0,11
LL 0,21 0,04
PL 0,02 0,17
LW 0,16 0,00
SW 0,14 0,01
WP 0,20 0,08
OB 0,01 0,05
PR 0,00 0,23
LDR 0,03 0,02
PV 0,05 0,05
LWR 0,00 0,06
Figura 4.5 Graficul PCA cu evidenţierea tuturor
taxonilor
PCA graph for all taxa
Figura 4.6 Contribuţia
variabilelor la primii doi factori
The contribution of the variables
at the first two factors
STB
ST-4 -3 -2 -1 0 1 2 3
Factor 1: 39,89%
-4
-2
0
2
4
6
Fa
cto
r 2
: 2
4,8
8%
Figura 4.7 Graficul PCA cu evidenţierea celor doi
taxoni utilizând variabilele care au generat funcţia
discriminantă între taxoni
PCA graph for the two taxa using variables that
gave discriminant function among taxa
PU
CL
BS
PR
LWR
-1.0 -0.5 0.0 0.5 1.0
Factor 1 : 39,89%
-1.0
-0.5
0.0
0.5
1.0
Fa
cto
r 2
: 2
4,8
8%
PU
CL
BS
PR
LWR
Figura 4.8 Contribuţia
variabilelor la primii doi factori
The contribution of the variables
at the first two factors
Var. F1 F2
PU 0,42 0,00
CL 0,39 0,01
BS 0,04 0,41
PR 0,13 0,15
LWR 0,02 0,42

32
4.2.3.3 Analiza discriminantă
Pentru a putea evalua combinaţiile multivariate dintre caractere, s-a aplicat analiza
discriminantă pentru cele 15 variabile pe datele tuturor celor 19 populaţii. În urma analizei s-a
obținut funcția discriminantă între cei doi taxoni care furnizează semnul plus (+) pentru stejarul
pedunculat şi semnul minus (-) pentru stejarul brumăriu:
557,9-161,1*PU-40,7*CL-6,5*PR+13,2*BS-2,3*LWR
Specific acestei funcții discriminante este faptul că, în cadrul acestei cercetări a fost luat în
analiză, pentru prima dată, descriptorul CL (culoarea de pe dosul frunzelor). Testarea funcţiei
discriminante s-a realizat pentru stejarul pedunculat şi stejarul brumăriu utilizând arborii celor doi
taxoni din pădurea de amestec Griviţa, validarea acesteia realizându-se 100% la stejarul pedunculat
şi 94% la stejaul brumăriu. Testarea şi validarea unor astfel de funcţii s-au realizat în cazul
diferenţierii lui Quercus prinus de Q. alba (Ledig et al. 1969), a stejarului pedunculat de gorun
(Kremer et al. 2002, Ponton et al. 2004, Kelleher et al. 2004), a gorunului de stejarul pufos
(Grossoni et al. 1998, Bruschi et al. 2003b) sau a stejarului brumăriu de stejarul pedunculat (Curtu
et al. 2009).
Variabilele care au arătat putere de discriminare sunt PU, CL, PR, BS și LWR. Variabila
PR, alături de PU și BS au avut un rol important de diferenţiere a stejarului pedunculat de gorun
într-un studiu cu privire la distribuţia stejarului pedunculat şi a gorunului în sud-estul Angliei
(Wigston, 1975). Ținând cont de faptul că PR (rata pețiolului) se calculează pe baza valorilor PL și
LL, se confirmă faptul că atât la nivelul lungimii pețiolului, cât și la nivelul dimensiunilor frunzelor,
cei doi taxoni se pot diferenția, stejarul brumăriu având în general frunze mai mari decât stejarul
pedunculat.
❖ Analiza discriminantă pe unităţi infraspecifice
Atunci când analiza discriminantă s-a aplicat la nivel intraspecific, variabilele care au
contribuit la discriminarea unităților infraspecifice sunt complet diferite de cele de la nivel
interspecific.
Quercus robur
La stejarul pedunculat, analiza discriminantă reliefează faptul că variabilele care contribuie
semnificativ (p<0,05 și Parțial Lambda cu valori cuprinse între 0,92-0,95) la diferențierea unităților
infraspecifice sunt strâns legate de elementele de descriere ale unităților infraspecifice
corespunzătoare, și anume: lungimea laminei (LL), proporția lățimii lobilor în raport cu lungimea
laminei (LWR), adâncimea sinurilor (SW), numărul de lobi (NL), ponderea adâncimii lobilor
(LDR), procentul de nervare a sinurilor (PV) și numărul de nervuri intercalare (NV). Pentru
diferențierea unitaților infraspecifice la stejarul pedunculat se recomandă utilizarea funcțiilor de mai
jos, care au fost testate și validate 100%.

33
FCT1: 968,0-128,8*LL-32,88*LWR+63,6*SW-52,8*NL+20,4*LDR-9,5*PV+86*NV, generează
semnul minus (-) pentru f. multilobata și semnul plus (+) pentru f. acutifolia; FCT 2 : -704,2-
29,7*LL+10*LWR+2,9*SW+11,6*NL+10,5*LDR-5,8*PV+37,7*NV, generează semnul minus (-)
pentru var. glabra f. heterophylla, f. acutifolia, f. macrophylla, f. vulgaris și var. puberula f.
rotundiloba; FCT3: 929,6+16,8*LL-15,6*LWR-30,4*SW+19,2*NL-6,6*LDR-0,2*PV+5,3*NV,
generează semnul minus (-) pentru f. macrophylla și semnul plus (+) pentru. f. multilobata, f.
vulgaris și f. acutiloba; FCT4: -945,4 -81,4*LL -27,0*LWR +75,8*SW+8,2*NL+23,9*LDR-
7,2*PV+107*NV, generează semnul minus (-) pentru var. glabra f. heterophylla, f . vulgaris și
pentru var. puberula f. acutiloba, și semnul plus (+) pentru var. glabra f. acutifolia; FCT5: -839,3
-1,8*LL +13*LWR -22,6*SW+118,8*NL-8,9*LDR+24,3*PV-227,6*NV, generează semnul minus
(-) pentru var. glabra f. heterophylla, f . multilobata, f. macrophylla și semnul plus (+) pentru var.
puberula f. acutiloba; FCT6: -841,9 -96,2*LL -45*LWR +89,6*SW-
22,3*NL+37,2*LDR+3,6*PV-16,4*NV, generează semnul minus (-) pentru var. glabra f.
multilobata, f. vulgaris și pentru var. puberula f. acutiloba.
Pentru încadrarea corectă a fiecărei unități infraspecifice în parte, fiecare dintre acestea
trebuie să respecte cumulativ semnul fiecărei funcții discriminate care a clasificat corect 100% din
cazuri. De exemplu, un arbore ale cărui valori ale variabilelor morfologice descriptive sunt
introduse în FCT 2, FCT 4 și FCT 5 și dau cumulativ semnul minus (-) este încadrat la var. glabra f.
hererophylla.
Quercus pedunculiflora var. atrichoclados
La stejarul brumăriu, analiza discriminantă reliefează faptul că variabilele care contribuie
semnificativ (p<0.05 și Parțial Lambda cu valori cuprinse între 0,93-0,96) la diferențierea unităților
infraspecifice sunt, ca și la stejarul pedunculat, strâns legate de elementele de descriere ale
unităților infraspecifice corespunzătoare, și anume: lățimea lobilor (LW), adâncimea sinurilor (SW),
lungimea laminei (LL), numărul de lobi (NL), rata pețiolului (PR) și lungimea pețiolului (PL).
FCT1 : 1595-15,2*LW+125,9*SW-64,9*LL-30,6*NL-119,9*PR+85,5*PL generează semnul minus
(-) pentru f. typica subf. goniolobula și pentru f. maxima subf. stellatoides, subf. simplex și subf.
Frainettoides; FCT2 : 716,6+6,9*LW-215,8*SW-77,8*LL+21,8*NL-57,2*PR+44,7*PL generează
semnul minus (-) pentru f. typica subf. obtusiloba și pentru f. maxima subf. acutiloba și subf.
simplex. FCT3 : 175,9-14,8*LW-119,1*SW+96,3*LL-37,6*NL+25,6*PR-45,1*PL generează
semnul plus (+) pentru f. typica subf. goniolobula și pentru f. maxima subf. stellatoides și subf.
Simplex; FCT4 : -1352,3-14,2*LW-50,1*SW+121,3*LL+17*NL+169,9*PR-91,3*PL generează
semnul plus (+) pentru f. typica subf. goniolobula și pentru f. maxima subf. Acutiloba; FCT5 :
457,8-10,2*LW+40,6*SW-38,2*LL+52,4*NL-111,8*PR+66,8*PL generează semnul minus (-)
pentru f. maxima subf. stellatoides și subf. acutiloba.

34
4.2.3.4 Analiza cluster
Dintre metodele „aglomerative”, cel mai des întâlnită în studiile de morfologie și taxonomie
este metoda înlănțuirii complete (engl. Complete Linkage) (Bruschi et al. 2003a). În cadrul
prezentului studiu, această analiză a fost realizată pentru toate populaţiile, inclusiv grupul de stejari
pedunculați (GRR) și stejari brumării (GRP) din pădurea de amestec Grivița utilizând ca variabile
cei 15 descriptori ai frunzei (Figurile 4.9. și 4.10).
0 1 2 3 4 5 6 7
Linkage Distance
PAU
TIM
RES
VJM
LEP
GRP
PUN
BRA
POG
SNA
VIS
MIT
URZ
BAN
GRR
NOR
POD
RUP
LER
Figura 4.9 Analiza cluster pentru toate populaţiile de ST și STB utilizând cei cinci descriptori (PU, CL,
PR%, BS și LWR%) generați de funcţia discriminantă
Cluster analysis for all populations of ST and STB using the five descriptors (PU, CL, PR%, BS și
LWR%) generated by the discriminant function
Se observă crearea a două clustere principale, clusterul 1 (de sus în jos), care grupează
majoritar populații de stejar pedunculat și clusterul 2, care grupează aporape în aceeași proporție
populații de stejar brumăriu, dintre cele 8 în total, sunt prezente 3 populații de stejar pedunculat.
Prin utilizarea tuturor variabilelor în analiză, nu s-a obținut o grupare clară a celor taxoni analizați,
și nici pe regiuni geografice. Analiza Cluster s-a realizat și cu utilizarea doar a variabilelor care au
generat diferențe semnificative, observându-se o grupare tot în două clustere principale, cu o
grupare pe taxoni doar la nivelul subclusterelor inferioare din ramură. Introducând din nou în
analiza cluster toate populaţiile, cu utilizarea doar a variabilelor generate de funcţia discriminantă
(PU, CL, PR, BS și LWR) s-au format două clustere, clusterul 1 doar cu populațiile pure de stejar
pedunculat din centrul, nord-vestul, estul și sud-estul țării, clusterul 2, cu 2 subclustere, unul format
din toate grupurile de stejar brumăriu și cel de-al doilea subcluster alcătuit din populațiile de stejar
pedunculat din sudul, vestul și sud-vestul țării.

35
Figura 4.10 Distribuția spațială a celor trei grupuri de Q. robur și Q. pedunculiflora în România, rezultate în
urma Analizei Cluster [harta de bază –Harta Ecoregiunilor Forestiere din România (Gancz & Pătrăşcoiu
2000)
Q. robur and Q. pedunculiflora three groups spatial distribution across Romania performed by Cluster
Analysis [base map – Romania Forest Ecoregion map (Gancz & Pătrăşcoiu 2000)]
4.2.3.5 Sinteza evaluării morfologice comparative stejar pedunculat - stejar brumăriu după
descriptorii frunzelor
La nivel intraspecific, dacă la stejarul pedunculat au fost identificate 8 dintre cele 9 unități
infraspecifice consemnate în literatura de specialitate, la stejarul brumăriu au fost identificate 7
dintre cele 10. Nu a fost identificat niciun stejar pedunculat var. glabra f. parvifolia și niciun
exemplar de stejar brumăriu care să fie asimilat cu var. atrichoclados f. parvifolia, var. virescens f.
macrophylla și subf. lobulosa.
Cel mai prezent în ecoregiunea 1.B - Păduri cu Q. robur, Q. cerris, Q. frainetto și alte
specii, pe coline mici și câmpii, stejarul pedunculat prezintă 7 dintre cele 8 unități infraspecifice, iar
pe un transect de la nord la sud, numărul acestora scade, ajungând în sud până la 5 (RES, VJM și
PAU). Stejarul brumăriu este întâlnit mai ales în ecoregiunea 3.A – Silvostepă cu stejari xerofiți,
doar populațiile Punghina și Viișoara se regăsesc în alte ecoregiuni precum 4.B - Păduri din lunca
înaltă a Dunării cu stejar pedunculat, frasin și alte specii (populația Punghina), respectiv în
ecoregoiunea 3B – Stepă (fără vegetație forestieră) (populația Viișoara). Față de celelalte populații,

36
stejarul brumăriu din populațiile din sudul și sud-vestul țării tării (PUN și BRA) prezintă o
frecvență mai mică a f. typica subf. obtusiloba, iar în populațiile SNA și BAN sunt întâlnite doar
câte 3 unități infraspecifice (42,8% din totalul tipurilor de unități infraspecifice identificate la
nivelul taxonului).
La nivel interspecific, diferențe semnificative la nivelul celor doi taxoni (p<0,5) s-au
înregistrat la nivelul a 11 parametri morfologici foliari dintre cei 15 analizați, excepție făcând
numărul de lobi (NL), numărul de nervuri intercalare (NV), adâncimea sinului (SW) și procentul de
nervare a sinurilor (PV). Totuși, trebuie menționat faptul că, în ceea ce privește valoarea
descriptorului SW, limita inferioară a intervalului de valori la stejarul brumăriu este mai mică față
de cea a stejarului pedunculat. Altfel spus, daca la stejarul brumăriu pot fi sinuri care să ajungă
chiar până aproape de nervura principală a frunzei, la stejarul pedunculat acestea nu ajung până la
mai mult de 6 mm față de aceasta.
Cea mai mare diferențiere între cei doi taxoni se constată la nivelul descriptorilor
morfologici foliari, precum pubescența (PU) și culoarea de pe dosul frunzelor (CL). Dacă stejarul
pedunculat este glabru, foarte rar prezintă peri simpli, valoarea medie a PU este de 1,03, stejarul
brumăriu prezintă peri predominant fasciculați, cu un grad de pubescență ridicat (PU= 4,52). Pe de
altă parte, culoarea brumărie a frunzei constituie o strategie adaptivă în vederea reducerii
transpirației la plante (Apostol et al. 2017). Diferența de culoare de pe dosul frunzelor, dar și
adaptarea diferită la secetă a celor două specii ar mai putea rezulta dintr-o posibilă diferență între
cele două specii referitoare la dizolvarea (la stejarul pedunculat) și persistența (la stejarul brumăriu)
a cerii cristaloide (Gülz & Müller 1992). Cei doi descriptori morfologici foliari (PU și CL)
caracterizează și diferențiază clar stejarul brumăriu de stejarul pedunculat.
În ceea ce privește pubescența, și mai ales tipul de părozitate, exemplarele desemnate
atipice, posibil hibrizi introgresivi, prezintă peri fasciculați și stelați în același timp, cei din urmă
fiind specifici gorunului. În studiul de față, valoarea PU de 1,03 pentru Q. robur a fost similară cu
cea raportată într-un studiu similar din Franța; frunze glabre (Dupouey et al. 1990) sau cu peri
simpli unicelulari, identificate în alte studii din Anglia (Wigston 1975), din Franța (Bacilieri et al.
1996), unde a fost înregistrată o valoare de 1,2 sau de 0,2 (Ponton et al. 2004).
Forma bazei frunzelor (BS) variază de la auriculat la puternic auriculată, și chiar răsfrânt
auriculat la Q. robur (Stanescu et al. 1997), și de la cordat auriculată la slab auriculat sau auriculat,
și chiar răsfrânt auriculat la Q. pedunculiflora și corespunde formei specifice speciilor descrise în
literatură (Dumitriu - Tătăranu 1960, Șofletea & Curtu 2007).
Valoarea LL pentru Q. robur de peste 11 cm se încadrează în valorile menționate în
literatură (Bacilieri et al. 1996, Bašić et al. 2007, Boratynski et al. 2008, Ponton et al. 2004). Chiar
dacă multe specii, în condiții xerofitice prezintă frunze micșorate , tocmai ca o strategie de
supraviețuire prin reducerea suprafeței de transpirație, stejarul brumăriu este atipic, prezentând atât

37
în silvostepă, cât și în stepă (populația Viișoara) frunze cu dimensiuni mari, cu valori cuprinse în
intervalul 7 și 19 cm, și cu o valoare medie la nivelul taxonului de 12 cm. Numărul mediu al
perechilor de lobi la Q. robur și Q. pedunculiflora a fost similar, cu valori medii de 5 perechi de
lobi (interval de valori între 3 și 7) și corespunde cu cel consemnat în literatură (Negulescu &
Săvulescu 1957, Dumitriu - Tătăranu 1960, Stănescu et al. 1997, Șofletea & Curtu 2007). Numărul
de nervuri intercalare (NV) prezintă valori medii similare la cei doi taxoni studiați, de 4,3 Acești doi
descriptori NL și NV nu contribuie la discriminarea celor doi taxoni
Lungimea pețiolului (PL) este un alt descriptor important ce diferențiază cei doi taxoni, la
stejarul pedunculat fiind mai scurt, comparativ cu stejarul brumăriu. La nivelul acestui descriptor,
amplitudine de variație (CV% = 32.56%), pentru Q. robur este în concordanță cu rezultatele unui
studiu realizat în Bulgaria (Broshtilov 2006), în care CV% a variat 37,3% și 44,7%. Lungimea
medie a pețiolului (PL) pentru Q. pedunculiflora, de sub 1 cm, se încadrează în partea de sus a
variației înregistrate în literatură (Negulescu & Savulescu 1957, Șofletea & Curtu 2007).
Pentru Q. robur, lățimea medie a lobului la punctul de lățime maximă a laminei (LW) a
variat între 2,11 cm și 6,11 cm, indicând valori ușor mai mici decât cele înregistrate în studiile din
Polonia (Boratynski et al. 2008) și Bosnia (Bašić et al. 2007). Valoarea medie a LW la stejarul
brumăriu este cu 3,6 cm mai mare față de cea a stejarului pedunculat.
4.3 Analiza fenologiei
Cercetările fenologice au fost efectuate în suprafața intensivă de cercetare Grivița, în
arboretul mixt de cvercinee în care s-au urmărit fenofazele înfrunzirii și înfloririi la 300 de arbori,
precum și în două plantaje (livezi semincere): unul de proveniențe de stejar pedunculat, situat la
Mândrești (Ocolul Silvic Tecuci), respectiv altul de stejar pedunculat în amestec cu stejar brumăriu,
la Cobadin (Ocolul Silvic Murfatlar).
4.3.1 Analiza fenologiei de primăvară privind înfrunzirea și înflorirea
4.3.1.1 Analiza fenologiei de primăvară privind înfrunzirea în pădurea de amestec Grivița
În ceea ce privește analiza fenologiei la speciile coabitante în pădurea de amestec Grivița, s-
a urmărit suprapunerea sau nu a fenofazelor, atât pentru înfrunzire, cât mai ales pentru înflorire, în
scopul clarificării existenței barierei temporale fenologice între cele patru specii, dar mai ales între
stejarul pedunculat și stejarul brumăriu, care să reprezinte o premisă, pentru a formula ipoteza că
taxonii nu sunt izolați reproductiv, ceea ce ar oferi din această perspectivă informații despre
posibilitatea producerii hibridărilor interspecifice.
Astfel, s-au efectuat observații fenologice în patru ani (2008, 2009, 2015 și 2016) asupra a
300 de arbori, aceiași care au fost evaluați și din punct de vedere al descriptorilor morfologici ai
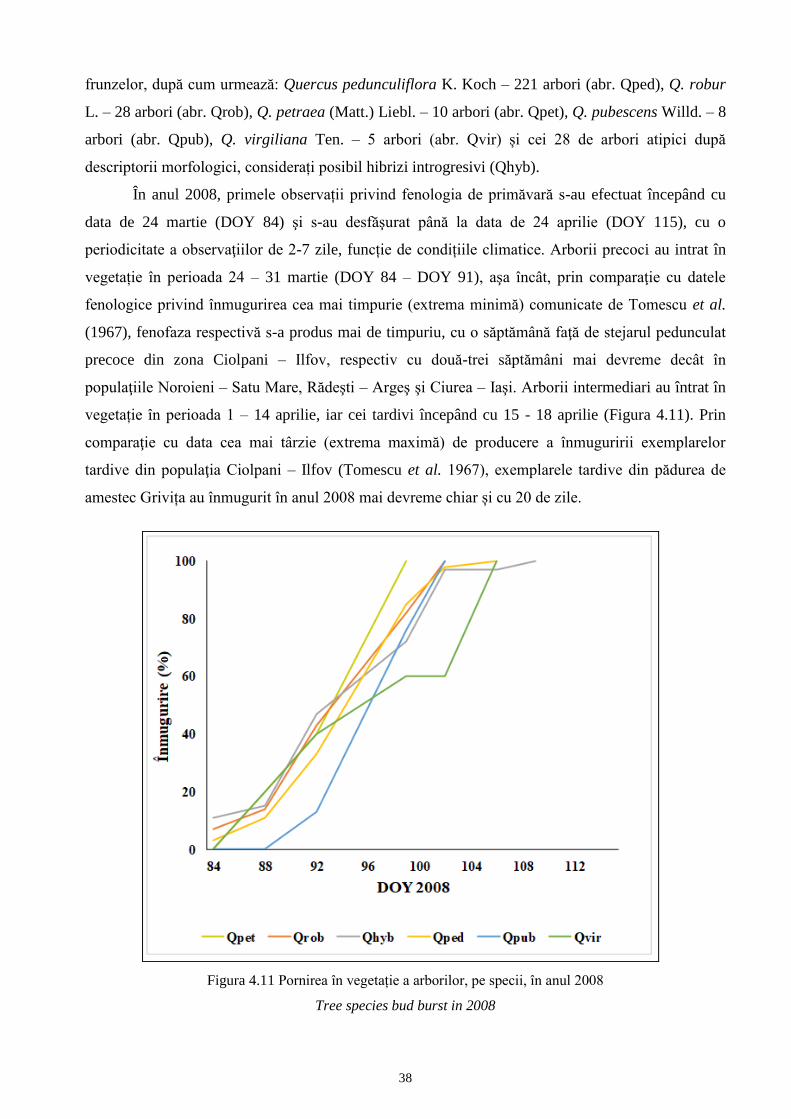
38
frunzelor, după cum urmează: Quercus pedunculiflora K. Koch – 221 arbori (abr. Qped), Q. robur
L. – 28 arbori (abr. Qrob), Q. petraea (Matt.) Liebl. – 10 arbori (abr. Qpet), Q. pubescens Willd. – 8
arbori (abr. Qpub), Q. virgiliana Ten. – 5 arbori (abr. Qvir) și cei 28 de arbori atipici după
descriptorii morfologici, considerați posibil hibrizi introgresivi (Qhyb).
În anul 2008, primele observații privind fenologia de primăvară s-au efectuat începând cu
data de 24 martie (DOY 84) şi s-au desfăşurat până la data de 24 aprilie (DOY 115), cu o
periodicitate a observaţiilor de 2-7 zile, funcție de condițiile climatice. Arborii precoci au intrat în
vegetație în perioada 24 – 31 martie (DOY 84 – DOY 91), aşa încât, prin comparaţie cu datele
fenologice privind înmugurirea cea mai timpurie (extrema minimă) comunicate de Tomescu et al.
(1967), fenofaza respectivă s-a produs mai de timpuriu, cu o săptămână faţă de stejarul pedunculat
precoce din zona Ciolpani – Ilfov, respectiv cu două-trei săptămâni mai devreme decât în
populaţiile Noroieni – Satu Mare, Rădeşti – Argeş şi Ciurea – Iaşi. Arborii intermediari au întrat în
vegetație în perioada 1 – 14 aprilie, iar cei tardivi începând cu 15 - 18 aprilie (Figura 4.11). Prin
comparaţie cu data cea mai târzie (extrema maximă) de producere a înmuguririi exemplarelor
tardive din populaţia Ciolpani – Ilfov (Tomescu et al. 1967), exemplarele tardive din pădurea de
amestec Grivița au înmugurit în anul 2008 mai devreme chiar și cu 20 de zile.
Figura 4.11 Pornirea în vegetație a arborilor, pe specii, în anul 2008
Tree species bud burst in 2008
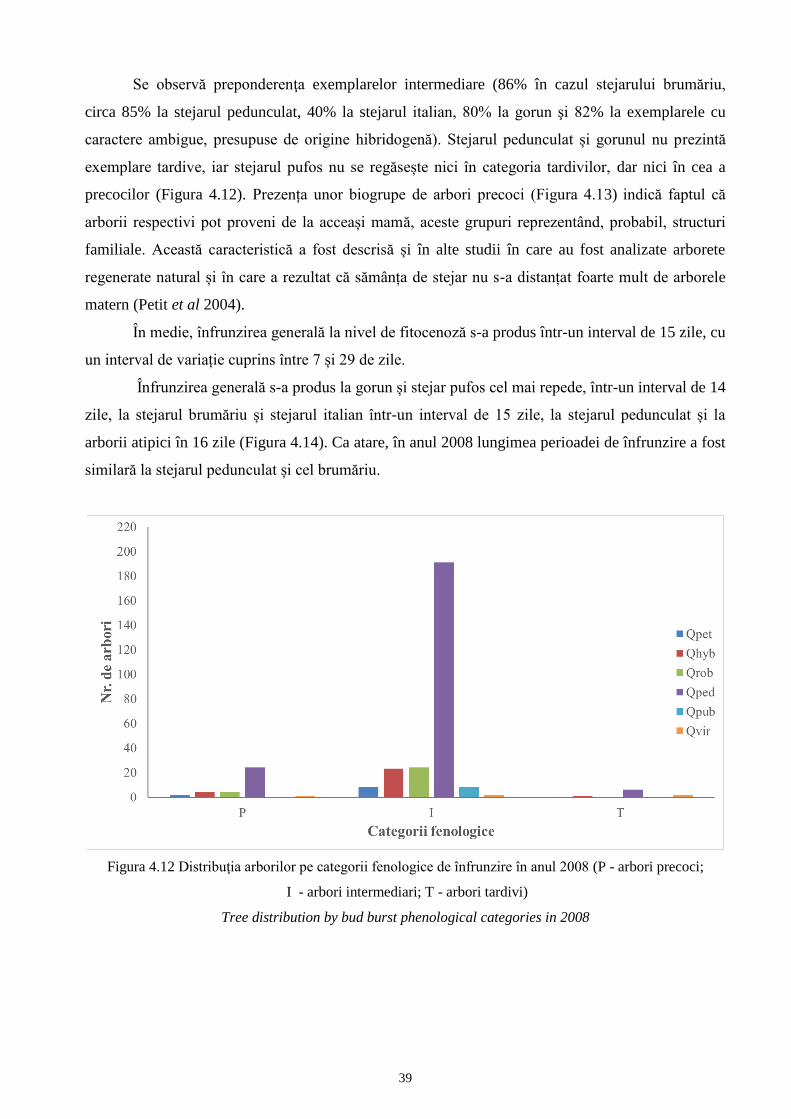
39
Se observă preponderenţa exemplarelor intermediare (86% în cazul stejarului brumăriu,
circa 85% la stejarul pedunculat, 40% la stejarul italian, 80% la gorun şi 82% la exemplarele cu
caractere ambigue, presupuse de origine hibridogenă). Stejarul pedunculat și gorunul nu prezintă
exemplare tardive, iar stejarul pufos nu se regăsește nici în categoria tardivilor, dar nici în cea a
precocilor (Figura 4.12). Prezența unor biogrupe de arbori precoci (Figura 4.13) indică faptul că
arborii respectivi pot proveni de la acceași mamă, aceste grupuri reprezentând, probabil, structuri
familiale. Această caracteristică a fost descrisă și în alte studii în care au fost analizate arborete
regenerate natural și în care a rezultat că sămânța de stejar nu s-a distanțat foarte mult de arborele
matern (Petit et al 2004).
În medie, înfrunzirea generală la nivel de fitocenoză s-a produs într-un interval de 15 zile, cu
un interval de variație cuprins între 7 și 29 de zile.
Înfrunzirea generală s-a produs la gorun și stejar pufos cel mai repede, într-un interval de 14
zile, la stejarul brumăriu și stejarul italian într-un interval de 15 zile, la stejarul pedunculat și la
arborii atipici în 16 zile (Figura 4.14). Ca atare, în anul 2008 lungimea perioadei de înfrunzire a fost
similară la stejarul pedunculat și cel brumăriu.
Figura 4.12 Distribuţia arborilor pe categorii fenologice de înfrunzire în anul 2008 (P - arbori precoci;
I - arbori intermediari; T - arbori tardivi)
Tree distribution by bud burst phenological categories in 2008
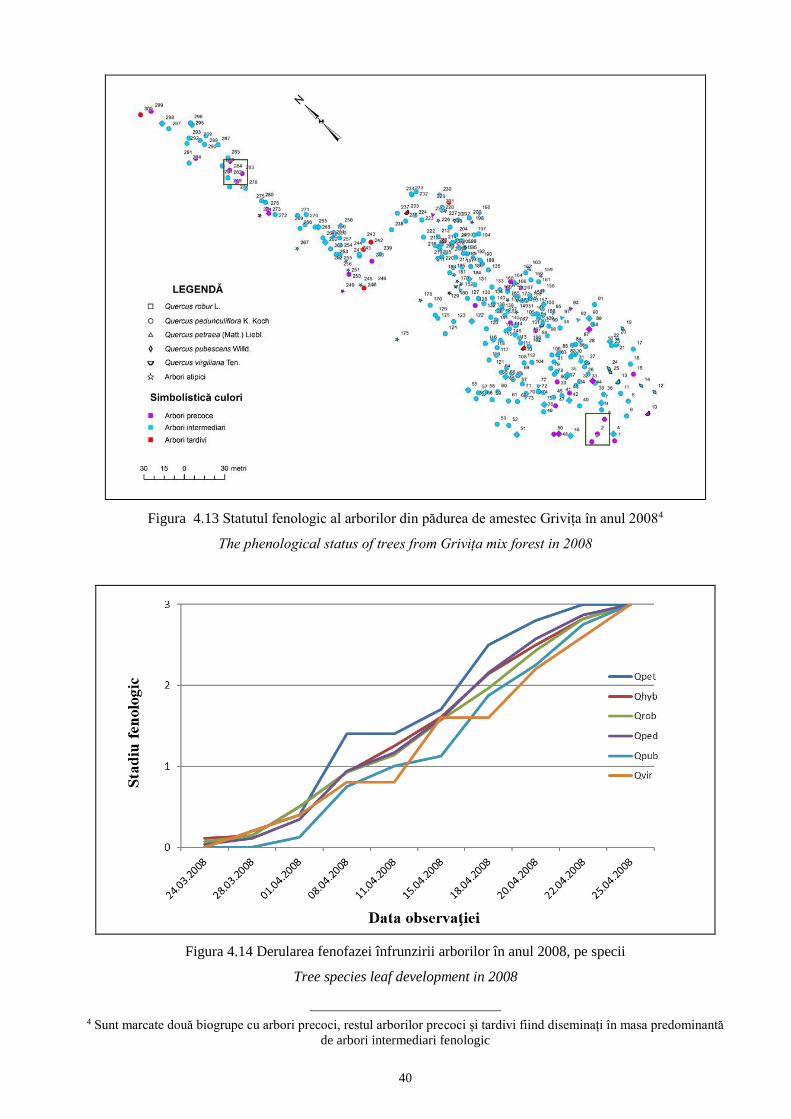
40
Figura 4.13 Statutul fenologic al arborilor din pădurea de amestec Grivița în anul 20084
The phenological status of trees from Grivița mix forest in 2008
Figura 4.14 Derularea fenofazei înfrunzirii arborilor în anul 2008, pe specii
Tree species leaf development in 2008
4 Sunt marcate două biogrupe cu arbori precoci, restul arborilor precoci și tardivi fiind diseminați în masa predominantă
de arbori intermediari fenologic

41
În primăvara anului 2009, primele observații privind fenologia de primăvară s-au efectuat
începând cu data de 11 aprilie şi s-au desfăşurat până la data de 4 mai, cu o periodicitate a
observaţiilor de 2 - 4 zile, funcție de condițiile climatice. Arborii precoci au intrat în vegetație în
perioada 11 – 14 aprilie, cei intermediari în perioada 15 – 17 aprilie, iar cei tardivi începând cu 18
aprilie. Se observă și de această dată preponderenţa exemplarelor intermediare (77% în cazul
stejarului brumăriu, circa 89% la stejarul pedunculat, 80% la stejarul italian, şi 93 % la exemplarele
cu caractere ambigue, presupuse de origine hibridogenă). Excluzând stejarul brumăriu și
exemplarele atipice, toți ceilalți taxoni nu prezintă exemplare tardive. Stejarul pufos nu prezintă
exemplare precoce, în schimb gorunul prezintă predominant exemplare precoce (60%). Ca și în anul
anterior de observații, se observă biogrupe cu arbori precoci. În medie, înfrunzirea generală la nivel
de fitocenoză s-a produs într-un interval de 10 zile, cu un interval de variație cuprins între 7 și 17
zile. Înfrunzirea generală s-a produs cel mai repede la stejarul italian (9 zile) și cel mai târziu la
stejarul pufos (11 zile), ceilalți taxoni înfrunzind în intervalul mediu la nivel de fitocenoză (10 zile).
Chiar dacă cei doi taxoni analizați prezintă același interval de înfrunzire, ca și în primăvara anului
2008, stejarul pedunculat nu a avut exemplare tardive.
În anul 2015, deplasările în teren au început în data de 31 martie (DOY 90) şi s-au
desfăşurat până la data de 8 mai (DOY 128), cu o periodicitate a observaţiilor de 3-5 zile. Arborii
precoci au intrat în vegetație în perioada 31 martie - 4 aprilie (DOY 90 – DOY 94), cei intermediari
în perioada 5 – 8 aprilie (DOY 95 – DOY 98), iar cei tardivi în perioada 9 – 20 aprilie (DOY 99 –
DOY 110) (Figura 4.24). Se observă preponderenţa exemplarelor intermediare (75% în cazul
stejarului brumăriu, circa 89% la stejarul pedunculat, 60% la stejarul italian, 40% la gorun, 80% la
stejarul pufos şi 68 % la exemplarele cu caractere ambigue, presupuse de origine hibridogenă).
Stejarul pedunculat, stejarul pufos și stejarul italian nu prezintă exemplare precoce. În medie,
înfrunzirea generală la nivel de fitocenoză s-a produs într-un interval de 15 zile, cu un interval de
variație cuprins între 9 și 23 de zile. Înfrunzirea generală s-a produs la gorun cel mai repede, într-un
interval de 12 zile, la stejarul brumăriu, stejarul italian și arborii atipici într-un interval de 13 zile, la
stejarul pedunculat în 15 zile, iar la stejarul pufos în 16 zile. Dacă în toți anii de observații, pe lângă
arborii intermadiari, stejarul brumăriu a prezentat atât arbori precoci, cât și tardivi, stejarul
pedunculat, fie nu a prezentat arbori tardivi (2008 și 2009), fie precoci (2015). De remarcat este însă
faptul că, odată cu lipsa exemplarelor precoce de stejar pedunculat, se observă formarea de biogrupe
de stejar brumăriu.
Următorul an (2016), deplasările în teren au început în data de 24 martie şi s-au desfăşurat
până la data de 5 mai, cu o periodicitate a observaţiilor de 3-5 (7) zile. Funcție de data la care
arborii au înregistrat stadiul I, aceștia au fost încadrați ca precoci (24 martie – 4 aprilie),
intermediari (5 – 12 aprilie) și tardivi începând cu 13 aprilie. Distribuția arborilor pe categorii

42
fenologice s-a realizat ținând cont de condiţia de preponderenţă a acestora, având ca argument
evaluări anterioare publicate în acest sens (Chesnoiu et al. 2009).
Ca o consecinţă a modalităţii de delimitare a categoriilor fenologice legate de înmugurire se
constată preponderenţa exemplarelor intermediare (85% în cazul stejarului brumăriu, circa 89% la
stejarul pedunculat, 60% la stejarul italian, 70% la gorun şi 75% la exemplarele cu caractere
ambigue, presupuse de origine hibridogenă). Stejarul pufos nu a prezentat variabilitate în ceea ce
privește înmugurirea, toate exemplarele înmugurind într-un interval foarte scurt de timp. Totodată,
din analiza de ansamblu a structurii fenologice referitoare la înfrunzire s-a constatat suprapunerea
fenofazei de declanşare a înmuguririi la stejarul brumăriu, gorun şi exemplarele prezumtiv hibride.
Stejarul brumăriu, taxonul cel mai bine reprezentat în fitocenoză, a înmugurit integral în
perioada 24 martie – 13 aprilie, cu o preponderență în perioada 6 – 8 apilie (DOY 97 – DOY 99),
corespunzătoare exemplarelor intermediare fenotipic însă, prezintă arbori specifici tuturor celor trei
categorii fenologice de înfrunzire. Stejarul pedunculat este şi el cel mai bine reprezentat de
exemplarele intermediare, însă prezintă arbori şi din categoriile precoci și tardivi. Stejarul pufos are
doar arbori intermediari fenologic. Stejarul italian şi gorunul au arbori fie precoci (40%, respectiv
30%), fie doar intermediari. Ca stejarul brumăriu, stejarul pedunculat și exemplarele atipice au
arbori corespunzători celor trei categorii fenologice. Se remarcă, de asemenea, biogrupe de arbori
precoci în cadrul fitocenozei. Din analiza duratei medii a perioadei de înfrunzire în primăvara
anului 2016, în pădurea de amestec Grivița nu au rezultat diferenţe semnificative între taxonii de
cvercinee analizaţi. În medie, fenofaza respectivă s-a derulat în circa 19 zile la Qped, Qrob și Qhyb,
18 zile la Qpet și Qvir și 17 zile la Qpub.
S-a constatat însă că, indiferent de apartenenţa taxonomică, durata de înfrunzire descreşte de
la precoci spre tardivi. Altfel spus, exemplarele care pornesc mai repede în vegetaţie au o dinamică
mai lentă a înfrunzirii decât cele la care fenofaza respectivă se declanşază mai târziu. Acest fapt
poate fi pus în legătură cu mersul vremii, întrucât temperaturile ridicate înregistrate şi acumulate la
un moment dat determină înfrunzirea explozivă a arborilor, inclusiv a celor tardivi.
4.3.1.2 Analiza fenologiei de primăvară privind înflorirea în pădurea de amestec Grivița
În anul 2008, înflorirea cvercineelor din pădurea de amestec Grivița a început în data de 20
aprilie (DOY 111), la un interval de 27 de zile față de intrarea în vegetație a primelor exemplare
(arborii precoci), și după o perioadă cu precipitații abundente (11,3 mm în 15 aprilie), urmată de
zile cu temperaturi medii cuprinse între 130C și 180C. În primele două zile au înflorit aproximativ
54% dintre arbori, corespunzători tuturor taxonilor evaluați, în următoarele două zile au înflorit și
ceilalți arbori, înregistrându-se astfel o înflorire explozivă.
Pentru înflorirea generală, stejarul pedunculat, stejarul pufos și stejarul italian au avut nevoie

43
de 3-4 zile, gorunul de 5 zile, iar stejarul brumăriu și arborii atipici de 6 zile, realizându-se în ziua
de 25 aprilie o înflorire generală a tuturor taxonilor evaluați în fitocenoza Grivița. Această
suprapunere a fenofazei înfloririi scoate în evidență lipsa unei bariere prezigotice reproductive la
nivel interspecific, ceea ce explică posibilitatea interfecundării, și deci, a prezenței posibililor
hibrizi introgresivi în această pădure de amestec.
În primăvara anului 2009, primele exemplare de cvercinee au înflorit începând cu data de 22
aprilie (DOY 112), la un interval de 11 zile față de intrarea în vegetație a primelor exemplare
(arborii precoci), și după 7 zile consecutive cu temperturi peste 130C, iar ultimele exemplare au
început să înflorească în data de 4 mai (DOY 124), deci la un interval de 12 zile. Dintre cei 300 de
arbori, 16 (5%) nu au înflorit, manifestând androsterilitate (11 stejari brumării, 4 arbori atipici și 1
stejar pedunculat). Procentul arborilor sterili se încadrează în intervalul de 2-8%, semnalat într-un
studiu multianual, în care a fost evaluat un arboret de amestec de stejar pedunculat și gorun
(Bacilieri et al. 1994).
Dintre arborii fertili, 51% au început să înflorească în data de 22 aprilie, toți taxonii fiind
reprezentați de exemplare care au eliberat polen începând cu această dată. Dispersia polenului la
nivel de fitocenoză a durat în medie 6 zile (5 zile la stejarul pedunculat și stejarul pufos, 6 zile la
stejarul brumăriu, stejarul italian și arborii atipici și 8 zile la gorun).
Arborii precoci au început să înflorească în perioada 22 – 29 aprilie, iar cei tardivi în
perioada 29 aprilie – 2 mai. De precizat faptul că, în data de 23 aprilie s-a înregistrat o temperatură
minimă negativă (-10C) care a afectat parțial inflorescențele arborilor de pe liziera pădurii și pe cele
din partea superioară a coroanelor arborilor. Faptul că în jurul datei de 1 mai, peste 96% din arbori
au dispersat polen, se explică prin prezența unei perioade de 4 zile (27 – 30 aprilie) în care s-au
înregistrat din nou temperaturi peste 130C, precedată însă și de precipitații semnificative în data de
25 aprilie (9,9 mm).
În primăvara anului 2015, primele exemplare de cvercinee au înflorit începând cu data de 15
aprilie (DOY 105), la un interval de 15 zile față de intrarea în vegetație a primelor exemplare
(arborii precoci), și după 7 zile consecutive cu temperturi peste 100C, iar ultimele exemplare au
început să înflorească în data de 5 mai (DOY 125), deci la un interval de 20 zile (Figura 4.15).
Dintre cei 300 de arbori, 10 (3%) nu au înflorit, manifestând androsterilitate (6 stejari brumării, 2
arbori atipici și 1 stejar pedunculat).
Dintre arborii fertili, doar 2% au început să înflorească în 5 aprilie, cei mai mulți, 55% au
început să înflorească în data de 10 aprilie, toți taxonii fiind reprezentați de exemplare care au
eliberat polen începând cu această dată. Perioada de începere a dispersiei polenului la nivel de
fitocenoza a durat în medie 12 zile (15 zile la stejarul pedunculat și stejarii atipici, 20 de zile la
stejarul brumăriu, 7 zile la stejarul italian și stejarul pufos și 9 zile la gorun).
44
În perioada înfloririi (15 aprilie – 5 mai) s-au înregistrat 5 zile cu temperaturi negative. În
plin proces de înflorire, în data de 23 aprilie, ca și în anul 2009, s-a înregistrat o temperatură
minimă negativă (-30C) care a afectat semnificativ inflorescențele arborilor de pe liziera pădurii și
pe cele din partea superioară a coroanelor arborilor. Faptul că în jurul datei de 1 mai, peste 93% din
arborii tuturor speciilor au dispersat polen, se explică prin prezența unei perioade de 7 zile (24 – 30
aprilie) în care s-au înregistrat temperaturi peste 140C și de prezența precipitațiilor în zilele de 28 și
29 aprilie (1,7 mm, respectiv 2,5 mm).
În primăvara anului 2016, primele exemplare de cvercinee au înflorit începând cu data de 13
aprilie (DOY 104), la un interval de 19 zile față de intrarea în vegetație a primelor exemplare
(arborii precoci), și după 9 zile consecutive cu temperaturi 150C, și peste această valoarea, iar
ultimele exemplare au început să înflorească în data de 5 mai (DOY 126), deci la un interval de 22
zile (Figura 4.16). Dintre cei 300 de arbori, 8 arbori (2,6%) nu au înflorit, manifestând
androsterilitate (6 stejari brumării și 2 arbori atipici).
Dintre arborii fertili, cei mai mulți, 27% au început să înflorească în data de 22 aprilie (DOY
113), toți taxonii find reprezentați de exemplare care au eliberat polen începând cu această dată.
Începutul dispersiei polenului la nivel de fitocenoză a durat în medie 16,5 zile (22 de zile la stejarul
brumăriu, stejarul pedunculat și stejarii atipici, 6 zile la stejarul italian, 11 zile la stejarul pufos și
16 zile la gorun).
În perioada înfloririi (13 aprilie – 5 mai) nu s-au înregistrat temperaturi negative. În jurul datei
de 5 mai, peste 93% din arborii tuturor speciilor au dispersat polen, un factor important care a
stimulat înflorirea intensivă fiind și prezența precipitațiilor din perioada 3-5 mai (între 2,5 și 10,7
mm).
4.15 Începutul înfloririi arborilor pe specii în anul
2015
The beginning of tree flowering by species in
2015
4.16 Începutul înfloririi arborilor în anul 2016, pe
specii
The beginning of tree flowering by species in
2016
45
4.3.1.3 Analiza fenologiei de primăvară privind înfrunzirea în plantajele Mândrești - Tecuci și
Cobadin - Constanța
În cadrul Plantajului Mândrești s-au făcut observații în anii 2015 și 2016. În anul 2015,
deplasările în teren au început în data de 31 martie 2015 şi s-au desfăşurat până la data de 5 mai
2015, cu o periodicitate a observaţiilor de 3-5 zile. Clonele precoce au intrat în vegetație înainte de
30 martie, cele intermediare în perioada 5 – 8 aprilie, iar cele tardive după această ultimă dată
(Figura 4.17, Figura 4.18). Se observă preponderenţa clonelor intermediare, dintre cele 42 de clone
37 (90,5%) sunt intermediare, celelalte 5 clone fiind precoce (1%) sau tardive (8,5%).
În anul 2016, deplasările în teren au început în data de 30 martie 2016 şi s-au desfăşurat
până la data de 28 aprilie 2016, cu o periodicitate a observaţiilor de 3-5 zile. Clonele precoce au
intrat în vegetație înainte de 30 martie, cele intermediare în perioada 31 martie – 9 aprilie, iar cele
tardive, după această ultimă dată (Figura 4.17).
În anul 2016, în plantajul Cobadin au fost evaluați 212 rameți aparținând la 16 de clone de
stejar brumăriu și 17 clone de stejar pedunculat. Primele înregistrări fenologice s-au efectuat în
plantaj începând cu data de 22 martie 2016 şi s-au desfăşurat până la data de 3 mai 2016, cu o
periodicitate a observaţiilor de 3-5 (7) zile (Figura 4.19). Clonele precoce au intrat în vegetație până
la data de 22 martie, cele intermediare în perioada 23 martie – 11 aprilie, iar cele tardive, după
această ultimă dată. (Figura 4.20).
Figura 4.17 Derularea fenofazei înfrunzirii clonelor
de stejar pedunculat în anii 2015 și 2016 în plantajul
Mândrești
Pedunculate oak clones leaf development in the years
2015 and 2016 in Mândrești seed orchard
Figura 4.18 Distribuţia clonelor pe categorii
fenologice de înfrunzire în anul 2015 în plantajul
Mândrești
Clones distribution by bud burst phenological
categories in 2015 in Mândrești seed orchard
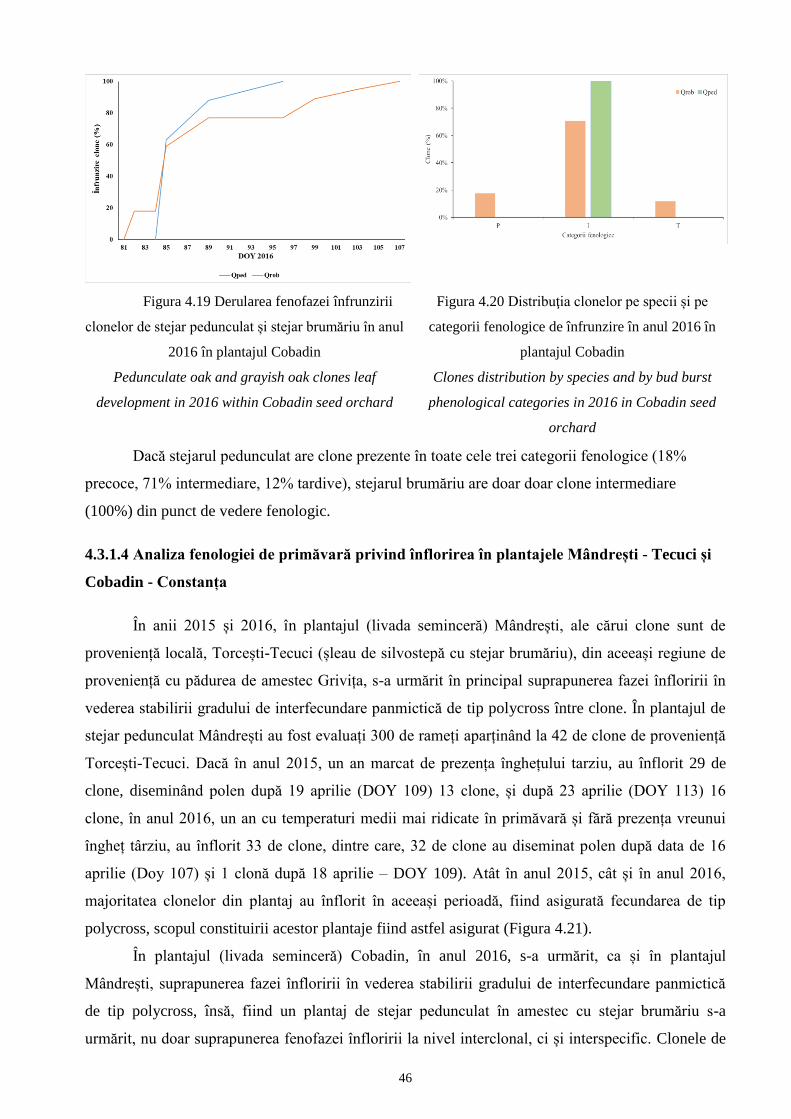
46
Figura 4.19 Derularea fenofazei înfrunzirii
clonelor de stejar pedunculat și stejar brumăriu în anul
2016 în plantajul Cobadin
Pedunculate oak and grayish oak clones leaf
development in 2016 within Cobadin seed orchard
Figura 4.20 Distribuţia clonelor pe specii și pe
categorii fenologice de înfrunzire în anul 2016 în
plantajul Cobadin
Clones distribution by species and by bud burst
phenological categories in 2016 in Cobadin seed
orchard
Dacă stejarul pedunculat are clone prezente în toate cele trei categorii fenologice (18%
precoce, 71% intermediare, 12% tardive), stejarul brumăriu are doar doar clone intermediare
(100%) din punct de vedere fenologic.
4.3.1.4 Analiza fenologiei de primăvară privind înflorirea în plantajele Mândrești - Tecuci și
Cobadin - Constanța
În anii 2015 și 2016, în plantajul (livada seminceră) Mândrești, ale cărui clone sunt de
proveniență locală, Torcești-Tecuci (șleau de silvostepă cu stejar brumăriu), din aceeași regiune de
proveniență cu pădurea de amestec Grivița, s-a urmărit în principal suprapunerea fazei înfloririi în
vederea stabilirii gradului de interfecundare panmictică de tip polycross între clone. În plantajul de
stejar pedunculat Mândrești au fost evaluați 300 de rameți aparținând la 42 de clone de proveniență
Torcești-Tecuci. Dacă în anul 2015, un an marcat de prezența înghețului tarziu, au înflorit 29 de
clone, diseminând polen după 19 aprilie (DOY 109) 13 clone, și după 23 aprilie (DOY 113) 16
clone, în anul 2016, un an cu temperaturi medii mai ridicate în primăvară și fără prezența vreunui
îngheț târziu, au înflorit 33 de clone, dintre care, 32 de clone au diseminat polen după data de 16
aprilie (Doy 107) și 1 clonă după 18 aprilie – DOY 109). Atât în anul 2015, cât și în anul 2016,
majoritatea clonelor din plantaj au înflorit în aceeași perioadă, fiind asigurată fecundarea de tip
polycross, scopul constituirii acestor plantaje fiind astfel asigurat (Figura 4.21).
În plantajul (livada seminceră) Cobadin, în anul 2016, s-a urmărit, ca și în plantajul
Mândrești, suprapunerea fazei înfloririi în vederea stabilirii gradului de interfecundare panmictică
de tip polycross, însă, fiind un plantaj de stejar pedunculat în amestec cu stejar brumăriu s-a
urmărit, nu doar suprapunerea fenofazei înfloririi la nivel interclonal, ci și interspecific. Clonele de
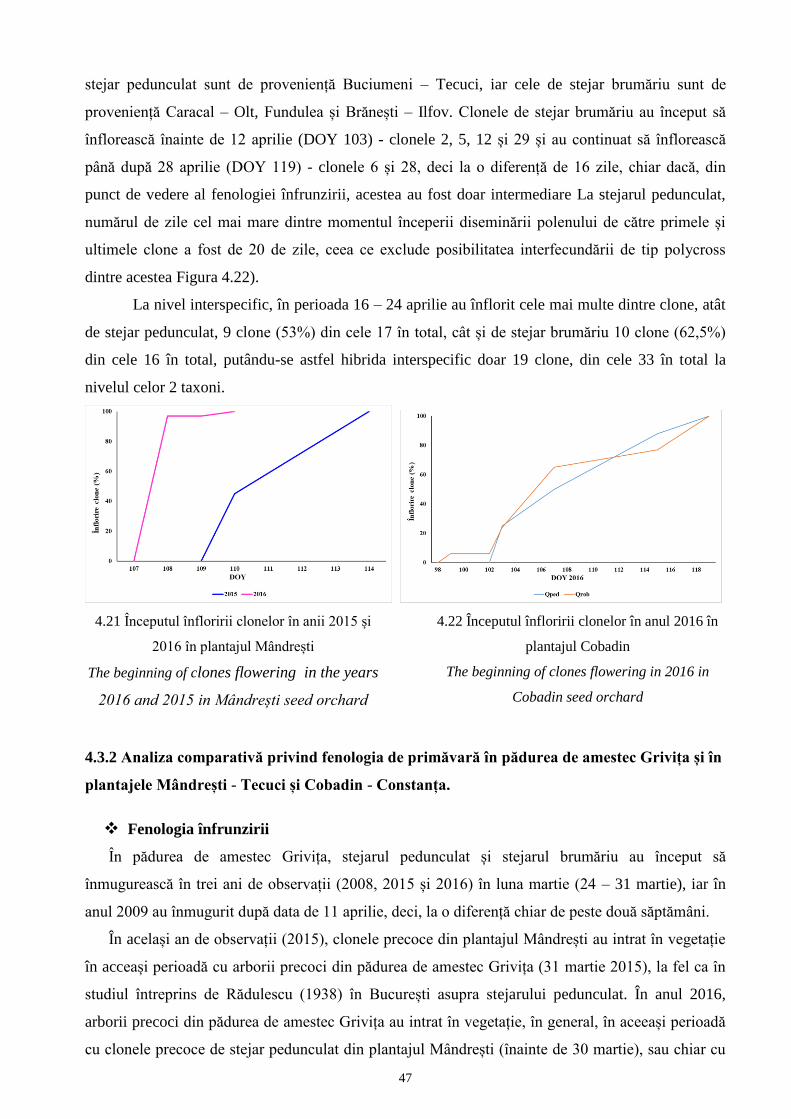
47
stejar pedunculat sunt de proveniență Buciumeni – Tecuci, iar cele de stejar brumăriu sunt de
proveniență Caracal – Olt, Fundulea și Brănești – Ilfov. Clonele de stejar brumăriu au început să
înflorească înainte de 12 aprilie (DOY 103) - clonele 2, 5, 12 și 29 și au continuat să înflorească
până după 28 aprilie (DOY 119) - clonele 6 și 28, deci la o diferență de 16 zile, chiar dacă, din
punct de vedere al fenologiei înfrunzirii, acestea au fost doar intermediare La stejarul pedunculat,
numărul de zile cel mai mare dintre momentul începerii diseminării polenului de către primele și
ultimele clone a fost de 20 de zile, ceea ce exclude posibilitatea interfecundării de tip polycross
dintre acestea Figura 4.22).
La nivel interspecific, în perioada 16 – 24 aprilie au înflorit cele mai multe dintre clone, atât
de stejar pedunculat, 9 clone (53%) din cele 17 în total, cât și de stejar brumăriu 10 clone (62,5%)
din cele 16 în total, putându-se astfel hibrida interspecific doar 19 clone, din cele 33 în total la
nivelul celor 2 taxoni.
4.21 Începutul înfloririi clonelor în anii 2015 și
2016 în plantajul Mândrești
The beginning of clones flowering in the years
2016 and 2015 in Mândrești seed orchard
4.22 Începutul înfloririi clonelor în anul 2016 în
plantajul Cobadin
The beginning of clones flowering in 2016 in
Cobadin seed orchard
4.3.2 Analiza comparativă privind fenologia de primăvară în pădurea de amestec Grivița și în
plantajele Mândrești - Tecuci și Cobadin - Constanța.
❖ Fenologia înfrunzirii
În pădurea de amestec Grivița, stejarul pedunculat și stejarul brumăriu au început să
înmugurească în trei ani de observații (2008, 2015 și 2016) în luna martie (24 – 31 martie), iar în
anul 2009 au înmugurit după data de 11 aprilie, deci, la o diferență chiar de peste două săptămâni.
În același an de observații (2015), clonele precoce din plantajul Mândrești au intrat în vegetație
în acceași perioadă cu arborii precoci din pădurea de amestec Grivița (31 martie 2015), la fel ca în
studiul întreprins de Rădulescu (1938) în București asupra stejarului pedunculat. În anul 2016,
arborii precoci din pădurea de amestec Grivița au intrat în vegetație, în general, în aceeași perioadă
cu clonele precoce de stejar pedunculat din plantajul Mândrești (înainte de 30 martie), sau chiar cu

48
4 zile mai târziu (4 aprilie). Tot în anul 2016, în plantajul Cobadin, clonele precoce (doar de stejar
pedunculat) au intrat în vegetație până la data de 22 martie, cu opt zile mai devreme față de clonele
precoce din plantajul Mândrești, sau chiar cu până la două sâptămâni mai devreme față de unii
arbori precoci din pădurea de amestec Grivița.
❖ Fenologia înfloririi
În ceea ce privește înflorirea arborilor din pădurea de amestec Grivița, se poate spune că în
toți cei patru ani de observații există perioade de suprapunere a fenofazei înfloririi generale la toți
taxonii, iar această perioadă este de obicei în jurul datei de 22 aprilie. În anii mai călduroși, precum
2015 și 2016, au fost și exemplare care au început să înflorească încă din 13 – 15 aprilie, însă în toți
anii niciun exemplar nu a mai înflorit după data de 5 mai. În anul 2009, când arborii au intrat în
vegetație chiar și cu două săptămâni mai târziu față de ceilalți ani, înflorirea generală a avut loc tot
în aceeași perioadă (22 aprilie), putându-se constata faptul că arborii care intră în vegetație mai
târziu prezintă o dinamică fenologică ridicată de creștere și dezvoltare.
În plantajul Mândrești, clonele de stejar pedunculat de proveniență Torcești – Tecuci, din
aceeași regiune cu pădurea de amestec Grivița, au înflorit în masă în jurul aceleiași perioade, 19-23
aprilie în 2015 și 16–18 aprilie în 2016. În plantajul Cobadin, clonele de stejar pedunculat au
înflorit mai devreme chiar și cu o săptămână față de cele din plantajul Mândrești și față de arborii
de stejar pedunculat din pădurea de amestec Grivița. Diferența dintre clone privind înflorirea în
plantajul Cobadin a fost și de 20 de zile, probabil ca urmare a influenței originii diferite a clonelor.
Parte din clonele de stejar brumăriu au înflorit anterior arborilor de stejar brumăriu din pădurea de
amestec Grivița, chiar cu 10 zile, altele în aceeași perioadă, sau chiar cu două săptămâni mai târziu.
Distanțele temporale mari între perioadele de înflorire ale clonelor exclud posibilitatea
interfecundării de tip polycross dintre acestea, așa încât scopul constituirii acestei culturi speciale,
de a produce semințe cu însușiri genetice superioare (Șofletea 2005), nu poate fi atins.
4.3.3 Interpretarea corelativă a datelor din cei 4 ani de observații fenologice în pădurea de
amestec Grivița
❖ Calculul corelațiilor privind înfrunzirea
Referitor la proporția exemplarelor intermediare fenologic în raport cu înfrunzirea, la stejarul
pedunculat și cel brumăriu, observațiile din cei patru ani relevă o diferențiere între cei doi taxoni.
Astfel, stejarul pedunculat manifestă constanță, deoarece în perioada evaluată exemplarele
intermediare s-au situat între 85 și 89%, în timp ce la stejarul brumăriu variația a fost mai mare,
între 75 și 86%. Ponderea exemplarelor intermediare a fost mult mai mică la stejarul brumăriu în
anii 2009 și 2015, de 79% și respectiv, 75%. În ceea ce privește corelația înfrunzirii dintre anii de
observații la cei 300 de arbori evaluați, se poate spune ca aceasta este puternic semnificativă,

49
respectiv semnificativă (2015 - 2016), însă cu valori relativ scăzute ale coeficientului de corelație
Spearman. Se poate concluziona faptul că nu toți arborii eșantionați intră în vegetație în aceeași
perioadă a anilor diferiți (Bacilieri et al. 1994).
Intrarea în vegetație, pe specii și între anii de observații, prezintă la stejarul brumăriu, în
toate cazurile corelație pozitivă și puternic semnificativă. Stejarul pedunculat prezintă în toate
cazurile corelație nesemnificativă și chiar negativă între anii 2015 și 2016. Rezultă astfel că,
fenologia înfrunzirii la stejarul brumăriu din pădurea Grivița manifestă o constanță mai mare decât
în cazul stejarului pedunculat. Această afirmație rezultă din analiza generală care indică localizarea
pădurii Grivița în zona de contact cu silvostepa. Gorunul și stejarii atipici prezintă corelație pozitivă
și puternic semnficativă între anii 2008-2009 și 2009-2016. Stejarul pufos prezintă corelație
nesemnificativă între anii 2008 - 2015. Stejarul italian prezintă corelație directă și nesemnificativă
în toate cazurile. Se poate observa că numărul mic de exmplare analizate la nivelul celorlalți taxoni,
în afară de stejarul brumăriu, influențează valorile și semnificația corelațiilor.
❖ Calculul corelațiilor privind înflorirea
În ceea ce privește corelația înfloririi dintre anii de observații la cei 300 de arbori evaluați, se
poate spune ca aceasta este puternic semnificativă, însă cu valori relativ scăzute spre medii ale
coeficientului de corelație Spearman. Acest fapt subliniază faptul că destul de mulți arbori înfloresc
aproximativ în aceeași perioadă a anului, respectiv își păstrează categoria fenologică, aceasta având
și un determinism genetic. Atât stejarul pedunculat, cât și stejarul brumăriu prezintă corelație
semnificativă și foarte semnificativă între perioadele înfloririi din ani diferiți.
❖ Calculul corelațiilor între categoria fenologică de înfrunzire și cea de înflorire, pe ani de
observații
În toți anii, stejarul brumăriu și arborii atipici au prezentat corelație semnificativă și puternic
semnificativă între cele două fenofaze studiate, iar stejarul pedunculat doar în anul 2015. Prin
urmare, momenul deschiderii mugurilor influențează momentul înflorii arborilor.
4.3.4 Interpretarea corelativă a datelor din anii de observații fenologice în plantajele Cobadin
și Mândrești
Atât pentru înfrunzire, cât și pentru înflorire, corelația între clonele de stejar brumăriu și cele
de stejar pedunculat din plantajul Cobadin este nesemnificativă. Semnificativă este însă corelația
înfrunzire-înflorire dintre clonele de stejar pedunculat. Corelația nesemnficiativă dintre clonele de
stejar brumăriu poate fi explicată prin originea diferită a acestora (Caracal – Fundulea – Brănești).
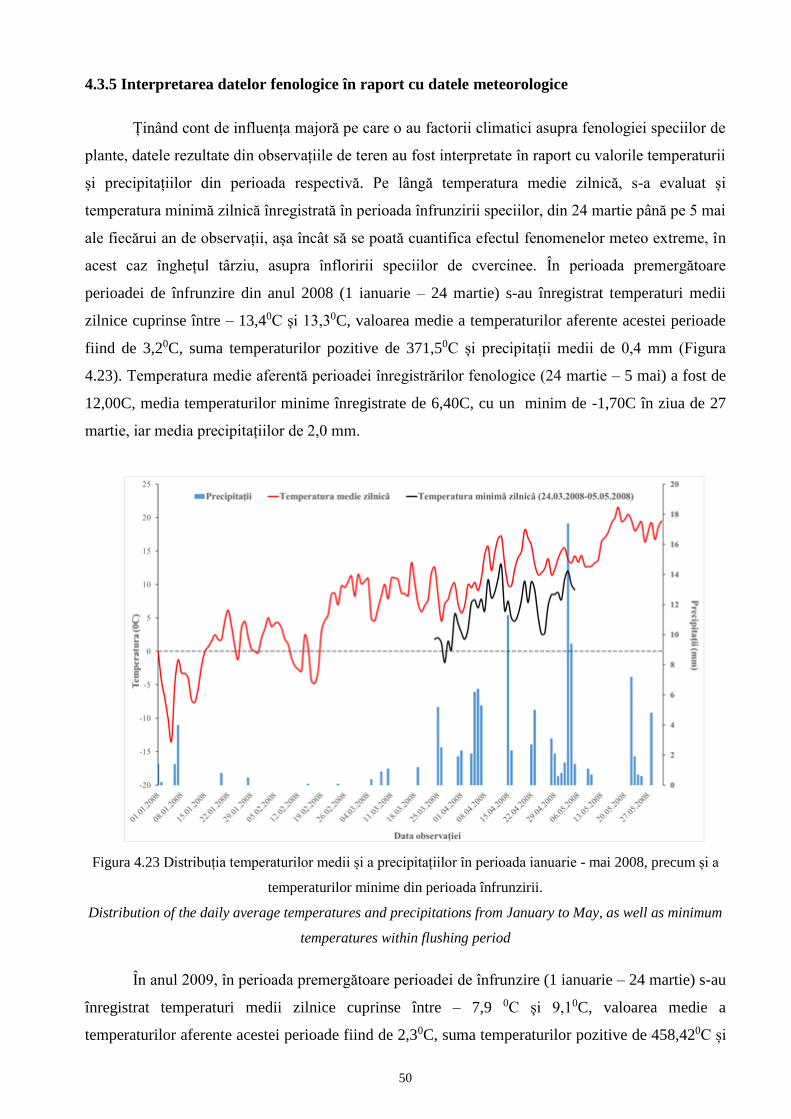
50
4.3.5 Interpretarea datelor fenologice în raport cu datele meteorologice
Ținând cont de influența majoră pe care o au factorii climatici asupra fenologiei speciilor de
plante, datele rezultate din observațiile de teren au fost interpretate în raport cu valorile temperaturii
și precipitațiilor din perioada respectivă. Pe lângă temperatura medie zilnică, s-a evaluat și
temperatura minimă zilnică înregistrată în perioada înfrunzirii speciilor, din 24 martie până pe 5 mai
ale fiecărui an de observații, așa încât să se poată cuantifica efectul fenomenelor meteo extreme, în
acest caz înghețul târziu, asupra înfloririi speciilor de cvercinee. În perioada premergătoare
perioadei de înfrunzire din anul 2008 (1 ianuarie – 24 martie) s-au înregistrat temperaturi medii
zilnice cuprinse între – 13,40C și 13,30C, valoarea medie a temperaturilor aferente acestei perioade
fiind de 3,20C, suma temperaturilor pozitive de 371,50C și precipitații medii de 0,4 mm (Figura
4.23). Temperatura medie aferentă perioadei înregistrărilor fenologice (24 martie – 5 mai) a fost de
12,00C, media temperaturilor minime înregistrate de 6,40C, cu un minim de -1,70C în ziua de 27
martie, iar media precipitațiilor de 2,0 mm.
Figura 4.23 Distribuția temperaturilor medii și a precipitațiilor în perioada ianuarie - mai 2008, precum și a
temperaturilor minime din perioada înfrunzirii.
Distribution of the daily average temperatures and precipitations from January to May, as well as minimum
temperatures within flushing period
În anul 2009, în perioada premergătoare perioadei de înfrunzire (1 ianuarie – 24 martie) s-au
înregistrat temperaturi medii zilnice cuprinse între – 7,9 0C și 9,10C, valoarea medie a
temperaturilor aferente acestei perioade fiind de 2,30C, suma temperaturilor pozitive de 458,420C și

51
precipitații medii de 1,1 mm. Temperatura medie aferentă perioadei înregistrărilor fenologice (24
martie – 5 mai) a fost de 11,80C, media temperaturilor minime înregistrate de 4,70C, cu minime sub
00C în perioada 26 - 27 martie (-3,20C) și pe 23 aprilie (-1,00C), iar media precipitațiilor de 0,7 mm.
În anul 2015, în perioada premergătoare perioadei de înfrunzire (1 ianuarie – 24 martie) s-au
înregistrat temperaturi medii zilnice cuprinse între – 13,6 0C și 9,60C, valoarea medie a
temperaturilor aferente acestei perioade fiind de 2,60C, suma temperaturilor pozitive de 355,460C și
preciptații medii de 0,9 mm. Temperatura medie aferentă perioadei înregistrărilor fenologice (24
martie – 5 mai) a fost de 12,20C, media temperaturilor minime înregistrate de 4,20C, cu minime sub
00C în perioada 24 - 25 martie (-1,70C, respectiv -0,20C), 4 - 5 aprilie (-2,60C, respectiv -2,20C), 10,
15, 20 și respectiv 22 - 24 aprilie, cu temperaturi chiar sub -30C. Media precipitațiilor aferentă
aceleiași perioade a fost de 1,6 mm. În anul 2016, în perioada premergătoare perioadei de înfrunzire
(1 ianuarie – 24 martie) s-au înregistrat temperaturi medii zilnice cuprinse între – 11,30C și 15,20C,
valoarea medie a temperaturilor aferente acestei perioade fiind de 4,20C, suma temperaturilor
pozitive de 453,070C și precipitații medii de 1,0 mm.
Temperatura medie aferentă perioadei înregistrărilor fenologice (24 martie – 5 mai) a fost de
13,90C, media temperaturilor minime înregistrate de 6,70C, cu un minim în data de 26 martie de –
2,90C. Media precipitațiilor în aceeași perioadă a fost de 1,9 mm. În ceea ce privește influența
temperaturilor pozitive, atât cuantumul acestora de la 1 ianuarie și până la momentul înmuguririi
(Stadiul I), cât și al numărului de zile cu temperaturi peste 100C, se poate spune că în toți anii de
observații suma temperaturilor pozitive a fost peste 3550C, iar numărul de zile cu temperaturi peste
100C a fost de 3 în anul 2015 și peste 10 zile în ceilalți ani. De altfel, și suma temperaturilor
pozitive în anul 2015 a fost cea mai mică dintre toți anii. Pentru intrarea în vegetație a stejarului
pedunculat, Tomescu & Bălănică (1953) au semnalat necesitatea unui număr de 12 zile cu
temperaturi peste 100C. Se poate spune că temperatura joacă cel mai important rol în înmugurire,
fapt demonstrat și prin testarea mai multor modele fenologice (Fu et al. 2012).
4.3.6 Analiza xilologică a speciilor de cvercinee din pădurea de amestec Grivița
4.3.6.1 Activitatea cambială și formarea xilemului în anul 2016
În data de 29 martie (DOY 89), chiar dacă nu a început activitatea cambială, la arborele Q.
pedunculiflora 18 s-au putut observa 3-4 celule cambiale. Până la data de 4 aprilie (DOY 95),
numărul celulelor cambiale observate a crescut la 9-10 (Figura 4.24b). Arborii de origine prezumtiv
hibridogenă au prezentat același start al activității cambiale. La celelalte 3 specii, Q. pubescens, Q.
petraea și Q. robur, diviziunea cambială a început mai târziu, între 4 și 10 aprilie, și a continuat
până la sfârșitul lunii iulie la majoritatea speciilor.

52
Fig. 4.24 Formarea xilemului la Quercus pedunculiflora
(a) – creșterea floemului și a xilemului în 2016 (Ew –lemn timpuriu, Lw – lemn târziu, Xyl – xilem, Ph
- floem); (b) - celulele cambiale active în data de 4 aprilie (Cc); (c) - vase din lemnul timpuriu
aflate în proces de dezvoltare în data de 24 aprilie
Development of the xylem in Quercus pedunculiflora: a – xylem and phloem increment
in 2016 (Ew - earlywood, Lw - latewood, Xyl - xylem, Ph-phloem); b - active cambium cells (Cc)
on 4th April; (c) - earlywood vessels in development process on 24th April
Formarea peretelui celular secundar, prin depunerea de lignină a început în a doua jumătate
a lunii aprilie la toate speciile. Procesul de lignificare a continuat la lemnul timpuriu, cambiul a
produs celule noi, formând lemnul târziu. Primele vase ale lemnului târziu au început să se formeze
la mijlocul lunii mai, cu o creștere intensă la sfârșitul lunii iunie și începutul lunii iulie însă, au fost
și arbori la care acestea au început să se formeze chiar și la începutul lunii iulie, continuând să se
extindă până la data de 22 iulie (DOY 204). Pentru Q pedunculiflora 18, Q pubescens 20 și Q robur
295, creșterea radială a încetat în data de 10 iunie (DOY 162). Pentru ceilalți arbori dezvoltarea
xilemului a continuat până pe 22 iulie (DOY 204). Procesul de lignificare a continuat până la
sfârșitul lunii august - începutul lunii septembrie (Figura 4.25).
4.3.6.2 Evaluări comparative între fenologia înfrunzirii, senescența și fenologia cambială
În vederea diferențierii taxonilor, s-au analizat și comparat parametrii anatomici ai acestora
(fibre de lemn, vase). Dezvoltarea celulelor din lemnul timpuriu a avut loc la mijlocul lunii aprilie
(DOY 105), la cele mai multe specii, cu excepția exemplarului prezumtiv hibridogen 168, la care
dezvoltarea vaselor a început mai devreme cu aproximativ 10 zile (DOY 95) și a arborelui Q.
pubescens 20, la care dezvoltarea vaselor a început mai târziu, cu aproximativ 5 zile (DOY 110).
Începutul activității cambiale s-a înregistrat după 29 martie. Variabilitatea fazelor formării
xilemului este înfluențată de condițiile climatice din sezonul de vegetație (Gričar 2013,
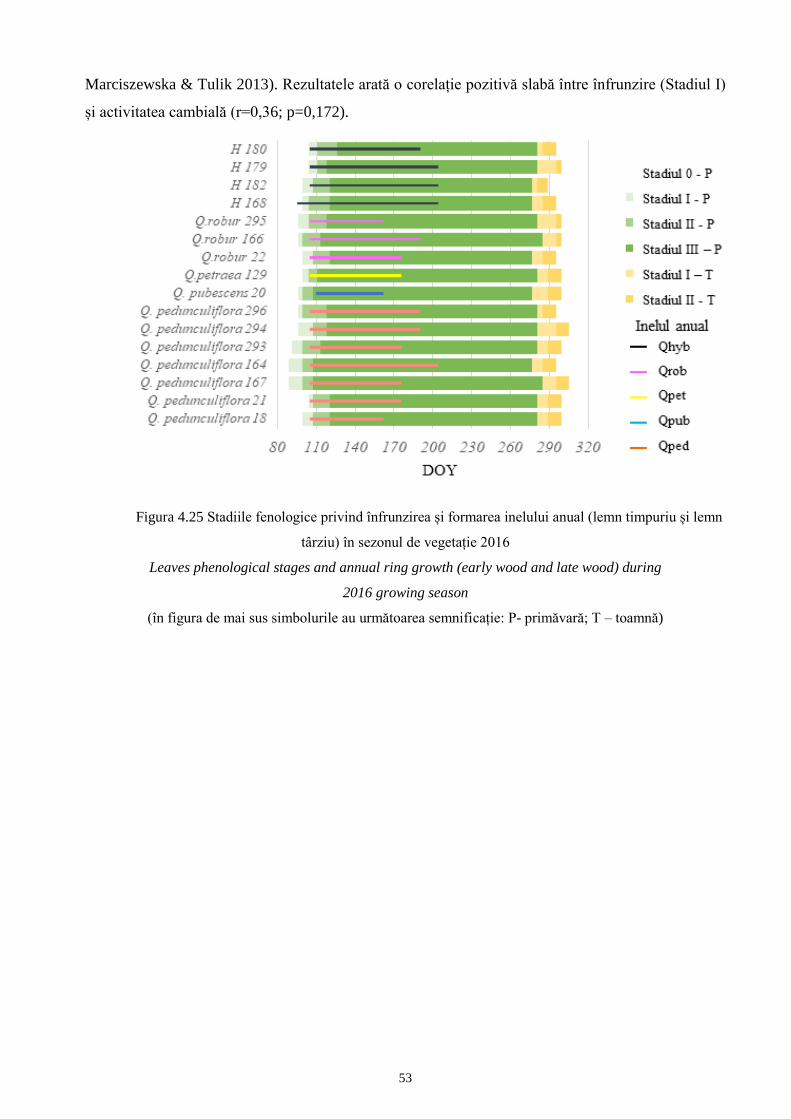
53
Marciszewska & Tulik 2013). Rezultatele arată o corelație pozitivă slabă între înfrunzire (Stadiul I)
și activitatea cambială (r=0,36; p=0,172).
Figura 4.25 Stadiile fenologice privind înfrunzirea și formarea inelului anual (lemn timpuriu și lemn
târziu) în sezonul de vegetație 2016
Leaves phenological stages and annual ring growth (early wood and late wood) during
2016 growing season
(în figura de mai sus simbolurile au următoarea semnificație: P- primăvară; T – toamnă)

54
5. CONCLUZII. CONTRIBUȚII ORIGINALE. DISEMINAREA
REZULTATELOR. DIRECȚII VIITOARE DE CERCETARE
5.1 Concluzii generale
5.1.1 Concluzii privind discriminarea taxonomică prin descriptori ai frunzelor a stejarului
pedunculat şi stejarului brumăriu
➢ Cu privire la evaluarea variabilităţii taxonomice infraspecifice şi a variabilității
fenotipice (intra- şi interpopulaţionale) la nivelul descriptorilor morfologici foliari.
▪ La nivel intraspecific, stejarul pedunculat prezintă variabilitate mai mare din punct de
vedere al numărului de unități infraspecifice identificate, față de stejarul brumăriu. Dacă la
stejarul pedunculat au fost identificate 8 dintre cele 9 unități infraspecifice consemnate în literatura
de specialitate, la stejarul brumăriu au fost identificate 7 dintre cele 10 unități. Anumite varietăți și
forme nu respectă întocmai descrierea din literatura de specialitate, mai ales forma maxima,
subforma frainettoides, care prezintă cel mai adesea frunze obovate, și nu eliptice.
▪ Pe un transect de la sud la nord, numărul unităților infraspecifice la nivelul populațiilor de
stejar pedunculat crește. Predominant întâlnit în ecoregiunea 1B –„Păduri cu stejar pedunculat,
gârniță, cer și alte specii, pe coline mici și câmpii”, un tip de ecoregiune favorabilă acestui taxon
din punct de vedere al condițiilor pedo-staționale, și mult mai rar în ecoregiunea 3A – „Silvostepă
cu stejari xerofiți” (Reșca și Grivița), variabilitatea infrataxonică rămâne aceeași, nefiind afectată
de condițiile mai defavoravile.
▪ Ca urmare a evaluării interpopulaționale, s-au identificat populații valoroase, cu
variabilitate morfologică ridicată, un exemplu fiind populația Braniştea Catârilor - Olt, respectiv
populații cu variabilitate redusă, un exemplu fiind populația Snagov.
➢ Cu privire la identificarea unor caractere de diferențiere între cei doi taxoni, utilizând
analize statistice multivariate.
▪ Funcțiile discriminante obținute cu ajutorul Analizei discriminante permit atât diferențierea
celor doi taxoni evaluați, la care diferențe se înregistrează la nivelul descriptorilor PU, CL, BS, PR
şi LWR, cât și a infrataxonilor acestora. Astfel, în ceea ce privește infrataxonii de stejar
pedunculat, caracterele ce contribuie la diferențiere sunt: lungimea laminei (LL), proporția lățimii
lobilor în raport cu lungimea laminei (LWR), adâncimea sinurilor (SW), numărul de lobi (NL),
ponderea adâncimii lobilor (LDR), procentul de nervare a sinurilor (PV) și numărul de nervuri
intercalare (NV), iar lățimea lobilor (LW), adâncimea sinurilor (SW), lungimea laminei (LL),
numărul de lobi (NL), rata pețiolului (PR) și lungimea pețiolului (PL) sunt caractere de diferențiere
specifice infrataxonilor de stejar brumăriu.

55
▪ Pe baza variabilelor care alcătuiesc funcţia discriminantă dintre cei doi taxoni (PU, CL,
BS, PR şi LWR), Analiza Componentelor Principale a evidențiat formarea celor doi nori de puncte
corespunzători celor doi taxoni evaluați, iar Analiza cluster a grupat populațiile în două mari
clustere: clusterul 1 doar cu populațiile pure de stejar pedunculat din centrul și sud-estul țării, și
clusterul 2, care la rândul lui prezintă 2 subclustere: primul format din toate grupurile de stejar
brumăriu și cel de-al doilea alcătuit din grupurile de stejar pedunculat din sud-estul țării.
5.1.2 Concluzii privind analiza fenologiei de primăvară la stejarul pedunculat şi stejarul
brumăriu
➢ Cu privire la analiza fenologiei de primăvară în scopul identificării derulării
fenofazelor antezei. Ipoteze de analiză: suprapunerea fenofazelor înfloririi ar putea face
posibil fluxul de gene între cei doi taxoni, ceea ce ar presupune posibilitatea interfertilităţii
potenţiale, iar decalajul fenofazelor ar putea sugera individualitatea celor doi taxoni ca specii.
▪ Arborii care pornesc mai repede în vegetaţie au o dinamică mai lentă a înfrunzirii decât cei
la care fenofaza respectivă se declanşază mai târziu. Acest fapt poate fi pus în legătură cu mersul
vremii, întrucât temperaturile ridicate înregistrate şi acumulate la un moment dat determină
înfrunzirea explozivă a arborilor, inclusiv a celor tardivi.
▪ Suprapunerea fenofazei înfloririi scoate în evidență lipsa unei bariere prezigotice
reproductive la nivel interspecific, ceea ce explică posibilitatea interfecundării, și în consecință, a
hibridării, permițând contopirea taxonilor în viitor. Pe de altă parte, datele morfologice sugerează
existența a două entități biologice bine individualizate, context în care deducem existența altor
mecanisme biologice care facilitează împerecherile preferențial la nivel intraspecific, în contextul
în care fenologia infloririi nu concură la separarea celor doi taxoni.
▪ În anii de observații majoritatea clonelor din plantajul Mândrești au înflorit în aceeași
perioadă, fiind asigurată fecundarea de tip polycross, scopul constituirii acestor plantaje, de
manifestare a fenomenului de heterozis, fiind astfel asigurat.
▪ Pentru a întra în vegetație, cvercineele au nevoie în general de o sumă a temperaturilor
pozitive, de la 1 ianuarie și până în momentul înmuguririi, de peste 3550C, precum și de un număr
de peste 10 zile cu temperaturi mai mari de 100C în perioada dinaintea înmuguririi.
▪ După iernile mai călduroase, în care nu se înregistează temperaturi sub -100C, și deci,
plantele nu trec printr-o perioadă de ger, intrarea în vegetație este mai târzie, chiar dacă suma
temperaturilor pozitive este mare și chiar dacă sunt înregistrate și precipitații.
▪ Corelația semnificativă și/sau puternic semnificativă dintre cele două fenofaze, înfrunzirea
și înflorirea arborilor de stejar brumăriu, permite prognozarea momentului diseminării polenului
de către florile mascule în populațiile și plantajele evaluate.

56
▪ Păstrarea categoriei fenologice de înflorire între anii de observații, atât la stejarul
pedunculat, cât și la stejarul brumăriu, presupune că această fenofază are și un determinism
genetic.
▪ Diferențele temporale mari privind înflorirea clonelor din plantajul Cobadin scot în
evidență importanța introducerii criteriului fenologic, și nu doar cel fenotipic, la alegerea arborilor
plus din care se prelevează altoaiele în vederea constituirii descendenților vegetativi pentru
viitoarele plantaje.
➢ Cu privire la evaluarea fazelor formării inelului anual și coroborarea cu categoriile de
arbori precoci, intermediari și tardivi.
▪ Formarea lemnului târziu începe după finalizarea procesului de înflorire.
▪ Perioadele formării inelului anual la stejarul pedunculat și stejarul brumăriu sunt
asemănătoare.
➢ Cu privire la identificarea de populații valoroase, după criteriul diversității fenotipice,
sau cel al existenței unor fenotipuri cu potențial ridicat de adaptare la condițiile climatice
locale, inclusiv pentru formularea de recomandări privind strategia de conservare a
genofondurilor de stejar pedunculat și stejar brumăriu din România.
▪ Un caz particular, populația Viișoara, singura fitocenoză din stepă prezintă o variabilitate
morfologică foarte ridicată, chiar în aceste condiții extreme de vegetație. În acest context este
iminentă conservarea in situ și ex situ a acestei populații valoroase în condițiilor climatice viitoare
din silvostepă și se recomandă instalarea uni plantaj cu proveniențe din această fitocenoză, în
vederea asigurării necesarului de material de reproducere în cadrul aceleiași regiuni de
proveniență.
▪ Conservarea in situ, ex situ și migrația asistată a proveniențelor valoroase de cvercinee, cu
plasticitate și adaptabilitate sporită, și cu capacități bune de producție este o condiție prealabilă
pentru asigurarea stabilității ecosistemelor forestiere în contextul schimbărilor climatice și al
influenței antropice.
▪ Ca o comparație a înfloririi clonelor de stejar pedunculat din cele două plantaje, Mândrești
și Cobadin, se poate afirma că suprapunerea fenofazelor în plantajul Mândrești se explică prin
instalarea acestuia în aceeași regiune de proveniență cu arboretul de unde s-au prelevat ramurile
altoi, pe când, în plantajul Cobadin clonele de stejar pedunculat de proveniență Buciumeni (H2) au
fost instalate în altă regiune de proveniență (I), acestea manifestând un comportament diferit. Se
confirmă astfel importanța transferului materialului forestier de reproducere în cadrul aceleiași
regiuni de proveniență.
▪ În aceleași condiții ecologice, clonele de stejar brumăriu de origini diferite (Caracal,
Fundulea și Brănești) au înflorit în perioade diferite, ceea ce confirmă din nou importanța utilizării
materialului forestier de reproducere doar în cadrul aceleiași regiuni de proveniență.

57
5.2 Contribuții originale
➢ Realizarea unei ample analize multivariate la nivelul descriptorilor morfologici foliari ai
stejarului pedunculat și stejarului brumăriu, cu evaluări intraspecifice și interspecifice.
➢ Identificarea pentru ambii taxoni, atât la nivel interspecific, cât și, în premieră națională și
internațională, la nivel intraspecific, de funcții discriminante ce se pot utiliza în vederea
determinării nu numai a speciei, ci și a varietăților. În cadrul funcției de discriminare a celor doi
taxoni a fost validat pentru prima dată aportul pe care îl are descriptorul CL (culoarea de pe dosul
frunzei).
➢ Realizarea în premieră a unei sinteze morfologice comparative, validată statistic, după
descriptorii frunzelor stejar pedunculat - stejar brumăriu.
➢ Recomandarea introducerii unor elemente noi de descriere morfologică a frunzelor
anumitor forme și varietăți.
➢ Realizarea de planșe herborizate cu tipurile de unități infraspecifice identificate la ambii
taxoni; cîte un dosar complet va fi depus atât la Herbarul Facultăţii de Silvicultură şi Exploatări
Forestiere Braşov – BVS, cât și la Herbarul Institutului Naţional de Cercetare Dezvoltare în
Silvicultură „Marin Drăcea” Bucureşti - BUCF.
➢ Fundamentarea unei metodologii de evaluare fenologică precisă, ușor de utilizat, care
poate fi adaptată și la alte specii. De altfel, în vederea diseminării rezultatelor tezei, această
metodologie a fost propusă ca protocol de lucru internațional în cadrul proiectului internațional
Trees4Future fiind adoptată deja de INRA (French National Institute for Agricultural Research).
➢ Evaluarea în premieră a fenologiei de primăvară, în special a înfloririi în plantaje cu
scopul identificării suprapunerii antezei clonelor. O suprapunere a înfloririi ar presupune o
hibridare de tip polycross care, asigură fenomenul de heterozis, iar o diferențiere dintre
perioadelel de înflorire ar presupune o consangvinizare.
➢ Determinarea corelațiilor multianuale intra- și interspecifice dintre fenofazele deschiderea
mugurilor foliari și fenofaza antezei care susțin utilizarea primei fenofaze pentru prognozarea
momentului înfloririi.
➢ Realizarea în premieră a analizelor xilologice la cvercinee în vederea coroborării acestora
cu fenologia, inclusiv cu senescența.
➢ Elaborarea de recomandări privind regulile de transfer și comercializare a materialului
forestier de reproducere fundamentate pe baza rezultatelor variabilității intra- și interspecifice ale
fitocenozelor studiate, precum și pe cea a rezultatelor analizelor fenologice.
➢ Recomandarea introducerii criterului fenologic la alegerea arborilor plus din arboretele
constituite ca materiale de bază pentru producerea materialelor forestiere de reproducere.

58
5.3 Diseminarea rezultatelor
A. Lucrări publicate în reviste ISI
Apostol (Chesnoiu), E.N., Curtu, A.L., Daia, L.M., Apostol, B., Dinu C.G., Șofletea N., 2017. Leaf
morphological variability and intraspecific taxonomic units for pedunculate oak and grayish oak
(genus Quercus L., series Pedunculatae Schwz.) in Southern Carpathian Region (Romania).
Science of the Total Environment, DOI:10.1016/j.scitotenv.2017.05.274. FI=4,900; SRI= 1,900.
B. Lucrări publicate în reviste indexate în baze de date internaționale (BDI)
1. Apostol - Chesnoiu, E.N., Curtu, A.L., Şofletea, N., 2015. Structura taxonomică
intraspecifică într-un complex de cvercinee din estul României, la contactul cu zona silvostepei
externe, Revista de Silvicultură și Cinegetică. 37, 47-51.
2. Enescu, M., Chesnoiu, E.N., Şofletea, N., Curtu, A. L, 2010. Leaf Morphology in Quercus
Robur L. Genetic Resources Across Romania. Bulletin of the Transilvania University of Braşov.
3(52), II, 47-54.
3. Chesnoiu, E. N., Șofletea, N., Curtu, A. L., Toader, A., Radu, R., Enescu, M., 2009. Bud
burst and flowering phenology in a mixed oak forest from Eastern Romania. Annals of Forest
Research. 52, 199-206.
4. Curtu, A.L., Şofletea, N., Toader, A.V., Enescu, M., Moldovan, C., Chesnoiu, E. N., 2009.
Stejarul brumăriu: specie sau unitate intraspecifică a stejarului pedunculat?. Revista Pădurilor, anul
124. 5, 24-30.
C. Lucrări prezentate la simpozioane și conferințe internaționale
1. Apostol E.N., Sidor C., 2017. Spring phenology in relation to global warming in Quercus
species. The 6th Scientific Conference of ICP Forests, 16-17 May, Bucharest, Romania.
2. Apostol (Chesnoiu) E.N., Semeniuc A., Dinu C.G., Curtu Al.L. Şofletea N., 2016. Leaves
and xylem phenology of oaks, a tool of evaluate forest ecosystems from Eastern Carpathians
tableland in the context of climate change. Forum Carpaticum 2016, Future of the Carpathians:
Smart, Sustainable, Inclusive, September 28 – 30, 2016, Bucharest, Romania.
3. Ducci F., Pâques L.E., Kowalczyk J.3, Proietti R.1, Rzońca M., Santi F.2, Monteverdi M.C.,
Chesnoiu E., 2015. Exploration of new techniques/tools for medium/high throughput phenotyping of
Phenology. Trees4Future Final Conference, Brussels, 4-6 Aprilie 2016, DOI: 10.13140/RG.2.1.1170.4725.
4. Chesnoiu (Apostol) E.N., Şofletea N., Dinu C.G., Curtu Al.L. 2014. Spring phenology
within two white oak seed orchards in Romania. The 13th international symposium / prospects for
the 3rd millenium agriculture, 25th-27th September 2014, University of Agricultural Sciences And
Veterinary Medicine Cluj-Napoca

59
5. Chesnoiu (Apostol) E.N., Şofletea N., Curtu Al.L. 2014. Quercus spring phenology within
a mixed oak forest from Eastern Romania. International Symposium „Forest and Sustainable
Development" Brașov, Romania 24-25 October.
6. Pârnuţă, Gh., Tudoroiu, M., Chesnoiu, E. N., Stuparu, El., Budeanu, M., Şofletea, N.,
Curtu, L. A., Daia, M., 2010. Conservation and sustainable management of forest genetic resources
in Romania-Adaptation of species in extreme site conditions. In TREEBREEDX Seminar on
Genetic Variability and Adaptive Potential of Temperate and Boreal Forest Trees Species,
Bucureşti , România, 23-25 February 2010
7. Chesnoiu, E.N., Șofletea, N., Curtu, L. A., 2010. Phenological evaluations within a mixed
oak forest from Eastern Romania. Padurea si dezvoltarea durabilă, Facultatea de Silvicultură și
Exploatări Forestiere Brașov, 15-16 Octombrie, Brașov, România
8. Chesnoiu, E.N., Şofletea, N., Curtu, L. A., Toader, A., Radu, R., Enescu M., 2008.
Bud burst and flowering phenology in a mixed oak forest from Eastern Romania. "Sustainable
Forestry in a Changing Environment" (ICAS – 75 years of activity)" ICAS, 23-25 Octombrie,
București, România.
5.4 Direcții viitoare de cercetare
Din punct de vedere al variabilității morfologice studiate, rezultatele obținute susțin statutul
taxonomic de specie al stejarului brumăriu, însă, dată fiind lipsa unei bariere prezigotice între cei
doi taxoni, care face posibilă hibridarea dintre aceștia, pe viitor trebuie continuate cercetările
privind hibridarea naturală interspecifică și evaluarea structurii genetice a acestora, în vederea
certificării diferențierii clare a lor, ca urmare a speciației ecologice. De asemenea, este imperios a se
extinde rețeaua fenologică în scopul identificării a cât mai multor arbori plus în populații adaptate
condițiilor climatice în schimbare și promovării celor mai valoroase populații, în vederea
constituirii de materiale de bază pentru lucrările de împădurire și de resurse genetice forestiere
pentru conservarea in situ.
Având în vedere că arborii tardivi au un potențial ridicat de producere de semințe cu o
periodicitate mai mică decât exemplarele tipice, datorită riscului mai redus de vătămare a florilor de
către înghețurile târzii, identificarea și selectarea acestora ca arbori plus, pentru utilizarea
materialului seminologic în practica silvică, trebuie să constituie o prioritate. De importanță
științifică și practică este identificarea și descrierea în viitor a formelor ”tardiflora” cu ajutorul
markerilor moleculari și utilizarea acestor metode avansate de certificare a provenienței materialului
forestier de reproducere valoros. Totodată, se impune continuarea cercetărilor cu privire la
mecanismul producerii activității cambiale, în corelație cu fenologia de primăvară, spre a oferi
informații utile pentru managementul durabil și eficient al ecosistemelor în care apar cei doi taxoni.

60
Bibliografie (selecție)
1. Antonova, G.F. and Stasova,V.V., 1997. Effects of environmental factors on wood formation in larch (Larix sibirica
Ldb.) stems. Trees. 11, 462 – 468.
2. Apostol - Chesnoiu, E.N., Curtu, A.L., Şofletea, N., 2015. Structura taxonomică intraspecifică într-un complex de
cvercinee din estul României, la contactul cu zona silvostepei externe, Revista de Silvicultură și Cinegetică. 37, 47-
51.
3. Apostol E.N., Curtu, A.L., Daia, L.M., Apostol, B., Dinu C.G., Șofletea N., 2017. Leaf morphological variability
and intraspecific taxonomic units for pedunculate oak and grayish oak (genus Quercus L., series Pedunculatae
Schwz.) in Southern Carpathian Region (Romania). Science of the Total Environment, acceptat la publicare.
4. Armăşescu S., Tabrea A., 1974. Cercetări asupra caracteristicilor dimensionale şi silvoproductive ale arboretelor de
stejar brumăriu şi stejar pufos. Analele I.C.A.S. 31(1), 207 – 220.
5. Athanasiadis, N., Tonkov, S., Atanassova, J. Bozilova, E., 2000. Palynological study of Holocene sediments from
Lake Doirani in northern Greece. Journal of Paleolimnology. 24(3), 331-342.
6. Bacilieri R., Labbe T., Kremer A., 1994. Intraspecific genetic structure in a mixed population of Quercus
petraea (Matt.) Liebl. and Q. robur L. Heredity 73 (2), 130-141.
7. Bacilieri, R., Ducousso, A., Kremer, A., 1995. Genetic, morphological, ecological and phenological differentiation
between Quercus petrea (Matt.) Liebl. and Quercus robur L. in a mixed stand of Nortwest of France. Silvae
Genetica. 44(1), 1-10.
8. Bacilieri, R., Ducousso, A., Kremer, A., 1996. Comparison of morphological characters and molecular markers for
the analysis of hybridization in sessile and pedunculate oak. Annals of Forest Science, nr.53, 79-91.
9. Badea, O., 1998. Fundamente dendrometrice şi auxologice pentru monitoringul forestier. Rezumatul Tezei de
doctorat, Suceava, 35 p.
10. Bašić N., Kapić J., Ballian D., 2007. Morfometrijska analiza varijabilinosti svojstava lista hrasta lužnaka (Querqus
robur L.) na području Sjeverne, Izvorni znanstevni članak, Sumar. inst. Jastrebar. 42(1), 5-18.
11. Bălănică T., Tomescu A., 1953. Dare de seamă asupra observaţiilor fenologice forestiere efectuate în anul 1949.
Analele I.C.A.S. 13(1), 71 – 80.
12. Beldeanu E., 2008. Specii de interes sanogen din fondul forestier. Editura Universităţii Transilvania Braşov.
13. Becker, M., Levy, G., 1990. Le point sur l'écologie comparée du chêne sessile et du chêne pédonculé. Revue
Forestière Française. 42, 48-54.
14. Beldie, Al., 1952. Genul Quercus. In: Nyarady, E. (ed.): Flora R.P.R. vol. I, Editura Academiei R.P.R., pp. 224 –
261.
15. Beldie, Al. şi Cretzoiu, P., 1941. Studiu sistematic al gorunului din România. Analele ICAS. 7 (1), 38-49.
16. Beldie A., 1977. Flora Ramâniei. Determinator ilustrat al plantelor vasculare, vol. I. Editura Academiei R.S.
România, pp. 81-85.
17. Bândiu, C-tin, Smejkal, G. M., Vişoiu-Smejkal, D., 1995. Pădurea seculară - Cercetări ecologice în Banat. Editura
Mirton, Timişoara, pp. 151-152.
18. Boratynski, A., Marcysiak, K., Lewandowska, A., Jasinsk, A., Iszkulo, G., Burczyk, J., 2008. Differences in leaf
morphology between Quercus petraea and Q. robur adult and young individuals. Silva Fennica. 42(1), 115–124.
19. Borazan A., Babac M., 2003. Morphometric leaf variation in oaks (Quercus) of Bolu, Turkey. Ann. Bot. Fennici.
40, 233 – 242.
20. Bordács, S., Popescu, F., Slade, D.,Csaikl, U. M., Lesur, I., Borovics, A., Kézdy, P., König, A. O., Gömöry, D.,
Brewer, S., Burg, K., Prtit, R. B., 2002. Chloroplast DNA variation of white oaks in northern Balkans and in the

61
Carpathian Basin. Forest Ecology and Management 156, 197-209.
21. Borlea, G. F., 1998. Forest reserves and their research in Romania. In Proceedings of the COST Action E. pp. 67-86.
22. Borza, Al., 1937. Quercus pedunculiflora C. Koch, un stejar nou pentru România. Bul. Grăd. Bot. Muz. Bot., Univ.
Cluj, XVI. 1-4, 55-62.
23. Broshtilov, K., 2006. Quercus robur L. leaf variability in Bulgaria, Plant Genetic Resources Newsletter. 147, 64-71.
24. Bruschi, P., Vendramin, G.G., Bussotti, F., Grossoni, P., 2000. Morphological and molecular differentiation
between Quercus petraea (Matt.) Liebl. and Quercus pubescens Willd. (Fagaceae) in Northern and Central Italy.
Annals of Botany. 85, 325-333.
25. Bruschi, P., Vendramin, G.G., Bussotti, F., Grossoni, P., 2003a. Morphological and molecular diversity among
Italian populations of Quercus petraea (Fagaceae), Annals of Botany, Vol. 91, pp. 707-716.
26. Bruschi, P., Grossoni, P., Bussotti, F., 2003b. Within- and amomg-tree variation in leaf morphology of Quercus
petraea (Matt.) Liebl. natural populations. Trees. 7, 164-172.
27. Carella, R. 2013. First observation of Quercus pedunculiflora K. Koch in the Italian Peninsula-short
communication. Journal of Forest Science. 59 (3), 130-135.
28. Chesnoiu, E. N., Șofletea, N., Curtu, A. L., Toader, A., Radu, R., Enescu, M., 2009. Bud burst and flowering
phenology in a mixed oak forest from Eastern Romania. Annals of Forest Research. 52, 199-206.
29. Ciocârlan, V., 2000. Flora ilustrată a României. Editura Ceres, Bucureşti.
30. Crabbe R., Dash J., Rodriguez-Galiano V., Janous D., Pavelka, M., Mareka M., 2016. Extreme warm temperatures
alter forest phenology and productivity in Europe. Science of The Total Environment, 563–564, pp: 486–495,
http://dx.doi.org/10.1016/j.scitotenv.2016.04.124
31. Crăciunesc, I., Vornam, B., Leinemann, L., Finkeldey, R., Șofletea, N., Curtu, A.L., 2015. High Genetic
Differentiation among European White Oak Species (Quercus spp.) at a Dehydrin Gene. Notulae Botanicae Horti
Agrobotanici Cluj-Napoca. 43(2), 582-588.
32. Crăciunesc, I., 2013. Evaluarea nivelului de hibridare naturală la specii autohtone de cvercinee: analiză de caz în
rezervația naturală Bejan-Deva. Teză de doctorat.
33. Curtu A.L., 2006. Patterns of genetic variation and hybridiyation in a mixed oak (Quercus spp.) forest. Cuvillier
Verlag Gottingen, 159 p.
34. Curtu, A.L., Gailing, O., Finkeldey, R., 2007. Evidence for hybridization and introgression within a species-rich oak
(Quercus spp.) community. BMC Evolutionary Biology. 7, 218,
35. Curtu, A.L., Şofletea, N., Toader, A.V., Enescu, M., Moldovan, C., Chesnoiu, E. N., 2009. Stejarul brumăriu:
specie sau unitate intraspecifică a stejarului pedunculat? Revista Pădurilor, anul 124. 5, 24-30.
36. Curtu, A.L., Craciunesc, I., Enescu C.M., Vidalis, A., Sofletea, N., 2015. Fine-scale spatial genetic structure in a
multi-oak-species (Quercus spp.) forest. iForest – Biogeoscience and Forestry. 8, 324-332.
37. Davis P. H. 1965–1988. Flora of Turkey and the East Aegean Islands, Vol: 1–10, Edinburgh University Press,
Edinburgh.
38. Doniţă, N., Popescu, A., Paucă-Comănescu, M., Mihăilescu, S., Biriş, I. A., 2005. Habitatele din România. Editura
Tehnică Silvică, Bucureşti.
39. Dubrovský M., Trnka M., Holman I.P., Svobodová E., Harrison P.A., 2015. Developing a reduced-form ensemble of
climate change scenarios for Europe and its application to selected impact indicators. Climatic Change 128: 169–
186. http://dx.doi.org/10.1007/s10584-014-1297-7.
40. Dumitriu - Tătăranu, I., 1954. Asupra prezenţei şi răspândirii stejarului pufos şi a celui brumăriu, precum şi a
câtorva specii lemnoase rare în pădurile Ocolului Silvic Huşi. Revista Pădurilor. 11, 534-539.
41. Dumitriu - Tătăranu, I., 1960. Arbori şi arbuşti forestieri şi ornamentali cultivaţi în R.P.R.. Editura Agro-Silvică, pp.

62
574-600.
42. Dupouey, J.-L., Fougère V., Kremer, A. 1990. Variabilité génétique des Chênes sessile et pédonculé estimée à l`aide
de marqueurs morphologiques et moléculaires”. Rev. For. Fr. XLII-2, pp. 198-204.
43. Dupouey, J.L., Badeau, V.,1993. Morphological variability of oaks (Quercus robur L, Quercus petraea (Matt)
Liebl, Quercus pubescens Willd) in northeastern France: preliminary results, Annals of Forest Science. 50(1), 35s-
40s.
44. Enescu, V., 1993. A test of half-sib progenies of greyish oak, Quercus pedunculiflora K Koch, Annals of Forest
Science, Vol 50(1), 439s-443s.
45. Enescu, M., Chesnoiu, E.N., Şofletea, N., Curtu, A. L, 2010. Leaf Morphology in Quercus Robur L. Genetic
Resources Across Romania. Bulletin of the Transilvania University of Braşov. 3(52), II 47-54.
46. Enescu, C. M., Curtu, A. L., Șofletea N., 2013. Is Quercus virgiliana a distinct morphological and genetic entity
among European white oaks? Turkish Journal of Agriculture and Forestry 37,632-641.
47. Fortini, P., Antonecchia, G., Di Marzio, P., Maiuro, L., Viscosi, V, 2015. Role of micromorphological leaf traits and
molecular data in taxonomy of three sympatric white oak species and their hybrids (Quercus L.). Plant Biosystems.
149(3), 546-558.
48. Franjić, J., Liber, Z., Škvorc, Ž., Idžojtić, M., Šoštarić, R., Stančić, Z., 2006. Morhological and molecular
differentiation of the Croatian populations of Quercus pubescens Willd. (Fagaceae). Acta Societatis Botanicorum
Poloniae. 75(2), 123-130.
49. Fu, Y.H., Campioli, M., Deckmyn, G., Janssens, I.A., 2012. The Impact of Winter and Spring Temperatures on
Temperate Tree Budburst Dates: Results from an Experimental Climate Manipulation. PLoS ONE 7(10): e47324.
doi:10.1371/journal.pone.0047324.
50. Gailing, O., 2008. QTL analysis of Leaf Morhological Characters in a Quercus robur full-sib family (Q. robur x Q.
robur ssp. slavonica). Plant Biology. 10, 624-634.
51. Gancz, V., Pătrăşcoiu, N., 2000. Cartografierea ecosistemelor forestiere din România prin mijloace GIS şi de
teledetecţie. Revista Pădurilor. 2, 35–40.
52. Georgescu C. C., Cretzoiu, P., 1941. Consideraţiuni sistematice asupra speciei Quercus pedunculiflora K. Koch în
România. Analele ICEF. I (VII), 3-37 + 3 planşe + 25 fig. + 4 tab.
53. Georgescu, C.C., Lupe, L., Cretzoiu, P., 1942. Răspândirea stejarului brumăriu (Quercus pedunculiflora K. Koch).
[Natural distribution of Grayish oak (Quercus pedunculiflora C. Koch).] Analele Icas. 8, 165–172.
54. Georgescu C. C., colab., 1943. Noi contribuţii la sistematica speciei Quercus pedunculiflora în România. Revista
Pădurilor. 7(8), 293-300.
55. Georgescu, C.C., Morariu, I., 1948. Monografia stejarilor din România. Universul, Bucureşti, 26p + 11 planşe.
56. Georgescu C.C., Petrescu M., Purcelean St., 1954. Studii asupra transpiraţiei câtorva specii de şleau. Analele
I.C.A.S. 15(1), 447-457.
57. Gömöry, D., Yakovlev, I., Zhelevs, P., Jedinakova, J., Paule, L., 2001. Genetic differentiation of oak populations
within the Quercus robur/Quercus petraea complex in Central and Eastern Europe. Heredity. 86, 557-563.
58. Govaerts, R., Frodin, D. G. 1998. World checklist and bibliography of Fagales, Royal Botanical Garden Kew.
59. Gričar J., 2013. Influence of Temperature on Cambial Activity and Cell Differentiation in
Quercus Sessiliflora and Acer Pseudoplatanus of Different Ages, Drvna Industrija 64 (2) 95-105.
60. Grossoni, P., Bettini, D., Bruschi, P., Bussotti, F., Lumini, M.B., 1998. Studio macro e micromorfologico di
Quercus petraea (Matt.) Liebl. e di Quercus pubescens Willd. della macchia di Berignone-Tatti (Volterra, Italia
Centrale) [Toscana]. Monti e Bosch 49( 2), 25-30.
61. Gülz, P. G., & Müller, E., 1992. Seasonal Variation In The Composition of Epicuticular Waxes of Quercus Robur

63
Leaves. Zeitschrift für Naturforschung C,.47(11-12), 800-806.
62. IPCC, 2013. Climate change 2007: the physical science basis. Technical report. In: Solomon S., Qin D., Manning
M., Chen Z., Marquis M., Averyt K.B., Tignor M., Mille H.L. (eds). Contribution of working group I to the fourth
assessment report of the intergovernmental panel on climate change. Ambridge University Press, Cambridge, United
Kingdom and New York, NY, USA. 996 p.
63. Kelleher, C.T., Kelly, D.L., Hodkinson, T.R., 2004. Species status, hybridisation and geographic distribution of Irish
populations of Quercus petraea (Matt.) Liebl. and Q. robur L.. Watsonia 25, 83-97.
64. Kissling, P., 1997. Les poils des quatre espèces de chênes du Jura (Quercus pubescens. Q. petraea. Q. robur et Q.
cerris). Bericht der Schweizerischen Botanischen Gesellschaft. 87, 1-18.
65. Kleinschmit, J., 1993. Intraspecific variation of growth and adaptive traits in European oak species, Annals of Forest
Science. 50, 166-185.
66. Kremer, A., Dupouey, J.L., Deans, J.D., Cottrel, J., Csaikl, U.M., Finkeldey, R., Espinel, S., Jensen, J.S.,
Kleinschmit, J., Van Dam, B., Ducousso, A., Forrest, I., De Herdia, U.L., Lowe Andew, J., Tutkova, M., Minro,
R.C., Badeau, V., 2002. Leaf morphological differentiation between Quercus robur and Quercus petraea is stable
across western European mixed stand. Annals of Forest Science. 59, 777-787.
67. Kubitzki, K., 1993. Fagaceae In: Kubitzki, K., Rohwer, J. G., Bittrich, V., (Eds.), The families and genera of
vascular plants. Spinger, Berlin, pp. 301-309.
68. Kucharczyk, H., Kucharczyk, M., Stanisławek, K., Fedor, P., 2012. Application of PCA in Taxonomy Research –
Thrips (Insecta, Thysanoptera) as a Model Group, Principal Component Analysis - Multidisciplinary Applications,
Dr. Parinya Sanguansat (Ed.), ISBN: 978-953-51-0129-1, InTech, Available from:
http://www.intechopen.com/books/principal-component-analysis-multidisciplinary-applications/application-ofpca-
in-taxonomy-research-thrips-insecta-thysanoptera-as-a-model-group
69. Lechowicz, M.J., 1995. Seasonality of flowering and fruiting in temperature forest trees. Canadian J. Bot. 73 (2),
175-182.
70. Ledig, F.T., Wilson, R.W., Duffield, J.W., Maxwell, G., 1969. A discriminant analysis of introgression between
Quercus prinus L. and Quercus alba L, Bulletin of the Torrez Botanical Club, Vol. 96, No. 2, pp. 156-163.
71. Ledig, F.T., 1993. Genetics of oaks species and the spectre of global climate change, Annals of Forest Science, Vol
50, Suppl 1, pp. 456s-460s.
72. Lefort, F., Lally, M., Thompson, D., Douglas, G.C., 1998. Morphological traits, microsatellite fingerprinting and
genetic relatedness of elite oaks (Q. robur L.) at Tullynally, Ireland, Silvae Genetica 47( 5-6), 257-262.
73. Marciszewska K. Tulik M., 2013. Hydraulic Efficiency and Safety of Xylem Sap Flow in Relation to Water Stress
in Woody Plants, Intech, http://dx.doi.org/10.5772/56656.
74. Martiník, A., Dobrovolny, L., & Palátová, E., 2014. Tree growing space and acorn production of Quercus robur.
Dendrobiology, 71.
75. Masato, I. 2009. Variation in leaf morphology of Quercus crispula and Quercus denata assemblages among contact
zones: a method for detection of probable hybridization, ournal of forest Research, 14, 240-244.
76. Masarovičová, E., 1991. Leaf Shape, Stomata Density and Photosynthetic Rate of the Common Oak. Biologia
Plantarum. 33(6), 495-500.
77. Mehrnia, M., Nejadsattari T., Assadi, M., Mehregan I. 2013. Taxonomic study of the genus Quercus L. Sect.
Quercus in the Zagros forests of Iran. J. Bot. 19 (1), 62-74.
78. Menzel, A., Sparks, T. H., Estrella, N., Koch, E., Aasa, A., Ahas, R. and Chmielewski, F. M. 2006. European
phenological response to climate change matches the warming pattern. Global change biology, 12(10), 1969-1976.
79. Negulescu, E., Săvulescu, AL., 1957. Dendrologie. Editura Agro-Silvică de Stat Bucureşti, 457p.

64
80. Negulescu, E.G., Stănescu, V., 1964. Dendrologia, cultura şi protecţia pădurilor. Editura Didactică şi Pedagogică,
Bucureşti, 500 p.
81. Norby R.J., Hartz - Rubin J.S., Verbrugge M.J., 2003. Phenological responses in maple to experimental
atmospheric warming and CO2 enrichment. Global Change Biology 9, 1792–1801.
82. Ozturk, M., Ozcelik, H., Sakcali, S., Guvensen, A., 2004. Land degradation problems in Euphrates Basin, Turkey.
EnviroNews, Newsletter of ISEB India. 10(3), 7-9.
83. Pârnuţă, Gh., Stuparu, El., Budeanu, M., Scărlătescu,V.,Marica, Fl. M., Lalu, I., Filat, M., Tudoroiu, M., Lorenț, A.,
Şofletea, N., Curtu, A-L., Daia, M.,Nica, M. S., Teodosiu, M., Chesnoiu, E. N., Pârnuţă, P., Mirancea, I., Marcu,
C., 2011. “Catalogul naţional al resurselor genetice forestiere”, Editura Silvică, 526 p.
84. Pârnuţă, Gh., Budeanu, M., Stuparu, El., Scărlătescu,V., Chesnoiu, E. N., Tudoroiu, M., Filat, M., Nica, M. S.,
Teodosiu, M., Lorenț, A., Daia, M., Dinu, G.C, 2012. Catalogul Naţional al Materialelor de Bază pentru producerea
Materialelor Forestiere de Reproducere. Editura Silvică, 334 p.
85. Petit R.J., Bodenes C., Ducousso A., Roussel G., Kremer, A., 2004. Hybridization as a mechanism of invasion in
oaks. New Phytologist 161 (1), 151-164.
86. Ponton, S., Dupouey, J-L., Dreyer, E., 2004. Leaf morphology as species indicator in seedlings of Quercus robur L.
and Q. petraea (Matt.) Liebl: modulation by irradiance and growth flush. Annals of Forest Science. 61, 73-80.
87. Popa, I., Semeniuc, A., 2009. Posibilităţi de evaluare a fazelor de formare a inelului anual prin tehnici de xilologie,
Revista pădurilor. 4, 13-16.
88. R Core Team 2016. R: A language and environment for statistical computing. R Foundation for Statistical
Computing. Vienna. Austria. URL https://www.R-project.org/.
89. Rădulescu A.V., 1938. Observațiuni fenologice la speciile lemnoase din București, fãcute în sezonul de vegetație
1937. Analele ICAS 4 (1), 231-247.
90. Rossi, S., Deslauriers, A., Anfodillo, T., 2006. Assessment of cambial activity and xylogenesis by microsampling
tree species: and example at the alpine timberline. IAWA Journal 27, 383-394.
91. Sanda, V., Ollerer, K., Burescu, P., 2008. Fitocenozele din România - Sintaxonomie, Structură şi Evoluţie. Editura
Ars Docendi, Universitatea din Bucureşti, 576 p.
92. Sârbu, I., Ștefan, N., Oprea, A.,. 2013. Plante vasculare din România. Editura victor B. Victor, București, 1320p.
93. Schwarz, O., 1964. Quercus L. In: Tutin, T.G., Heywood,V.H., Burges, N.A., Valentine, D.H., Walters, S.M. and
Webb, D.A. (Eds.) Flora Europaea, vol. 1: Lycopodiaceae to Platanaceae. Cambridge University Press, Cambridge,
pp. 61-64.
94. Stănescu, V., Şofletea, N., Popescu, O., 1997. Flora forestieră lemnoasă a României, Editura Ceres, Bucureşti, pp.
174-213.
95. Steinhoff, S.S., 1997. Results of Quercus hybridization workfrom 1989 to 1996 at Escherode (Quercus petraea
(Matt.)Liebl. and Quercus robur L.). In: Steiner, K. C. (ed.) Diversity and Adaptation in Oak Species. The
Pennsylvania State University, Pennsylvania, pp. 156±164.
96. Simota C., Dumitru S., Vizitiu O., Coteţ V., Ignat P., Mateescu E., Alexandru D.,
Cofas E., 2014. Ghid de bune practici agricole pentru atenuarea efectului schimbărilor
climatice asupra agriculturii. ICPA, ANM, USAMV Bucureşti. Document disponibil la:
www.icpa.ro/documente/ADER%20511_ghid.pdf
97. Șofletea, N., Curtu, L., 2001. Dendrologie. Editura Pentru Viață, vol II, 300p.
98. Şofletea, N., 2005. Genetică şi ameliorarea arborilor. Editura “Pentru Viaţă”, Braşov, 455 p.
99. Şofletea, N., Curtu, L., 2007. Dendrologie. Editura Universităţii Transilvania, Braşov, 418 p.
100. Șofletea, N., Enescu, C.M., Curtu, A.L., 2011. Small-scale morphological descriptors analysis in four romanian

65
oak stands reported to Series lanuginosae simk. Bulletin of the Transilvania University of Braşov Series II:
Forestry • Wood Industry • Agricultural Food Engineering. 4 (53) 2, 77-84.
101. Teodosiu, M., 2004. Studiu privind starea principalilor parametri climatici în cuprinsul rețelei de supraveghere
intensivă a ecosistemelor forestiere (nivel II). Referat ştiinţific parţial ICAS, 40 p.
102. Toader, A., Şofletea, N., Curtu, Al. L., 2009. Variaţia genetică izoenzimatică a stejarului pedunculat (Quercus
robur L.) şi stejarului brumăriu (Quercus pedunculiflora K. Koch.) din România, Proceedings of the Biennial
International Symposium Forest and Sustainable Development, Editura Universităţii Transilvania din Braşov, pp.
1-8.
103. Tomescu, A., 1964. Fazele periodice de vegetație la speciile forestiere în anul 1961, în R. P . R.. Analele I.C.A.S.
24(1), 29 -50.
104. Tomescu A., Florescu I.I., Mihalache A., Ștrimbei M., Avramescu C., 1967. Cercetãri fenologice la
principalele specii forestiere autohtone din RSR - Sinteza pentru perioada 1956-1965. Centrul de
documentare tehnică pentru economia forestieră, București, 221 p.
105. Tomescu, A., 2000. Evaluation of Holocene pollen records from the Romanian Plain. Review of Palaeobotany and
Palynology. 109(3), 219-233.
106. TREES4FUTURE - Designing Trees for the Future (coordonat de L.E.Pâques (INRA, Orléans, FR), 2016.
Alternative methodologies for phenology assessment in forest trees, http://www.trees4future.eu/uploads/t4f-
training-
activities/Annex%20I%201%20Alternative%20methodologies%20for%20phenology%20assessment%20in%20for
est%20trees.pdf, pp. 4-6.
107. Viscosi, V., Fortini, P., Slice, D.E., Loy, A., Blasi, C., 2009a. Geometric morphometric analyses of leaf variation
in four oak species of the subgenus Quercus (Fagaceae). Plant Biosystems - An International Journal Dealing with
all Aspects of Plant Biology: Official Journal of the Societa Botanica Italiana, pp. 143-575.
108. Viscosi, V., Lepais, O., Gerber, S., Fortini, P., 2009b. Leaf morphological analyses in four European oak species
(Quercus) and their hybrids: A comparison of traditional and geometric morphometric methods. Plant
Biosystems. 143(3), 564-574.
109. Vuckovic, B., 1984. A new habitat of Quercus pedunculiflora C. Koch in Serbia (Yugoslavia).] Zbornik radova
Instituta za sumarstvo i drvnu industriju. 22–23, 111–113.
110. Wigston, D.L., 1975. The distribution of Quercus robur L, Q. petraea (Matt) Liebl and their hybrids in south-
western England. The assessment of the taxonomic status of populations from leaf characters. Watsonia. 10,345-
369.
111. Wright, K., 2015. Corrgram: Plot a Correlogram. R package version 1.8. https://CRAN.R-
project.org/package=corrgram
112. ***2012. Forest National Inventory of Romania.
113. *** Herbarul Facultăţii de Silvicultură şi Exploatări Forestiere Braşov – BVS.
114. *** Herbarul Institutului Naţional de Cercetare-Dezvoltare în Silvicultură “Marin Drăcea” Bucureşti - BUCF.
115. *** IBMSPSS software.
116. *** 2008. STATISTICA 8, StatSoft Inc., Tulsa, OK, USA.
117. ***2009. UK Governments indicators of climate change (http://www.forestry.gov.uk/fr; Department for
Environment, Food and Rural Affairs.
118. *** 2007. WinFOLIA, Regent Instruments INC.

66
SCURT REZUMAT
SHORT SUMMARY
Scopul studiului a constat în identificarea și validarea unor caracteristici (elemente)
discriminatorii relevante între Q. robur și Q. pedunculiflora.
În cadrul tezei de doctorat s-au realizat evaluări fenotipice la nivelul descriptorilor
morfologici foliari ai populaţiilor autohtone de stejar pedunculat şi stejar brumăriu (atât la nivel
interspecific, cât și, în premieră, la nivelul unităților infraspecifice), precum și analize fenologice și
xilologice. Cu ajutorul analizelor fenotipice s-au putut valida o serie de caracteristici morfologice ce
contribuie semnificativ la diferențierea celor doi taxoni analizați, cea mai mare diferențiere între cei
doi taxoni se constată la nivelul descriptorilor morfologici foliari, precum pubescența (PU) și
culoarea de pe dosul frunzelor (CL).
Analiza fenologiei a scos în evidență lipsa unei bariere prezigotice care să împiedice
hibridarea celor doi taxoni analizați, așa încât există alte mecanisme care contribuie la menținerea
identității morfologice a celor doi taxoni. De asemenea, din punct de vedere al analizelor xilologice,
nu există diferențe în ceea ce provește perioadele formării inelului anual la stejarul pedunculat și
stejarul brumăriu.
The goal of the present study was to identify and validate relevant discriminatory traits
(elements) between Q. robur and Q. pedunculiflora.
In the thesis, the phenotypic evaluations were made at the foliar morphological descriptors
of the natural population of pedunculate oak and grayish oak (both interspecifically and, for the first
time, at the level of infraspecific units) as well as phenological and xylological analyzes. With the
help of phenotypic analyzes, a number of morphological characteristics were validated, which
significantly contribute to the differentiation of the two taxa analyzed, the greatest difference
between the two taxa is found at the level of leaves morphological descriptors, such as pubescence
(PU) and the color on the back of the leaf (CL).
The analysis of phenology has highlighted the lack of a prezygotic barrier that prevents
hybridization of the two taxa analyzed, so there are other mechanisms that help maintain the
morphological identity of the two taxa. Also, from the point of view of the xylological analyzes,
there are no differences in the formation of the annual ring between the pedunculate oak and the
grayish oak.

67
CURRICULUM VITAE
Date personale
Nume: CHESNOIU (căs. APOSTOL)
Prenume: Ecaterina Nicoleta
Adresa: str. Galata nr. 55, loc. Voluntari, jud. Ilfov
Data și locul nașterii: 23.02.1982, Drăgășani, jud. Vâlcea
Cetățenie: română
Starea civilă: căsătorită
Telefon: 0721818827
E-mail: [email protected]
Experiență profesională:
2007 - prezent: Institutul Naţional de Cercetare - Dezvoltare în Silvicultură „Marin Drăcea“ (fost
ICAS), cercetător științific în cadrul Colectivului de Genetică Forestieră și Ameliorarea Arborilor
Educație:
2010 – 2012: Universitatea de Arhitectură și Urbanism „Ion Mincu” din București, Master în Peisaj
și Teritoriu
2008 – 2010: Universitatea „Gh. Asachi” din Iași, Master în Managementul Administrării
Afacerilor (MBA)
2001 – 2006: Universitatea „Transilvania” din Brașov, Facultatea de Silvicultură și Exploatări
Forestiere
Publicații:
A. Lucrări publicate în reviste indexate în baze de date internaționale (ISI & BDI): 8 (4 ca prim
autor)
B. Lucrări prezentate la simpozioane și conferințe internaționale: 10
C. Cărți publicate: 2

68
CURRICULUM VITAE
Personal data
Surname: CHESNOIU (marr. APSOTOL)
First name: Ecaterina Nicoleta
Address: 55 Galata Street, Voluntari City, Ilfov County
Date an place of birth: 23.02.1982, Drăgășani, Vâlcea County
Nationality: romanian
Marital status: married
Phone: 0721818827
E-mail: [email protected]
Profesional experience:
2007 - present: Naţional Institute for Research and Development in „Marin Drăcea“ (former ICAS),
scientific researcher within Genetics and Tree Breeding Department
Education:
2010 – 2012: University of Architecture and Urbanism “Ion Mincu” Bucharest, Master degree in
Landscape and Territory
2008 – 2010: University “Gh. Asachi” of Iași, Master degree in Management Business
Administratiom (MBA)
2001 – 2006: University “Transilvania” of Brașov, Faculty of Silviculture and Forest Engineering
Publications:
A. Research articles published in journals indexed in international data bases (ISI & BDI): 8 (4 as
first author)
B. Papers presented at national and international conferences and simposya: 10
C. Books published: 2