Ă a activitĂŢii cardiace aspecte teoretice - umfcv.ro nervoasa a aparatului cv_26 oct... ·...
TRANSCRIPT

Lucrări practice Fiziologia aparatului cardio-vascular
1
REGLAREA NERVOASĂ A ACTIVITĂŢII CARDIACE
ASPECTE TEORETICE Principala funcţie a pompei cardiace este de a propulsa un volum de sânge în mica şi marea circulaţie. Acest volum de sânge se numeşte volum sistolic sau volum bătaie şi trebuie să aibă o valoare suficient de mare pentru a asigura nevoile de oxigen şi substanţe nutritive ale organismului. Volumul expulzat într-un minut se numeste debit cardiac şi păstrarea lui la valori care să satisfacă cerinţele metabolice ale organismului se face prin intervenţia unor mecanisme complexe de reglare şi control. Cum volumul de sânge pompat de inimă (debitul cardiac) depinde de frecvenţa bătăilor inimii şi de volumul expulzat cu fiecare bătaie, adaptarea inimii se realizează prin controlul activităţii de pacemaker şi a celei contractile, adică a activităţii electice şi mecanice a inimii. (Milnor W., 1989). Mecanismele care intervin în controlul activităţii inimii sunt clasificate în intrinseci (care ţin de inimă) şi extrinseci. • Mecanismele intrinseci de reglare a cordului
a. mecanismul heterometric – Frank-Starling de adaptare la presarcină crescută, în care modificările debitului cardiac sunt determinate de alungirea fibrelor cardiace. Acest mecanism nu asigură componenta presională, fiind un mecanism fiziopatologic de adaptare la suprasolicitări;
b. mecanismul homeometric de adaptare la postsarcină, la aceeaşi lungime iniţială a fibrelor. Reglarea se realizează prin mecanisme de modificare a contractilităţii miocardului şi prin modificarea frecvenţei cardiace.
Mecanismele dependente de inimă (intrinseci) intervin în puţine situaţii fiziologice, în fapt, controlul activităţii cardiace realizându-se pe cale nervoasă şi umorală. • Mecanismele extrinseci de reglare cardiacă sunt: − mecanisme nervoase care au la bază arcuri reflexe şi eferenţe vegetative
simpatice şi parasimpatice; − mecanisme umorale.
Mecanismele nervoase sunt mecanisme reflexe şi presupun parcurgerea unui arc anatomic reflex care cuprinde:
1. zonele receptoare (reflexogene) la nivelul cărora, prin intermediul baro şi chemoreceptorilor, se percepe modificarea care declanşează mecanismul;
2. căile nervoase aferente, care transportă stimulii la centrii de control din SNC; 3. centrii cardiovasculari care emit comanda în funcţie de nevoile organismului; 4. căi eferente amielinice care au pe traseu un ganglion, unde se face sinapsa
dintre neuronul pre şi postganglionar; 5. efectorul, care poate fi: − celula pacemaker, în cazul în care este necesară modificarea de frecvenţă; − fibra contractilă pentru adaptarea forţei de contracţie; − fibra musculară netedă din peretele vaselor pentru adaptarea tonusului
vascular (vasoconstricţie sau vasodilataţie).

Fiziologia aparatului cardio-vascular Lucrări practice
2
1. Receptorii cardiovasculari Receptorii implicaţi în reglarea activităţii inimii şi vaselor sunt prezenţi în întregul sistem cardiovascular, dar cu precădere la nivelul unor zone reflexogene “strategice” (sinocarotidiană, cardio-aortică, a venelor mari, etc.). Ei au rol de traductori ai modificărilor presionale – baroreceptori şi/sau ai modificărilor compoziţiei biochimice a sângelui – chemoreceptori. Baroreceptorii care participă la reglarea cardio-vasculară sunt situaţi în: − artere - baroreceptori arteriali care se găsesc în crosa aortică, la emergenţa
arterelor subclaviculare şi în sinusul carotidian, deasupra bifurcaţiei carotidei primitive, pe carotida internă. Sunt stimulaţi de distensia pereţilor arteriali, produsă cel mai frecvent de creşterea presiunii arteriale. Baroreceptorii sinocarotidieni şi aortici sunt stimulaţi de modificările presiunii arteriale sistemice. Creşterea presiunii arteriale sistemice (sau compresiunea carotidelor deasupra bifurcaţiei) măreşte frecvenţa impulsurilor până la un anumit nivel, proporţional cu creşterea presiunii. Aceste impulsuri determină un reflex depresor care are ca finalitate: scăderea debitului cardiac (prin scăderea frecvenţei şi a contractilităţii cardiace) şi scăderea rezistenţei periferice vasculare. Scăderea presiunii arteriale (sau compresia pe carotida comună) stimulează baroreceptorii sinocarotidieni şi aortici, determinând un efect opus, caracterizat prin tahicardie, vasoconstricţie, creşterea presiunii şi a debitului cardiac - reflex presor.
− atrii - baroreceptorii atriali. Sunt situaţi subendocardic, endocardul atrial fiind zona cardiacă cu cea mai bogată inervaţie şi sunt de tip A (descarcă mai ales în sistola atrială) şi de tip B (descarcă tardiv în cursul diastolei atriale, sincron cu perioada de umplere maximă cu sânge a atriilor). Frecvenţa de descărcare a receptorilor atriali depinde direct proporţional de presiunea venoasă. Stimularea receptorilor din atriul drept (AD) produce tahicardie prin reflexul Bainbridge realizat pe căi aferente vagale şi căi eferente simpatice, care influenţează frecvenţa de descărcare a nodulului sinoatrial. Reflexul Bainbridge are rolul de a preveni acumularea sângelui în vene, atrii şi în circulaţia pulmonară. Distensia atrului stâng (AS) produce un răspuns asemănător, având aceeaşi explicaţie.
− peretele ventriculilor - baroreceptorii ventriculari sunt reprezentaţi de mecanoreceptori, stimulaţi de distensia ventriculară, mai puţin numeroşi decât cei atriali. Ei produc efecte depresoare, dar numai în cazul destinderilor foarte mari ale ventriculului stâng.
− circulaţia pulmonară - baroreceptorii din circulaţia pulmonară sunt localizaţi în adventicea trunchiului arterei pulmonare şi a ramurilor dreaptă şi stângă ale acesteia. Sunt stimulaţi de distensia patului vascular pulmonar, determinând vasodilataţie cu hipotensiune arterială şi bradicardie, adică reflexe depresoare. În condiţii experimentale aceşti receptori pot fi stimulaţi de injectarea în circulaţia pulmonară a veratridinei, fenilbiguanidei sau a serotoninei.
− zone din afara aparatului cardiovascular: receptori alveolari, receptori pentru durere, proprioreceptori, receptori mezenterici. o

Lucrări practice Fiziologia aparatului cardio-vascular
3
o Baroreceptorii mezenterici sunt mecanoreceptori cu structură
asemănătoare corpusculilor Pacini, cu rol posibil în reglarea circulaţiei viscerale şi care, prin stimulare, iniţiază reflexe cardiovasculare de tip depresor. Existenţa lor este dovedită experimental prin experienţa lui Goltz, când stimularea unei anse intestinale determină reflex oprirea cordului.
Chemoreceptorii. Sunt prezenţi în regiuni strategice ale sistemului arterial (chemoreceptorii periferici) sau la diferite niveluri ale nevraxului (chemoreceptorii centrali). Ei sunt stimulaţi de modificările presiunilor parţiale ale gazelor şi pH-ului sanguin, nefiind exclusă şi influenţa componentelor biochimice plasmatice. − Chemoreceptorii periferici: sinocarotidieni (aflaţi la bifurcaţia carotidei
primitive, în corpusculul sau glomusul carotidian) au cea mai bogată irigaţie pe unitatea de greutate. Chemoreceptorii aortici sunt situaţi în apropierea crosei aortice. Stimulul principal pentru activarea chemoreceptorilor este scăderea pO2, dar stimularea lor este determinată şi de creşterea pCO2. Chemoreceptorii aortici, spre deosebire de cei sinocarotidieni, nu sunt activaţi de modificarea pH-ului, fiind chiar deprimaţi. Creşterea temperaturii are efect stimulant asupra chemoreceptorilor, iar scăderea temperaturii efect inhibitor. Stimularea chemoreceptorilor determină creşterea ventilaţiei şi secundar, creşterea frecvenţei cardiace şi a presiunii sanguine (efect presor).
− Chemoreceptorii centrali – cei mai cunoscuţi sunt cei bulbari, situaţi pe faţa ventrală, în apropierea rădăcinilor nervilor cranieni IX, X şi XI. Sunt sensibili la modificarea pH-ului extracelular şi a lichidului cefalorahidian (LCR).
2. Căile aferente ale receptorilor cardiovasculari Căile de legătură ale receptorilor cardiovasculari cu centrii de reglare cardiovasculară sunt realizate prin ramuri ale nervului vag sau ale altor nervi parasimpatici. − baroreceptorii din crosa aortică sunt legaţi prin fibre ale nervului vag (nervul
Cyon-Ludwig) de centrul cardioinhibitor şi vasodepresor. − baroreceptorii din sinusul carotidian sunt legaţi prin fibre ale glosofaringianului
(nervul Hering) de centrul cardioinhibitor şi vasodepresor din bulb. Nervul vag şi glosofaringian se mai numesc şi nervi tampon. Stimularea lor electrică produce bradicardie, vasodilataţie şi hipotensiune arterială, iar secţionarea lor produce creşterea tensiunii arteriale până la valori de 300/200 mmHg (“hipertensiune neurogenă sau de defrenare”).
− chemoreceptorii ventriculari sensibili la anoxie şi acidoză au aferenţe aparţinând sistemului simpatic, care ajung la centrii medulari vasoconstrictori sau cardioacceleratori.
3. Centrii de reglare cardiovasculară Centrii nervoşi implicaţi în reglarea activităţii cardiace sunt situaţi la diferite nivele ale sistemului nervos central. Cei mai importanţi centri se găsesc în trunchiul cerebral şi în măduva toraco-lombară. − centrul cardioinhibitor este localizat în bulb, mai ales în nucleul ambiguu şi
mai puţin în nucleul dorsal al vagului şi în nucleul tractului solitar. Centrul cardioinhibitor este conectat cu baroreceptorii din sinusul carotidian şi crosa aortică, iar stimularea sa electrică produce bradicardie şi scăderea debitului cardiac prin eferenţele vagale.

Fiziologia aparatului cardio-vascular Lucrări practice
4
− centrul vasomotor este localizat în formaţiunea reticulată, într-o zonă difuză, situată între obex şi nucleii vestibulari şi de la tavanul ventriculului IV până la piramidele bulbare. Stimularea electrică a regiunilor rostrale şi laterale ale centrului vasomotor (zona presoare) produce vasoconstricţie, tahicardie şi hipertensiune arterială, pe când stimularea unor zone mai restrânse situate în jurul obexului (zona depresoare) produce bradicardie, vasodilataţie şi hipotensiune arterială. Centrul vasomotor, prin cele două zone ale sale (presorie şi depresorie), modulează activitatea cordului şi vaselor, acţionând numai asupra centrilor vasoconstrictori medulari, cărora le stimulează sau le inhibă activitatea.
− centrii medulari se găsesc în coarnele intermedio-laterale ale măduvei toracolombare (T1-L2). Centrul cardioaccelerator este localizat în principal în segmentele T1 şi T2, dar la inervaţia inimii mai participă şi fibre simpatice cu originea în segmentele T3-T6. Secţionarea măduvii spinării în regiunea cervicală produce o scădere rapidă a presiunii arteriale, vasodilataţie periferică, fenomen numit “şocul spinal”. Dacă se păstrează nervul frenic şi eferenţa simpatică preganglionară, situaţia revine la normal în câteva zile.
4. Eferenţele centrilor de reglare cardiovasculară − eferenţa parasimpatică. Fibrele vagale eferente fac sinapsă în celulele
postganglionare localizate la nivelul inimii, în vecinătatea nodulului sinoatrial (NSA) şi a celui atrioventricular (NAV). Inervaţia vagală a musculaturii ventriculare şi a fasciculului His este foarte slab reprezentată. Vagul drept inervează mai ales zona NSA, stimularea sa producând un reflex vagal evidenţiat prin bradicardie sau chiar oprirea inimii pentru câteva secunde în diastolă. Vagul stâng influenţează mai ales conducerea atrioventriculară, deoarece se distribuie la nodulul atrioventricular. Stimularea vagală, în afară de bradicardie şi întârzierea conducerii atrioventriculare mai produce o uşoară scădere a forţei de contracţie prin efecte inotrop negative (scăderea presiunii maxime ventriculare şi scăderea presiunii în diastolă). Nervul vag induce creşterea rezervelor de glicogen şi fosfaţi macroergici la nivelul miocardului, realizând un efect protector asupra inimii. o Eferenţa parasimpatică are 2 neuroni: protoneuronul localizat în centrul
cardioinhibitor şi deutoneuronul localizat în musculatura atrială. Există o permanentă descărcare a potenţialelor de acţiune prin nervii vagi care deprimă activitatea cordului. De aceea, denervarea parasimpatică a cordului, realizată prin metode chirurgicale sau farmacologice (cu atropină) produce tahicardie (frecvenţa cardiacă creşte de la 70 la 150/min.).
o Efectele parasimpaticului se realizează prin eliberarea mediatorului numit acetilcolină la nivelul terminaţiunilor nervoase. Acetilcolina acţionează asupra receptorilor muscarinici (M2) membranari cu eliberare de GMPc, urmată de scăderea activităţii adenilatciclazei şi a AMPc. Activitatea inhibitorie la nivelul miocardului se datorează hiperpolarizării indusă de creşterea permeabilităţii pentru K+ a membranei celulare.
o Efectul deprimant al vagului şi acetilcolinei poate fi contracarat prin blocarea receptorilor M cu atropină.

Lucrări practice Fiziologia aparatului cardio-vascular
5
− eferenţa simpatică. Căile simpatice îşi au originea în cordoanele intermediolaterale ale primelor 5-6 segmente toracice medulare. Sinapsa cu neuronii postganglionari se face la nivelul ganglionilor cervico-dorsali, de unde pleacă fibrele postganglioanare sub forma nervilor cardiaci (superior, mijlociu şi inferior), care inervează sistemul excitoconductor şi fibrele contractile. Fibrele simpatice drepte se repartizează mai ales în ţesutul nodal, afectând în special frecvenţa, în timp ce, cele stângi se distribuie cu precădere în micardul contractil, amplificându-i activitatea Ca şi în cazul parasimpaticului, există o descărcare permanentă de impulsuri prin nervii simpatici cardiaci. Denervarea simpatică, realizată pe un cord în prealabil denervat vagal, determină scăderea frecvenţei cardiace (la om de la 150 la 100/min.). Efectele simpaticului pe cord se datorează eliberării mediatorului numit noradrenaliă (epinefrină) care acţionează pe receptorii beta1. Noradrenalina stimulează toate proprietăţile miocardului şi mobilizează rezervele de glicogen şi fosfaţi macroergici, acţiunea sa fiind de mai lungă durată decât a acetilcolinei. Efectele sale se manifestă prin creşterea permeabilităţii membranei celulare pentru Na+ şi Ca2+, cu depolarizarea acesteia. o
Reţineţi • SNV parasimpatic via nervul vag şi mediatorul său acetilcolina
au efecte deprimante asupra proprietăţilor miocardului. În condiţii de stimulare vagală normală, acest lucru este benefic, având efecte protectoare asupra inimii. În caz de stimulare vagală puternică, apare bradicardia severă sau chiar oprirea inimii.
• SNV simpatic prin mediatorii săi, adrenalina si noradrenalina are efecte stimulatoare asupra proprietăţilor miocardului. Aceste efecte sunt benefice pentru adaptarea organismului în condiţii de efort, emoţii, stare de “alertă”, când cordul trebuie să pompeze volume mai mari de sânge. Dacă simpaticul domină în mod cronic activitatea inimii, atunci se instalează tahicardia, tulburările de excitabilitate şi hipertensiunea arterială. În aceste situaţii trebuie administrate beta blocante.
TIPURI DE REFLEXE CARDIOVASCULARE – reflexe depresoare şi presoare
• Reflexe cu originea în baroreceptorii arteriali − creşterea presiunii arteriale medii sau celei diferenţiale produce distensia
pereţilor arterelor mari şi determină stimularea baroreceptorilor din sinusul carotidian şi din crosa aortică. Răspunsul la această stimulare este de tip depresor, iar reflexul declanşat se numeşte un reflex depresor. Impulsurile generate de baroreceptori sunt conduse prin nervii tampon până la centrul cardioinhibitor şi până la zona depresorie a centrului vasomotor, cărora le stimulează activitatea. Stimularea centrului cardioinhibitor produce prin intermediul nervilor vagi, bradicardie şi scăderea forţei de contracţie a miocardului, ceea ce duce la scăderea debitului cardiac. Stimularea zonei depresorii a centrului vasomotor produce deprimarea centrilor vasoconstrictor şi cardioaccelerator medulari, determinând vasodilataţie şi bradicardie.

Fiziologia aparatului cardio-vascular Lucrări practice
6
Scăderea debitului cardiac şi vasodilataţia determină scăderea presiunii arteriale, ceea ce face ca baroreceptorii să-şi scadă frecvenţa de descărcare.
− scăderea presiunii arteriale produce scăderea, până la dispariţie a descărcărilor baroreceptorilor, ceea ce blochează centrul cardioinhibitor şi zona depresorie a centrului vasomotor şi stimulează astfel, indirect, centrii cardioaccelerator şi vasoconstrictor medulari, determinând tahicardie, vasoconstricţie şi creşterea presiunii arteriale. Reflexul este de tip presor
• Reflexe cu originea în baroreceptorii atriali – stimularea baroreceptorilor atriali prin creşterea presiunii venoase produce tahicardie şi vasodilataţie, având ca rezultat scăderea presiunii venoase. În plus, stimularea baroreceptorilor atriali mai determină inhibarea secreţiei de vasopresină din nucleii supraoptic şi paraventricular hipotalamici, ceea ce duce la creşterea diurezei şi la scăderea presiunii venoase. Baroreceptorii atriali par a fi punctul de plecare al reflexului Bainbridge, care constă în apariţia tahicardiei la injectarea rapidă intravenoasă de soluţii saline sau de sânge la animale anesteziate.
• Reflexul Bezold-Jarisch – injectarea de veratridină sau de de nicotină în ramurile arterelor coronare care irigă ventriculul stâng, produce bradicardie, vasodilataţie şi hipotensiune arterială, prin stimularea chemoreceptorilor ventriculari. Declanşarea acestui reflex în infarctul de miocard are rol protector asupra inimii lezate, şi o menajează de o activitate prea intensă. Uneori poate fi prea puternic şi atunci, reflex, poate determina prăbuşirea tensiunii arteriale şi chiar să oprească inima. Reflexul dispare dacă se secţionează nervul vag. Un reflex depresor asemănător cu reflexul Bezold-Jarisch, din punct de vedere al efectelor cardiovasculare şi al mecanismului de producere, apare la distensia mecanică a ventriculului stâng.
• Chemoreflexul pulmonar – injectarea în circulaţia pulmonară a veratridinei, fenilbiguanidei sau serotoninei, ori distensia mecanică a patului vascular pulmonar, stimulează chemo, respectiv mecanoreceptorii din circulaţia pulmonară care, la rândul lor stimulează, prin intermediul unor aferenţe vagale, centrii cardioinhibitor şi vasodepresor, determinând bradicardie, vasodilataţie şi hipotensiune arterială.
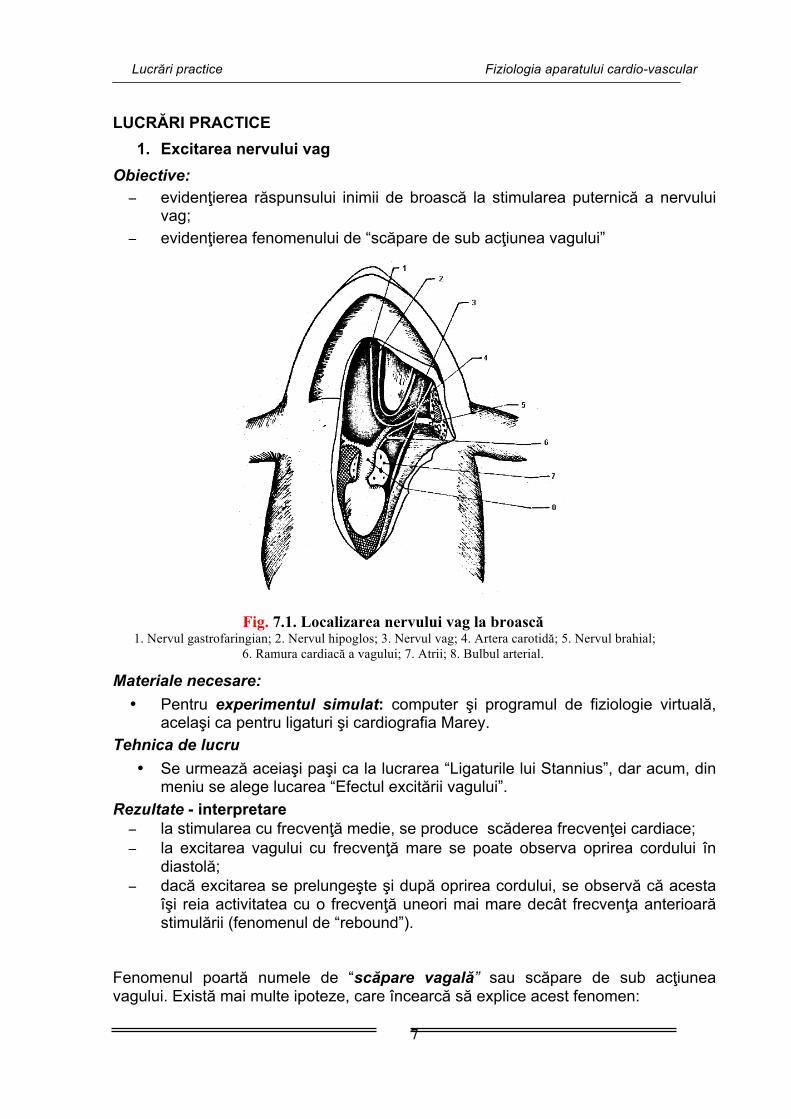
Lucrări practice Fiziologia aparatului cardio-vascular
7
LUCRĂRI PRACTICE
1. Excitarea nervului vag Obiective: − evidenţierea răspunsului inimii de broască la stimularea puternică a nervului
vag; − evidenţierea fenomenului de “scăpare de sub acţiunea vagului”
Materiale necesare: • Pentru experimentul simulat: computer şi programul de fiziologie virtuală,
acelaşi ca pentru ligaturi şi cardiografia Marey. Tehnica de lucru
• Se urmează aceiaşi paşi ca la lucrarea “Ligaturile lui Stannius”, dar acum, din meniu se alege lucarea “Efectul excitării vagului”.
Rezultate - interpretare − la stimularea cu frecvenţă medie, se produce scăderea frecvenţei cardiace; − la excitarea vagului cu frecvenţă mare se poate observa oprirea cordului în
diastolă; − dacă excitarea se prelungeşte şi după oprirea cordului, se observă că acesta
îşi reia activitatea cu o frecvenţă uneori mai mare decât frecvenţa anterioară stimulării (fenomenul de “rebound”).
Fenomenul poartă numele de “scăpare vagală” sau scăpare de sub acţiunea vagului. Există mai multe ipoteze, care încearcă să explice acest fenomen:
Fig. 7.1. Localizarea nervului vag la broască 1. Nervul gastrofaringian; 2. Nervul hipoglos; 3. Nervul vag; 4. Artera carotidă; 5. Nervul brahial;
6. Ramura cardiacă a vagului; 7. Atrii; 8. Bulbul arterial.

Fiziologia aparatului cardio-vascular Lucrări practice
8
• intrarea în activitate a centrilor inferiori de automatism ; • stimularea vagală repetată, prelungită duce la eliberarea acetilcolinei
depozitată în veziculele de la fibrele postganglionare şi epuizarea acesteia. Absenţa mediatorului va duce la dispariţia efectului inhibitor şi inima îşi reia activitatea;
• umplerea ventriculară crescută, care se produce în timpul opriri cordului determină alungirea fibrelor miocardice, cu deschiderea unor canale ionice favorizând apariţia potenţialelor de acţiune;
2. Reflexul Goltz La nivelul mezenterului există mecanoreceptori care pot fi stimulaţi prin tracţiune, lovire sau distensie. Excitaţia este transmisă prin măduvă (căile medulare ascendente) la nucleul dorsal al vagului, de unde, pe căi eferente vagale ajunge la cord, determinând efecte deprimante, care pot ajunge până la oprirea cordului. Existenţa unor astfel de reflexe depresoare poate fi pusă în evidenţă cu ajutorul experienţei descrise de Goltz. Mod de lucru – pentru experiment pe broască
• Se foloseşte o broască nespinalizată, decapitată, dar cu bulbul intact, care se fixează pe planşeta de disecţie în decubit dorsal.
• Se execută o breşă în musculatura abdominală şi se expune o ansă intestinală împreună cu mezenterul. Se mai face o breşă în peretele toracic, pentru evidenţierea cordului.
Rezultate • După 10-15 minute de expunere la aer a ansei intestinale, interval necesar
pentru uscarea parţială şi sensibilizarea acesteia la stimuli mecanici, se loveşte ansa intestinală cu bagheta de sticlă sau se tracţionează mezenterul.
• Se observă, după un timp de latenţă scurt (1-2 secunde), o scădere a frecvenţei contracţiilor cardiace sau chiar oprirea cordului în diastolă. Reflexul dispare după atropinizare. o
Reflexele vagale viscero-vegetative (în acest caz cu punct de plecare digestiv) urmate de bradicardie severă sau stop cardiac se pot produce la om: − în cursul intervenţiilor chirurgicale pe abdomen, dacă se execută
manipulări brutale ale viscerelor abdominale; − dacă anestezia nu este suficient de profundă; − dacă în preanestezie nu s-a administrat atropină.
În cazul declanşării unui astfel de reflex trebuie administrată atropină.
3. Reflexul Dagnini-Aschner sau oculo-cardiac • Compresiunea globilor oculari produce excitarea terminaţiilor senzitive ale
trigemenului. Excitaţia ajunge la nucleul trigemenului şi de aici, prin formaţiunea reticulată, la centrul cardioinhibitor. Acesta determină creşterea frecvenţei de descărcare a impulsurilor prin nervii vagi şi consecutiv bradicardie şi scăderea vitezei de conducere a impulsurilor prin nodulul atrioventricular.
• Reflexul se manifestă numai în situaţia în care frecvenţa cardiacă este crescută. Din acest motiv, pentru a demonstra practic, în laborator, acest reflex se vor efectua 20-30 de genuflexiuni, notând frecvenţa cardiacă după

Lucrări practice Fiziologia aparatului cardio-vascular
9
acest efort fizic (trebuie să fie mai mare de 120/1minut, dar să nu depăşească 170/1 minut).
• Atenţie! Se recomandă efectuarea practică a reflexului de către doi examinatori: un examinator comprimă globii ocolari şi al doilea examinator supraveghează frecvenţa cardiacă.
o Se execută o compresiune moderată, prin intermediul pleopelor, asupra globilor oculari, timp de 15-20 secunde, ceea ce determină o scădere a frecvenţei cardiace faţă de valorile anterioare.
o Scăderea frecvenţei cardiace după compresiunea globilor oculari depinde de reactivitatea vegetativă a subiectului. La subiecţii cu tonus vagal accentuat, în urma efectuării acestei manevre, se poate ajunge până la sincopă vagală, pe când la subiecţii cu tonus simpatic crescut se observă numai o scădere minoră a frecvenţei cardiace.
În tahicardiile paroxistice atriale, compresiunea globilor oculari, produce scăderea frecvenţei cardiace sau chiar revenirea la ritmul sinusal. Acest efect se datorează deprimării reflexe a focarului ectopic atrial şi scăderii vitezei de conducere prin nodulul atrioventricular.
În tahicardiile paroxistice ventriculare, compresiunea globilor oculari este ineficientă, pentru că inervaţia vagală a ventriculilor este foarte slab reprezentată anatomic şi funcţional. Prin urmare, manevra de compresiune a globilor oculari reprezintă o metodă de diagnostic diferenţial între tahicardiile paroxistice atriale şi cele ventriculare şi o metodă de tratament în tahicardiile atriale.
FIZIOLOGIE INTEGRATIVĂ Manevrele vagale. Manevrele vagale sunt probe clinice prin care sunt stimulate zone reflexogene implicate în reflexe care au drept cale eferentă nervul vag. Acest nerv se distribuie viscerelor din torace şi abdomen (inimă, plămân, tub digestiv, sistem excretor, etc.), deci stimularea vagală va produce efecte vagotonice şi la aceste organe. În practica medicală manevrele vagale sunt efectuate pentru influenţarea proprietăţilor miocardului, în special pentru modificarea frecvenţei cardiace. Nervul vag are origine în bulb. Primul neuron se găseşte în nucleul dorsal al vagului şi în nucleul ambiguu. Fibrele preganglionare lungi coboară în mediastin şi fac sinapsă cu al doilea neuron, situat în ganglionii parasimpatici localizaţi în peretele inimii, în aproprierea nodulului sinoatrial (NSA) şi a nodulului atrioventricular (NAV). Fibrele postganglionare sunt scurte şi se distribuie musculaturii atriale şi structurilor supraventriculare ale sistemului excito-conductor: NSA şi 1/3 superioară a joncţiunii atrioventriculare. Există fibre vagale şi în musculatura ventriculilor, mai ales în ventriculul stâng, dar stimularea vagală nu are efecte clinice la acest nivel. Nervul vag prezintă fenomenul de teritorializare: nervul vag drept se distribuie predominant la nivelul NSA, iar nervul vag stâng se distribuie predominant la nivelul NAV. Mediatorul parasimpatic la nivelul cordului este acetilcolina, iar receptorii colinergici sunt de tip muscarinic, M2 şi M3.

Fiziologia aparatului cardio-vascular Lucrări practice
10
Efectele stimulării vagale sunt: • Scăderea automatismului NSA exprimată prin scăderea frecvenţei cardiace; • Scăderea conducerii potenţialelor de acţiune de la celulele pacemaker din
NSA la miocardul din jurul acestuia; • Creşte perioada refractară a ţesutului miocardic din jurul NSA; • Creşte durata conducerii potenţialelor de acţiune în NAV; • Creşte perioada refractară la nivelul celulelor pacemaker din NAV; • Încetineşte conducerea prin musculatura atrială.
Proprietăţile miocardului sunt influenţate prin stimulare vagală astfel: • Efect cronotrop negativ: scade frecvenţa cardiacă, posibil până la oprirea
cordului la indivizi hiperactivi sau cu leziuni miocardice severe. Este posibilă manifestarea fenomenului de “scăpare” de sub influenţa vagului, dar există şi situaţii în care apare stop cardiac neresuscitabil.
• Efect inotrop negativ - scade forţa de contracţie a miocardului urmată de scăderea debitului cardiac şi a tensiunii arteriale;
• Efect dromotrop negativ - scade viteza de conducere în miocard; • Efect batmotrop negativ - scade excitabilitatea miocardului; • Efect tonotrop negativ - scade tonusul muscular; • Scade consumul de oxigen al miocardului.
Concluzii. • Sistemul nervos parasimpatic scade frecvenţa cardiacă, scade viteza de
conducere prin NAV şi scade forţa de contracţie a miocardului cu 20-30%. • Sistemul nervos parasimpatic protejează miocardul ventricular în tulburările de
ritm supraventriculare, cu frecvenţă crescută, prin reducerea numărului potenţialelor de acţiune transmise de la atrii la ventriculi.
• Sistemul nervos parasimpatic se comportă ca un mecanism de protecţie a cordului faţă de suprasolicitări.
În repaus, la nivelul cordului predomină efectele sistemului nervos parasimpatic faţă de cel simpatic.
Exemple de manevre vagale
• Masajul sinusului carotidian. • Compresiunea globilor oculari (reflexul Dagnini-Aschner). • Manevra Valsalva: inspir forţat urmat de expir forţat cu glota închisă, timp de
15-20 secunde. • Aplicarea la nivelul feţei al unui prosop cu gheaţă sau apă rece. • Aplicarea pe o suprafaţă mare a toracelui a unui prosop umed rece. • Imersia corpului în apă rece. • Stimularea esofagului prin ingestie de apă rece sau bol alimentar dur. • Inspir profund lent. • Tuse provocată, strănut, efort de defecaţie. • Stimularea faringelui pe peretele posterior până la provocarea vărsăturii. • Compresia abdominală cu coapsele flectate pe abdomen sau cu palmele
cadrului medical.

Lucrări practice Fiziologia aparatului cardio-vascular
11
Există o semnificativă variabilitate interindividuală a răspunsului reflex în cazul stimulării unor zone reflexogene. Reflexul vagal cu stop cardiac şi sincopă, uneori neresuscitabil, poate să apară la unii indivizi prin: compresie puternică a sinusului carotidian, prin lovituri de tip karate, prin imersia bruscă a corpului în apă rece, lovirea testiculelor, pensarea colului uterin, puncţie venoasă, puncţie pleurală, chirurgia abdomenului, lovituri la “plex”, şocuri emoţionale puternice, zgomote foarte puternice, etc.
Indicaţii ale manevrelor vagale. Manevrele vagale sunt indicate în scop: Terapeutic:
• pentru reducerea frecvenţei cardiace în tahicardia parosistică supraventriculară (TPSV) sau convertirea frecvenţei cardiace la ritm sinusal. Manevrele sunt eficiente în tahicardia cu reintrare nodală şi în tahicardia asociată cu sindrom de preexcitaţie.
• pentru creşterea blocajului potenţialelor de acţiune la NAV şi reducerea gradului de transmitere la ventriculi în fibrilaţia atrială şi în flutter-ul atrial.
Diagnostic: • Evidenţierea sindromului de preexcitaţie pe ECG. • Evidenţierea flutter-ului atrial mascat de TPSV. • Tahicardiile ventriculare nu răspund la manevrele vagale. • Manevrele vagale nu sunt eficiente în TPSV în cazul unor leziuni severe
instalate (ischemie, neuropatie vegetativă, etc.). Contraindicaţii ale manevrelor vagale. Manevrele vagale sunt contraindicate la:
• Boli coronariene, infarct miocardic acut, hipotensiunea arterială, hipovolemie. • Subiecţi cunoscuţi cu hipervagotomie. • Sportivi de performanţă care, prin antrenamente, au dezvoltat reactivitate
vagală crescută şi prin aceasta manifestă un bloc atrio-ventricular funcţional. • Pacienţi cu sindrom de hipersensibilitate sinocarotidiană. • Pacienţi cu tratament digitalic, posibil supradozaj digitalic.
LECTURĂ SUPLIMENTARĂ - MEDIATORII CHIMICI AI SISTEMULUI NERVOS VEGETATIV Acetilcolina – mediatorul sistemului parasimpatic, produce efecte deprimante asupra tuturor proprietăţilor cordului (efecte cronotrop, dromotrop, inotrop, batmotrop şi tonotrop negative), acţionând asupra receptorilor muscarinici.
• Efectele cronotrop negative ale acetilcolinei se explică prin creşterea permeabilităţii membranei celulelor “P” (pace-maker) pentru K+, ceea ce determină hiperpolarizare prin ieşirea K+ din celulă şi apariţia unui eflux de K+ în cursul depolarizării lente diastolice. Hiperpolarizarea determină depărtarea potenţialului diastolic maximal de nivelul critic de declanşare şi scăderea consecutivă a frecvenţei de descărcare a impulsurilor. Efluxul de K+ în timpul depolarizării lente diastolice determină contrabalansarea influxului lent, depolarizant de Na+ şi Ca2+, care tinde să aducă potenţialul de membrană la nivelul critic de declanşare, producând astfel scăderea vitezei de depolarizare lentă diastolică şi deci a frecvenţei de descărcare a

Fiziologia aparatului cardio-vascular Lucrări practice
12
impulsurilor (Sabău M., 1999). • Efectele inotrop negative ale acetilcolinei se explică prin scurtarea duratei
potenţialului de acţiune al fibrelor miocardice, realizată pe seama platoului, ceea ce reduce influxul de Ca2+ ce precede sistola. S-a determinat prin studii efectuate cu microelectrozi pe un singur canal de Ca2+ (tehnica de patch-clamp), că acetilcolina blochează canalele de Ca2+, cu atât mai puternic cu cât acestea erau mai intens activate de stimularea adrenergică.
Catecolaminele – adrenalina şi noradrenalina produse de medulosuprarenală, ajung în torentul circulator şi au efecte asemănătoare celor eliberate la nivelul terminaţiunilor nervoase simpatice.
• Ele stimulează proprietăţile fundamentale ale miocardului, acţionând asupra receptorilor miocardici. Noradrenalina influenţează atât receptorii α, cât şi β. Stimularea α-receptorilor se asociază cu producerea de IP3 (inozitol trifosfat) şi DAG (diacil glicerol). IP3 eliberează Ca2+ din reticulul sarcoplasmic şi induce un efect inotrop pozitiv. DAG activează proteinkinaza C ce menţine contracţia fibrei netede în platou (Rosen M., et al., 1991).
• Stimularea β-receptorilor activează adenilatciclaza şi producerea de AMPc. Activarea proteinkinazei A dependentă de AMPc se manifestă prin efecte metabolice ce duc la creşterea lipolizei şi a glicogenolizei. Stimularea transportului Ca2+ la nivelul sarcolemei şi a reticulului sarcoplasmic se manifestă prin efecte inotrop pozitive (Sabău M., 1999).
• Efectele catecolaminelor asupra fenomenelor electrice şi mecanice ale inimii sunt mediate în special prin receptorii β1 localizaţi la nivel sarcolemal. De asemenea, efectele cronotrope pot fi mediate prin β1-receptorii de la nivelul NSA.
• Efectele cronotrop pozitive se explică prin creşterea permeabilităţii membranei celulelor pace-maker pentru Na+ şi Ca 2+, ceea ce accelerează panta depolarizării lente diastolice.
• Efectele dromotrop pozive se produc prin creşterea vitezei de conducere prin NAV şi fasciculul His, datorită creşterii permeabilităţii celulelor acestor structuri pentru Na+ şi Ca2+ .
• Efectele inotrop pozitive se realizează prin stimularea receptorilor β1 miocardici, ceea ce produce activarea în interiorul acestor celule a adenilatciclazei, proces care necesită energie furnizată de GPT. Adenilatciclaza determină transformarea ATP în AMP ciclic, care acţionează asupra proteinkinazei ciclic AMP-dependente, disociind-o în 2 subunităţi catalitice şi 2 subunităţi regulatoare. Una din subunităţile catalitice, proteinkinaza A, fosforilează proteinele canalului de Ca2+, determinând creşterea influxului de Ca2+ în sistolă de 3-4 ori, stimulând astfel contractilitatea.
• Proteinkinaza A mai determină fosforilarea unei proteine, fosfolambanul, care activează pompa de Ca2+ din reticulul sarcoplasmic, determinând creşterea vitezei de recaptare a Ca2+ în reticul, în perioada care precede diastola ventriculară. Rezultă scurtarea fazei de relaxare, dar şi creşterea cantităţii de Ca2+ din reticul disponibilă pentru următoarele sistole, ceea ce favorizează stimularea inotropismului.
• Proteinkinaza c-AMP dependentă fosforilează şi fracţiunea I a troponinei, reducând afinitatea troponinei C pentru Ca2+, ceea ce scurtează perioada de relaxare a miocardului, fenomen esenţial pentru prevenirea tetanizării cordului la frecvenţe crescute de contracţie. AMP ciclic împreună cu complexul Ca2+-modulină activează fosforilkinaza, care produce degradarea glicogenului, furnizând astfel energia necesară pentru viitoarele cicluri de contracţie-relaxare. AMP ciclic este degradat în celulă sub acţiunea fosfodiesterazei, enzimă care poate fi blocată de substanţe din clasa metilxantinelor (cafeină, teofilină). Astfel, metilxantinele determină, ca şi stimularea β-adrenergică, creşterea concentraţiei AMPc intracelular.
• Stimularea receptorilor β1 determină şi vasodilataţie coronariană, creând condiţii bune de de lucru ale miocardului.

Lucrări practice Fiziologia aparatului cardio-vascular
13
FIŞĂ DE LUCRU ÎN LABORATOR 1. Stimulaţi moderat vagul şi explicaţi ce se întâmplă. ........................................................................................................................................ 2. Stimulaţi în continuare vagul cu stimuli puternici şi explicaţi ce se întâmplă. …………………………………………………………………………………...... 3. Continuaţi stimularea vagală şi după încetarea activităţii inimii şi explicaţi ce se întâmplă. ........................................................................................................................................ 4. Desenaţi o cardiogramă în stare de repaus. Marcaţi cele trei momente de la întrebările de mai sus şi evidenţiaţi modificarile frecvenţei şi amplitudinii contracţiilor. ………………………………………………………………………………………………...... 5. Numiţi tipul de reflex pe care l-aţi declanşat şi descrieţi arcul reflex. ........................................................................................................................................ 6. Enumeraţi câteva situaţii în care poate apărea reflexul vagal cu stop cardiac şi sincopă. ………………………………………………………………………………………………...... 7. Descrieţi reflexul Goltz la broască şi precizaţi arcul reflex. ………………………………………………………………………………………………...... 8. Enumeraţi situaţiile în care un reflex depresor de tip Goltz se poate declanşa la om şi indicaţi modalităţile de prevenire ........................................................................................................................................ 9. Descrieţi reflexul Dagnini-Aschner şi precizaţi arcul reflex ………………………………………………………………………………………………....... 10. Explicaţi ce este o manevră vagală şi enumeraţi câteva

Fiziologia aparatului cardio-vascular Lucrări practice
14
………………………………………………………………………………………………...... 11. Enumeraţi câteva indicaţii ale manevrelor vagale şi discutaţi despre efectele lor ………………………………………………………………………………………………...... 12. Explicaţi ce efecte are stimularea vagului asupra proprietăţilor miocardului ………………………………………………………………………………………………...... 13. Enumeraţi receptorii implicaţi în reglarea activităţii cordului şi vaselor şi precizaţi tipul de reflex declanşat prin stimularea fiecăruia. ………………………………………………………………………………………………...... 14. indicaţi centrii nervoşi implicaţi în reglarea activităţii cordului şi vaselor şi precizaţi tipul de reflex declanşat prin stimularea fiecăruia. ………………………………………………………………………………………………...... 15. Descrieţi sumar inervaţia parasimpatică a inimii. ………………………………………………………………………………………………...... 16. Descrieţi sumar inervaţia simpatică a inimii.

Lucrări practice Fiziologia aparatului cardio-vascular
15
………………………………………………………………………………………………...... 17. Numiţi mediatorii chimici ai SNV parasimpatic şi simpatic, receptorii asupra cărora acţionează şi efectele obţinute. ………………………………………………………………………………………………...... 18. Numiţi 3 substanţe care pot bloca, prin inhibiţie competitivă efectele mediatorilor SNV simpatic şi parasimpatic. ………………………………………………………………………………………………...... 19. Numiţi efectorii prin care se realizează reglarea activităţii cordului şi vaselor. ………………………………………………………………………………………………...... 20. Precizaţi care sunt parametrii cardio-vasculari care trebuie adaptaţi la diferite variaţii ale mediului intern sau extern. ………………………………………………………………………………………………...... 21. Prezentaţi sistematizat mecanismele de acţiune pentru SNV simpatic şi parasimpatic. ………………………………………………………………………………………………...... 22. Descieţi şi exemplificaţi un reflex depresor. ………………………………………………………………………………………………...... 23. Descieţi şi exemplificaţi un reflex presor. ………………………………………………………………………………………………......